
Abstract
Recent theoretical accounts of emotion regulation assign an important role in this process to the prefrontal cortex, yet there is little relevant data available to support this hypothesis. The current study assessed the relation between individual differences in asymmetric prefrontal activation and an objective measure of uninstructed emotion regulation. Forty-seven participants 57 to 60 years old viewed emotionally arousing and neutral visual stimuli while eye-blink startle data were collected. Startle probes were also presented after picture presentation to capture the persistence or attenuation of affect following the offset of an emotional stimulus. Subjects with greater relative left-sided anterior activation in scalp-recorded brain electrical signals displayed attenuated startle magnitude after the offset of negative stimuli. This relation between resting frontal activation and recovery following an aversive event supports the idea of a frontally mediated mechanism involved in one form of automatic emotion regulation.
Psychologists have long recognized the importance of emotion regulation, and have attempted to describe its form and functional role (e.g., Freud, 1920/1966). Modern conceptualizations of emotion regulation have also underscored the implications of dysregulated emotion in mental illness (Gross & Munoz, 1995). The clinical importance of emotion regulation can be seen by reviewing the diagnostic criteria for Axis I and II psychopathology in the fourth edition of the Diagnostic and Statistical Manual of Mental Disorders (DSM-IV; American Psychiatric Association, 1994); many of these criteria include disturbances in regulatory processes. Cole, Michel, and Teti (1994) listed several examples of psychological symptoms arising from emotion dysregulation, encompassing many DSM-IV diagnostic categories. These symptoms include inappropriate affect, chronic worry, avoidance or constriction of emotion, extreme emotional lability, and sustained negative affect. Indeed, a lack of emotion regulation may be seen as a crucial component of depressive and anxiety disorders. For example, Nolen-Hoeksema (1991) has shown that some depressives seem to lack the ability to “turn off” negative emotions. It is possible that a subset of depressives lack a relatively automatic regulatory mechanism present in nondepressives.
The broad construct of emotion regulation may be profitably parsed into those regulatory mechanisms that are under voluntary control and those that occur relatively automatically. 1 Most of the extant literature has dealt with voluntary emotion regulation (e.g., Gross, 1998; Gross & Levenson, 1993, 1997; Jackson, Malmstadt, Larson, & Davidson, 2000), because it is relatively easy to instruct participants to suppress, enhance, or maintain an emotional response or expression. These studies have examined emotion regulation in carefully controlled experimental environments, typically following fairly specific instructions to the participant (e.g., to suppress, maintain, or enhance a brief emotional reaction, or to focus on suppression of emotional experience or emotional behavior). However, in addition to examining voluntary emotion regulation, it is crucial to develop approaches to study more automatic forms of emotion regulation, processes that may be more pervasive yet more difficult to capture experimentally. One experimental approach is to study emotion regulation in the absence of explicit regulatory instructions, as it occurs following the offset of an affective challenge. Indeed, one parameter of “affective style” (Davidson, 1998) is uniquely well suited to highlight individual differences in automatic regulatory processes: duration of emotional response following emotional provocation. Relatively fast recovery following a negative-affect elicitor may be one index of individual differences in automatic emotion regulation—that is, regulation that occurs in the absence of specific intentions to suppress negative emotion.
Recent studies conducted in our laboratory suggest that baseline prefrontal cortex (PFC) activation may be a good predictor of emotion regulation processes. In a small preliminary study with young adults, Larson, Sutton, and Davidson (1998) found that relative right-sided resting frontal brain activation (i.e., more activation in the right than the left frontal electrode site) was associated with startle potentiation following the offset of negative stimuli, supporting the idea that PFC asymmetry is an important predictor of recovery following the presentation of a negative affective stimulus. Jackson, Burghy, Hanna, Larson, and Davidson (2000) found that relative left-sided baseline frontal activation was associated with ability to voluntarily suppress negative emotion (for methods involved in this emotion regulation paradigm, see Jackson, Malmstadt, et al., 2000). More recent functional magnetic resonance imaging (fMRI) work by our group (Dalton, Kalin, & Davidson, 2002) has shown that right PFC is activated during threat of shock, a finding in line with a previous positron emission tomography (PET) experiment (Hsieh, Stone-Elander, & Ingvar, 1999) that examined anticipation of an impending but unpredictable unpleasant stimulus. In another recent fMRI study, Ochsner, Bunge, Gross, and Gabrieli (2002) reported increases in left lateral PFC activation when participants were asked to regulate (“reappraise”) emotional reactions to highly negative scenes. These studies suggest that individual differences in emotion regulation may be reliably indexed by PFC activation, and that both right- and left-sided neural circuits are implicated in these regulatory processes. These data are also in accord with the corpus of frontal lesion data, which suggest that patients with damage to particular sectors of the PFC have problems regulating emotion (Damasio, 1994; Robinson & Downhill, 1995).
