
Abstract
OBJECTIVES: We tested the hypothesis that mastoid volume buffers the rate of change in middle ear pressure caused by transmucosal, inert gas exchange.
STUDYDESIGN: Twelve monkeys were randomly assigned to group 1 or group 2. Right ears of group 1 had sham surgery and of group 2 had obstruction of the mastoid antrum. Before and after surgery, the time constant for transmucosal N2O exchange was estimated from N2O breathing experiments. The hypothesis predicts that the postoperative time constant measured for right ears of group 2 but not group 1 is greater than that measured before surgery.
RESULTS: Mastoid antrum block significantly decreased right middle ear volume but did not affect the time constant for transmucosal N2O exchange.
CONCLUSION: A mastoid gas-reserve function is not supported by the experimental data.
SIGNIFICANCE: These results for monkeys and the theory developed to explain the effect of mastoid volume on transmucosal inert gas exchange suggest that the results for previous experiments in humans interpreted as evidencing a mastoid gas-reserve function are consistent with alternative explanations.
One hypothesis advanced to explain this relationship is that the mastoid airspace behaves as a gas reserve, thereby lessening the rate of pressure decrease caused by the ME to blood transfer of N2. 6 Specifically, direct measurements show that the ME N2 partial pressure exceeds that of the surrounding mucosa (blood) by approximately 50 mm Hg. 7 In the absence of gas resupply during eustachian tube (ET) openings, that gradient will cause a net N2 efflux from the ME and will drive total ME pressure to subatmospheric pressure levels sufficient to provoke mucosal inflammation and effusion. 8 For a relatively fixed-volume system such as the ME, adding or removing a specified volume of gas at a given rate (as in transmucosal transfers) will cause a greater rate of pressure change in small volume systems compared with large volume systems. Thus, at identical N2 efflux rates and ET opening frequencies, larger-volume MEs will have a corresponding lesser rate of total pressure change and a lesser probability of achieving the threshold underpressure that precipitates OME by hydrops ex vacuo. However, like many aspects of ME physiology, this hypothesized buffering mechanism has not been examined critically. 6
An increase in ME pressure was reported for anesthetized patients breathing gas mixtures that included N2O, and this was attributed to gradientdriven N2O transfer from blood to ME. 9,10 Like N2, N2O is an inert gas whose rate of exchange across the ME mucosa is primarily perfusion limited. 11 The hypothesis of a gas-reserve function for the mastoid can be tested using data for the ME pressure response to established blood-ME N2O pressure gradients under the expectation that the rate of pressure change is greater for MEs with smaller mastoid volumes. In one such study of patients with OME, no change in ME pressure was observed for ears with “nonaerated” mastoids. 12 In contrast, 3 studies of patients without otoscopic or tympanometric evidence of concurrent ME disease reported a greater rate of pressure increase for ears with smaller mastoid volumes. 13–15 These studies were all done for N2O breathing in the surgical setting where the inherently poor control over variables that affect the rate of ME pressure change such as gas mixture, mucosal blood perfusion, body temperature, extant partial-pressure gradients, and forced tubal opening frequency could have contributed to the ME pressure behavior. Also, subject age and OME history can independently affect certain parameters (eg,. arterial N2O transients and mucosal perfusion) that control the rate of transmucosal N2O exchange, 16,17 but the confounding effects of these factors were not considered in the data analyses for any of the experiments.
In the experiment described here, the effect of mastoid volume on the blood to ME exchange rate of N2O was studied in cynomolgus monkeys under well-controlled conditions. The monkey was chosen for this experiment because of the documented similarity in ME anatomy and function between old-world monkeys and humans. 18 Specifically, and in contrast to canid and rodent species, monkeys have a mastoid antrum and a well-developed, mastoid air-cell system. Also, monkeys were used previously to model gas exchange between ME and blood. 11,19 ME volume was reduced in one group of animals by unilateral blockage of the mastoid antrum, while a second group served as a sham-operated control. The time constant for N2O exchange was estimated from the data acquired during breathing experiments that used gas mixtures containing 5% N2O. 19 The expectations under the hypothesis of a mastoid gas-reserve function are that the time constant for transmucosal N2O exchange (as a surrogate for N2 exchange) measured for the antrum blocked MEs is greater than that measured in (1) the same ears before surgery, (2) the contralateral, control MEs, and (3) the sham-surgery MEs.
MATERIALS AND METHODS
Protocol
Twelve juvenile cynomolgus monkeys weighing between 2.8 and 4.2 kg were used in the study. The animals were quarantined for 45 days and were examined by otoscopy and tympanometry to document bilaterally normal ME status. Over a 1-month period, 2 experiments (separated by at least 1 week) were conducted using a 5% N2O gas mixture, and baseline transmucosal N2O time constants were calculated from these data. Then, the animals were randomly assigned to 1 of 2 equal groups (group 1, control group, sham surgery; group 2, experimental group, mastoid antrum block), and the designated surgical procedure was performed on the right ear as described later. Weekly, the ears of all monkeys were examined by otomicroscopy and tympanometry to assess postoperative ME status. After documentation of a normal right tympanogram, a postoperative experiment was performed on all animals using a 5% N2O gas mixture and repeated approximately 1 week later. Postoperative time constants for transmucosal N2O exchange were calculated from these data. This study was performed in accordance with the PHS Policy on Humane Care and Use of Laboratory Animals, the NIH Guide for the Care and Use of Laboratory Animals, and the Animal Welfare Act (7 USC et seq). The study protocol was approved by the Animal Care and Research Committee at the Children's Hospital of Pittsburgh.
