
Abstract
The proteolytic activity of gelatinase A, a member of the matrix metalloproteinase (MMP) family, is considered to be a critical factor in tumor cell penetration of the extracellular matrix. To express catalytic activity, however, gelatinase A requires activation by another MMP, membrane type 1-matrix metalloproteinase (MT1-MMP). The head and neck squamous cell carcinoma cell line, UM-SCC-1, forms a quiescent monolayer atop collagen unless stimulated with epidermal growth factor (EGF; 3.5 nmol/L), which induces single cell invasion within 48 hours. To determine the role of the MT1-MMP/gelatinase A protease system in an in vitro stromal invasion model, expression vectors for MT1-MMP and gelatinase A were transfected into UM-SCC-1 (SCC-1/MT and SCC-1/gelA, respectively). SCC-1/MT tumor cells were found to invade in the absence of growth factor stimulation. Additionally, these cells displayed shorter onset to invasion and penetrated deeper into the collagen gel with EGF stimulation than did control vector transfectants. SCC-1/gelA cells similarly demonstrated invasion in the absence of EGF and a heightened invasive potential under EGF-stimulated conditions. These results suggest that the MT1-MMP/gelatinase A protease system participates in squamous cell carcinoma invasion of collagenous matrices. (Otolaryngol Head Neck Surg 1999;121:337-43.)
The extracellular matrix (ECM) is a mesh of proteinaceous and carbohydrate macromolecules that must be cleaved by tumor cells before the cells can invade surrounding tissues and form metastatic colonies in distant organs and lymph nodes. 1 Identification of the enzyme or enzymes responsible for ECM degradation and cancer cell invasion may lead to new therapeutic modalities directed at limiting tumor invasion at local, regional, and distant sites. 2 Although unlikely to be curative, such therapy may prolong life in patients found to be unresectable or may serve as an adjunct to conventional palliative therapy.
Previous studies have demonstrated that a family of Zn++-dependent endopeptidases, the matrix metalloproteinases (MMPs), are highly expressed in most cancers. The most convincing evidence for the role of MMPs in invasion comes from studies using inhibitors. A family of naturally occurring inhibitors, the tissue inhibitors of metalloproteinases (TIMPs), specifically TIMP-1, TIMP-2, and TIMP-3, block the proteolytic activity of all known MMPs. Both in vitro and in vivo data suggest that tissue invasion by tumor cells is reduced in the presence of TIMP-1 and TIMP-2. 3,4 Expression vectors for TIMP-1 in a human gastric cell line and TIMP-2 in transformed rat embryo cells inhibited metastasis in laboratory animals. 5 Work from our laboratory has implicated MMPs, but not the plasminogen-plasminogen activator system in head and neck squamous cell carcinoma (HNSCC) invasion of dermal stroma. 6
Although several MMPs have been implicated in the invasive program, gelatinase A is considered to be a critical enzyme in tumor cell-mediated ECM dissolution. 1 Like most MMPs, gelatinase A requires proteolytic cleavage of its amino terminal end to display catalytic activity. Recent studies have identified a new class of membrane associated metalloproteinases capable of matrix degradation and activation of progelatinase A, called the membrane-type MMPs. Four members have been identified to date: MT1-, MT2-, MT3-, and MT4-MMP. 7,8 Among the MT-MMPs, MT4 is distinguished from the others because of its low sequence homology compared with the homologies of the other MT-MMPs, and it is unclear whether it can activate gelatinase A. 7 Although the role of the MT-MMP family in tumor cell invasion remains unclear, in situ hybridization studies and immunolocalization studies in breast, colon, gastric, and head and neck cancer have identified the presence of MT1-MMP overexpression. 9,10 These studies have localized MT1-MMP to tumor cells at the tumor-stromal interface. The presence of MT2-MMP and MT3-MMP has not been identified with the same frequency. 10 The pathophysiologic role of MT1-MMP in tumor cell invasion, however, remains poorly understood.
MT1-MMP is thought to mediate tumor cell invasion directly by proteolyzing ECM components as well as indirectly through gelatinase A activation. The substrate specificity of MT1-MMP appears to be similar to that of interstitial collagenase. It has been shown to cleave proteoglycans, fibronectin, vitronectin, and laminin-1, as well as collagen (types I, II, and III), into typical 3/4 and 1/4 fragments. 11 The ability of MT1-MMP to convert progelatinase A to gelatinase A further increases the spectrum of tumor cell-directed proteolysis. Furthermore, because MT1-MMP is anchored to the cell membrane and gelatinase A is known to have cell surface receptors, 12 it is thought that these 2 proteases focus the degradation cascade close to the cell surface. Other MMPs are secreted into the interstitial space without localized activation or enzymatic activity; thus their role in tumor cell migration is less clear. Therefore the MT1-MMP/gelatinase A protease system is considered to play a key role in tumor cell-directed cell proteolysis and migration. 13 To examine the role of these enzymes in tumor cell-mediated ECM invasion, populations of an HNSCC cell line, UM-SCC-1, were transfected with either gelatinase A or a catalytically active form of MT1-MMP. Transfected cells were then assessed for invasive potential in vitro on a type I collagen construct.
