
Abstract
Inner ear fluid homeostasis is critical to normal auditory and vestibular function. Disturbance of this delicate fluid balance can result in overaccumulation of endolymph in the inner ear, or endolymphatic hyrdops (EH). The resultant changes in the membranous labyrinth seen histologically with EH are believed to account for the classic tetrad of symptoms associated with Ménière's disease (MD): fluctuating hearing loss, tinnitus, vertigo, and aural fullness.
The pathogenesis of EH is unclear. The underlying problem is presumed to be either one of overproduction or malabsorption of endolymph. Dysregulation of fluid production or fluid transport into the endolymphatic space can lead to EH. Systemic hormones, such as vasopressin (VP), may play a role. This has been demonstrated in the animal model. 1 - 6 This relationship between VP and MD also has been demonstrated clinically. Plasma concentrations of VP were significantly elevated in patients with MD. 4 , 7 , 8
There are many described methods to induce experimental EH in animals. Histological EH has been induced in guinea pigs by chronic administration of VP through subcutaneous mini-osmotic pumps. 6 Alternatively, malabsorption of endolymph by the endolymphatic sac can result in EH. Surgical obstruction of the endolymphatic duct and sac is a well-established model of EH in guinea pigs. 9 - 12
Currently, the clinical diagnosis of MD is one of exclusion based on the patient's history, progression of symptoms, physical examination, audiogram, and electronystagmography. An MRI is often obtained to rule out retrocochlear pathology such as a vestibular schwannoma. The MRI is generally not used to evaluate the presence of EH or inflammation. No definitive diagnostic test exists for MD.
Surgically induced EH has been evaluated by MRI in the animal model. At 1.5 tesla (T), the perilymph preferentially enhanced with gadodiamide, facilitating the distinction of compartments within the membranous labyrinth. 13 - 15 With the advancement of MRI scanners and increasing magnetic field strength, greater ultrastructural detail has been seen radiographically. Use of intratympanic gadolinium and optimization of the inversion time have recently demonstrated improved cochlear imaging and differentiation of endolymph, perilymph, bone, and air. 16 Surgically induced EH in guinea pigs has been quantified and evaluated in vivo by MRI at 4.7 T, and again administration of gadolinium contrast assisted in the distinction of cochlear compartments and the presence of membrane rupture. 17 Recently, human subjects with MD underwent MRI at 3.0 T, demonstrating in vivo MD-associated changes of the membranous labyrinth manifested as loss of definition of the cochlear spaces. 18 , 19
The MRI appearance of VP-induced EH has not been previously reported. Our experimental goal in this small pilot study was to characterize the cochlear changes seen on 3.0-T MRI associated with either the administration of high-dose systemic VP or surgical ablation of the endolymphatic sac.
Methods
Subjects
Twelve healthy male albino Hartley guinea pigs weighing between 300 and 500 g were utilized. A small sample size was used because this was a pilot study. All animals were handled according to the standards and protocols of the Institutional Animal Care and Use Committee at the University of North Carolina at Chapel Hill, and the Guide for the Care and Use of Laboratory Animals published by the National Institutes of Health.
Experimental Design
In group 1, five guinea pigs received chronic administration of VP ([Arg]8-VP, Sigma Chemical, St Louis, MO) at a rate of 1000 μU/kg/min (2.86 ng/kg/min) by subcutaneous mini-osmotic pumps (model 2002; Alzet Corp., Palo Alto, CA). The rate of VP administration was selected on the basis of prior studies yielding maximal induction of EH. 6 The pumps were implanted surgically in the subcutaneous tissues on the back between the scapulae. Gadolinium contrast was administered intravenously, and postcontrast MR images were obtained at days zero, seven, and 14 after initiation of systemic VP administration.
In group 2, seven guinea pigs underwent surgical ablation of the right endolymphatic duct and sac. Two animals died during surgery. The five surviving guinea pigs underwent MRI at eight and 12 weeks postablation surgery. Contrast-enhanced coronal 3.0-T MR images were obtained four hours after administration of intravenous gadolinium. Two additional guinea pigs were utilized as control animals and did not undergo the ablation or VP pump implantation procedures.
Surgical Procedures
Mini-osmotic pumps were surgically implanted subcutaneously under sterile conditions. A 1-cm skin incision was made between the scapulae, and the pump was inserted into a subcutaneous pocket developed with a hemostat. The skin was reapproximated with running suture.
