
Abstract
Involvement of the intrinsic laryngeal muscles in generalized neuromuscular disorders is infrequent. 5 Myotonic dystrophy, or Steinert's disease, is an autosomal dominant multisystem degenerative disease characterized by myotonia and progressive muscular weakness. 6 It represents one of the most common inherited muscle disorders with an estimated prevalence of 2.1 to 14.3 per 100,000 worldwide, yet laryngeal involvement is rare. 7 There are few references in the literature regarding laryngeal involvement in other neuromuscular disorders such as arthrogryposis, scapuloperoneal muscular atrophy, oculopharyngeal muscular dystrophy, and Kearns-Sayre syndrome. 5 It is difficult to study the morphology of the laryngeal muscles with such rare involvement in generalized neuromuscular diseases. One patient with Kearns-Sayre syndrome did not have clinical laryngeal involvement. 5 On examination of her intrinsic laryngeal muscles after death, there was noted to be characteristic red ragged fibers seen in Kearns-Sayre syndrome. All intrinsic laryngeal muscles were involved but less so than the extraocular muscles. 5 In biopsy specimens of laryngeal muscles from patients suffering from myotonic dystrophy, Duchenne's muscular dystrophy, and oculopharyngeal muscular dystrophy, there were minimal variations from the general morphological pattern of the specific muscular dystrophic process. 8 However, although the laryngeal muscles underwent the same alterations, they preserved the cross striations in almost all cases. This recalcitrance to injury was interpreted as an explanation for the prolonged function of the laryngeal muscles even in severe cases of these degenerative diseases. 8
In normal adult limb skeletal muscle, the myonuclei are postmitotic. Satellite cells reside under the muscle basal lamina and are quiescent unless the muscle is injured. We recently showed that normal, uninjured adult laryngeal muscles continue to add myonuclei, adding approximately 2 myonuclei per every 1,000 myofibers in a 24-hour period. 9 Given the difference in the uninjured baseline myonuclear status, 9 the response of laryngeal myonuclei to denervation, in terms of time course and location of response, seemed warranted. In this study, we examined the effect of unilateral recurrent laryngeal nerve section on the process of myonuclear addition using bromodeoxyuridine uptake as a marker of cell division.
MATERIALS AND METHODS
Adult New Zealand white rabbits were obtained from Bakkon Rabbitry (Viroqua, WI) and housed in the University of Minnesota animal facility. All procedures were approved by the Animal Care and Use Committee at the University of Minnesota and adhered to the National Institutes of Health guidelines for use of animals in research.
Nerve section was performed on 16 normal adult rabbits. The rabbits were anesthetized with an intramuscular injection of 1:1 ketamine and xylazine (10 mg/kg:2 mg/kg, respectively). The left recurrent laryngeal nerve in each animal was dissected free of other neck structures, and a 2-cm section was removed. The incision site was closed using nylon sutures. One week or 24 weeks after surgery, 8 rabbits received intraperitoneal injections of bromodeoxyuridine (brdU) in sterile saline at a dose of 50 mg/kg body weight every 2 hours for 12 hours, followed by a 24-hour brdU-free period. The muscles from 4 additional normal rabbits were labeled with brdU in the same manner as controls. Twenty-four hours after the first brdU injection, the rabbits were given an overdose of barbiturate anesthesia. The entire larynx was dissected free of surrounding tissue, and the left epiglottis was removed with a scalpel to verify the injured side. The larynges were embedded in tragacanth gum and frozen in methylbutane chilled to a slurry on liquid nitrogen. The larynges were serially sectioned at 12 μm. Every tenth section was immunostained for expression of brdU (Chemicon, Temecula, CA; 1:100) and dystrophin (Vector Laboratories, Burlingame, CA; 1:20) to ascertain if the labeled nuclei were within the muscle sarcolemma and therefore were myonuclei. Every 30th section was stained with hematoxylin and eosin. Every 20th section was immunostained for neonatal MyHC. For double-labeling experiments, tissue sections were quenched with hydrogen peroxide, incubated with blocking serum, followed by biotinavidin blocking reagent (Vector Laboratories), and incubated using a primary antibody to dystrophin. The sections were rinsed in phosphate-buffered saline (PBS) and incubated with the Vectastain peroxidase ABC kit (Vector Laboratories). The peroxidase was developed using 3,3′-diaminobenzidine and hydrogen peroxide. After a PBS rinse, the sections were incubated in 2 N HCl for 1 hour at 37°C, followed by neutralization in borate buffer and a PBS rinse. The sections were incubated using a primary antibody to brdU, rinsed in PBS, incubated with the alkaline phosphatase ABC kit (Vector Laboratories), and reacted with the alkaline phosphatase black substrate kit. All brdU myonuclear counts were performed on brdU-dystrophin–labeled sections. The dystrophin immunostained brown, and the brdU-positive nuclei were black. BrdU labeling and myofiber number were quantified using the Bioquant Nova Prime morphometry system (Bioquant Inc, Nashville, TN).
