
Abstract
Although the literature enlists “self-protection” to explain widely varied behavior (e.g., startle, defense mechanism, hoarding), the concept is not defined, nor are its varied modes classified. Few textbooks even index the term. To help remedy this odd neglect, the author postulated the protective motive as a biologically given “thrust to wholeness,” modified by experience and dependent on social support. Utilizing mechanisms of homeostasis, defense, prevention, healing, and “outreach” (i.e., wholeness expressed), the motive disposes the individual to maintain and express the functional integrity of body and self (including “all that is mine”). Seemingly unrelated phenomena (e.g., immune response, quest for immortality, forgiveness, sharing, urge to procreate) are thus afforded common explanation. Prospectively, the thrust to wholeness affords conceptual unity, parsimony, and pedagogical utility. Also, it may furnish a conceptual anchor for well-being and its paradoxes, self-defeat and suicide.
Nothing evades our attention so persistently as that which is taken for granted…. Obvious facts tend to remain “invisible.”
—Gustav Ichheiser, Misunderstandings in Human Relations, 1949.
Although the literature presents self-protection as the explanatory motive of numerous processes in human and animal life, it is an odd fate for the motive that, unlike other motives (e.g., achievement, affiliation, cognitive dissonance, pleasure), its aim and characteristics have not been examined or defined. Even indexes rarely cite references to it. This obscurity and oversight possibly stem from its very popularity in everyday life as the “obvious cause” of numerous types of action, so that to elucidate might seem like making obvious the obvious. Ichheiser's (1949) statement above regarding “obvious facts” seems to characterize the inert fate of self-protection.
In the present article, I seek to remove this veil and to explain the pervasive activating dynamic of the protective motive in both human and nonhuman animals. Drawing on the literature, largely from psychology and the biological sciences, my objective is to illustrate five forms of protection: homeostasis, defense, prevention, healing, and outreach, a new concept that I introduce here.
Brief Historical Sketch
Many early thinkers recognized the central place of self-protection in survival, particularly the idea that humans and animals are innately averse to perceived threats to their bodily integrity. The Stoics, Zeno (336–265 BCE) and Chrysippus (276–206 BCE), taught that both human and nonhuman animals are innately oriented to preserve their lives and avoid injury (Reale, 1985). Thomas Aquinas (1225–1274) regarded the “natural inclination” in humans toward self-preservation, along with procreation and care of the young, as a fundamental principle of natural law (D'Entreves, 1959). For Spinoza (1677/2000), the term conatus referred to the tendency of all things to persist in their essential nature; for nonhuman organisms, conatus is the striving to persist, and for humans, it is the awareness of thus striving. Hobbes (1651/1958) asserted that people, fearing for their lives from internecine carnage, preferred to surrender their liberties for the benefits of safety under a ruler powerful enough to secure order. For both Spencer (1862) and Darwin (1859/1981), the struggle for existence is the explicit manifestation of the instinct of self-preservation.
For Sigmund Freud (1961), self-preservation represents the “true life instincts,” synonymous with sexual energy or Eros, energized by the germ cells. These instincts are first manifest in prompting the individual toward basic narcissism and in the promptings of hunger, fear, and avoidance of bodily harm, thus preserving the organism “for a comparatively long period (p. 34).” Second, Eros prompts the organism to procreate by uniting with germ cells external to the individual, thus bringing about vicarious immortality of the individual and direct preservation of the species.
Similar to Freud, but without pinpointing a specific endogenous force, Rogers (1963) proposed that animal organisms possess an “actualizing tendency,” a directive energy to maintain and safeguard their integrity, reproduce themselves, and fulfill their potential. In contrast to both Freud and Rogers, for B. F. Skinner (1974), behavior that enables species to survive and thrive is not determined by instinctive inner impulsions but is rather the adaptive outcome of diverse contingencies of their reinforcement history.
The Present State of Self-Protection
In the early decades of the 20th century, American psychology and medicine seriously promoted research on prevention of mental illness and other health problems, but interest waned in later decades. Recently, prevention has come of age (Coie et al., 1993; Heller, 1996; Kaplan, 2000; Reiss & Price, 1996). More recently, a special issue of American Psychologist (Weissberg & Kumpfer, 2003) was devoted to “Prevention that Works for Children and Youth.”
This reentry reflects the growing recognition by the public at large, the medical community, and the government that psychology as science and as helping profession may help prevent and allay human suffering and reduce the related economic burden. These concerns have brought psychology alongside other social and biological sciences and medicine to seek ways to prevent and overcome such health-related problems as obesity, drug addiction, sex-related disorders, stress, anger control, and prescription noncompliance (see, e.g., Health Psychology and Psychology and Health). Similar concerns are central in a number of books on people's protective but often complacent behavior in hazardous working conditions in industry (Hale & Glendon, 1987), in health-related behavior (Maddux & Rogers, 1983; Weinstein, 1993), and in everyday life (Weinstein, 1987).
Self-protection is cited in a few psychological reference works (e.g., Drever, 1956; Reber, 1985) as a motive that enters into broad areas of human behavior. However, I failed to find any reference that defines the term, classifies or explains its varied manifestations, or addresses the distinction between body protection and self-protection. The literature in social psychology does refer in various contexts to self-protection but the meaning appears limited to defense of the individual's identity or self-esteem. However, several texts in social psychology (e.g., Kenrick, Neuberg, & Cialdini, 1999; Lord, 1999) do discuss some aspects of self-protection in various contexts and refer to it in their indexes. In a related area, the Journal of Personality (Cramer & Davidson, 1998) devoted a whole issue to the long-neglected concept of defense mechanisms. These stirrings appear reflected as well in the Annual Review of Psychology, beginning in the 1990s, in the increased attention to self-protection, primarily as defense or prevention.
Despite these widely dispersed references to protective behavior, the literature is silent about the meaning and ultimate objective of the protective proclivity itself. 1 If elucidated, the concept would deepen understanding of its widespread but casual usage in the literature and provide theoretical and practical advantages for psychology and related fields. Toward this end, I bring together here five categories (modes) that share the common goal of assuring body and self “wholeness.”