Surgical Procedures
All surgical procedures were performed by an ENT surgeon (C.M.A.) in an operating room where sterile conditions were maintained. Before surgery, the monkey was sedated with ketamine (12 to 15 mg) and weighed, and ME status was evaluated bilaterally using otomicroscopy and tympanometry. Then, the monkey was anesthetized by an intramuscular injection of “monkey mix” (10 mg/kg, ketamine 2 mg/kg xylazine, and 0.3 mg/kg acepromazine). The area over the right ME was shaved and prepped, the left hind leg was shaved to safely ground a cautery unit, an intravenous line was established in the right leg, and a preoperative antibiotic (25 mg/kg Bactrim IM) was administered. The monkey was intubated with a 3–0 endotracheal tube and placed on the operating table, and the surgical site was scrubbed twice with povidone-iodine (Betadine) and draped. Lidocaine (1% with epinephrine 1:100,000) was injected into the postauricular sulcus. An incision was made through the skin and subcutaneous tissues overlying the sulcus, the auricle was retracted, and the external auditory canal and temporal line were identified. The underlying periosteum was incised and elevated to expose the mastoid cortex, and the antrum was opened at the McEven's triangle using a drill. Throughout, Bo-vie cautery was used to control bleeders.
For animals assigned to the control group (group 1), a piece of titanium mesh (1.3-mm TI; Synthes, Monument, MO) was cut to fit and cover the antrum at the subcortical level. Residual bone fragments were removed by rinsing with saline followed by suction, and then the area was flooded with lidocaine (1% with epinephrine 1:100,000) to reduce bleeding. The drill site was completely dried using suction and swabs, and hydroxyapatite cement (HAC) (Bone Source HA Cement; Osteogenics Inc, Richardson, TX) was prepared and placed over the titanium mesh to bring the surface to the level of the mastoid cortex.
Animals assigned to the experimental group (group 2) underwent a more extensive drilling. After surface preparation as described above, the air cells of the entire mastoid were exposed. Then, a drill was used to remove the periantral, sinodural, zygomatic, apical, and perisinus air cells, thereby completing a simple mastoidectomy. The short process of the incus was identified, and the titanium mesh was cut to fit into and cover the aditus ad antrum. The drill site was washed and suctioned to remove bone fragments, and the region was flooded with lidocaine. After the area was dried with suction and swabs, the mastoid cavity was filled with HAC to the level of the mastoid cortex. Heart rate, oxygen saturation, and body temperature were monitored throughout the surgical procedure.
For animals in both groups, a JP drain (TLSR Surgical Drainage System 7 F Round; Porex Surgical Inc, College Park, GA) was placed into the postauricular region, and the subcutaneous tissue was closed in layers with 4–0 Vicryl, finishing with 3–0 Prolene for the skin. A Tegaderm (3M Health Care, St Paul, MN) dressing was affixed with Mastisol to cover the auricular incision site. The JP vacuum drain tube was changed after 1 hour and removed 4 hours postoperatively. The monkey was observed for 1 hour after recovery from the anesthesia and then returned to the vivarium. An antibiotic (25 mg/kg Bactrim IM) was administered twice a day for 5 days. Sutures were removed at 10 days, and the ears were examined weekly by otomicroscopy and tympanometry.
N2O Breathing Experiments
N2O breathing experiments were conducted as previously described. 19 Briefly, the monkey was sedated with 30 mg Ketamine and anesthetized with “monkey mix.” The monkey was monitored for bilateral ME pressure, temperature, and vital signs over a 60-minute period (10-minute intervals) to allow for physiologic acclimation to the anesthetized state. Then, the monkey was intubated, and the endotracheal tube was placed online to the output of a Harvard respirator (model 661; South Natick, MA). The respirator was adjusted to deliver 30 cc gas per stroke at 40 strokes/min. The animal breathed room air for 60 minutes. Then, the intake to the respirator was switched to the experimental gas mixture, and for 60 minutes the animal breathed 5% N2O/20% O2/balance N2 at ambient pressure. Throughout the 120 minutes of controlled breathing, ME pressure was recorded bilaterally at 5–minute intervals using a clinical tympanometer (GSI-33 Middle Ear Analyzer, Model 1733; Littleton, MA). Vital signs and rectal temperature were recorded at regular intervals. At the termination of the experimental session and after documented recovery from the anesthesia, the monkey was returned to the vivarium.
Measurement of ME Volume
After collection of data for all N2O breathing experiments, a myringotomy was made in the anteroinferior quadrant of the tympanic membrane of the right and left ears. A probe was sealed in the ear canal and attached to a pressure transducer and, by closed valve, to a known test volume. The probe and ME were equilibrated to ambient pressure, and an overpressure was applied to the test volume. The valve was opened, and the system pressure was recorded. The volume of the ME and measurement system was calculated using Boyle's law, and ME volume was calculated by subtracting the known volume of the measurement system.
Mathematical Description of Transmucosal Inert Gas Exchange
Under certain conditions, the primary outcome variable for these experiments, change in ME pressure, is equal to the change in ME N2O pressure. We present a mathematical description that defines these conditions and the methods for calculating the time constant for simple, perfusion-limited N2O exchange between a fixed-volume, temperature-stable gas pocket such as the ME and local blood.