METHODS AND MATERIAL
Cell Culture
The human squamous carcinoma cell line UM-SCC-1 (generously provided by T. Carey, University of Michigan) was routinely maintained in minimal essential media supplemented with L-glutamine (2 mmol/L), penicillin (100 U/mL), and streptomycin (100 μg/mL) (all from Life Technologies, Gaithersburg, MD), as well as 10% heat-inactivated fetal bovine serum (Hyclone Laboratories Inc, Logan, UT). Cells were cultured in humidified 5% CO2/95% air at 37°C.
Cell Invasion Assays
Rat tail type I collagen (Sigma, St Louis, MO) was dissolved in 0.2% acetic acid at 2.2 mg/mL and dialyzed against Tris-buffered saline (pH 7.6) at 4°C. Collagen (0.8 mL) was gelled at 37°C for 40 minutes in 24-mm Transwell dishes (3 μm pore size; Corning Costar Corp, Cambridge, MA) before addition of 2 × 10 5 cells/well and media. Cells were grown to confluence, and recombinant human EGF (3.5 nmol/L; Collaborative Biomedical Products, Becton Dickinson Labware, Bedford, MA) was added to the lower chamber of the Transwell dishes.
To assess the role of proteinases in the invasive process, cells grown atop collagen gels were incubated with proteinase inhibitors that were added to the upper and lower compartments of the Transwell dishes. Media were changed every 3 days, and fresh inhibitors were added. The inhibitors used included BB-94 (Batimastat; [4-(N-hydroxyamino)-2R-isobutyl-3S-(thienylthiomethyl)-succinyl]-L-phenylalanine-N-methylamide; a gift of British Biotechnology, Oxford, UK) and recombinant tissue inhibitor of metalloproteinase-2 (TIMP-2; a gift of Amgen, Thousand Oaks, CA). TIMP-2 was dissolved in Hank's buffer (Life Technologies), whereas BB-94 was dissolved in dimethylsulfoxide (used at a final solvent concentration of 0.1%).
Northern Blot Analysis
For Northern blot analysis, total RNA (10 μg) was electrophoretically fractionated in 1% denaturing agarose gels and transferred to nylon membranes (Hybond N+; Amersham Life Science Corp, Arlington Heights, IL). Blots were prehybridized for 30 minutes at 68°C in QuikHyb (Stratagene, La Jolla, CA) and then hybridized for 3 hours at 68°C using [32P]dCTP-labeled cDNA probes (Rediprime Labeling System, Amersham Life Sciences Corp) for human EGF receptor, stromelysin-1, collagenase-1, gelatinase A, gelatinase B, or MT1-MMP using 1.4-, 0.8-, 0.3-, 0.75-, 2.0-, and 2.9-kilobyte cDNA fragments, respectively. Hybridized blots were rinsed twice in 2 × standard saline citrate (SSC)/0.1% sodium dodecylsulfate (SDS) at 68°C, washed for 30 minutes at 68°C in 0.1 × SSC/0.1% SDS, and subjected to audioradiography at −80°C for 4 to 24 hours. To assess RNA integrity and relative loading, the blots were probed with 36B4 or GAPDH.
Western Blot Analysis and Zymography
Western blots were performed as described. 14 In brief, samples were separated by 8.5% SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and immunoblotted with rabbit antiserum to either human interstitial collagenase or human stromelysin-1 antiserum (both gifts of H. Welgus, Washington University, St Louis, MO). Zymography was performed as described using 8.5% SDS-PAGE impregnated with gelatin (2 mg/mL). 15 MT1-MMP activity was assessed by its ability to process recombinant progelatinase A (gift of R. Fridman, Wayne State University, Detroit, MI) as described previously. 15
Vector Construction and Transfection of UM-SCC-1 Cells
The pCR3.1 plasmid (Invitrogen, San Diego, CA) for expression of ΔMT1-MMP and gelatinase A were constructed by manipulation of the cDNAs of the proteins as described. 16 In brief, full-length gelatinase A was generated by polymerase chain reaction from an HT-1080 cDNA library cloned into Lambda ZAP-Express (Stratagene) using oligonucleotides to ATG and UGA sites. The ΔLMT1-MMP insert was generated from an HT-1080 cDNA library cloned in Lambda ZAP-Express with the coding sequence for the cytosolic tail deleted as described. 16 Higher levels of expression were found with the cytosolic segment deleted. Cells were transfected with the pCR3.1 plasmid lacking an insert (control vector) or containing sequences encoding ΔMT1-MMP or gelatinase A using the LipofectAMINE method (Life Technologies). Transfected cells were cultured for 48 hours and then incubated for 24 hours with serum-free supernatant before assessment of protein expression by zymography as described above. Over-expressing clones derived from single cell colonies were chosen for the control vector group, ΔMT1-MMP- and progelatinase A-transfected cells.
Sample Fixation and Processing
Collagen gels were fixed in 2% glutaraldehyde and 1.5% paraformaldehyde in 0.1 mol/L sodium cacodylate buffer and then removed from the Transwell culture plates. Before being embedded in epoxy resins, the samples were cut into sections and postfixed in 1% osmium tetroxide (in 0.1 mol/L sodium cacodylate). Sections, 0.5 μm thick, were stained with toluidine blue before examination by light microscopy. Tumor cell invasion was assessed by light microscopy in a minimum of 4 to 6 randomly selected sections for each experimental sample.