Surgical ablation of the endolymphatic sac was performed by using aseptic techniques described previously. Body temperature was maintained with an electric heating pad and a rectal temperature probe. Intramuscular xylazine (5 mg/kg) with ketamine (20-40 mg/kg) and 0.25 percent to 2 percent inhaled isoflurane were administered for deep anesthesia. The soft tissues were infiltrated with 1 percent lidocaine with 1:100,000 epinephrine for local hemostasis. Under operative microscopy, a small craniotomy was created over the supraoccipital ridge with a high-speed otological drill. Blunt dissection and medial retraction of the sigmoid sinus were performed to expose the endolymphatic sac. The sac was then divided with a sharp pick, and a burr was used to drill into the vestibular aqueduct. Subsequently, the vestibular aqueduct was packed with bone wax. 9 , 11 , 15 , 20 The wounds were closed with running chromic suture, and topical antibiotic ointment was utilized in the perioperative period.
Magnetic Resonance Imaging
A precision vaporizer was utilized to administer 0.25 percent to 2 percent isoflurane anesthesia. Imaging was performed with continuous monitoring of respiratory rate and observation of mucous membrane color. All scans were performed with a 3.0-T MRI Siemens Allegra (Erlangen, Germany) head scanner equipped with a 2.1-cm custom surface coil according to the following protocol: gradient echo localizer sequence, transverse T2 images (3.23 min), and coronal spin echo (SE) T1 images prior to gadolinium, and four-hours postgadolinium (echo time [TE] = 20 ms, repetition time [TR] = 600 ms, 256 × 256 matrix, and 0.8-mm slice thickness, 20.33 min). The contrast agent gadolinium was utilized at a dose of 2.5 mL/kg (287 mg/mL) administered through the dorsal penile vein.