The pressure of any closed, gas-filled compartment such as the ME (e) is defined by the general gas law
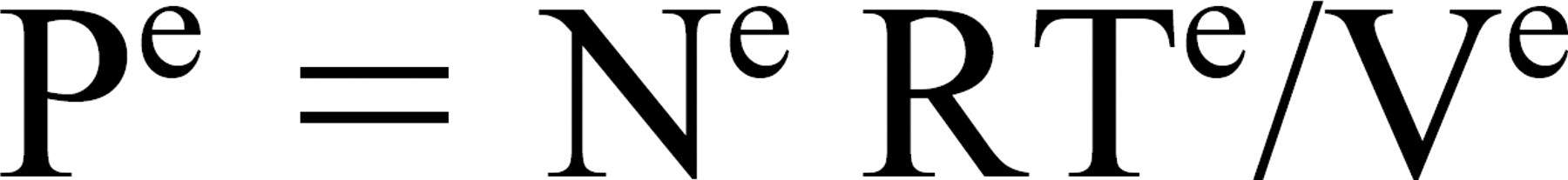
where Pe is pressure, Ve is volume, Ne is number of moles of gas, Te is temperature of the ME, and R is the general gas constant. Total ME pressure is equal to the sum of the partial pressures of the physiologic gases and any represented, nonphysiologic species (eg, N2O), and the change in total ME pressure is equal to the sum of the changes in the partial pressures of those gases
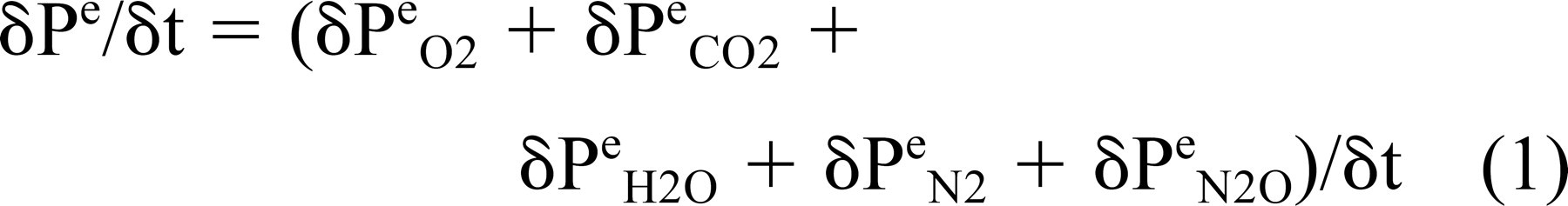
Under physiologic conditions and at constant blood O2 and CO2 pressures, the ME-blood partial-pressure gradient for those gases is approximately 0 mm Hg and the ME is saturated with water vapor. 7 Because there is no gradient to drive ME-blood gas exchange,
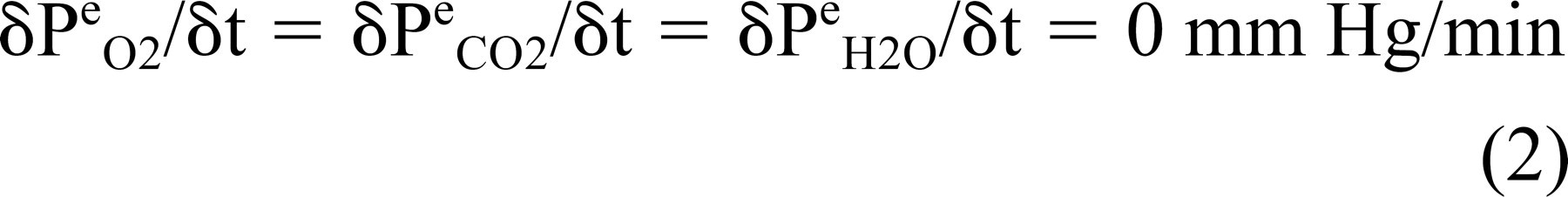
Direct measurement in animals show that the change in ME N2 pressure at ME-blood gradients as high as 50 mm Hg is not measurable in experiments lasting for 4 hours. 11 Consequently, in short experiments, the effective N2 exchange rate is 0 mm Hg/min, and Eq 2 reduces to
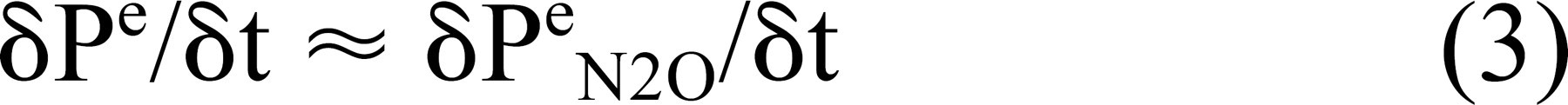
In the experiments described here, an N2O pressure in arterial blood is established at the onset of controlled breathing with the gas mixture. Under that condition, N2O will be transferred from blood to ME. The change in the number of moles of N2O in the ME is equal to the extant difference between the number of moles in the arterial (a) and venous (v) blood of the mastoid (m) and tympanum (t) mucosa