Immunolabeling for neonatal MyHC were performed as described earlier, except the primary antibody was used at 1:20 (Vector Laboratories). The number of brdU-labeled nuclei was determined as a percent of the number of myofibers in the microscopic fields counted. The data was analyzed for statistical significance using either a paired 2-tailed t test or an analysis of variance and Dunn's multiple comparison tests aided by the Prism and Statmate software (Graphpad, San Diego, CA). An F test was used to verify that the variances were not significantly different. Data were considered significantly different if P < 0.05.
RESULTS
In all muscles examined, a few brdU-positive cells were present within the connective tissue between myofibers. These cells could represent fibroblasts, lymphocytes, or satellite cells. Their identity was not investigated further. In the pectoralis major and tibialis anterior, an average of 1,000 muscle fibers were counted per section, and no brdU-positive myonuclei were found within the dystrophin-positive myofiber cross-sections. 9
In the thyroarytenoid muscle (TA), 6.11% ± 0.44% of the myofibers in cross-section had a brdU-positive nucleus at 1 week after denervation (Fig 1). On the contralateral side, 3.49% ± 0.91% of the myofibers had a brdU-positive nucleus. These were both statistically significant increases when compared with the control value of 0.14% ± 0.04%. The denervated side was also significantly increased compared with the contralateral side (Fig 2). At 24 weeks, the denervated TA was similar to the contralateral side, 0.41% ± 0.16% and 0.50% ± 0.09%, respectively (Fig 2). By 24 weeks, these were not statistically different from control values.
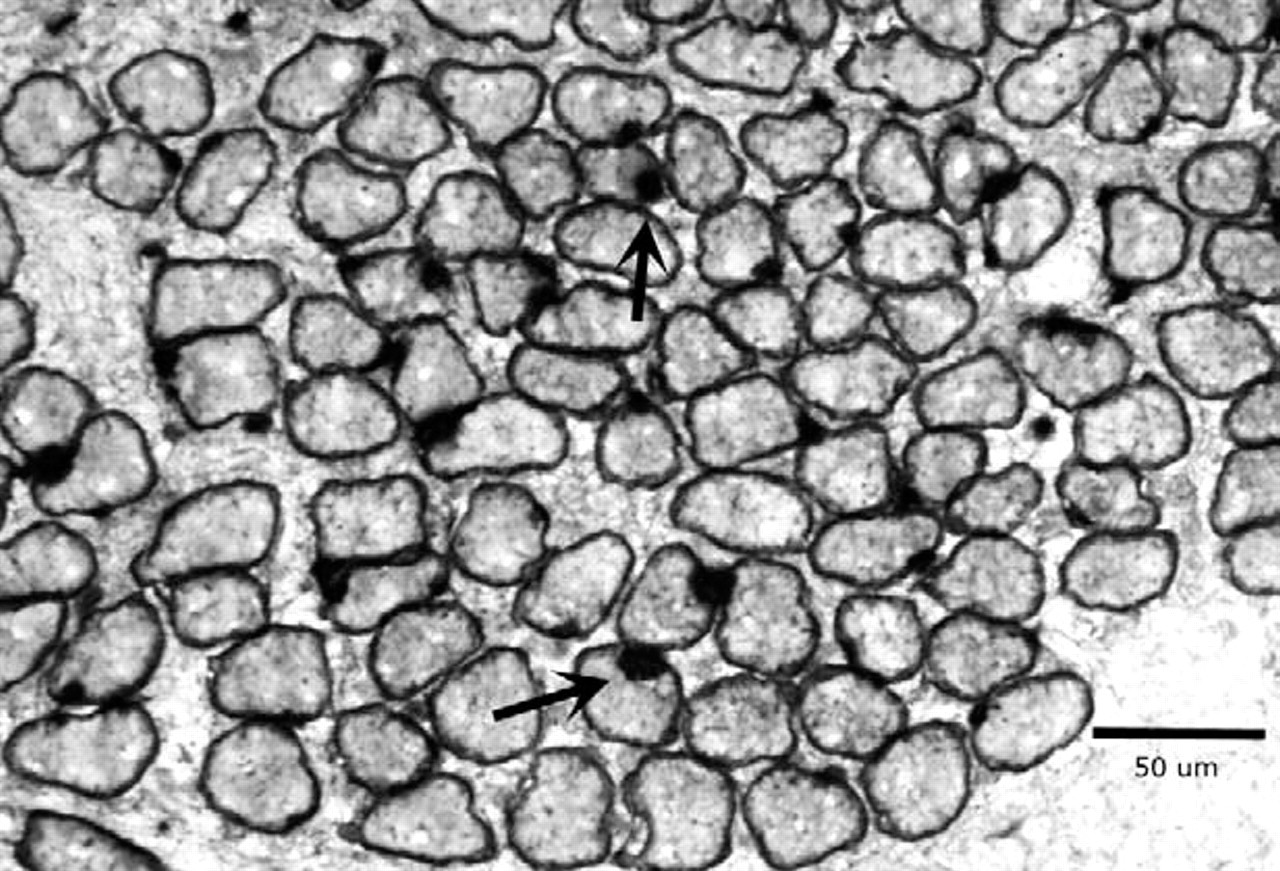