Meaning and Composition of the Protective Motive
The word protect derives from the Latin roots pro (before) and tegere (to cover), meaning to defend or prevent against harm. Although a helpful start, the latter stops short of conveying why anyone, human or nonhuman, tends to be “against harm.” Seeking insight, I examined literature in psychology, biology, ethology, anthropology, and sociology, where the term protection was used to explain the why of processes or actions in diverse contexts. However, beyond naming the term as if it had a well-known, clearly defined scientific meaning, no further elucidation was given.
It eventually became clear that, regardless of whether the phenomenon to be explained concerned breathing, startle, hoarding, healing, aggression, immune response, identity maintenance, alliance-making, or sun bathing—the protective aim was implicitly embedded in the context of the given actions and processes themselves. By asking of these phenomena, “What was their implicit end goal for the body or the self?” every instance appeared to meet in the same endpoint: to attain, maintain, or restore functional wholeness.
However, whereas body preservation pertained to automatic biological processes (e.g., reflex, sleep, digestion, immune response, blood clotting), self-conservation pertained to people's voluntary actions (e.g., identity maintenance, making excuses, affiliation, restoring self-assurance, self-actualization). Accordingly, despite their common end goals and some overlap between them, these two aspects of protection should be distinguished as follows.
By body preservation, I refer to the automatic engagement of biological mechanisms in the individual (human and nonhuman) to keep the anatomical organism whole and functioning with relatively undiminished effectiveness to fulfill its physiological needs. By self-conservation, I refer to the prompting within a person to maintain intact a felt center of experience and agency: “my self.” The latter comprises relatively stable but modifiable attributes which the self-as-agency, by managing its cognitive, emotional, and volitional resources, is prompted to maintain or reach toward optimally feasible wholeness. (See “The Me and the I” in James, 1892/1915, chap.12; and Osborne, 1996.) Thus, although distinguishable, the normal aim of both preservation and conservancy is to keep body and self optimally whole and functional.
A Taxonomy of Protective Modes
The goal here is twofold: First, to demonstrate that, despite the diversity of processes and behaviors that characterizes the protective impulse, they converge toward the same overall end, wholeness of body and self; and second, to provide a basis to formulate a comprehensive definition of the protective motive. I believe this foundation will provide insights into the character and scientific potential of the protective motive.
The taxonomy comprises five modes: homeostasis, defense, prevention, healing, and outreach. Each mode is exemplified below under two aspects relating respectively to preservation of the body and conservation of the self.
Homeostasis: Engendering and Sustaining Individual Wholeness
Body Homeostasis
Cannon (1929) proposed the now commonly accepted understanding that, to survive as an intact, functioning entity, each organism maintains a relatively stable internal milieu or homeostasis. This includes the functions of respiration, temperature stability, restoration, the sleep cycle, drinking, feeding, metabolism, micturition, and defecation. Despite much advance on these functions since Cannon's publication, the full details of the life-maintaining processes of homeostasis are not yet fully known (Koshland, 2002). However, there seems to be general agreement that they operate according to a genetic pattern for each species and are expressed uniquely in each individual.
Recently, Recordati (2003) offered the hypothesis that homeostasis is achieved by differential action of the hypothalamus on the autonomic nervous system. When stimulated by exteroceptive information, the hypothalamus acts on the sympathoadrenal network that activates the visceral apparatus to enable appropriate behavioral interaction with the external environment. On the other hand, when stimulated by proprioceptive (i.e., interoceptive) signals, the hypothalamus acts on the parasympathetic system to prepare the visceral apparatus to direct actions of the organism upon itself, thereby maintaining internal stability. Thus, in contrast to Cannon's (1929) view that functional stability is achieved by the autonomic nervous system as a whole, Recordati's hypothesis specifically narrows it to the parasympathetic portion.
Besides the latter adaptive internal and external accommodations, the organism normally maintains a fairly even emotional state. Surges in hedonic tone, either pleasant or unpleasant, tend to fade automatically by the operation of an “opponent process” in the central nervous system, thereby averting their disorganizing physiological impact (Solomon & Corbit, 1974). Thus free of undue emotional distress, the organism can function more effectively both internally and externally in interaction with the environment.
Homeostasis Can't Do it Alone
Despite the exquisitely integrated mechanisms for maintaining body wholeness, homeostasis requires outside assistance to adequately perform its operations. Research has long established that intake of victuals is not alone sufficient for the human infant to thrive. When infants are nourished without being warmly held, cooed to, and lovingly stroked with motherly attention, they show signs of depression and facial rigidity; smiling and crying diminish or disappear, and physiological disturbances such as erratic sleep and diarrhea set in (Ribble, 1943; Spitz, 1949). Similar behavioral pathology is induced in monkeys when deprived of “contact comfort” from a mother or surrogate (Harlow & Suomi, 1974). Rat pups deprived of maternal tactile stimulation for a day reveal a rise in corticosterone and related stress hormones, but a return to tactile contact even by human stroking restores the stress hormones to normal (van Oers, de Kloet, Whelan, & Levine, 1998). The effective functioning of homeostasis, thus, depends on positive social input, a necessity that pertains as well, as discussed later, to the succeeding protective modes.
Self-Homeostasis
At least since James (1892/1915), it has been commonly held that humans generally seek to maintain a stable valuation of the self, to project a preferred image to others, and to resist interference with this aim (see, e.g., Buss, 1996; Higgins, 1989). However, unlike the relatively fixed neurochemical processes in body homeostasis, the range of reasons and actions available to maintain stability of the self varies widely in response to situational contexts. Two-year-olds, for instance, may steadfastly oppose ordinary requests or demands with “No!” as if to proclaim “this is where I stand.” Such negativity, termed reactance by Brehm (1966), characterizes adults as well when asserting their preferences and choices, thus proclaiming their selfhood. McGuire and McGuire (1988) similarly reported that American children in Grades 5 to 11, in answer to the question, “Who are you?” referred to those attributes that distinguished them from others, such as hair color, height, weight, or birthplace, thus ostensively asserting their identity.
Similarly, although seemingly unrelated, the adult Kwakiutl of British Columbia, by engaging in their culturally accepted potlatch (i.e., the transfer and exchange of huge gifts during feasts), maintained their self-image or enhanced their prestige (Gil-del-Real & Brown, 1980). Again, people seek affirmation of their self qualities by comparing their looks, capabilities, health, or achievements with others whose relevant features are like theirs but do not overshadow them (Festinger, 1954; Tesser, 1988). Just as young organisms require tender nurturance to function normally (see above), so too do adult humans warm to those who affirm their preferred self-image (Golwitzer, 1986).