Under magnification, manually drawn regions of interest (ROI, 10 pixels average size) were used to evaluate cochlear enhancement. ROIs were placed on a minimum of three slices at three different locations per slice within the perilymphatic and endolymphatic spaces. ROIs were also drawn in the brainstem to serve as a control of signal intensity and to ensure that techniques of acquisition and measurement were uniform throughout the study. Statistical analysis was performed by utilizing a two-tailed Mann-Whitney U test.
Histological Studies
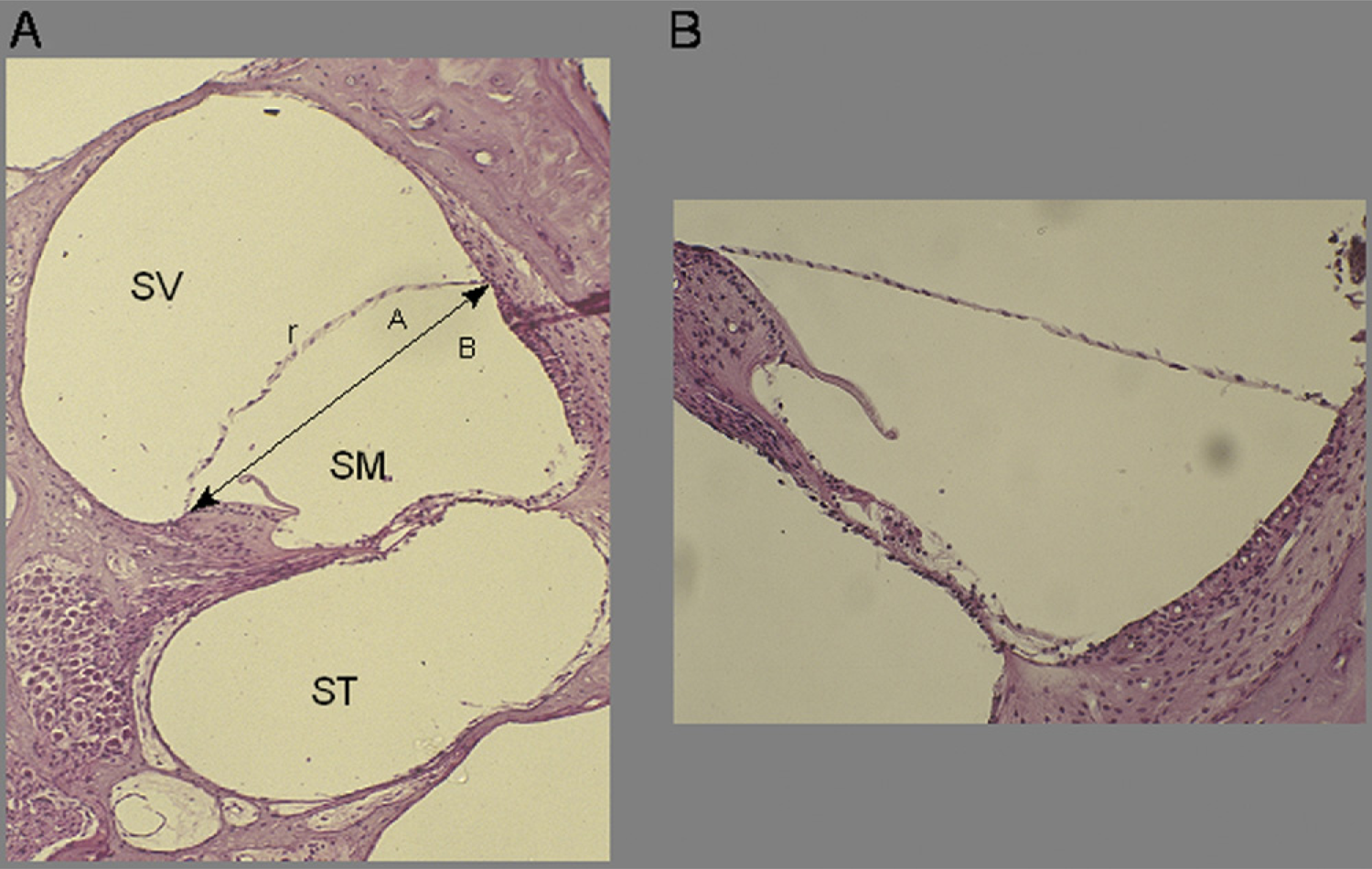
All guinea pigs were euthanized under general anesthesia by a transcardiac saline wash followed by administration of 4 percent paraformaldehyde in physiological buffered saline. The cochleae were immediately harvested, and subsequent temporal bone fixation was performed with 10 percent formalin buffer for 10 days or more. The guinea pig temporal bones were then decalcified with 5 percent trichloroacetic acid and dehydrated in a graded ethanol series. Specimens were then embedded in paraffin, and prepared blocks were cut into three 10-μm midmodiolar sections, stained with hematoxylin-eosin, and viewed with a light microscope for evaluation. The slices were digitized to a computer and analyzed with Scion Image software. Increase ratios (IRs) were calculated to assess cross-sectional area of the scala media by using the formula IR (%) = 100 × (area A/area B). 6