Cross-section of thyroarytenoid muscle from adult rabbit 1 week after denervation demonstrating integration of brdU-labeled myonuclei into myofibers. Black arrows indicate brdU-labeled myonuclei within dystrophin-labeled sarcolemma. Magnification bar is 50 μm.
In the posterior cricoarytenoid muscle (PCA), 2.59% ± 0.61% of the myofibers in cross-section had a brdU-positive nucleus at 1 week after denervation. This was significantly elevated over both the control and contralateral sides. On the contralateral side, 1.18% ± 0.30% of the myofibers had a brdU-positive nucleus, a significant increase compared with the control value of 0.13% ± 0.01% (Fig 2). At 24 weeks, the denervated numbers were similar to the contralateral side, 0.34% ± 0.12% and 0.46% ± 0.22%, respectively (Fig 2), which were not statistically significant compared with controls.
In the lateral cricoarytenoid muscle (LCA), 2.72% ± 0.82% of the myofibers in cross-section had a brdU-positive nucleus at 1 week after denervation. This was significantly elevated over the control and contralateral sides. On the contralateral side, 1.09% ± 0.25% of the myofibers had a brdU-positive nucleus. The control value was 0.48% ± 0.08% (Fig 2). At 24 weeks, the denervated side was similar to the contralateral side, 0.37% ± 0.13% and 0.41% ± 0.14%, respectively (Fig 2), and not statistically different from control values.
At 1 week, neonatal MyHC expression was significantly elevated compared with the contralateral control side and to muscles from normal rabbits (Fig 3). In fact, almost every myofiber seen in cross-section 1 week after recurrent laryngeal nerve section was neonatal MyHC positive. The number of myofibers positive for neonatal MyHC decreased by 24 weeks but appeared to be elevated compared with controls (Fig 4). The majority of myofibers appeared to be normal in cross-sectional area compared with control, but small groups of either atrophic or regenerating myofibers positive for neonatal MyHC were seen (Fig 4).