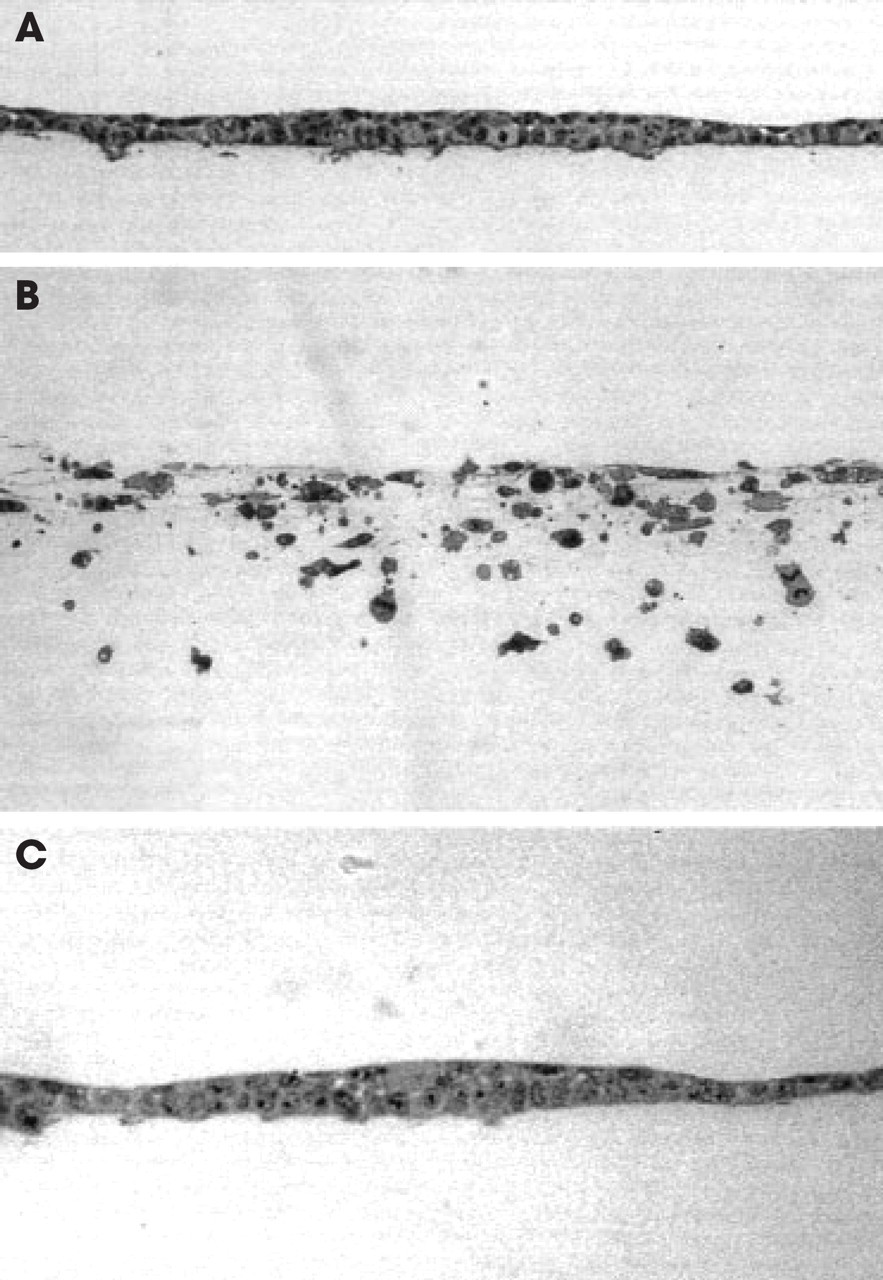
MMP inhibitors block EGF-stimulated (3.5 nmol/L) UM-SCC-1 invasion. Transverse light sections of cells cultured atop type I collagen gels were incubated for 6 days without exogenous growth factors (
RESULTS
EGF-induced Invasion
Confluent monolayers of UM-SCC-1 cultured atop type I collagen gels constitutively expressed EGF receptor (data not shown). In the presence of optimal doses of EGF (3.5 nmol/L), cells began to express a collagen-invasive phenotype at 48 hours, which peaked after a 6-day incubation period. Whereas unstimulated cells expressed no invasive behavior (Fig 1A), single cell invasion was obtained with cells continuously exposed to EGF (Fig 1B). The ability of UM-SCC-1 cells to penetrate a dense collagen barrier is consistent with expression of proteolytic activity.
Recent studies have suggested MMPs may regulate cell motility by proteolyzing either membrane-associated components or the underlying matrix as a prelude to displaying invasive behavior. 11,16 To determine the role of MMPs in regulating the expression of invasive behavior, UM-SCC-1 cells were stimulated with EGF to invade collagen matrices in the presence of MMP inhibitors. Both natural and synthetic MMP inhibitors (TIMP-2 and BB-94, respectively) completely blocked invasion though the type I collagen construct during a 6-day incubation period (Fig 1C).