Results
We obtained T1-weighted postcontrast cochlear images through MRI at 3.0 T. All cochleas demonstrated preferential perilymphatic enhancement. There was no significant change in the signal intensity of the brainstem control areas before or after treatment with surgery or chronic VP administration. MRIs at seven days post–chronic administration of VP resulted in increased cochlear enhancement compared with pre–VP administration images (Fig 1). Figure 1A demonstrates the cochlear appearance prior to VP administration, and Figure 1B shows its appearance one week after the initiation of continuous VP administration. Comparison of Figure 1A with 1B demonstrates increased signal intensity at the cochlear level. Magnified views of this increased whole cochlear signal intensity are seen in Figure 1C (pre-VP) and 1D (1 week post-VP). Contrast gadolinium concentration is greater in the perilymph (P) than it is in the endolymph (E). This phenomenon allows for visualization of the cochlear compartments on MRI (Fig 1C). In the surgical ablation group, we saw subjectively increased whole cochlear enhancement on MRI eight weeks after endolymphatic sac obliteration surgery compared with the normal nonsurgical contralateral cochlea of the same animal (Fig 2). In both groups, we also observed an increased signal within the vestibule and semicircular canals.
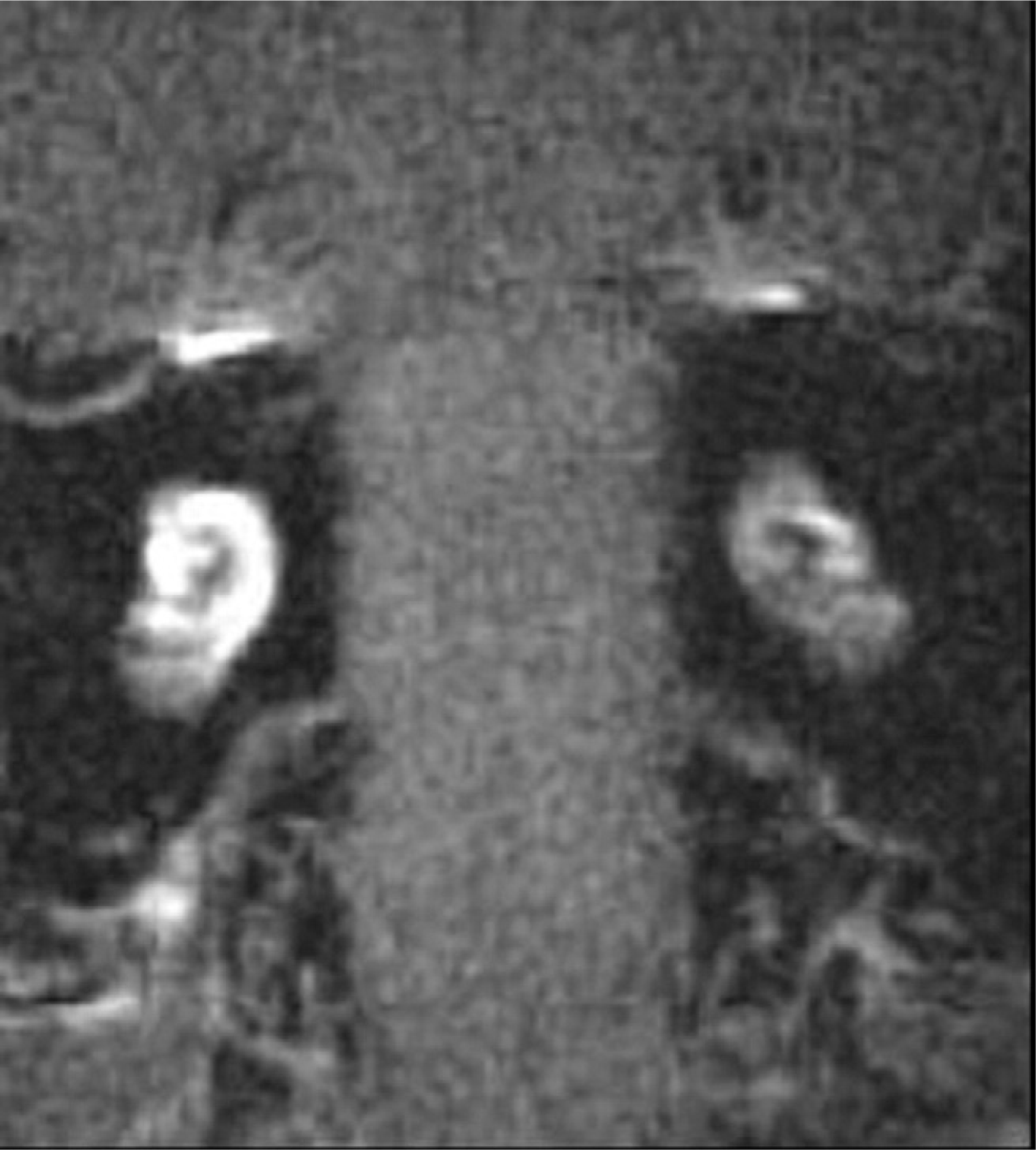
T1-delayed contrast MRI demonstrating increased cochlear enhancement eight weeks after surgical ablation of the right endolymphatic sac compared with the contralateral nonsurgical ear.