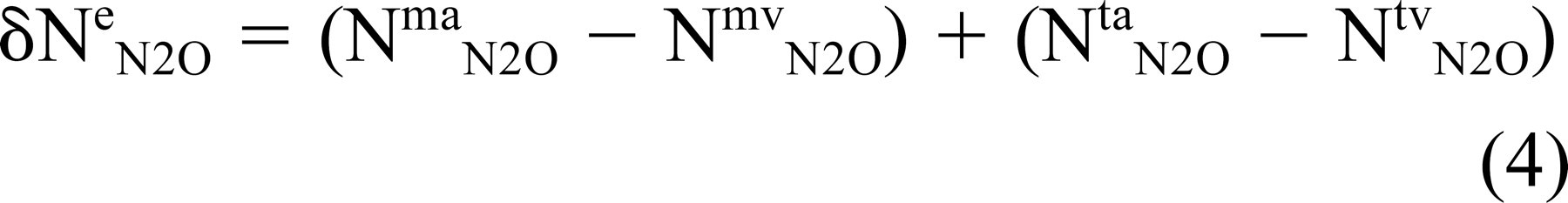
The number of moles of an inert gas in blood is directly related to gas partial pressure, gas solubility in blood (Sb g, equal for arterial and venous blood), and blood volume (Vb), or
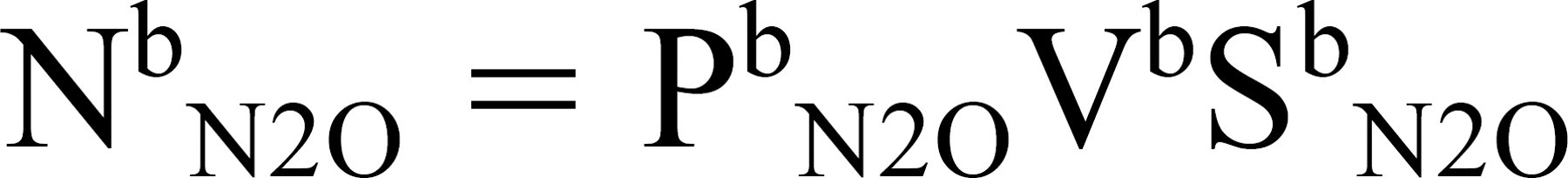
Substituting this and the expression for pressure from Eq 1 for moles of gas in Eq 4 and rearranging terms yields
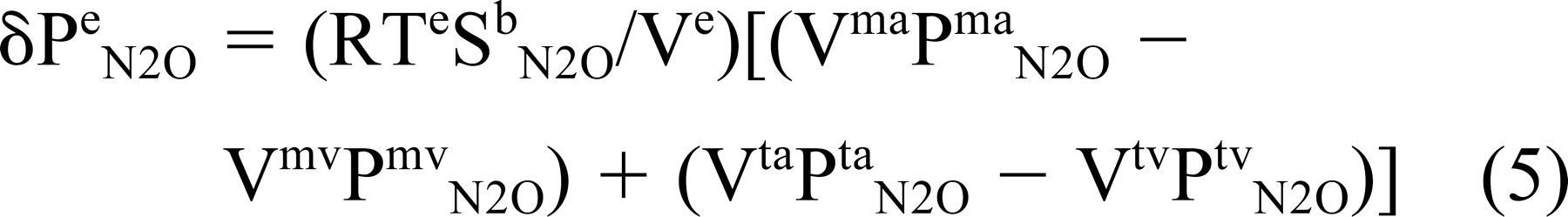
where Ve = Vm + Vt. Dividing both sides of this equation by a time interval (8t) and noting that Pma N2O = Pta N2O and by continuity Vt,ma/δt = Vt,mv/δt = volume blood flow through the bounding mucosa (Qt,m) yields
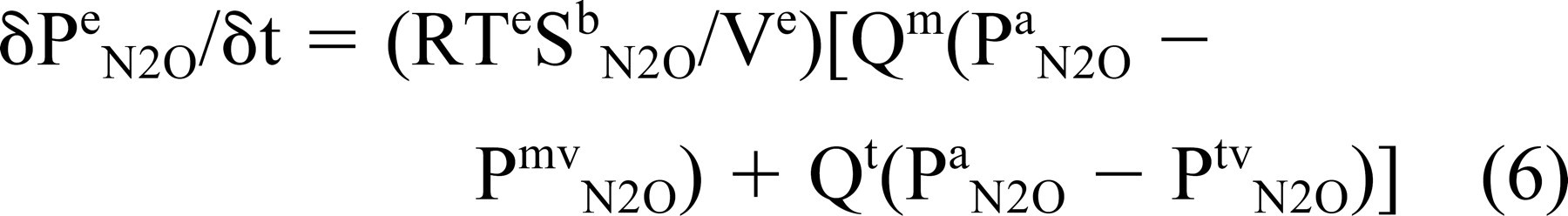
Under the experimental conditions, the pressure of N2O in the ME must at all times be less than or equal to that of the local arterial and venous blood, or
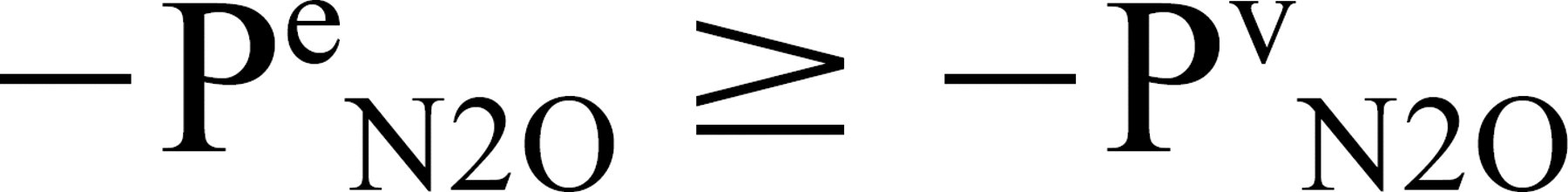
Adding Pa N2O to both sides of this inequality and multiplying by a constant of proportionality (Fe N2O) yields
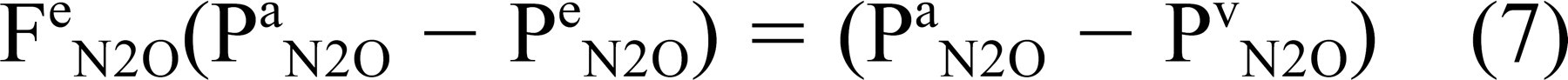
where 1 ≥ Fe N2O ≥ 0. Theory and direct measurements agree that air-phase gas exchange between tympanum and mastoid is very rapid and thus Pm N2O = Pt N2O = Pe N2O Substituting this result and that from Eq 7 into Eq 6 and rearranging terms yields a time constant for transmucosal N2O exchange as given by
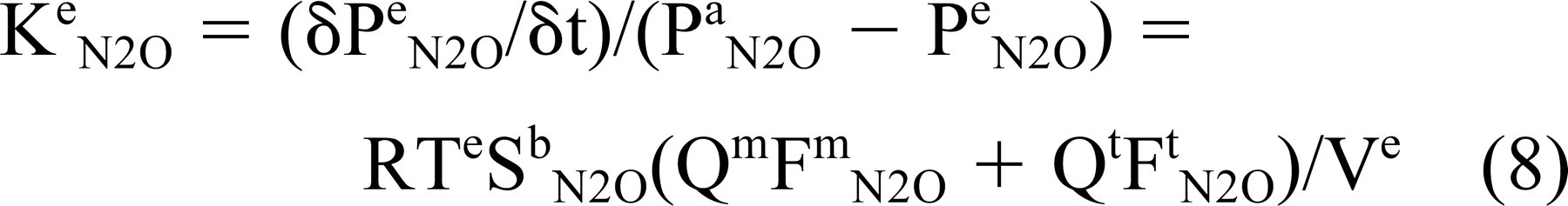
Therefore, at defined N2O pressure gradients, the rate of change in ME pressure is an inverse function of ME volume and a direct function of the sum of adjusted, regional blood flows.