MMP Expression During Invasion
The expression of MMPs in EGF-stimulated collagen invasion was characterized by isolation of total RNA from 6-, 12-, 24-, and 48-hour cultures and subjection to Northern analysis. Collagenase-1, gelatinase B, and stromelysin-1 were identified by Northern analysis, but gelatinase A, stromelysin-2, stromelysin-3, MT2-MMP, and MT3-MMP were not detected (data not shown). Moreover, serum-free supernatant collected from UM-SCC-1 cells in the presence or absence of EGF did not contain gelatinase A. Gelatinase A, however, is present at high concentrations in fetal bovine serum, and invasion assays were conducted in the presence of medium with 10% serum (see Methods). Serum-free supernatants were collected and assessed for collagenase-1, stromelysin-1, and gelatinase B by Western blot and zymography (Fig 2). These enzymes were not, however, present in the active form. Glycosylated forms of stromelysin-1 and collagenase-1 appear in the 2 distinct glycosylated forms separated by 2 to 3 kDa.
The MT1-MMP transcript could be detected at low levels in unstimulated UM-SCC-1; however, mRNA levels increased in a complex temporal fashion in response to EGF (data not shown). As a membrane-anchored enzyme, MT1-MMP protein could not be easily quantitated during the invasion assay, but the presence of catalytically active MT1-MMP protein could be detected by virtue of its ability to catalyze progelatinase A to its active form. Thus resting or stimulated UM-SCC-1 atop type I collagen gels was incubated with exogenous recombinant gelatinase A, the supernatant was recovered, and the processing of the proenzyme was assessed by gelatin zymography (data not shown). Progelatinase A conversion to its 62-kDa active species was significantly increased in the invading cultures, suggesting a role for the MT1-MMP/ gelatinase A protease system in collagen invasion.
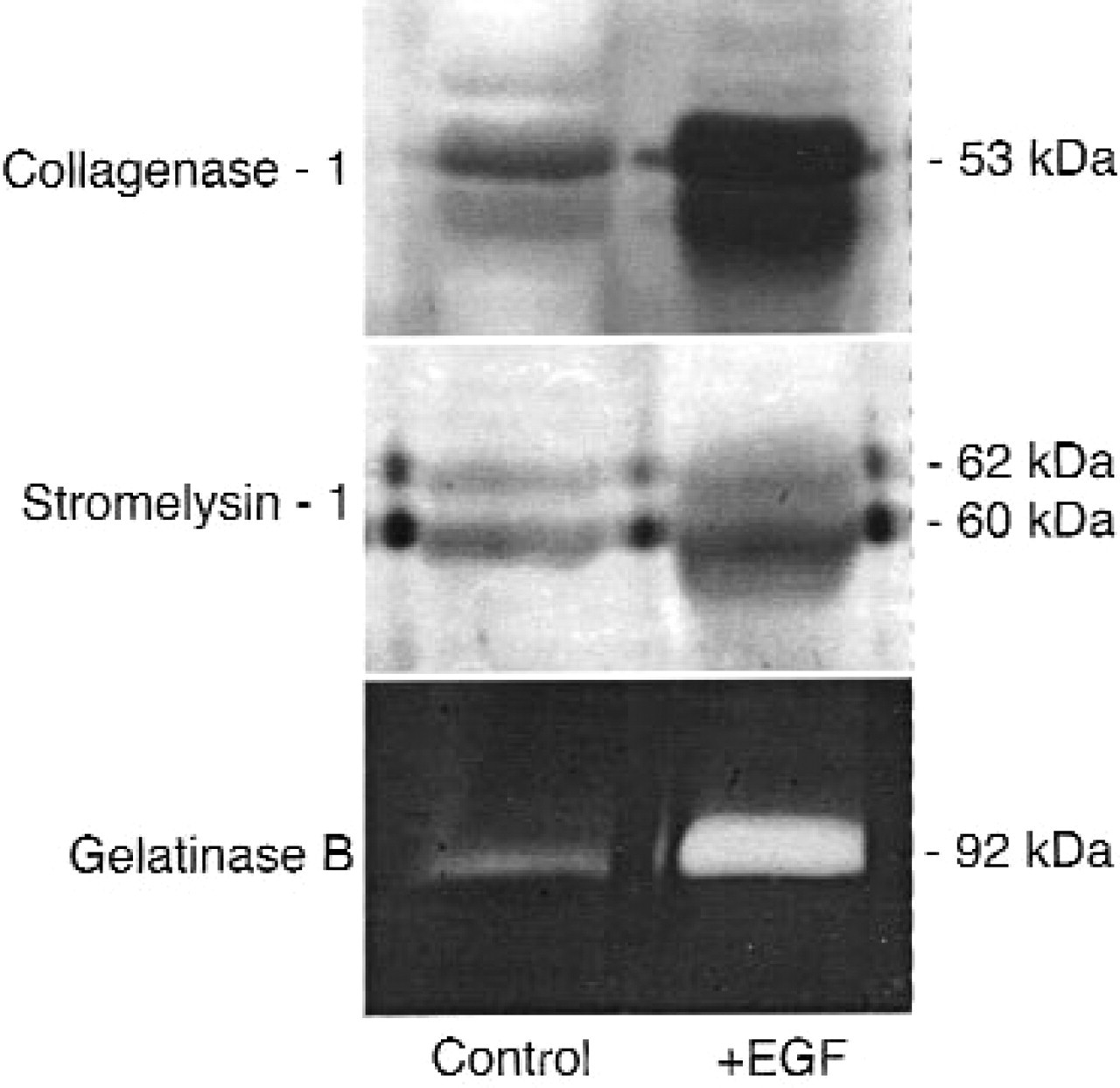
Collagenase-1, stromelysin-1, and gelatinase B are not in the catalytically active form in EGF-stimulated UM-SCC-1 cells. Western blot analysis (collagenase-1, stromelysin-1) and gelatin zymography (gelatinase B) of 24-hour, serum-free conditioned media from cells cultured atop collagen in the absence or the presence of EGF. Collagenase-1 and stromelysin-1 are detected as glycosylated and nonglycosylated doublets on Western blots. Results are representative of 2 or more independent experiments.