A powerful tool for studying the time course of affective responding (Davidson, 1998) is electromyographic (EMG) measurement of emotion-modulated startle (for a review, see Lang, 1995). Using such measures, it is possible to probe both during and after an affective challenge to determine both initial emotional reactivity and duration of emotional response. By inserting startle probes at different points during and after an emotional stimulus, we hoped to capture a feature of affective style that may be characteristic of relatively automatic emotion regulation.
In the current study, we first recorded 8 min of resting electroencephalographic (EEG) activity using a standard protocol we adopted in many of our previous studies (e.g., Tomarken, Davidson, Wheeler, & Kinney, 1992). We then measured participants' eyeblink startle magnitude during and following the presentation of affectively arousing and neutral pictures. Participants were instructed simply to watch the pictures; there were no explicit regulation instructions given. We hypothesized that eyeblink startle magnitude after offset of the stimulus would reflect automatic regulatory components of the emotional response, whereas the initial reactivity component of the response would be indexed by eyeblink magnitude during picture presentation. In keeping with the hypothesis of a prominent role of PFC circuitry in emotion regulation, we predicted that relatively left-sided baseline EEG activation would be associated with decreased responses to unpleasant pictures following, but not during, picture presentation.
METHOD
Participants
Forty-seven individuals (30 females, 17 males; age range: 57–60 years) were recruited from the Wisconsin Longitudinal Study, a long-term study of a random sample of 10,317 men and women who graduated from Wisconsin high schools in 1957 (more information about this sample is available on the Web at http://dpls.dacc.wisc.edu/wls/other_html/aboutwls.htm). All participants provided written informed consent according to the procedures set forth by the University of Wisconsin-Madison Human Subjects Committee. Left-handed individuals, as determined by a 13-item handedness questionnaire (those who endorsed 10 or more of 13 items as being performed primarily with the left hand; Chapman & Chapman, 1987), were excluded from participation.
Procedures
Participants were engaged in a multiday study involving several affect elicitation methods and several psychophysiological and immunological measures. The procedures described here took place on their first day in the laboratory. So that 29 channels of EEG activity could be recorded, participants were fitted with a stretch-lycra electrode cap (Electro-Cap, Inc., Eaton, Ohio) in accordance with the International 10-20 System. Vertical and horizontal electro-oculographic activity was recorded over the supra- and suborbit of one eye, and over the external canthus of each eye, respectively. Participants were also prepared for the collection of eyeblink startle data (two sensors over the suborbit of one eye and one forehead ground lead) and impedance cardiography data (not reported here). After baseline EEG data were recorded, participants were asked to view emotional and affectively neutral pictures on a 17-in. computer monitor. Participants were instructed to watch each picture the entire time it was on the screen, without turning away or closing their eyes. Following the experiment, sensors were removed and participants were orally debriefed.
Stimuli
Participants viewed 51 pictures from the International Affective Picture System (IAPS; Center for the Study of Emotion and Attention, 1995). Seventeen unpleasant (valence M=−1.19, arousal M= 1.17), 17 pleasant (valence M= 1.05, arousal M= 0.98), and 17 neutral (valence M= 0.00, arousal M=−1.56) pictures were chosen on the basis of standardized ratings (Lang, Bradley, & Cuthbert, 1995). 2 Each picture was presented for 6 s, with 14 s between pictures. On each trial, an acoustic startle probe (50-ms, 95-dB burst of white noise) was presented either during the stimulus (2.5 or 4.5 s after picture onset) or after stimulus offset (7 s after picture onset). Presentation of stimuli was counterbalanced with respect to picture valence and probe time.