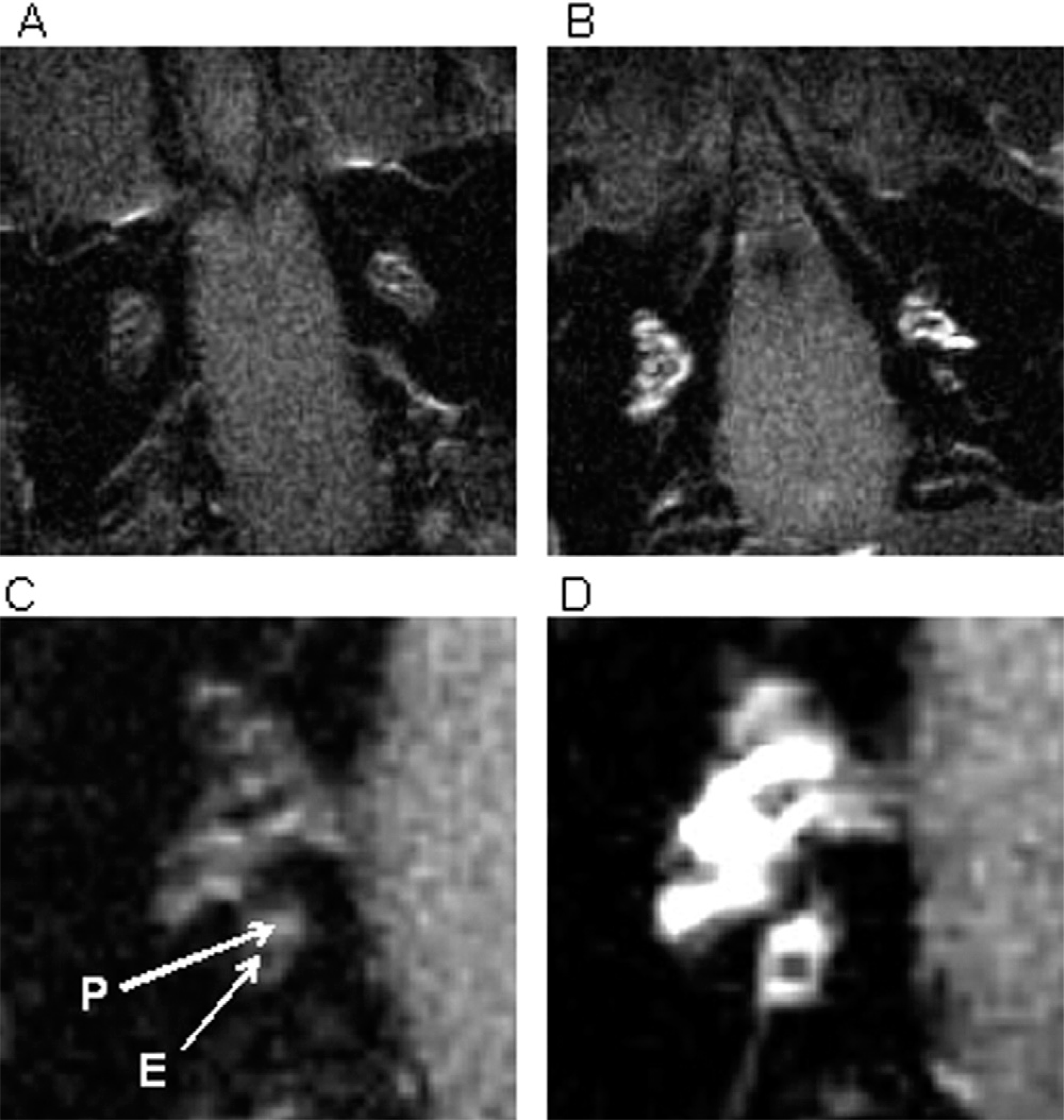
T1-weighted MR images of the cochlea before (
We were unable to consistently detect EH by quantitative size measurement of the cochlear compartments because of equipment limitations and the small size of the cochleae. We did, however, consistently demonstrate increased enhancement of the MRI signal. Specifically, in the endolymphatic sac ablation group, we measured a significant increase in ROI signal intensity of the endolymph following induction of EH of 45 percent (two-tailed Mann-Whitney U test, P = 0.0079). The perilymph enhancement also increased to a lesser degree but was still considered significant (two-tailed Mann-Whitney U test, P = 0.0317) (Fig 3). After chronic administration of VP, the ROI signal intensity of the endolymph increased significantly from baseline at one and two weeks (two-tailed Mann-Whitney U test, P = 0.0159 and 0.0079), with a 110 percent and 215 percent increase, respectively, in the endolymph. The ROI signal intensity of the perilymph also increased from baseline at one and two weeks at 70 and 150 percent, respectively. This result was not significant at one week (two-tailed Mann-Whitney U test, P = 0.0556) but was significant at two weeks (two-tailed Mann-Whitney U test, P = 0.0079) (Fig 4).
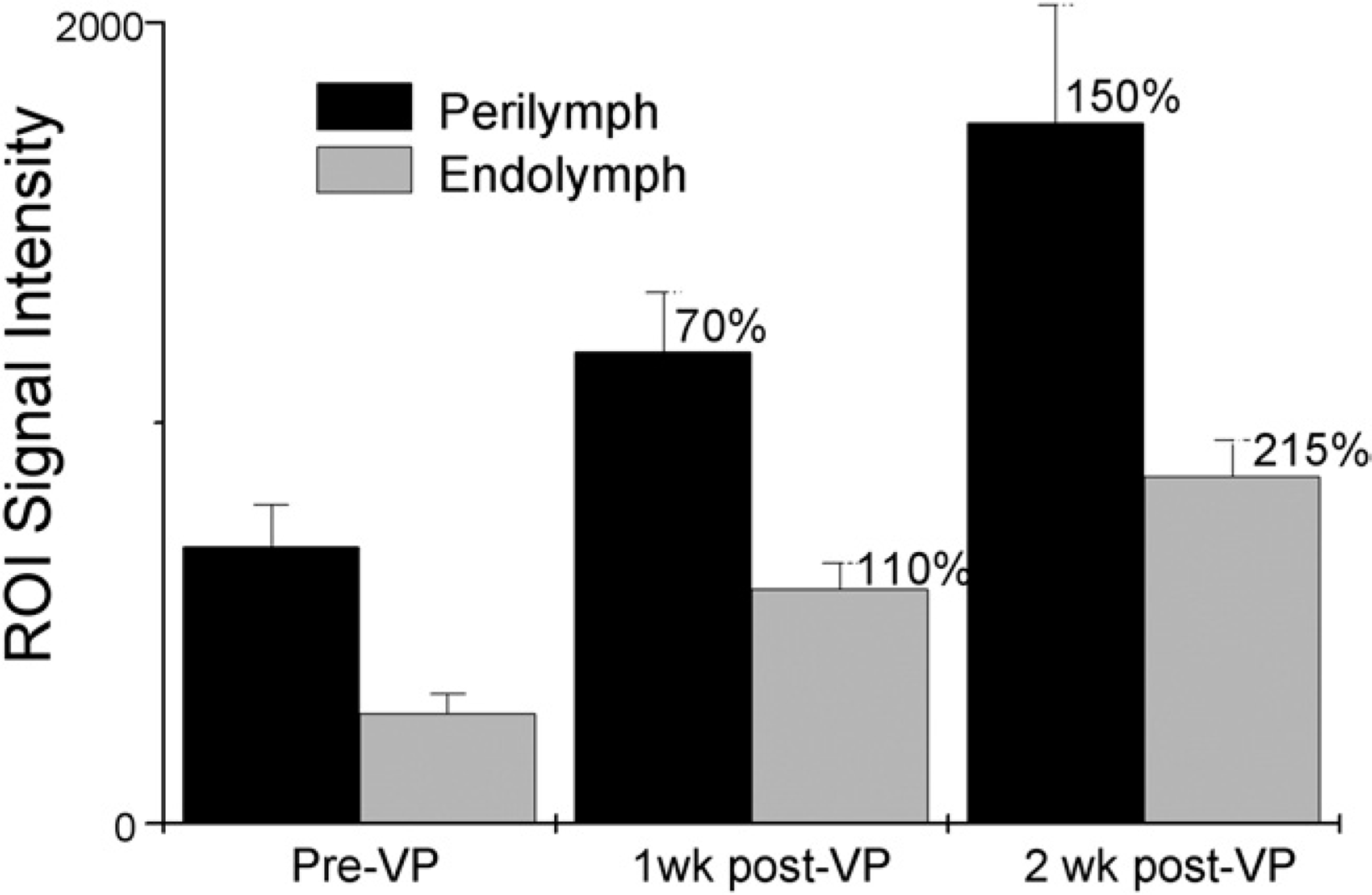
Increased region-of-interest signal intensity of perilymph and endolymph at one and two weeks after chronic administration of vasopressin. ROI, region of interest; VP, vasopressin.