Role for MT1-MMP in Invasion
To test the hypothesis that EGF induction of MT1-MMP is a critical element of the invasive phenotype, we augmented MT1-MMP cell surface activity using a eukaryotic expression vector for the transmembrane and catalytic portion of MT1-MMP. A cytosol-deleted, ΔMT1-MMP construct was made with the cDNA encoding the transmembrane and extracellular domain and was inserted into pCR3.1 in the sense direction (Fig 3). Previous studies from our laboratory have demonstrated that the MT1-MMP protein can efficiently proteolyze ECM components and activate gelatinase A with the cytosolic portion deleted. 16 The pCR3.1 vector uses a cytomegalovirus promoter to drive constitutive expression and contains a selectable marker for neomycin resistance. Selection using neomycin was applied for 3 weeks, and single cell clones of ΔMT1-MMP and control vector cells (pCR3.1 vector alone) were selected. Populations of transfected cells were tested for ΔMT1-MMP expression by addition of exogenous gelatinase A to unstimulated, transfected cells, and supernatants were analyzed by gelatin zymography. Conversion of progelatinase A to gelatinase A in the absence of EGF indicated proteolytically active ΔMT1-MMP protein. Clones with high (A), intermediate (B), or low (C) expression were selected (Fig 3).
The invasive potentials of ΔMT1-MMP clones (A, B, or C) were assessed under unstimulated and EGF-stimulated conditions. Invasion was graded by the same observer according to the number of cells within the matrix and depth of penetration. Cells were cultured atop type I collagen and assessed every 24 hours by phase microscopy for invasion. In the absence of EGF, UM-SCC-1 cells overexpressing ΔMT1-MMP invaded the collagen substratum during the 96-hour incubation time, whereas control vector cells remained a confluent monolayer (data not shown). Greater invasion was noted in the EGF-stimulated SCC-1/MT cells (clone A) compared with stimulated vector control cells (Fig 4). A relative decrease in invasive potential was noted in clones with decreasing expression of ΔMT1-MMP, such that SCC-1/MT clone B was more invasive than SCC-1/MT clone C, but less invasive than UM-SCC-1 clone A (data not shown).
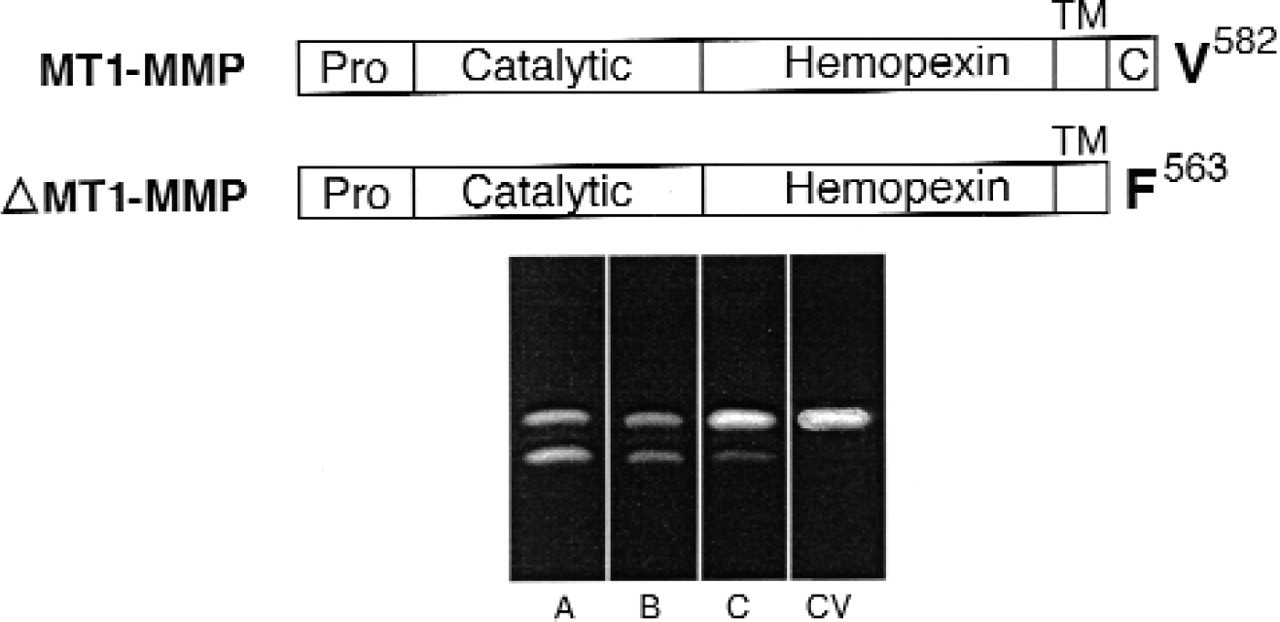
Expression and characterization of SCC-1/MT1 cells. Top, Domain alignments of wild-type MT1-MMP and ΔMT1-MMP Wild-type MT1-MMP is arranged into pro-, catalytic, hemopexin, transmembrane, and cytosolic domains. The ΔMT1-MMP construct is truncated at the edge of the cytosolic domain. Bottom, Zymography of recombinant gelatinase A (140 nmol/L) incubated with cells stably transfected with the ΔMT1-MMP construct (clones A, B, and C) or control vector (CV) cells atop collagen. Increased conversion of the zymogen to the lower molecular weight species indicates increased ΔMT1-MMP expression. Results are representative of 2 or more independent experiments.