There may appear to be no commonality between young children's negativism, older children's assertions of self-distinctness, adults' comparing themselves with others, the Kwakiutl potlatch, or seeking the company of others who affirm one's self-image, but the apparent disparity is erased when the various strategies are seen as manifesting the same compelling aim: to assure or enhance functional wholeness of the self. Without dependable internal standards of reference that constitute a stable, nonfractured self-image to make decisions or maintain smooth commerce with one's environment, self-guidance would hardly be possible. Not only the self alone but also others need to know what to expect from us. The thrust, therefore, to protect (i.e., conserve) the self impels the individual to maintain a relatively stable self-image (see Figure 1).
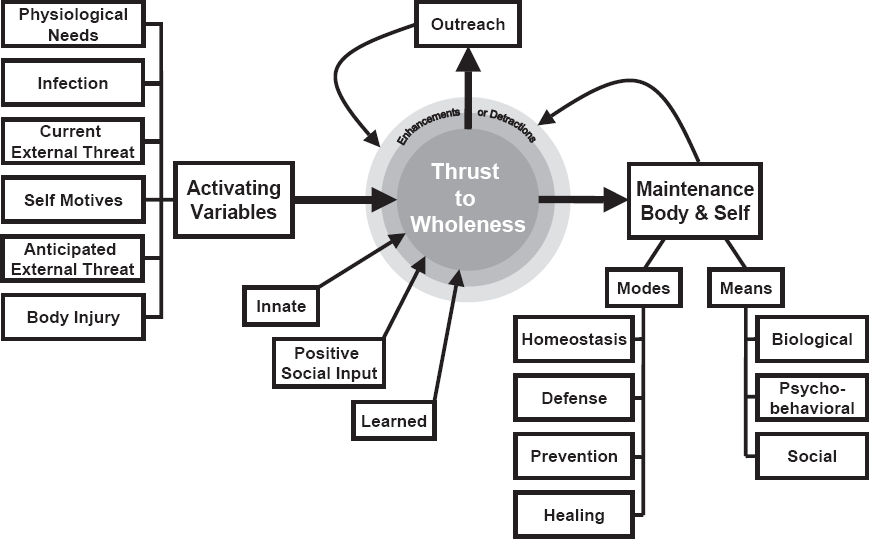
The thrust to wholeness: A schema of the protective motive.
The Melding of “Me and Mine”
Self-homeostasis is not restricted to the self as if it were a constricted, self-contained entity; the self, rather, extends beyond its boundary to include “all that is mine” (James, 1892/1915; see also ego extension [Allport, 1955] and self-expansion [Aron, & McLaughlin-Volpe, 2001]). These self-enlargements may include one's offspring and family, ethnic or religious identification, beliefs and values, or tangible possessions. To fulfill their best interest fulfills “mine” (see, e.g., Aron & McLaughlin-Volpe, 2001; Brewer, 1991). Even among animals, although the range of concern beyond the individuated organism is exceedingly more restricted than among humans, care for the young is often intensely dedicated, especially in avian and mammalian species (Masson, 1999). The protective motive, thus, normally thrusts for wholeness beyond the “separate me” to embrace “all that is mine.”
Defense: Resisting Body Injury and Self-Diminution
Defense of the Body
Whereas homeostasis is directed to the ongoing upkeep of wholeness of body and self, defense is directed to averting or counteracting injury to the body or self, an innate proclivity called harm avoidance (Murray, 1938). In the face of threat or imminent injury, swift subcortical processes are set in motion in the sympathoadrenal system and the hypothalamic-pituitary-adrenal axis (Bard, 1934; Cannon, 1929; Recordati, 2003). By this alarm response, the organism is aroused for whatever overt action appears necessary to cope with the danger (McEwen, 1998; Selye, 1976). Concurrently, autoreactive receptors of the immune system deployed in the lymphoid tissues innately recognize pathogenic microorganisms and unleash antibodies against the infectious invaders while the blood cells provide nutrients for tissue repair (Hoffman, Kafatos, Janeway, & Ezekowitz, 1999; see the “Healing and Soothing: Restoration to Wholeness” section below).
Behaviorally, in assessing risk, the general patterns and defensive actions among animals are similar (Blanchard, 1997). In a sudden confrontation with a feared predator, animals freeze all motion, but if the predator continues to approach, freezing gives way to escape or, if not possible, to exhibit a threatening stance. If the predator approaches still closer, even smaller (wild) animals may attack the larger. Organisms are thus enabled both by cellular processes and behavioral responses available within the organism's repertoire to swiftly muster resources to defend against internal and external threat.
Wariness of the “Bad-for-Me”
Preparedness theory (Seligman, 1971) suggests that evolution has predisposed humans and other animals to form aversive responses more readily toward certain stimuli and situations than to others. Experimental evidence for this aspect of the theory appears confirmed, suggesting that some behaviors appear evolutionarily significant (see review by McNally, 1987). Manifestations of insecurity include infant distress, apparently unlearned, at separation from the mother (Bowlby, 1969). Human adults, when eating alone, look up and around more often (Eibl-Eibesfeld, 1989, pp. 386–387), and they make more defensive appraisals of angry and fearful faces (Öhman, 2002). Even without conscious awareness, adults appear capable of evaluating certain external stimuli as bad (Chen & Bargh, 1999).
However, Öhman, Flykt, and Esteves (2001) caution that research has not shown conclusively that humans are prepared by evolutionary history to respond visually with fear to potentially harmful stimuli such as snakes and spiders. Even the avoidance by most infants of a drop-off is not innate, as shown by Adolph (2000), but rather the cumulative effect of learning the consequences of a drop-off upon losing balance for the postures in which they have had prior experience. These studies confirm the common understanding that action against threat rests variously on the perceptual preparedness toward fear-relevant stimuli conveyed by biological evolution and on learning of skills acquired by experience.