The inclusion of regional blood flow in the time constant calculation complicates the simple relationship between pressure change and volume suggested by Sade and Fuchs
3–5
to underlie pressure buffering by mastoid volume. To appreciate this, note that total volume blood flow for mastoid or tympanum can be decomposed as follows: Qt,m = (Qt,m/At,m)(At,m/Vt,m)Vt,m, where At,m and Vt,m are the surface area and volume of the tympanum or mastoid, respectively. Then, Qt,m/At,m is the average blood perfusion per unit surface area (Rt,m
Q/A), and At,m/Vt,m is the surface area-volume ratio (Rt,m
A/V) for the tympanum and mastoid compartments. These parameters and the volume and gradient ratio can be written in terms of the respective values for the tympanum such that Fm
N2O = UFt
N2O, Vm = XVt, Rm
Q/A = YRt
Q/A, and Rm
A/V = ZRt
A/V where U, X, Y, and Z are scaling constants. Substituting these relationships into Eq 8 and simplifying yields
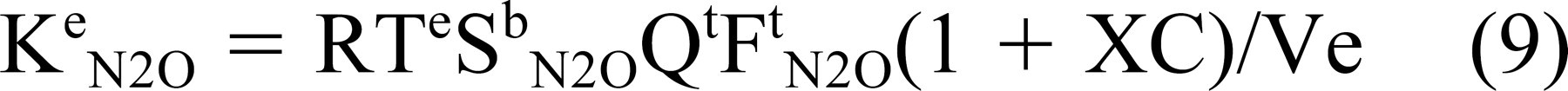
Hypothesis Testing
Time constants were calculated for each experiment using Eq 8:
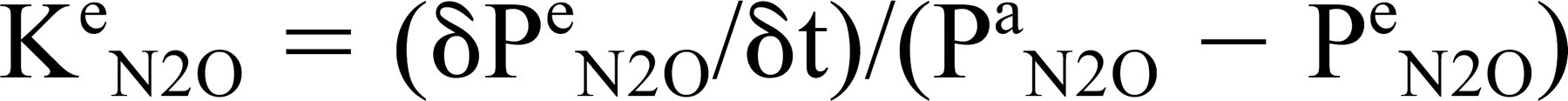
where 8Pe N2O/δt was estimated as the slope of the line relating ME pressure to time of N2O breathing (determined by linear regression) and Pa N2O -Pe N2O was estimated as the percent substitution of arterial N2 by N2O (ie, Pa N2O = 0.05 times blood N2 pressure = 390 mm H2O; Pe N2O ≥ 0 mm H2O). ME pressure as a function of N2O exposure time was modeled by iterating the derived equation (from Equations 3 and 8)
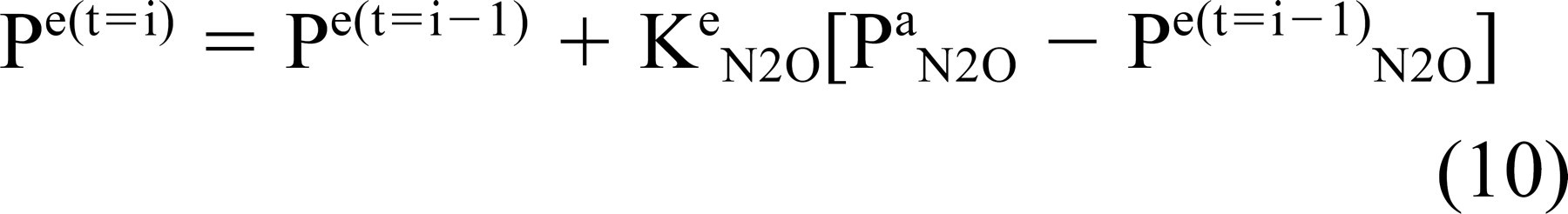
over increasing values of i. The predicted effect of mastoid volume on the time constant was modeled using EQ9 for different, assumed values of C.
The hypothesized mastoid gas-reserve function predicts that the time constant for transmucosal N2O exchange is less for an ME with a mastoid airspace (left ears of all animals, right ears of group 1) compared with an ME without a mastoid airspace (right ears of group 2). Time constants were compared between the preoperative and postoperative experiments for the right ears of group 1 and group 2 using a 1-tailed, paired Student's t test. Measured time constants were also compared with volume-adjusted time constants calculated using the model equation. Throughout, average ± SD values are used.
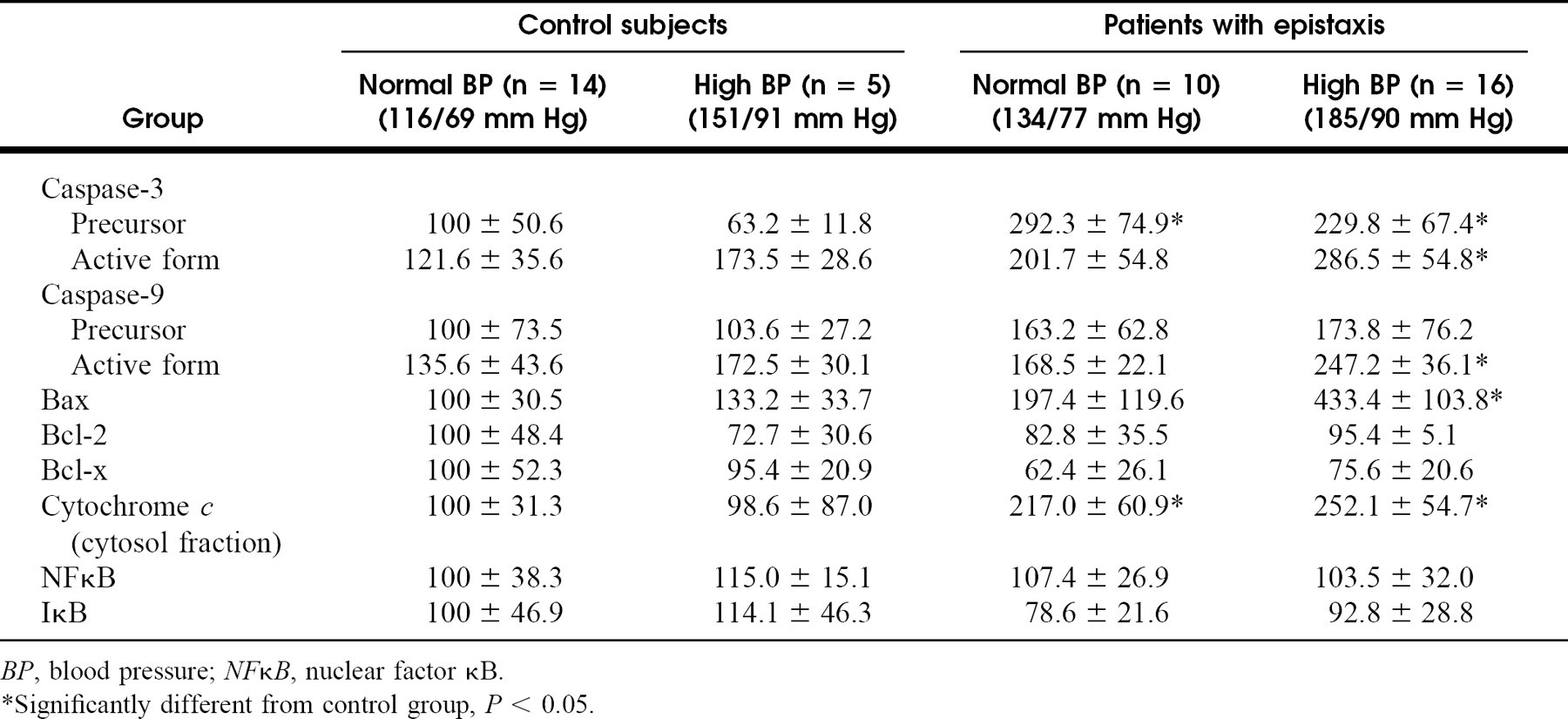
Body weight (Wt, kg), middle ear volume (Vol, ml), and average preoperative and postoperative time constant (K N2O) measured for the left and right ears of all animals in groups 1 and 2
RESULTS
After surgery, the right ear of all animals in both groups developed OME as documented by otomicroscopy and tympanometry. Postoperative experiments were conducted only after the typanograms for the respective MEs were normalized and no effusion was visualized by otomicroscopy. Usually, this occurred approximately 2 weeks earlier for right ears of group 1 animals compared with those of group 2. The average number of postoperative days before the first experiment was 29.8 ± 6.6 for group 1 and 44.8 ± 18.9 days for group 2, and before the second experiment was 40.5 ± 5.5 days for group 1 and 53.3 ± 17.4 days for group 2.