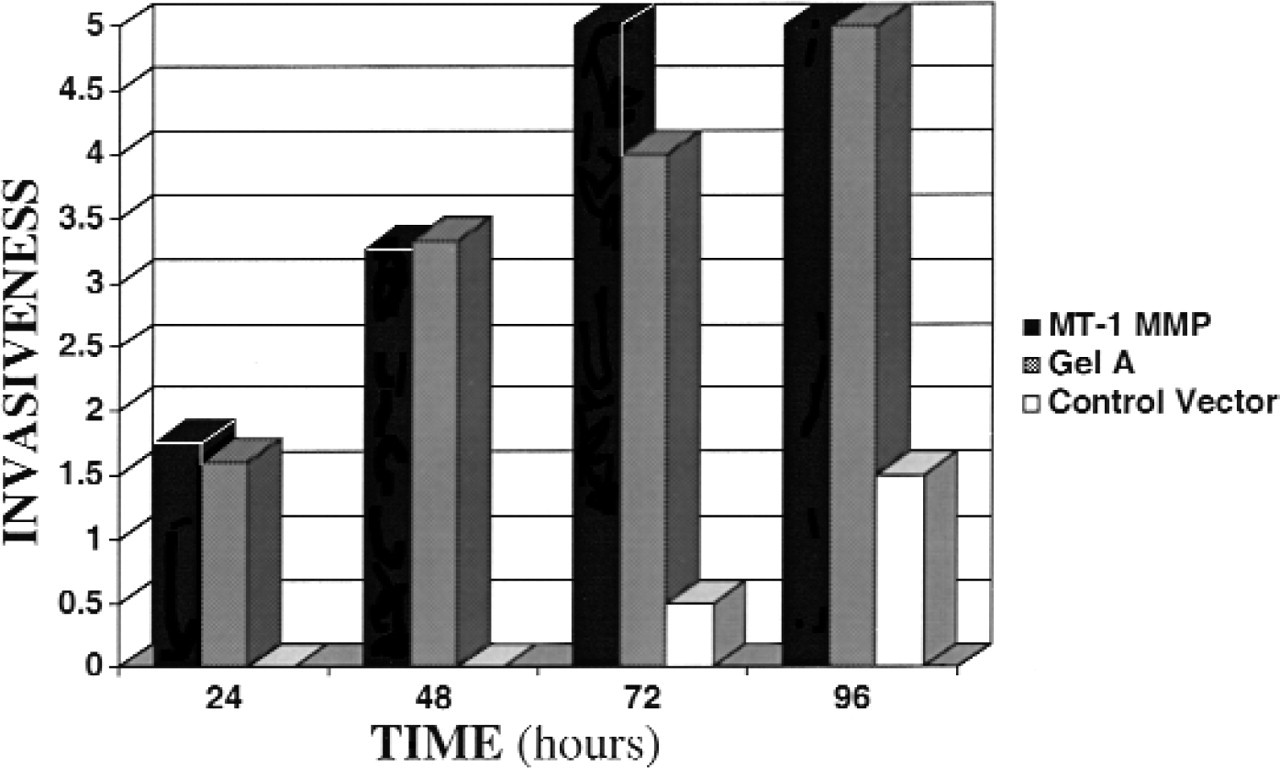
Invasive potential of transfected cell lines. SCC-1/MT (black), SCC-1/gelA (shaded), and control vector (white) transfected cells were assessed under EGF-stimulated conditions for 96 hours. Invasiveness was measured by phase microscopy at 24-hour intervals and scored according to number of cells invading and depth of penetration. Representative of 2 independent experiments using SCC-1/MT clone A and SCC-1/gelA clone A.