Defense of the Self
Beyond preservation of the tangible organism against injury or death, the protective motive in humans is manifest also in defending the intangible self against wholeness diminishment by personal failure or social disparagement. At least since James (1892/1915, Freud (1911/1959), and Adler (Ansbacher & Ansbacher, 1964), the consensus in psychology is that humans tend to be guarded against blame for their actions or for insults to their self-identity, dignity, status, or self-esteem (Buss, 1996; Fiske, 1993; Tesser, 1988). People tend to generate reasons (excuses) to persuade self and others of their rectitude, thus to undercut the affront and to conserve self-identity (Snyder & Higgins, 1988).
In clinical contexts, such wholeness-defensive measures have been interpreted as resulting in tension reduction and described variously as “safe feeling,” or rest (Ansbacher & Ansbacher, 1964), as self-sufficiency (Horney, 1950), or as security (Rogers, 1951). In the context of wholeness, the tension relief can be explained as likely arising from one's awareness that the body or self has emerged relatively undiminished structurally and functionally.
Unconscious defense of the self
Unconsciously, the self is conserved by defense mechanisms that are set off by anxiety against the threat of internal disruption from painful memories or from motives and desires felt as shameful (S. Freud, (1911/1959). Distorting one's motives and conflicts, or even reality itself, is a defensive device that enables the individual to maintain a sense of internal unity and thus to be able to present an outward appearance of internal calm (Cramer, 1998; A. Freud, 1936).
Interpersonal and social defense
At the interpersonal level, defense is manifest in the tendency for the individual to affiliate with others who, like oneself, are faced with a common danger (Schachter, 1959). In stressful conditions, such as threat to bodily safety, the impulse to affiliate is especially prominent in the female of some species, including the human, particularly during care of offspring, to tend the young and befriend others, thereby providing firmer ground for preserving themselves and their young (Taylor et al., 2000).
In the larger social arena, human and nonhuman animals (especially apes) seek security through surveillance and policing against internal insurrection and alien incursion (Allee, Emerson, Park, Park, & Schmidt, 1967). Despite the often bloody struggle for existence, Kropotkin (1902) garnered field observations, and Allee et al. provided experimental evidence that animals confronting threat band together for meeting the common needs of living in community (see Dugatkin, 1997). Among humans, Le Dantec (1918) similarly concluded on examining both the historical and contemporary records that people are never more drawn together in mutual defense to safeguard their group or uphold their group honor than when faced with a natural disaster or a hated enemy.
Prevention: Safeguarding against Injury
Prevention in Animals
In contrast to defenses against immediate threat, preventive behavior is directed toward future exigencies that preempt or dampen their force. However, compared with the broad-range impact of foresight in humans (see below), animal preventive behavior is limited to instinctive stereotyped preparations in advance of need. To facilitate reproduction and care of the young, female mammals prior to parturition tend to retreat to a secluded area, and birds prepare nesting areas according to the instinctive recipe of each species (Allee et al., 1967; Masson, 1999). Similarly, unvarying stratagems are common in the food hoarding of ants, bees, and squirrels, hibernation preparations among beavers and bears, seasonal migrations of various mammals, fish, and birds either to obtain food or to facilitate mating in nonobtrusive environments. Noteworthy, especially among mammals (including humans) and birds, is parental concern with the safety and training of offspring, thus assuring continuity of the organism and survival of the species (Bell, 2001; Griffin, 2001; Masson, 1999; Taylor et al., 2000).
Prevention in Humans
The resource specifically correlated with the ability to project the past as foresight into future possibilities, called “mental time travel” (chronesthesia), makes it possible to anticipate a broad range of potential threats and to take appropriate actions (Tulving, 2002). Prominent examples in everyday life include laying away provisions in time of plenty in anticipation of future scarcity, buying insurance policies, or taking prophylactic measures to avoid ill health. At the social level, individuals may prevent harm to kin and to valued others by alerting them to prospective dangers, or people may form group alliances against foreseen dangers. Chronesthesia, thus, constitutes a formidable instrument in conservancy of self and all that is mine.
Search for Perpetual Wholeness
Perhaps no planning evoked by mental time travel is more characteristic of humans than the near-universal practices to outwit oblivion at life's end. Elliot Smith's self-preservation theory (1919) provides abundant evidence from ancient texts and from more recent anthropological studies that the dread of mortality (i.e., the demise of wholeness) and the urge to prolong life are deeply embedded in the human psyche. This is reflected in rituals to ward off personal death and to animate the corpses of deceased relatives for life beyond the grave.
Kluckhohn (1942) proposed that such practices are magical expressions to defend against anxiety over anticipated dangers in the human condition (e.g., sickness, retaliation, famine, death) and, when expressed by many in words and actions, become a group's cohesive myths and rituals. Even in modern society, according to terror management theory (Goldenberg, Pyszcznski, Greenberg, & Solomon, 2000), the dread of death remains a potent force, although often expressed in subtle ways by preventive maneuvers against its inevitability (e.g., by doing good works that reflect important values of one's culture). One thus seeks to lay a firm basis to assure perpetual identity.
Healing and Soothing: Restoration to Wholeness
Unlike inert matter, human and nonhuman organisms, when damaged, do not stay put. The tissues trend back toward some semblance of wholeness. The self, too, when hurt appears to operate as if directed by an image of its pre-hurt state or an improvement of it (see Dubos, 1990; Greenwald, 1980). These restorative tendencies constitute the healing mode of the protective motive.
Healing the Body
Injury to the body evokes a healing response to restore the body to wholeness. At the moment of injury, inflammation occurs, shown in three classic signs: heat, redness, and swelling; later, pain and loss of function may be exhibited. These signs, detailed by Mitchell and Cotran (2003), involve a cascade of regulated events that function to (a) dilute and destroy invasive foreign matter such as bacteria, viruses, fungi, and other toxins, (b) debride necrotic tissues, and (c) set in motion the regeneration of injured cells and their replacement with new cells of the same type. Cytokines mediate the processes of both inflammation and the inducement of fibroblasts, which, in turn, stimulate synthesis of tissue-restoring collagen.
Chemical changes at the wound site initiate a rise in vascular permeability, permitting red blood cells to enter and causing the familiar vascular response of the inflammatory reaction. Released to the site from the blood are plasma proteins, neutrophils, and macrophages, and lymphocytes are deployed from the contiguous lymphoid tissues. Within 24 hours, the neutrophils and macrophages begin the process of phagocytosis (i.e., ingesting, killing, and degrading the ingested particles) and debriding the wound.