EEG and eyeblink startle data collection and reduction
Eight 1-min EEG trials (four with eyes open, four with eyes closed) were recorded using an electrode cap (see Tomarken, Davidson, Wheeler, & Doss, 1992, and Davidson, Jackson, & Larson, 2000, for information regarding EEG data collection and reduction). All channels (FPF1, FPF2, FP1, FP2, FZ, F3, F4, F7, F8, FC3, FC4, FC7, FC8, CZ, C3, C4, CP3, CP4, CP5, CP6, T3, T4, T5, T6, PZ, P3, P4, PO3, and PO4) were referenced to physically linked ears to reduce artifact associated with concurrently collected impedance cardiography data. Impedances at each ear sensor were matched to within 0.5 kς (see Dalton & Davidson, 1997, for methodological details). An EEG asymmetry score was calculated for each nonmidline electrode pair by subtracting the log-transformed power density value in the alpha band (8–13 Hz) for the left site from that for the right site, creating 13 laterality scores (e.g., lateral frontal asymmetry = log F8 − log F7). Positive asymmetry scores thus reflect greater left-sided activation (i.e., greater alpha band power density on the right than left). Laterality scores are preferable to data from individual sites as they control for individual anatomical differences such as differences in skull thickness and brain volume (see Tomarken, Davidson, Wheeler, & Kinney, 1992, for details).
Startle eyeblink data were acquired, processed, and standardized following the procedures described in Jackson, Malmstadt, et al. (2000). Integrated and rectified EMG signals were computer-scored and reviewed. Trials were excluded if they contained excessive noise during the 50-ms prestartle baseline epoch, or if the onset of the eyeblink reflex began less than 15 ms after the startle probe. Startle blink magnitudes (in microvolts) were calculated by subtracting the amount of integrated EMG at reflex onset from the maximum peak of integrated EMG between 20 and 120 ms following probe onset. Noise-free trials with no perceptible eyeblink reflex were given a magnitude of zero. Blink magnitudes were z-transformed within subjects to control for large individual differences in response amplitude and baseline EMG levels. A difference score representing emotion-modulated startle was computed at each probe time by subtracting standardized startle magnitude in the neutral-picture condition from that in the unpleasant-picture condition. Each difference score thus represents a negative reactivity score that is unaffected by individual differences in baseline startle magnitude.
RESULTS
Startle eyeblink magnitude
The effects of picture valence on startle eyeblink magnitude were assessed using a 3 (picture valence) × 3 (probe time) analysis of variance. There was a main effect for picture valence, F(2, 28) = 3.48, p= .047. There was no significant main effect for probe time, nor was there a significant Picture Valence × Probe Time interaction. There was a significant linear relationship for picture valence, F(1, 46) = 22.87, p < .001. Eyeblink magnitudes for unpleasant-picture trials (M= 0.17, SD= 0.42) were significantly larger than eyeblink magnitudes for neutral-picture trials (M= 0.00, SD= 0.37), t(46) = 2.64, p= .01, and pleasant-picture trials (M=−0.08, SD= 0.28), t(47) = 4.76, p < .001, replicating several previous studies of emotion-modulated startle (see Lang, 1995). There was a trend for eyeblink magnitudes to be smaller for pleasant-picture trials than for neutral-picture trials, t(49) = 1.97, p= .054.
Eeg-startle relations
Correlations were computed between eyeblink startle difference scores (startle magnitude in the unpleasant-picture condition minus startle magnitude in the neutral-picture condition; i.e., negative reactivity), both during picture presentation (Probes A and B) and after picture offset (Probe C), and each frontal asymmetry score (FPF2 – FPF1, FP2 − FP1, F4 − F3, F8 − F7, FC4 − FC3, FC8 − FC7). Probe A negative-reactivity scores were nonsignificantly correlated with frontal EEG asymmetry (rs ranged from −.09 to .26, ps > .15). Similarly, Probe B negative-reactivity scores were nonsignificantly correlated with frontal EEG asymmetry (rs ranged from −.08 to .20, ps > .27). Significant inverse correlations were found between negative-reactivity scores following picture offset (Probe C) and EEG asymmetry at frontal pole (FP2 − FP1; r=−.41, p= .02) and fronto-central (FC4 − FC3; r=−.36, p= .03) sites (see Fig. 1). The inverse correlations between negative-reactivity scores and frontal asymmetry indicate that increased relative left frontal activation is associated with decreased negative reactivity.