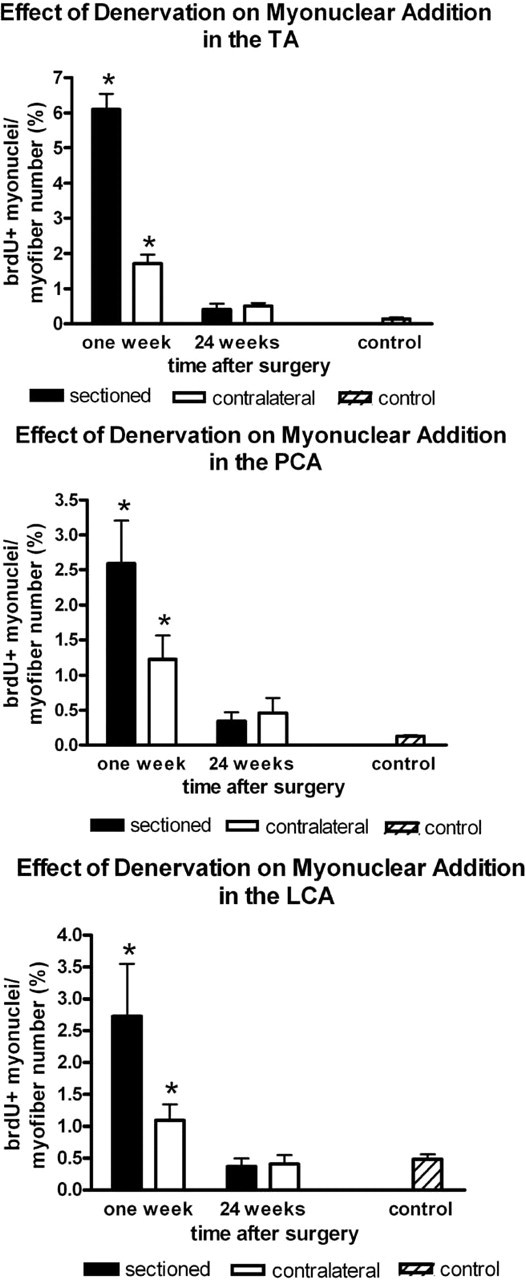
The percentage of myofibers, in cross-section, in the TA, PCA, and LCA muscles with a brdU-positive myonucleus indicating myonuclear addition at 1 week and 24 weeks after denervation in the denervated and contralateral sides compared with control. Asterisk signifies statistical significance of P < 0.05.
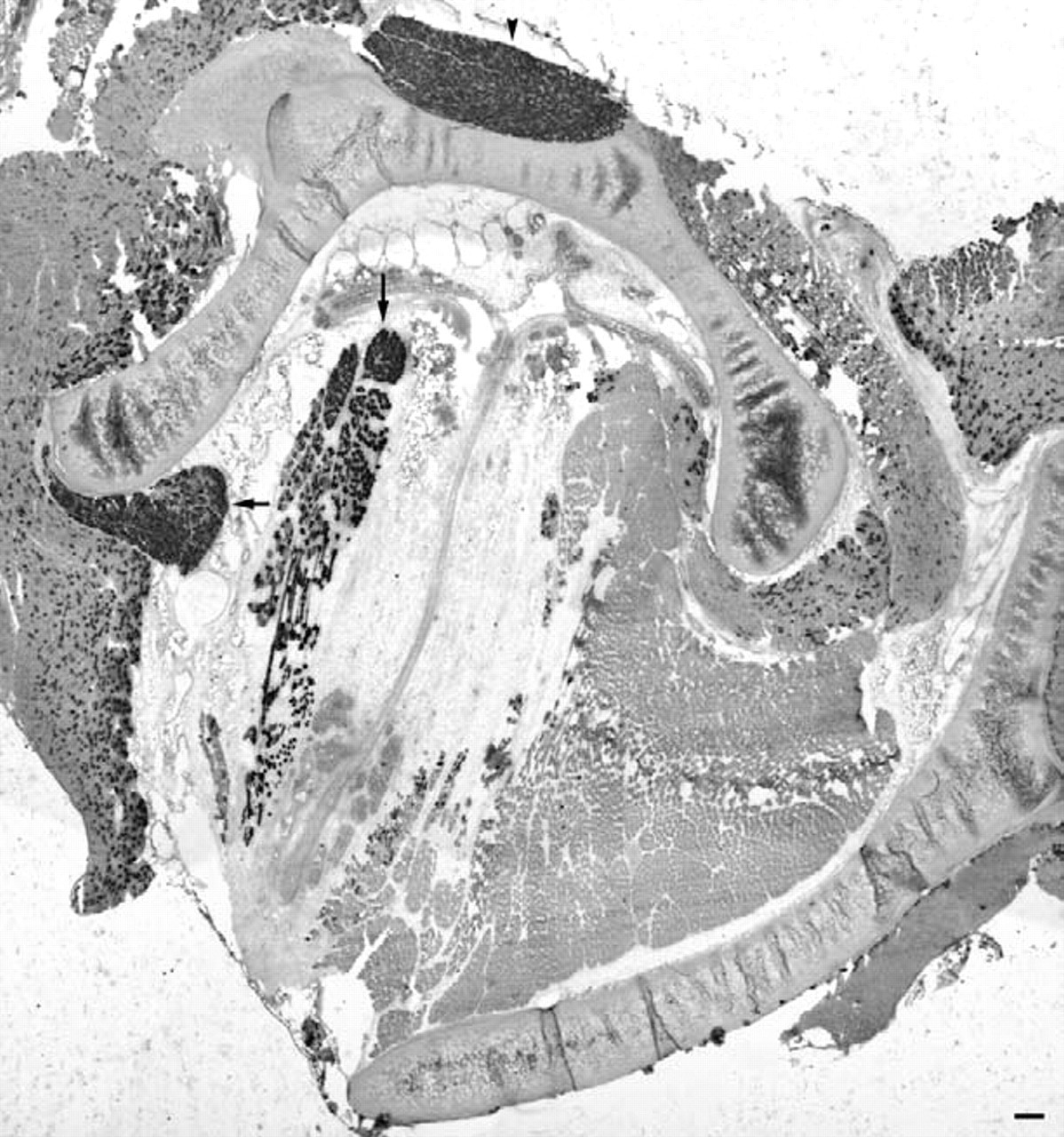
Cross-section of adult rabbit larynx 1 week after denervation showing increased expression of neonatal MyHC compared with contralateral side. Vertical arrow, TA; arrowhead, PCA; horizontal arrow, LCA. Magnification bar is 200 μm.
DISCUSSION
There was a clinical understanding of the uniqueness of the laryngeal muscles before understanding their unusual properties at a cellular level. The intrinsic laryngeal muscles have an adenosine triphosphatase activity profile similar to extraocular muscles (EOM). 10 In addition, contraction times of the intrinsic laryngeal muscles of adult rhesus monkeys were similar to those described for EOM and are exceedingly fast compared with limb skeletal muscle. 11 The intrinsic laryngeal muscles share many of the unique properties of the EOM including expression in adults of specific immature and unusual MyHC isoforms not seen in limb muscles, including neonatal and developmental MyHC. 12,13 These properties could be because of unique embryology, cranial nerve innervation, or to continuous myofiber turnover in adults.