Concurrently, cells from the surrounding tissues begin the mitotic process of regenerating the injured cell tissues, thus restoring the parenchyma. If, however, the defect is too extensive or the inherent repair capacity of the tissues is exceeded, collagen fibrils from connective tissue undergo fibrosis (i.e., form scar tissue) to bridge the gaps.
Amazingly, past experience plays a vital role in this defensive/healing drama. Not only do lymphocytes innately recognize the molecular patterns of pathogens, but once having unleashed antibodies against particular microorganisms, their ability upon subsequent invasions to recognize and eradicate them is heightened (Hoffman et al., 1999; Mitchell & Cotran, 2003). Cell deployments, however, vary greatly, depending on the organ injured, type and frequency of injury, health status and age of the individual, and genetic variations (Mitchell & Cotran, 2003). However, despite their bewildering specificities, they share the same endpoint: restoration of the body to full or approximate wholeness.
The Place of Social Support in Body Healing
As in the other modes, many studies have indicated that a lack of social support also adversely affects the healing function. For example, stress and negative emotions induced by a lack of social support appear to prolong infection and inflammation in a broad spectrum of conditions. These include cardiovascular disease, osteoporosis, and Type 2 diabetes, whereas close personal relationships, by their positive impact on the immune and endocrine systems, appear to guard against or diminish susceptibility to infection and inflammation (Kiecolt-Glaser, McGuire, Robles, & Glaser, 2002). Not only does receiving social support buffer against the negative effects on the immune system, but also individuals whose sociability facilitates high-quality social relationships are subject to decreased probability of developing a cold (Cohen, Doyle, Turner, Alper, & Skoner, 2003).
Why it is that stresses caused by insufficient or disruptive social relationships impact adversely on the immune and endocrine systems is not yet fully clear. But recent studies indicate that chronic stress, such as experienced by parents in caring for children with cancer, causes the healing effects of the inflammatory response to become injurious (Miller, Cohen, & Ritchie, 2002). Similarly, the positive effects of the inflammatory response are reversed by the periodic stress imposed upon submissive mice by placing an aggressive intruder into their cage (Avitsur, Stark, & Sheridan, 2001). In both studies, the high levels of cortisol associated with the prolonged stress appeared to cause glucocorticoid resistance in the inflammatory component of the immune cells. Thus, chronic diminishment of wholeness due to stress impaired the normally adaptive turn-off of the immune cells by the normally responsive regulation of the endocrine hormone.
Body Healing: Conclusion
In the present context, the above observations imply that close social relationships positively affect the quality and extent of self-wholeness. In turn, self-wholeness buffers stress, infection, and inflammation, and when the latter do occur, self-wholeness speeds the body's healing processes. Although the physiological mechanisms are not yet fully clear, it appears evident that wholeness of the body is helped or hindered in the measure of wholeness of the self. As the late René Dubos (1990) put it, “everything we experience becomes incarnated… in our physical and mental life.” (For a fine overview of mind-body interactions in health and disease, see Ray, 2004.)
Healing the Self
As noted earlier, regarding the defensive process of fending off affronts to self-regard by excuses and defense mechanisms, people are similarly inclined in periods of dented identity to utilize various stratagems to restore their integrity. Normal children when anxious may overcome distress by thumb-sucking (Foster, 1998), conversing with imaginary companions (Friedberg, 1995), or finding solace in clutching a security blanket (Ybarra, Passman, & Eisenberg, 2000). Similarly, to compensate for a sense of emptiness, adults may fill up with food (Cushman, 2003) or turn to the ages-old practice of finding a listening ear (Jackson, 1992; Pennebaker, 1990). In all these instances of challenges to wholeness, despite their variety, children and adults alike appear compelled by the same dynamic: to restore themselves to wholeness.
Even a common minor frustration, the “tip-of-the-tongue” phenomenon—the occasional inability to call to mind a desired word—may set off an intense effort to recapture the reclusive word, a type of “inarticulate psychosis,” as James put it (1892, p. 249). In the present context, this scrambling appears to reflect a need to reverse the affront to the individual's self-image as someone capable of freely producing words. If, finally, the sought-after word rises to memory, felt wholeness is restored and, at once, relief is regained (Brown & McNeill, 1966). The pinch into wholeness by a word-finding difficulty is insignificant compared with the deeply felt wounds from offenses by others; nonetheless, if healing occurs, it occurs in both instances by a restoration of the self to wholeness.
Forgiveness: Soothing the Wounds of Undeserved Hurt
Forgiveness is an individual's choice to abandon resentment and seek reconciliation with someone whose actions are felt as patently inconsiderate or unfair (see, e.g., Enright & Fitzgibbons, 2000). In so doing, both the forgiver and the person forgiven, but especially the former, experience lightening of mood, reduced rumination and distancing, and improved well-being. The benefits are significantly marked in close, strongly committed marital relations (Karremans, Van Lange, Ouwerke, & Kluwer, 2003). Even merely imagining forgiving someone for an actual wrong to oneself has been shown experimentally in immediate lowered electromyogram of the brow, lowered skin conductance, heart rate, and blood pressure from baseline (Witvliet, Ludwig, & Vander Laan, 2001).
Seen within the present schema, the latter findings suggest that the forgiving individual senses that repairing the rift between self and other would ease the disruptive burden of resentment on one's cognitive and emotional well-being, thereby inching the self back to wholeness. The benefits are especially heightened in close relations, because in the resentful, nonforgiving state, the other, currently alienated from one's extended self, can now be reincorporated into “all that is mine.” Why, then, do people often resist forgiving others (see, e.g., Enright & Fitzgibbons, 2000)?
Ironically, if the resister perceives forgiveness as symbolizing surrender to a deeply felt injustice, this route means not a return to but a retreat from wholeness. Here, the wholeness perspective, in line with discrepancy theory (Higgins, 1989), suggests that forgiveness implies to the resister a succumbing to injustice, thus more dissonant with the moral self than stoking the fire of righteous indignation. Hence, in such instances, by not forgiving, the individual may feel more protected, more whole, than by forgiving.