Table 1 lists the body weight for all animals and the ME volumes and N2O time constants (average of 2 preoperative and 2 postoperative measurements) for the left and right ears of animals in the control and experimental groups. Average body weight was 3.41 ± 0.47 kg for experimental animals and 3.15 ± 0.32 kg for control animals. There was a positive, linear relationship between the measured left ME volume and body weight (Ve = 0.56BodyWeight + 0.23, r = 0.64, P < 0.01). Average test-retest measurements of left ME volume were 2.02 ± 0.36 and 2.10 ± 0.40 ml (P = NS), and these measurements were highly correlated (r = 0.88). Average left and right ME volumes were 1.95 ± 0.34 and 1.83 ± 0.36 mL (P = NS) for control animals and 2.16 ± 0.36 and 1.18 ± 0.17 mL (P < 0.01) for experimental animals. The average preoperative and postoperative time constants were 0.0038 ± 0.0015 and 0.0045 ± 0.0010/min (P = NS) for the left ears, 0.0040 ± 0.0013 and 0.0047 ± 0.0015/min (P = NS) for the group 1 right ears, and 0.0034 ± 0.0010 and 0.0031 ± 0.0009/min (P = NS) for the group 2 right ears.
Figure 1 shows the average preoperative and postoperative values of right ME pressure as a function of 5% N2O breathing time for group 2 animals. Also shown is the pressure-time function predicted using the average time constant measured preoperatively for those ears and the functions predicted after adjusting that time constant for the postoperative ME volume reduction under the assumption that the mastoid acts as an ideal gas reserve (Ke N2O = 0.0078/min; Ve = 1.18 mL, Vt = 1 mL, C = 0) or a partial gas reserve (Ke N2O = 0.0046/min; Ve = 1.18 mL, Vt = 1 mL, C = 0.5.). There were no differences between the measured preoperative and postoperative pressure-versus-time functions. The function predicted by the preoperative time constant fit both data sets well. In contrast, the pressure-time functions predicted under the assumption of a gas-reserve function for the mastoid showed much greater rates of pressure change vis-a-vis the postoperative data.
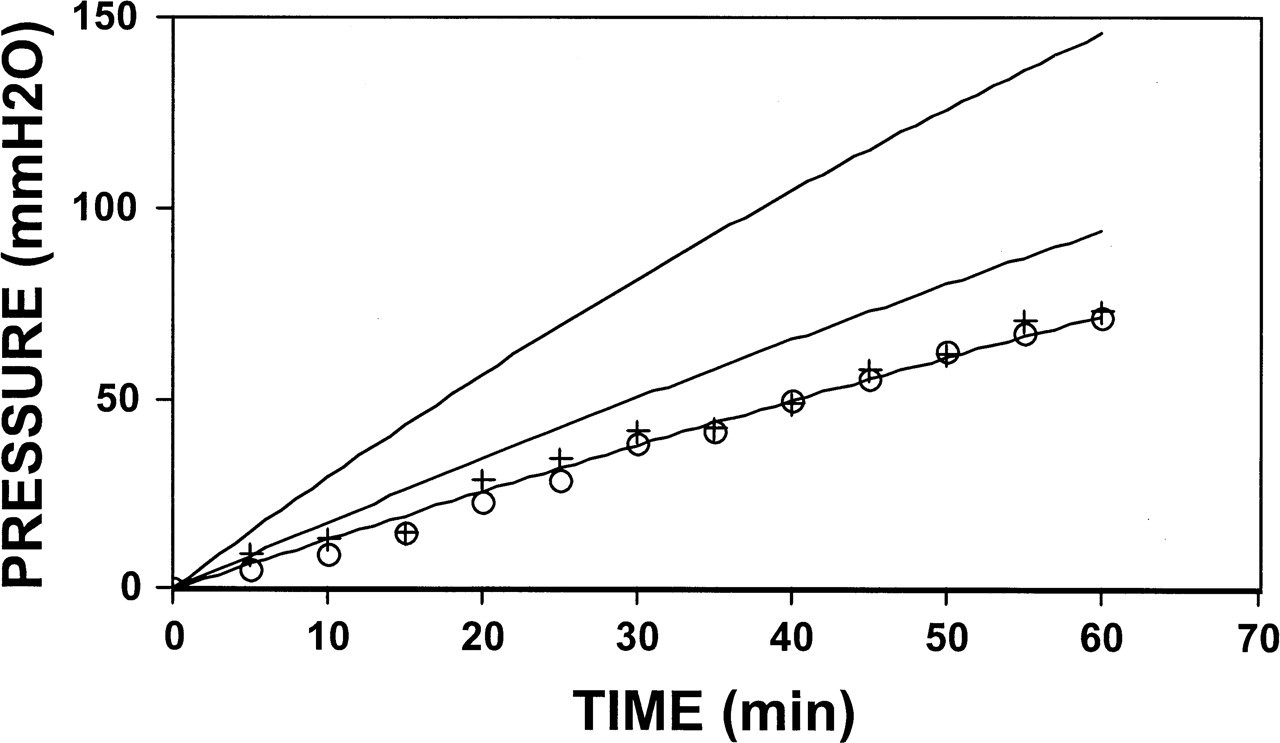
Average middle ear pressure-time functions for the N2O breathing experiments in right ears of group 1 animals performed before (circles) and after (plus signs) mastoid antrum blockage. Also shown is the predicted function assuming the time constant for those ears measured preoperatively (line fitted to data) and the functions predicted for the postoperative data after adjusting the time constant to conditions where the mastoid acts as an ideal (top line) or partial (middle line) gas reserve. See text for details.