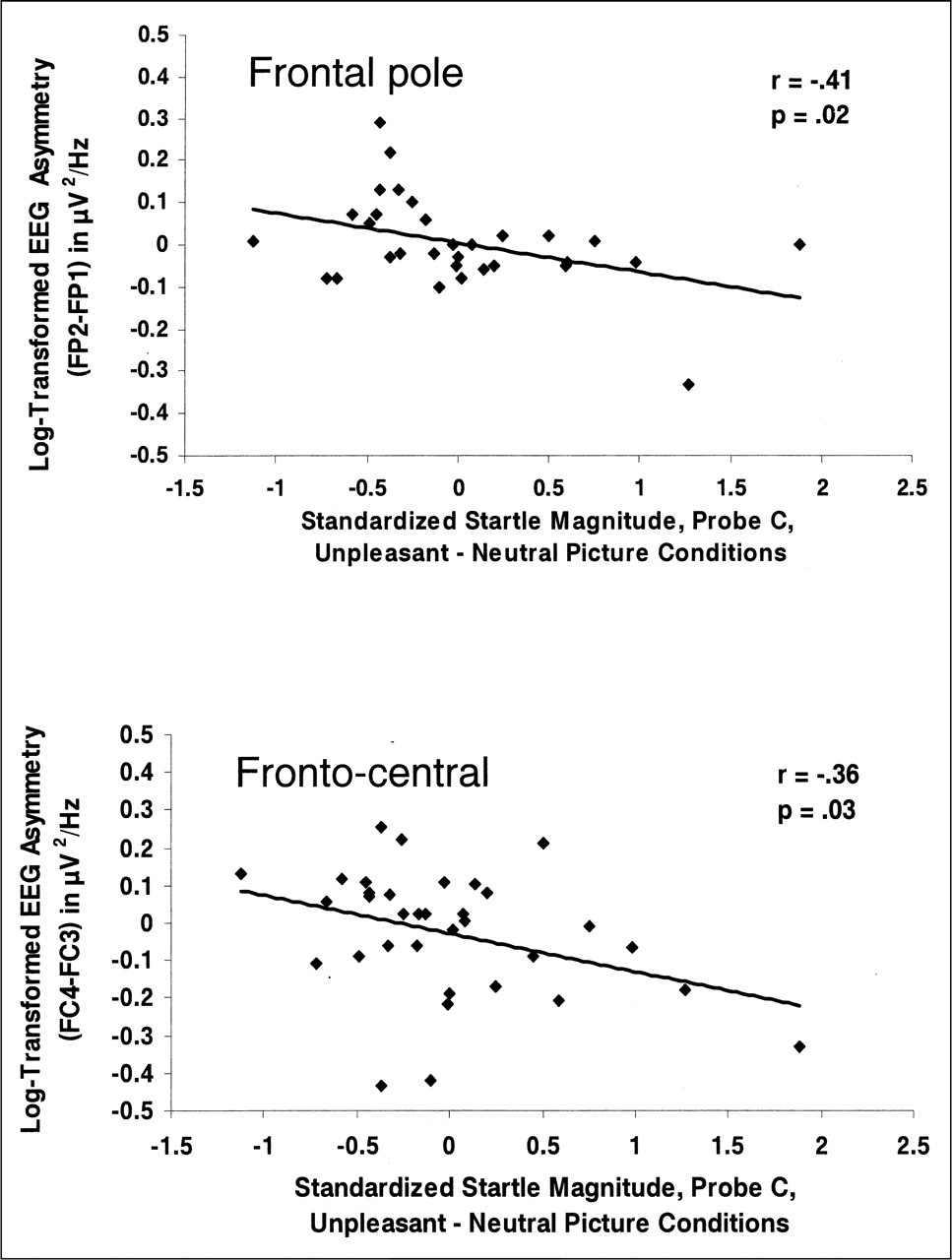
Baseline electroencephalographic (EEG) asymmetry (log right minus log left alpha power) as a function of standardized magnitude of the eyeblink startle to the late probe (Probe C). Results are shown separately for frontal pole (top panel) and fronto-central (bottom panel) sites. Higher numbers on the ordinate are associated with greater relative left-sided activation. Each graph also shows the correlation between EEG asymmetry and startle magnitude. Startle magnitude was calculated by subtracting magnitude in the neutral-picture condition from magnitude in the negative-picture condition.