Outreach: Wholeness Expressed
Given that the trust to functional wholeness is the ultimate objective of the protective motive, might it be that, to the extent that this aim is met, the thrust does not become inert but rather unceasingly maintains momentum in mental and behavioral activity? If so, by what signs might this outreach be recognized? Considering the question led me to draw on two bodies of evidence, particularly among humans, starting in the earliest period with spontaneously emitted behavior and followed at later maturation with self-directed action.
Body Outreach
At birth, humans appear endowed with energy to express their capacities spontaneously in a patterned sequence in accord with their stage of development. Although the early outreach actions of the infant have no specific direction, they appear nonetheless as the unreflective exercise of the capabilities made possible by the infant's level of wholeness. According to the careful studies of Gesell (1929, 1952), the human neonate spontaneously emits short, random movements and occasional smiles. Somewhat later, these are followed with mass neuromuscular activity and then followed successively by reaching, creeping, crawling, standing, walking, and exploring (see also Schachtel, 1959).
In these early activities, and even later in the child's more vigorous play, the neuromuscular actions do not appear directed to satisfy vital tissue needs but rather as energy emitted by the functional capacity of wholeness (cf. White's “effectance”; 1959). So impressed was Rogers (1963) by this “actualizing tendency” as an innate potential not only in humans but within each living organism that he ascribed it even to the lowly potato in a dark cellar that emits shoots groping toward light.
Even the child restricted by physical handicaps is not fully deterred from expressing some degree of motor activity in keeping with its maturational level (Gesell, 1929). Also, brain wounded soldiers, greatly handicapped, make efforts to test the limits of their movement capacity (Goldstein (1939/1995). This irrepressible urge to test their compromised limits led Goldstein to conclude that “[the] tendency to actualize its nature, to actualize ‘itself’ is the basic drive, the only drive by which the life of the organism is determined” (1939/1995, p. 162).
The latter observations and viewpoints led me to conclude that an initial sign of wholeness is manifest in overt neuromuscular motility. Somewhat later in development, signs of covert cognitive capacities can be inferred from the child's early discriminative abilities. For example, infants prefer complex figures over simple lines and the human face over geometric designs (Fantz & Yeh, 1979). Also, infants in their first year of life show surprise by looking reliably longer when an adult in their presence acts inconsistently with what she knows (Onishi & Baillargeon, 2005). 2 (However, Perner & Ruffman, 2005, question this early interpretation of the results.) This inferential capacity adds to a growing literature indicating that, in their first year of life, infants can perceive and reason about others' goals, dispositions, and emotions. Thus, the signs of outreach in the child can be noted both in its freely expressed actions and its discriminatory and inferential ability.
However, outreach is unique among the protective modes. Whereas the other modes, such as homeostasis or healing, provide direct avenues toward wholeness, the early manifestations of outreach appear initially as emergent from, but not contributing to, wholeness. These early motor stages nonetheless appear to be an essential precursor to the enhancement of wholeness, as discussed next.
Self-Outreach
When awareness of selfhood arises in the child as an autonomous center of control, the initially undirected manifestations of body outreach gradually come under the child's emerging cognitive capacity. No longer a mere vehicle for the outflow of energy, the child now couples perception of his or her own movements with the awareness that it can direct those movements to test how far and how well his or her competence will allow: “Can I do this?” (Piaget, 1952; Thelen, 1995).
The intrinsic urge to test its competence and overcome obstacles suggests that outreach directed by the self likely starts when the spontaneous outreach of infancy recedes and awareness of oneself as an agent emerges (see Allport, 1955; Schachtel, 1959). The child now tends to reach beyond innate competencies toward the exploration of untried avenues of self-expression, the things it likes to do (Deci, 1975; White, 1959). These tendencies, emerging from genetically based motor patterns in interaction with the child's unique experiences, have been variously called “striving for superiority” (Ansbacher & Ansbacher, 1964), “becoming” (Allport, 1955), “self-realization” (Goldstein, 1939/1995), “self-actualization” (Maslow, 1970; Rogers, 1951), “actualizing tendency” (Rogers, 1963), “emergence from embeddedness” (Schachtel, 1959), and “self-transcendence” (Frankl, 1966; Weiner, 1993).
The deaf and mute Helen Keller (1896) caught the irrepressible spirit of outreach in her words: “One can never consent to creep when one feels an impulse to soar.” So strong is the propelling force of self-outreach that, if blocked from expression, wholeness of body or self may be compromised, resulting in personality disturbance, diminished well-being, and interference with the normal process of socialization (see Ryan & Deci, 2000).
Mating and procreating as outreach
A significant illustration of outreach in the adolescent and adult life of the individual is provided respectively in the urge to conjugate and the desire to procreate. I referred earlier to Aquinas (D'Entreves, 1959) and to Freud (1924), who believed that the urge to procreate is an innate proclivity of self-preservation; and later Carl Rogers (1963) viewed procreation as an aspect of the built-in tendency of the organism to actualize itself. Seen, however, from the perspective of outreach, these conceptions conflate the biological or primary expression (sex) with the psychocultural or secondary end (procreation).
In its primary expression, the appetitive sexual outreach is not to procreate but to copulate. This is apparent when seen from the vantage of nonhuman animals where the copulative response is punctuated at sexual maturity (Beach, 1947). Procreation may result, but it seems unduly anthropomorphic to attribute the urge as aimed at duplicating the organism.
In humans, rather, where cultural influences and unique personal experience become embedded into self goals (Allport, 1955; Frankl, 1966; Weiner, 1993), an additional factor intervenes. When the individual's copulatory urge has progressed beyond the sexually ready state to a self goal to procreate and nurture offspring, this desire, as Beach's (1947) comparative research suggests, more likely reflects the shaping influences of one's culture, nurturing temperament, and personal experience. Only then, in this indirect way, does it seem appropriate to attribute the urge to procreate as a manifestation of self-preservation (Freud, 1924) or of self-actualization (Rogers, 1963). In humans, thus, the sexual urge may become either manifest as biologically based outreach as in other animals or transformed in humans into an expression of self-outreach to procreate and nurture an extension of the self.
Outreach redounds on wholeness
In keeping with the concepts of efficacy (Bandura, 1986) and effectance (White, 1959), it is likely that each episode of outreach, if successfully attained, enhances the individual's experience of his or her own body from the superior vantage of the level reached. Each such rise may enhance the individual's assuredness and impart enhanced readiness to stretch toward still higher levels or readiness to outreach in new areas. Concurrently, as suggested by the broaden-and-build theory (Fredrickson, 2001), each successful outreach evokes a positive glow that reflects back positively on the individual's general well-being and induces a readiness to engage in new areas of outreach. In this light, outreach appears not only as a dynamic emergent from wholeness but also as an inherent contributor to it (see Figure 1).