The EOM have a remarkable ability to add myonuclei and remodel in normal adult mammals, even in elderly individuals. 14–16 This is accomplished in EOM by maintaining a population of activated satellite cells in the absence of injury. Recently, the same phenomenon of a slow but continuous addition of myonuclei to existing myofibers in normal rabbit adult laryngeal muscle was shown. 9
Our study is the first to examine the response of satellite cells to laryngeal denervation. In contrast to decreased messenger RNA (mRNA) expression of cell cycle transcription factors,
17
there was a marked increase in myonuclear addition in the intrinsic laryngeal muscles through satellite cell activation.
9
This included a statistically significant difference in the denervated TA, PCA, and LCA compared with the contralateral side and control muscles. Although not as large as the denervated side, there was a significant difference on the contralateral side when compared with control animals at 1 week. The increased brdU-positive myonuclei were expected on the denervated side based on prior studies with denervated muscles.
18,19
The contralateral uninjured TA, PCA, and LCA myonuclear addition increases may have several explanations. Although unlikely, there may have been injury to the contralateral recurrent laryngeal nerve (RLN). In 1 case in which this inadvertently happened because of surgical exploration on both sides of the trachea, the animal awoke from anesthesia stridorous, necessitating euthanasia. It is also possible that sprouting occurs from the contralateral side; however, this type of sprouting did not occur in an examination of experimental denervation.