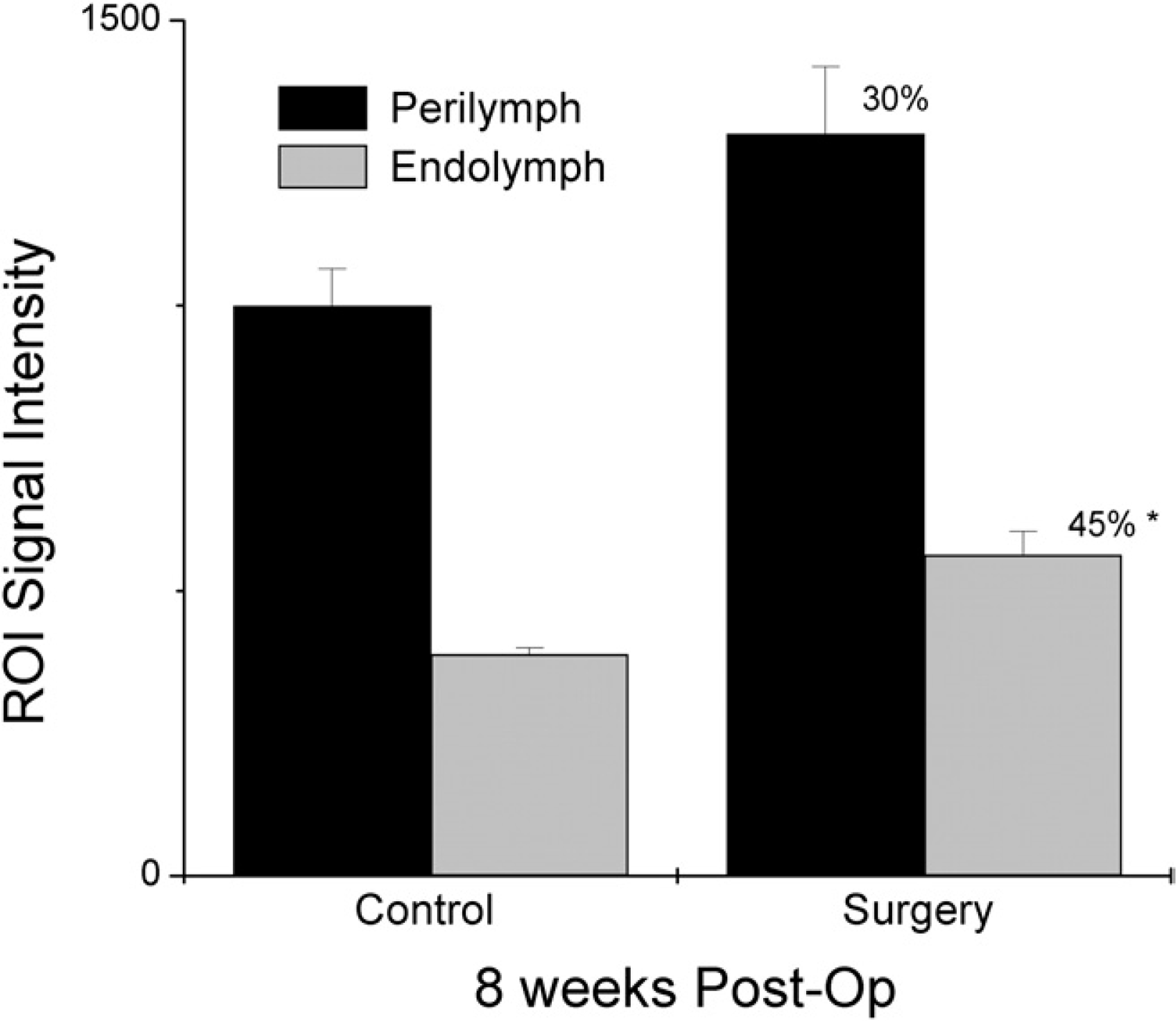
The region-of-interest signal intensity of the endolymph and perilymph increased significantly after endolymphatic sac surgery compared with the contralateral control cochlea of the same animal. Post-Op, postoperative; ROI, region of interest.
EH was quantified histologically by measuring IRs of the scala media by the formula IR (%) = (area A/area B) × 100 (Fig 5). Figure 5A shows histological analysis of the VP model, which demonstrated increased lengths of Reissner's membrane with IRs of the scala media of 15 percent to 35 percent. Histological analysis of the control cochleas did not demonstrate EH (Fig 5B). For comparison, these data are graphed with the results adapted from the IR% dose response curve by Takeda et al 6 (Fig 6). Using the highest dose of VP (1000 μU/kg/min or 2.86 ng/kg/min), we demonstrated a degree of IR similar to that seen in the previous study. Histological analysis of the surgical models demonstrated significant membrane rupture, making accurate measurement of the scala media IR impossible.
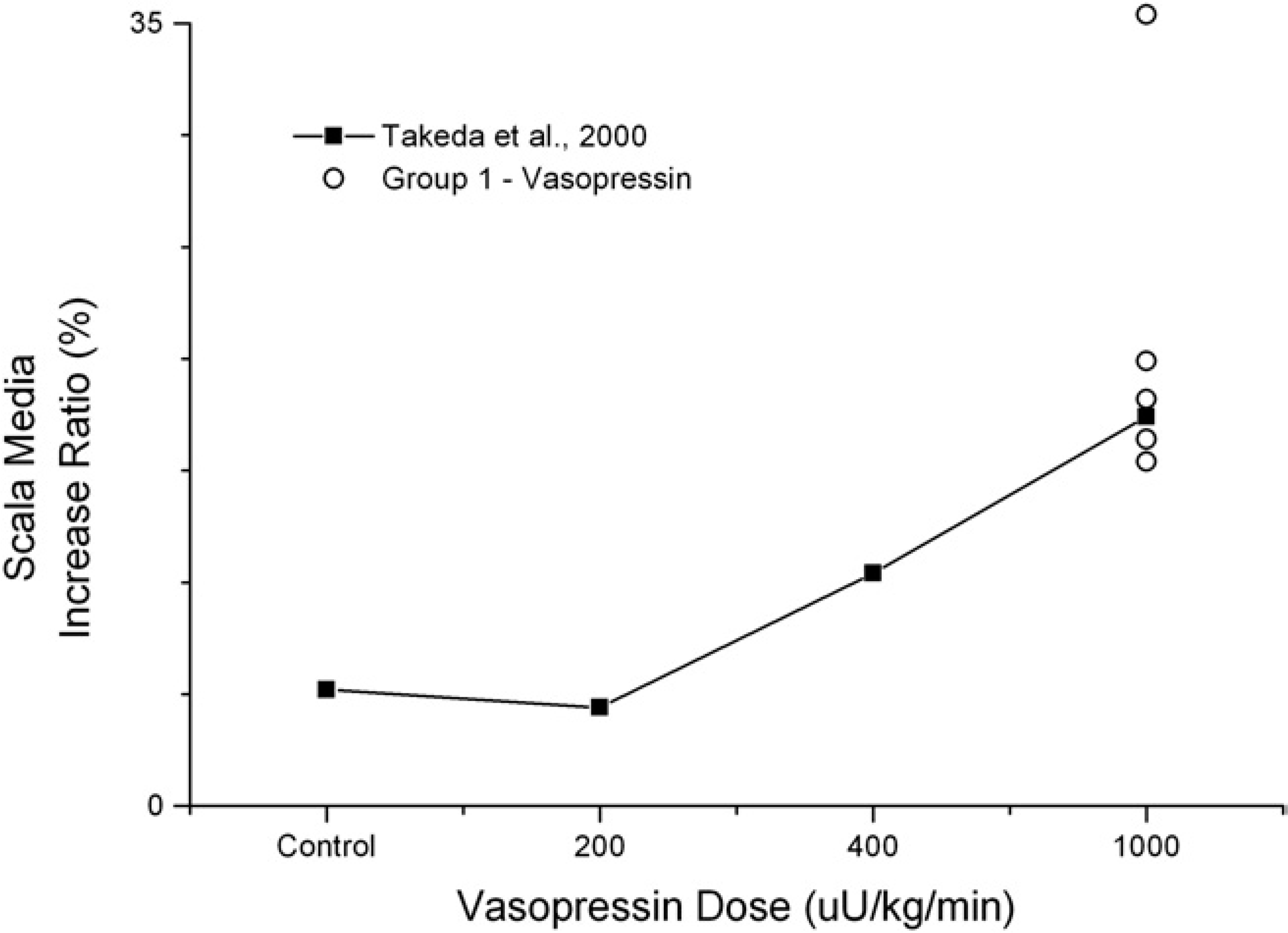
Plotting increase ratios of the scala media after chronic administration of vasopressin against original data adapted from Takeda et al 6 demonstrates a comparable degree of endolymphatic hydrops in the two studies.
(
Discussion
We have confirmed the previously reported finding that postcontrast cochlear MRI allows in vivo visualization and distinction of cochlear compartments. This is aided by the preferential enhancement of perilymph. The blood labyrinth barrier normally permits only small amounts of gadolinium into the scala media (endolymph), whereas the scala tympani and scala vestibuli (perilymph) have robust uptake of contrast agent. 