Outreach: Conclusion
The proclivity for organisms to express wholeness in overt behavior and, among humans, to reach self goals, recommends outreach for inclusion in the protective taxonomy. First, outreach flows from the state of wholeness made possible by the operation of the other protective modes; second, if outreach is blocked from expression, wholeness of body or self may be jeopardized; third, if outreach is successfully expressed, the consequence may redound to the enhancement of wholeness and thus further the well-being of body and self. Outreach, thus, merits a place as a mode in the protective taxonomy.
Defining the Protective Motive
Confronted by the many-sided character of the protective motive, my attempts to formulate a succinct yet comprehensive definition appeared insuperable. The task eased when I realized that, despite the different objectives of the respective modes, they are pointed toward the same ultimate agenda: to maintain functional wholeness of body and self. In this light, homeostasis, because of its continuous operation, plays the central role, whereas the other players (defense, healing, etc.), although readily available, are activated primarily when needed as maintenance supports. Outreach, however, not so neatly encapsulated, requires separate mention.
Bearing in mind these considerations, I propose this definition. The protective motive is the biopsychological thrust in human and nonhuman animals that requires social nurturance, is modified by experience, and induces biological mechanisms and self-directed actions to maintain, express, and enhance functional wholeness of body, self, and “all that is mine.”
Summary and Implications
Self-protection is often used in the literature of biology, psychology, and the social sciences as an explanatory motive, but the concept has until now remained undefined, an unusual fate for a widely used concept that alleges explanatory power. Seeking to clarify the construct, I have introduced here a taxonomy of its various operations (modes) organized around the construct, “thrust to wholeness.” Its central feature is the predisposition by nature and by learning to ensure the functional integrity of the body and, in humans, integrity of the self. These ends are achieved through four modes (homeostasis, defense, prevention, and healing), but the aim in each mode is identical: to maintain wholeness of body and self.
Beyond its dynamic property to maintain functional integrity of body and self, does the thrust to wholeness itself have an outward sign? Drawing on developmental psychology, I postulated that the thrust to wholeness is manifested in early infancy as the spontaneous emergence of patterned motor activity, preference for human faces and complex geometric shapes, and the ability of babies to detect deception. I call these spontaneous tendencies outreach.
In later infancy, with the further development of cognitive capacity and motor coordination, the relatively undirected energy of wholeness comes under the control of the self's wants, interests, and reasoning toward new potentials of competence. In turn, actions with positive outcomes empower the individual with a fuller sense of assuredness and autonomy.
Although the thrust toward wholeness is identical for both body and self, the means toward the respective ends can be either automatic or self-driven. Thus, whereas breathing, startle, and cell repair are normally involuntary biological processes to keep the body whole, keeping the self whole usually involves self-directed voluntary actions. I have therefore proposed that the two classes of protection be distinguished respectively as body preservation and self-conservation.
This article has emphasized that, although both classes of the protective motive reside in the individual, the motive cannot thrive in a social vacuum. The protective taxonomy outlined here has disclosed the indispensability of social support for the effective functioning of the protective modes. Although the various modes that enable the organism to function as a separate unit are rooted within the organism itself, nonetheless they cannot operate effectively in isolation, especially in the early period of life. However, throughout life as well, the individual's protective capacity is undermined (e.g., to maintain homeostasis or restore health after injury) without the input of caring others.
The self, thus, in “self-protection” is hardly possible without other selves. In turn, as the protective taxonomy repeatedly implies, being buoyed up by received support, the self reaches out vigorously to protect “all that is mine.” This force is especially salient in female mammals in the nurturing of offspring (Taylor et al., 2000). The thrust to wholeness, whether in human or animal, is thus fashioned to operate within the individual to receive and, beyond the individual, to share.
Finally, drawing on the processes and actions illustrated in the five modes, I formulated a definition of the protective motive as a dynamic within the individual to maintain and express functional wholeness. As far as I know, it is the only comprehensive yet succinct definition in the literature of this prepotent motive.
Benefits of the Protective Taxonomy
A Conceptual Lens
The protective taxonomy presented here does not introduce new data; rather, it organizes and explains existing evidence from diverse processes and actions in humans and animals. For example, it is well established that organisms strive to escape from danger or to hoard food against later need. In the present perspective, it is not enough to explain these behaviors by simply relying on the word protective. The word is acceptable, but I have suggested that, to advance our understanding, it must be grounded in the concept that the actions are impelled by innate and learned redispositions to initiate defensive and preventive actions and, furthermore, that underlying these is a directive impulse that propels them to a commonly shared end: Functional wholeness of body or self.
Similarly, the protective motive, understood as a thrust to wholeness, opens a view on vast areas of homeostasis, healing, and outreach where the motive has rarely been considered. Their inclusion together with defense and prevention demonstrates a new way of perceiving how widely differing behaviors can be explained by the relatively simple protective schema.
Conceptual Parsimony
By postulating wholeness as the dynamic thrust served by protective modes aimed at maintaining structural and functional integrity of the organism and the self, a theoretical path is cleared to see that many processes, although functionally disparate, are nonetheless equivalent in their protective capacity. In casting this light on human and animal behavior, the protective taxonomy offers a practical advantage of potentially encompassing more facts with fewer concepts. The ordering of autonomous processes side by side with deliberative actions under a few categories might rob the “luminous” status of the latter, but it has the advantage of reducing unconnected clutter. For example, automatic reflex and self-initiated self-handicapping would occupy the same taxon, defense. Similarly, instinctive animal hoarding and premeditated human alliance-making would share the same taxon, prevention. The value of such clustering lies in encapsulating vast factual and conceptual breadth under relatively few categories.
This taxonomic conception suggests a practical benefit, relatively simple to achieve, in enhancing handbooks and texts on motivation and the usefulness of indexes in general texts. References under “protective motive” would include subsections under each of the five modes, and each mode would be further subclassified under “self conservation” and “body preservation.” Not only would this arrangement help the inquiring reader to efficiently locate a particular topic but also, importantly, it would help both researcher and general reader to effectively grasp large areas of protectively equivalent information.