20
It is also possible that the contralateral laryngeal muscles work harder secondary to the paralysis on the denervated side, as seen in positron-emission tomography scanning,
21
where after unilateral paralysis the vocal fold of the uninjured side crosses the midline in an attempt to restore normal approximation of the vocal folds to normalize phonation and prevent aspiration. Satellite cells fuse with existing myofibers in response to injury conditions such as exercise and overloading.
22
This would correlate with the increased brdU-positive myonuclei of the contralateral TA compared with the contralateral PCA and LCA because this response would affect the TA to a greater degree. There was a trend toward the maintenance of increased myonuclear addition at 24 weeks after denervation, but this was not significantly different from control values. However, very little evidence of muscle atrophy was seen. This is in contrast to long-term denervation of limb muscle in which significant atrophy occurs by 24 weeks postdenervation.
23
It is also possible
Cross-section of the PCA muscle 24 weeks after denervation immunostained for the expression of neonatal MyHC. Arrow indicates a small group of myofibers of smaller cross-sectional area. Magnification bar is 50 μm.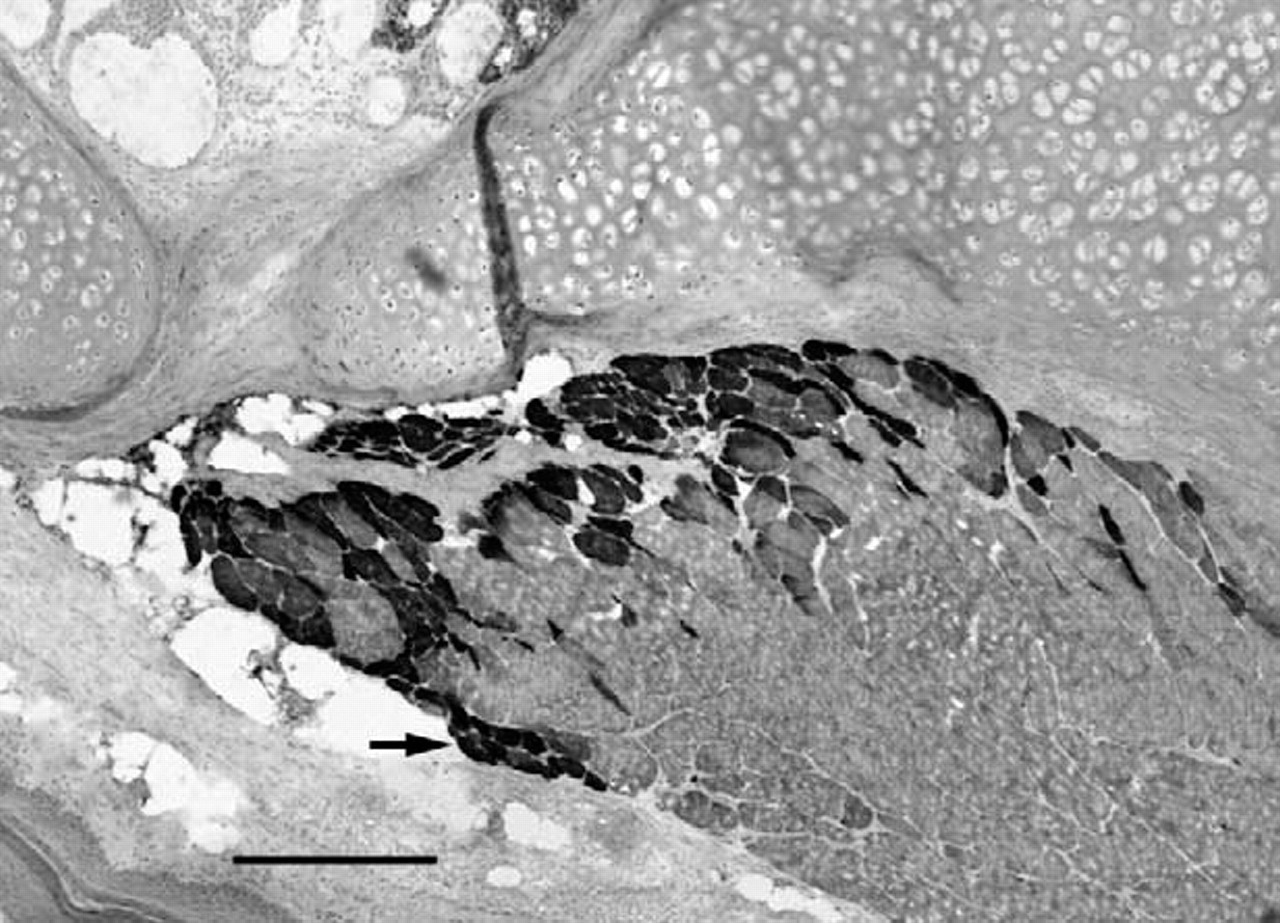
Numerous studies have investigated the specific MyHC isoforms in laryngeal muscles and their changes in expression after denervation. After recurrent laryngeal nerve section in the rat, type I MyHC was almost completely gone at 6 months with a concomitant increase in type II MyHC in the PCA. 24 The type IIL laryngeal MyHC, which is identical to extraocular MyHC, remained constant in the PCA and TA after denervation. Nonneuronal factors were postulated to explain the type IIL expression. 24 After denervation of rat laryngeal muscles, a decrease in type IIB and an increase in type IIA and IIX expression in the PCA and TA was observed; however, variable changes in type IIL occurred over the 6-month timecourse. 25 After recurrent laryngeal nerve section in the rat with immediate reanastomosis, there was a transformation in the TA from fibers coexpressing IIB/EO(IIL) to IIX. This was attributed to random cross-reinnervation showing that myosin expression in laryngeal muscle fibers is subject to neural regulation. 26
Laryngeal denervation affected cell-cycle regulation, resulting in an increase in the mRNA expression of MyoD and the cell-cycle inhibitors p21 and GADD45. This was interpreted as an inhibition of satellite cell proliferation with laryngeal muscle denervation in the first month after denervation. 17 However, it is noted that to date no studies have examined the response of satellite cells to laryngeal denervation. 17