13 Significant enhancement of the scala media may indicate rupture of Reissner's membrane, the physical barrier between the scala media and the scala tympani, as seen in our surgical group and VP group. In a population of experimentally damaged (noise trauma) cochleae, the scala media showed a significantly higher increase in signal intensity over the scala tympani and scala vestibule compared with intact cochlea. This suggested that damage to the cochlear membranes allowed mixing of the perilymph and endolymph as well as entry of contrast into the scala media. 13 In our endolymphatic sac ablation model, we measured a significant increase in ROI signal intensity in both the scala tympani and the scala media versus the control cochlea, but the result was less significant within the scala tympani (perilymph). The reasons behind this finding are unclear at present. Ruptured Reissner's membranes were observed histologically in several specimens in this group. One theory that explains this finding is that rupture of the Reissner's membrane allows mixing of inner ear fluids and contrast.
Our results support a role for VP as a mediator for inner ear fluid dynamics. The sample size in this pilot study was admittedly small, and larger studies are needed to further validate these findings. The mechanism by which VP permits increased passage of gadolinium into the membranous labyrinth is poorly understood. However, numerous studies have supported the role of VP activity at the molecular level in the inner ear, as well as its role in the development of EH. The VP receptor V2, previously thought to be unique to the kidney, has been identified in the inner ear. 21 Furthermore, VP receptor messenger RNA (mRNA) expression in the inner ear is downregulated by chronic application of VP. 1 Aquaporin-2 is a transmembrane water transport channel protein that plays a role in VP regulation. In the inner ear, aquaporin-2 mRNA expression was upregulated by VP and downregulated by local application of a V2 antagonist. 5 , 22 Furthermore, systemic and round window application of a V2 antagonist reduced EH histologically. 23 Our findings provide additional evidence that VP is acting on inner ear fluid dynamics at the molecular level.
Conclusions
This is the first study to demonstrate MRI evidence of dynamic cochlear fluid changes associated with high-dose chronic systemic administration of VP in guinea pigs. Our results from this pilot study support the theory that VP may be a mediator involved in the pathophysiology of MD. Furthermore, in vivo postcontrast MRI of the inner ear at 3.0 T demonstrated cochlear signal intensity changes associated with chronic systemic administration of VP and with surgical ablation of the endolymphatic sac. Larger studies are needed to further characterize and confirm these MRI findings. Understanding the in vivo MRI appearance of EH induced by various methods contributes to the future use of MRI as a tool in the diagnosis and treatment of MD.