Figure 2 shows the distribution of postoperative time constants for both ears of the 12 monkeys as a function of ME volume. Also shown are the predicted time constant-versus-volume functions for MEs where the mastoid behaves as an ideal (Ke N2O = 4.2 at Ve = 2.8 mL, C = 0) and partial gas reserve (C = 0.5) and where the mastoid has no gas-reserve function (C = 1). There is no apparent relationship between the measured time constants and ME volume for all ears or for ears within any subgroup (ie, all left ears; right ears of group 1 or group 2), a pattern not consistent with a gas-reserve function for the mastoid. In contrast, the predicted time constants under conditions where the mastoid acts as a gas reserve show a curvilinear decrease with increasing ME volume.
DISCUSSION
The mechanism underlying a possible effect of mastoid volume on ME pressure has had confusing presentations in the literature, with most descriptions implying a limiting role with respect to the total magnitude of pressure change 4,5,12–15 as opposed to a buffering effect on the rate of pressure change as described in this report. 6 An example of the former is provided by Sade and Fuchs: “The physiological role of the pneumatized mastoid can be seen to be a function of its volume, which according to Boyle's law, may share and damp middle ear pressure aberrations.” 5-9 Their description implies that larger mastoid volumes will maintain stable ME pressures at lesser deviations from ambient compared with smaller mastoid volumes. However, that expectation is not consistent with known physiologic mechanisms because, in the absence of gas resupply from an external source, ME pressure will be stable only when the blood-ME pressure gradients for all represented gases are equal to 0 mm Hg. Because the sum of the blood gas partial pressures is less than that of the ambient environment by approximately 50 mm Hg, 7 ME pressure will stabilize at that underpressure regardless of mastoid volume. 6
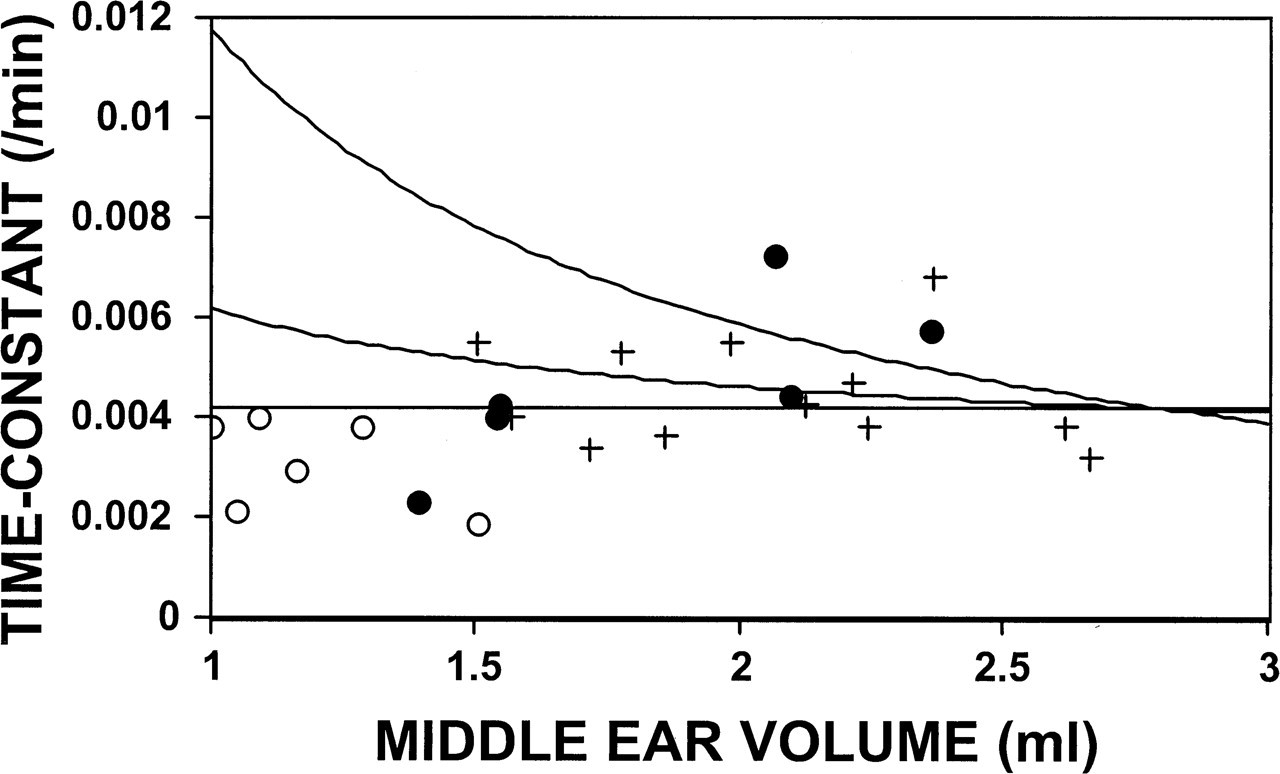
The postoperative time constants as a function of volume for the left (plus signs), group 1 right (filled circles), and group 2 right (open circles) middle ears. Also shown are the predicted functions assuming that the mastoid acts as an ideal (top curve) or partial (middle curve) gas reserve or has no gas-reserve function (bottom curve). See text for details.