Work is underway to quantify the results of the neonatal MyHC expression after denervation. Neonatal MyHC is expressed in muscle development but may also be expressed during denervation, regeneration, or atrophy. 27 An immunohistochemical classification of normal human interarytenoid muscles noted neonatal MyHC expression but only in a very few fibers. 27 In the PCA as early as 1 week after denervation, there was an increase in mRNA for neonatal MyHC but the increase was modest. 17 It should be noted that mRNA expression does not always correlate well with changes in protein expression. The increase in neonatal MyHC expression at 1 week after denervation in the TA, PCA, and LCA in our study encompasses expression in nearly every fiber in contrast to the modest increase seen at the mRNA level. 17 At 24 weeks, the small fibers in the TA expressing neonatal MyHC may represent regenerating or atrophying fibers.
The process of satellite cell activation, as supported by MyoD upregulation, 18 and myonuclear addition in denervated laryngeal muscles and the resulting maintenance of muscle size may, in part, explain the ability to successfully reinnervate laryngeal muscles in patients after long periods of denervation. 27 This phenomenon was seen after functional denervation caused by botulinum toxin treatment of extraocular muscles, with significant satellite cell activation and myonuclear addition. 28 Work in models of chronic muscle denervation suggests that both the denervated muscle and denervated nerve play a role in failure of adequate regeneration, even when a fresh nerve is used. 29 In addition, it is possible that the sparing of laryngeal muscle function in a wide array of muscle diseases may, in part, be explained by the ability of laryngeal muscles to remodel by continuous addition of myonuclei.
In summary, denervation of adult laryngeal muscles by section of the recurrent laryngeal nerve results in significant upregulation of myonuclear addition in the denervated muscles and a compensatory upregulation of myonuclear addition in the laryngeal muscles on the contralateral side compared with control muscles. Because denervation results in significantly increased myofiber remodeling compared with control laryngeal muscles, the upregulation of these processes may contribute to the minimal histological and morphometric changes that occur after nerve section, as well as the ability to be reinnervated successfully after long periods of denervation.