Role for Gelatinase A in Invasion
MT1-MMP has recently been described to directly degrade matrix components including type I collagen. To determine whether enhanced invasive potential of UM-SCC-1 can be induced with elevated levels of gelatinase A, we inserted the full-length cDNA of gelatinase A into the eukaryotic expression vector, pCR3.1, in the sense direction and transfected it into UM-SCC-1 cells as described above. Overexpression was determined by gelatin zymography (Fig 5). Clones of intermediate or low expression could not be successfully selected. High levels of gelatinase A expression resulted in substantial expression of the activated, 62-kDa species, despite the absence of MT1-MMP upregulation by EGF. The abundance of gelatinase A produced by SCC-1/gelA transfectants allows for the conversion of a small percent, but a large total quantity, of activated gelatinase A (Fig 5).
Three gelatinase A overexpressing clones were assayed on type I collagen using phase microscopy at 12-hour intervals during a 72-hour time course and compared with vector-only controls. As before, unstimulated cells exhibited some evidence of invasion, and stimulated overexpressing cells exhibited deeper penetration of the matrix and more areas of invasion (Fig 4).
DISCUSSION
The biomechanical program tumor cells use to penetrate the ECM is poorly understood. We have developed an in vitro invasion assay using acid-solubilized, fibrillar type I collagen gels that cannot be penetrated by HNSCC tumor cell lines, except under growth factor-stimulated conditions. In this article we show that invasion of this construct occurs in the presence of EGF and that the invasive phenotype is MMP dependent. In a previous report we demonstrated similar results using a nonviable dermal explant model. 6 Analysis of MMP proteins expressed during UM-SCC-1 invasion in vitro revealed that both gelatinase A and MT1-MMP were present and in the activated form. We found that overexpression of either ΔMT1-MMP or gelatinase A increased the invasive phenotype of UM-SCC-1, which suggests that the MT1-MMP/gelatinase A protease system participates in the proteolytic mechanism by which HNSCC tumor cells penetrate stromal barriers (primarily composed of type I collagen).
Analysis of MMP expression in UM-SCC-1 cells during growth factor-stimulated in vitro invasion identified the expression of MT1-MMP mRNA and the activation of gelatinase A. Of the MT-MMP family, MT1-, MT2-, and MT3-MMP are known to activate progelatinase A. Transcripts for MT2- or MT3-MMP, however, were not identified by PCR analysis (data not shown), nor was gelatinase A detected at the message or protein level in UM-SCC-1 cells. As a normal constituent of fetal bovine serum, however, gelatinase A was present during the invasion assays. All MMPs require cleavage of the prodomain before expressing proteolytic function and can be detected in their activated form by the decrease in molecular weight on Western blot or zymography. Although actively invading UM-SCC-1 cells expressed collagenase-1, stromelysin-1, and gelatinase B, none of these proteases was present in the processed form. MT1-MMP and gelatinase A, however, were both detected in the active form, which suggests a role for these enzymes in the invasive program.
The MT-MMP family of proteases was only recently identified as the membrane-anchored proteases capable of converting progelatinase A to the active, 62-kDa species. The MT-MMP family is currently thought to be important in tumor cell invasion because (1) it is capable of progelatinase A activation, (2) it has intrinsic matrix degrading function similar to that of collage-nase-1, and (3) as a membrane-bound protein it is thought to localize tissue degradation to the cell surface. 13 Although the activation mechanism of the MT-MMP family remains an area of investigation, this zymogen is known to be processed intracellularly and expressed only in the active form on the cell surface. 16 The importance of MT1-MMP localization to the cell surface has recently been demonstrated by Nakahara et al. 12 In this study an MT1-MMP mutant was constructed lacking the transmembrane domain, and although catalytically active, it was unable to localize to the cell surface. Compared with wild-type MT1-MMP, cells expressing soluble MT1-MMP showed a decreased degradative potential in vitro. 12