Quasi-stable ME pressures at near-ambient levels can be maintained by repeated introductions of gas volumes to the ME as occurs during ET openings. Therefore, the frequency of tubal openings and the volume gas transferred per tubal opening must be sufficient to balance the net gas loss due to transmucosal exchange. More rapid gas exchange rates will require more frequent tubal openings and/or greater volume transfers per opening to maintain near equivalence between ME and ambient pressures. Thus, the significance of a mastoid gas-reserve function to ME pressure regulation lies in its potential to decrease the demand placed on the ET tube for gas resupply. 6
Because under physiologic conditions the total ME-blood pressure gradient is attributable to the difference in N2 pressure, an effective gas-reserve function for the mastoid would be manifest as a lesser rate of transmucosal N2 exchange for larger mastoid volumes. 6 However, past studies showed that the transmucosal N2 exchange rate is very slow and not measurable in short duration experiments. 11 Therefore, in this experiment and in earlier studies, the transmucosal exchange rate of N2O, a nonphysiologic, inert gas characterized by a more rapid, perfusion-limited transmucosal exchange, was studied as a surrogate for N2 ex-change. 15–17,19
In this experiment, communication between the tympanum and mastoid was interrupted by surgical blockage of the mastoid antrum in the right ears of the 6 group 2 animals. The effectiveness of the procedure was demonstrated by the significantly lesser ME volume measured for the right ME vis-à-vis the left ME of group 2 animals. In contrast, the volumes of the right MEs for group 1 (sham surgery) were not different from the respective left MEs, and left ME volume for both groups was reproducible on repeated measurement. Time constants for transmucosal N2O exchange were measured for the left and right ears of all animals before and after the surgery using previously established techniques. 19 The average preoperative and postoperative time constants were not different for the left ears or for the right ears of either group. Also, for all ears (all left ears and the right ears of group 1 and group 2), there was no apparent relationship between the time constant and ME volume. These results are not consistent with the hypothesized gas-reserve function of the mastoid, which predicts a lesser time constant for increasing ME volume and an increase in the time constant after blockage of the mastoid antrum. Although the sample size for this experiment was small, establishing a 95% confidence interval about the postoperative values of the group 2 time constants excludes all predicted values for C < 0.6.
An ideal mastoid gas reserve would exchange gas only with the tympanum in the air phase. In the mathematical model presented earlier, this corresponds to the condition where the constant, C = 0. A less-efficient, but still functional, gas reserve is realized for all nonzero values of C < 1. C represents the relative volume of inert gas exchanged with blood per unit volume of mastoid and tympanum airspace. In physical terms, C is defined as the product of the mastoid-tympanum ratios of the respective surface area-to-volume ratio, blood flow per unit surface area, and the ratio of the arterial-venous to arterial-ME gas pressure gradient. The latter 2 terms represent the effective mucosal perfusion per unit surface area. From geometric considerations, the surface area-to-volume ratio is much greater for the partitioned mastoid airspace (ie, numerous interconnecting air cells) compared with that of the tympanum, and thus the effective mucosal perfusion per unit surface area must be much less for the mastoid than for the tympanum. 20 Moreover, these relationships must be maintained during the period of growth and development of the mastoid air cell system. These restrictive requirements make the hypothesized mastoid gas-reserve function unlikely in the absence of strong supporting data.
Earlier studies that attempted to test the mastoid gas-reserve hypothesis in humans by studying the rate of ME pressure change in response to breathing gas mixtures containing N2O were confounded by the presence of concurrent ME disease, 12 inclusion of both adults and children in the study population, 12,14 or the failure to document complete resolution of the residual effects of preexisting mucosal inflammation. 15 These factors can either decouple the ME pressure response from the volume rate of N2O exchange (eg, overt disease) 17 or differentially and independently affect the rate of N2O exchange (eg, age, subclinical disease). 16,17 The latter class of confounders is particularly troublesome for data interpretation because those factors can covary with mastoid volume while causing changes in the ME pressure response similar to those expected under the hypothesis of a gas-reserve function for the mastoid.
For example, a more rapid N2O blood saturation was reported for infants and children compared with adults. 18 This will establish a higher N2O blood-ME gradient earlier in children and during the time period over which the rate of ME pressure change is usually measured (typically the first 10 minutes after onset of breathing the anesthetic gas mixture). From Eq 8 [δPe N2O/δt = (Pa N2O -Pe N2O)Ke N2O], a larger blood-ME N2O gradient will cause a faster increase in ME pressure for equal N2O time constants. Because children have lesser mastoid volumes compared with adults, the interpretation of the results for studies that included wide age ranges 12,14 may erroneously attribute a faster observed rate of ME pressure increase to a smaller mastoid volume, when in fact the difference is a consequence of higher N2O driving pressures. Also, as defined by Eq 9 [Ke N2O = RTeSb N2OQtFt N2O(1 + XC)/Ve], the measured N2O time constant depends on the mucosal blood perfusion rate. Because small mastoid volumes in both children and adults have been related to OME susceptibility 15 and the accompanying mucosal inflammation can persist long after otoscopic and/or tympanometric documentation of disease resolution, a faster rate of ME pressure change during N2O breathing for smaller mastoids cannot be attributed with certainty to a gas-reserve function of the mastoid (versus increased mucosal perfusion secondary to residual inflammation). For these reasons, interpretations of the results of past clinical studies purportedly demonstrating a mastoid gas-reserve function should be reconsidered in light of these caveats.