Pedagogical Utility
If the approach suggested here for classifying the vast variety of instrumental actions and processes is broadly utilized, it can help build a cumulative science constituted of fewer, more manageable concepts. The assembling of automatic processes (e.g., cell repair, hunger, startle, aggressive defense) and voluntary actions (e.g., excuse making, forgiveness, self-presentation, self-actualization) under the higher order aims of the protective modalities (from homeostasis to outreach) simplifies the writing and teaching of psychology, and possibly, too, related sciences. Instead of the current proliferation of ad hoc “and… and” concepts lacking a binding tie between them, the broad rubrics of self-preservation and self-conservation and their associated modes would facilitate the development in psychology and related sciences of a more orderly conceptual gestalt. Students, faced with neater, conceptually interconnected disciplines, would more smoothly learn and more readily remember (see Hintzman, 1986; Wallace, West, Ware, & Dansereau, 1998).
Whither Wholeness?
I have proposed here that a protective impulse in humans and animals directs innate biological mechanisms and learned behaviors toward maintaining functional wholeness of body and, for humans, the self. However, wholeness, the headstone of self-protection, raises questions and new vistas that beckon attention.
What Makes Wholeness Fail?
Limitations and breakdowns
Despite my portrayal of self-protection as pervasive and its effects positive, it is not an inerrant master. Limitations and breakdowns reduce the range of the generally successful wholeness-serving modes. The pains associated with babies' teething, the debilitating effects of autoimmune diseases (e.g., multiple sclerosis, rheumatoid arthritis), or the wide-ranging negative effects of chronic stress on mental functioning and emotional well-being appear to lack counteractive utilities of defense or healing; nor is the conscious self ordinarily able to counteract the sabotaging rage of an unconscious and relentlessly punishing superego.
Thus, although the protective modes help make life and the pursuit of personal goals possible, they do so imperfectly (Black, 2006a). Interdisciplinary studies in the biological sciences and medicine and in psychology and the social sciences on the causes of these shortcomings would be fascinating not only in their own right but also might contribute insights to reduce the tragic failures of self-protection on the job and in everyday life (see Hale & Glendon, 1987; Maddux & Rogers, 1983; Weinstein, 1987; Weinstein, 1993).
The seduction of wholeness
If the protective motive impels the organism and the self toward wholeness, how is one to explain the paradox, particularly in humans, of wholeness-defeating actions and even suicide (see Baumeister, 1990; Baumeister & Scher, 1988; Berglas & Baumeister, 1993)? To unravel this conundrum, I propose elsewhere (Black, 2006b) that the impulse to keep body and self protected remains uninterrupted even where the individual engages in acts that appear to the observer as obviously self-defeating or body damaging (see Adler, 1930/1937). The great protector, abetted by various easements (magical thinking, past successes, bravado) becomes itself seduced into pursuing unrealistic risks. Negative outcomes may result, affecting wholeness and well-being adversely, but the thrust to wholeness persists.
What Happens to Wholeness in Old Age and Extremity?
If, as posited here, the thrust to wholeness pervades the organism both as body and as self, by what mechanisms does this dynamism recede as the tempo of life slows and the individual approaches the final days? The protective impulse in the very old seems to retreat before the superior force of organismal degeneration. Perhaps, contrary to appearances, the retreat itself represents a built-in neurochemical process that shields the dying person (or nonhuman animal) from experiencing the trauma of decline. Exploration of this inevitable state would contribute to the final chapter in biology and psychology's book of the living (see Chong, McDonald, & Strauss, 2004; Lachman, 2005).
How Are Conflicts Settled between Body and Self?
Presumably, since self-conservancy does not appear until after the self arises a year or two after birth (Lewis, 2003) and, hence, may be relatively weak against the built-in biologically based power of body preservation, how are conflicting aims between them “settled”? For instance, if body homeostasis demands sleep, by what mechanism does the self override this insistence with its own contrary demand, say, to complete a wholeness-fulfilling task? Or, vice versa, how does the body ultimately overturn the self's veto with an unstoppable soporific shutdown?
Possibly, neuroimaging techniques might unravel the mechanisms of such conflicts in neural traffic. The prospect for deriving therapeutic insights from such research for application in disorders such as obsessive or suicidal ideation is exciting. Answers here may shed important insights on the thrust to wholeness.
Conclusion
In providing an analysis of the wide-ranging guises of the protective impulse, I have sought to bring order and definition to its heretofore loose usage in the literature. The immediate implication of this work is for this crucial motive to emerge from its murky station and be accorded the clarity that its penetration into animal and human life warrants. Beyond defensive and preventive actions in which the literature has often utilized self-protection, albeit as a mere detail, its central role should more pointedly extend as well into homeostasis, healing, and outreach. In each of these modes, as I have suggested, it is clarifying to distinguish self-conservancy from self-preservation. From this broad conceptual base, the protective motive offers a perspective for explaining far more diverse phenomena than are now perceived outside its lens.
Ultimately, the most fundamental insight may come from investigations into the neuronal and learning mechanisms that govern the various modes of the protective thrust, including those governing spontaneous and self-directed outreach. Acquiring knowledge of these mechanisms may unlock insights into functional wholeness whose benefits can now only be dimly envisioned.
Footnotes
1
After completing this work, I found what at first appeared to be an exception to the unexamined concept of protection. In her analysis of the implicit meanings of “protection,” especially used as the guiding goal of the nursing profession, ![]() unraveled one salient dimension, “vigilance,” that occurs primarily in the two interpersonal contexts of nursing, namely, managing and communicating with the patient. Thus, although her concern is with protection, it is extraneous to the present focus on the endogenous imperatives, biological and psychological, operating within the individual human or animal.
unraveled one salient dimension, “vigilance,” that occurs primarily in the two interpersonal contexts of nursing, namely, managing and communicating with the patient. Thus, although her concern is with protection, it is extraneous to the present focus on the endogenous imperatives, biological and psychological, operating within the individual human or animal.
2
After the experimenter, E, places a toy in position A, it is then moved to position B when E leaves the room. Upon returning, E seeks to retrieve the toy from position B, thus acting out of keeping with E's initial act; hence, the infants' surprise.