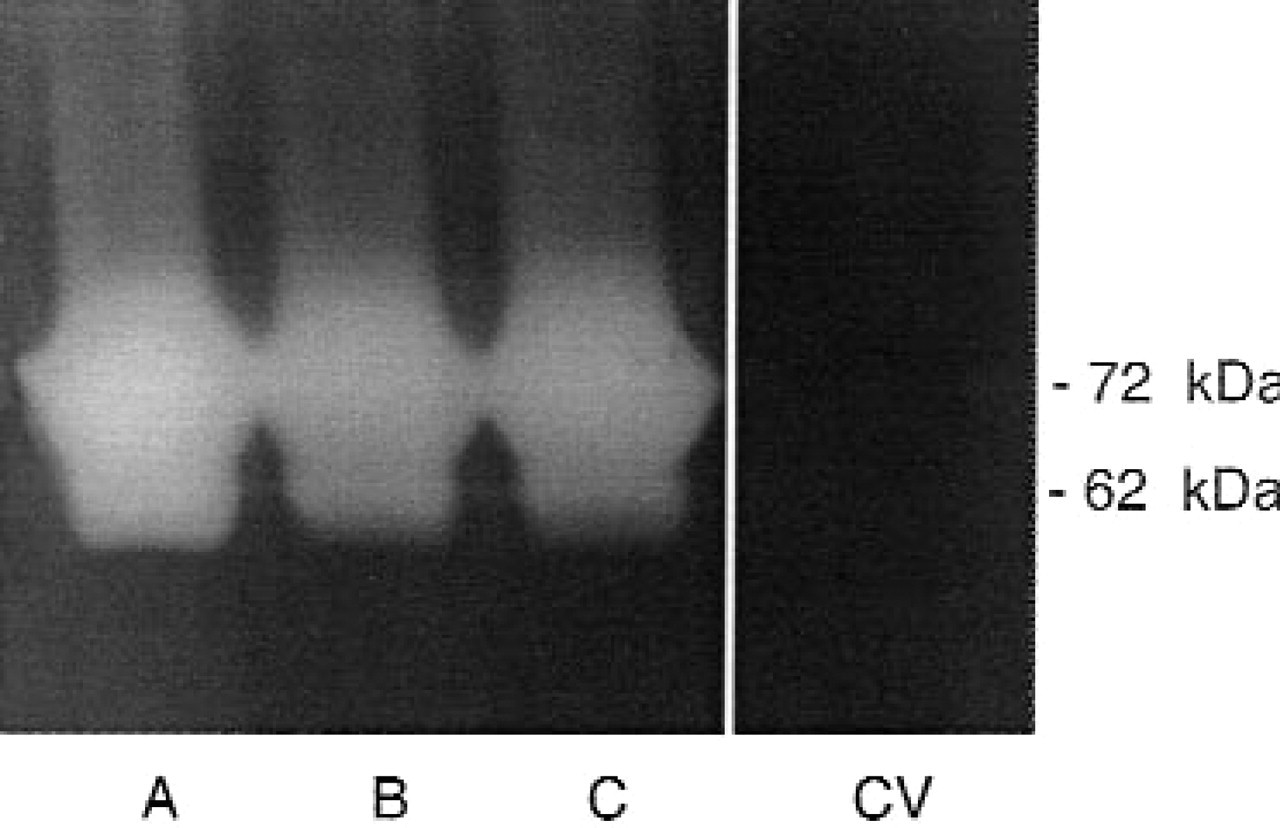
Expression and characterization of SCC-1/gelA transfectants. Zymography of 12-hour, serum-free conditioned media from cells stably transfected with the gelatinase A construct (clones A, B, or C) or control vector (CV). Results are representative of 2 or more independent experiments.
In the absence of growth factors, wild-type UM-SCC-1 or control vector cells failed to display a minimally invasive phenotype. Addition of EGF, however, stimulated invasion coincident with MT1-MMP expression. A full-length MT1-MMP expression vector was constructed but could not be successfully transfected into UM-SCC-1. Overexpression of ΔMT1-MMP (cytosol tail deleted) led to spontaneous invasion under unstimulated conditions and accelerated invasion (compared with vector controls) in the presence of EGF. Moreover, higher levels of SCC-1/MT overexpression positively correlated with increased invasive potential in the 3 different clones. Previous immunochemical in situ studies have found elevated levels of activated gelatinase A and increased MT1-MMP (but not MT2- or MT3-MMP) expression in 100% of HNSCC tumor samples examined. 10 Although MT1-MMP was initially thought to be overexpressed in stromal cells at the tumor-stromal interface, recent in situ studies have localized MT1-MMP transcript and protein to the tumor cells at the advancing edge of tumors. Our findings suggest the MT1-MMP expression or gelatinase A is sufficient for stromal invasion.
Because MT1-MMP is capable of degrading type I collagen and activating progelatinase A, we investigated whether gelatinase A participates in collagen matrix invasion. Full-length gelatinase A was overexpressed in UM-SCC-1, and the invasive was potential assessed. Because of the gross excess of progelatinase A, a substantial quantity of active gelatinase A was present in the active form even in the absence of EGF. SCC-1/gelA cells invaded under resting conditions and displayed elevated invasive behavior when stimulated with EGF compared with control vector cells. Gelatinase A is known to localize to the cell membrane by binding to cell surface receptors. 12 Although direct MT1-MMP degradation of the ECM remains probable in this in vitro assay, these results suggest that the MT1-MMP/ gelatinase A cascade mediates HNSCC invasion through collagenous matrices. The relative importance of these 2 enzymes in the invasive program requires further investigation.
The increased expression of MMP genes is recognized as a tissue-specific phenomenon in physiologic and pathologic processes. For example, studies have shown gelatinase B to be the critical enzyme in metastatic behavior of transformed rat embryo cells. 17 Stromelysin-1 and stromelysin-2 participate in keratinocyte migration in human wound healing. 18 The coexpression of several MMPs appears to be a common feature of malignancies. In addition to MT1-MMP, increased expression of collagenase-1, stromelysin-2, and stromelysin-3 has been identified in HNSCC tumor samples. It is likely that MT1-MMP and gelatinase A work together in a complex manner to degrade the ECM and facilitate invasion.
Results from this study show that overexpression of the MT1-MMP/gelatinase A protease system can increase the invasive potential of HNSCC in vitro. This work also supports the possible role of MT1-MMP and gelatinase A as potential markers of aggressive disease, as previously suggested by in situ studies. 9,10 Pharmacologic agents have recently been developed that target MMPs. 2 It is possible that anti-MMP therapy could reduce the rate of tumor cell invasion or metastasis in patients with head and neck cancer.