To assess the relations between negative reactivity and power density at individual sites (as opposed to using the asymmetry metric just reported), we regressed whole-head power from each individual site to remove the effects of individual differences in skull thickness (see Davidson, Jackson, & Larson, 2000, for a description of this method). Correlations between the residual power densities at the individual sites and negative-reactivity scores at each probe time were all nonsignificant. For a regional contrast, we also computed the correlation between parietal asymmetry (P4 − P3) and negative reactivity at Probe C, r= .24, p > .18. This correlation was significantly different from both of the correlations between frontal asymmetry and Probe C negative reactivity reported in the previous paragraph, ts > 2.35, ps < .03.
Negative-reactivity scores at the three probe times were uncorrelated with each other (rs ranged from −.12 to .05, ps > .52).
DISCUSSION
In the current study, we found that automatic, uninstructed emotion regulation, as indexed by eyeblink startle magnitude following unpleasant-stimulus offset, was significantly inversely correlated with relative left-sided resting frontal EEG activation. After picture offset, there were significant inverse correlations between frontal pole and fronto-central resting EEG asymmetry and difference scores for eyeblink startle magnitude (unpleasant-picture condition minus neutral-picture condition). In contrast, these EEG asymmetries did not predict negative reactivity during picture presentation. In addition, these correlations were significantly stronger than the nonsignificant correlations between EEG and startle reactivity at parietal sites.
These findings suggest that baseline frontal EEG is related to duration of negative affect following emotional provocation, rather than to emotional reactivity during stimulus presentation. Although some theorists believe that initial reactivity and subsequent regulation are aspects of the same affective process (e.g., Frijda, 1986), in the current study asymmetry in baseline prefrontal activation predicted only postpicture startle reactivity. This finding is consistent with the notion that the initial reaction to an emotional picture and the lingering response that persists following picture offset are governed by dissociable mechanisms. The results reported here suggest that it is possible, using physiological measures with very fast temporal resolution, to experimentally disentangle emotional reactivity and emotion regulation. Experiments of this type should help in developing clearer constructs of the different types of emotional regulation. Individual variations in the affective parameters described by Davidson (1998) may result from individual differences in initial reactivity, the effects of explicit attempts to regulate emotion, or more automatic regulatory processes; in any case, the final result is to determine in part the quality, intensity, and duration of various emotional episodes. By focusing on a single aspect of affective style, such as duration of an emotional response following the offset of an emotion-eliciting stimulus, researchers may begin to parse the automatic components of emotion regulation from explicit attempts to suppress or enhance an emotional response.
Voluntary and automatic emotion regulation, although empirically separable, are likely to interact both during an emotional experience and over longer periods of time. For example, Davidson (1998) has suggested that those processes that were once voluntary may become automated with increasing use, an idea that echoes the work of James (1890/1981) and that has found empirical support in the literature on skill acquisition (for a review, see Wegner & Bargh, 1998). Indeed, cognitive psychologists have studied extensively the complementary roles of automaticity and consciousness in information processing (e.g., Posner & Snyder, 1975). Automatic and voluntary emotion regulation may share a similar relationship, with voluntary emotion regulation strategies becoming automatic over time (thus allowing increased efficiency and reduced monitoring of responses to emotional challenges), and with automatic regulatory processes taking precedence over voluntary regulatory processes when the individual experiences an extremely intense emotional response.
It is crucial to recognize that biological predispositions underlying emotional reactivity strongly influence the environment within which automatic and voluntary emotion regulation processes develop. In our laboratory, we are currently examining the relations between various forms of automatic and voluntary emotion regulation in order to determine the degree to which individual differences in these processes are predicted by measures of brain activation and initial emotional reactivity. A key aspect of our theoretical approach in this exercise in construct explication is our belief that the most fruitful target of study for emotion regulation researchers interested in parsing automatic and voluntary emotion regulation is the brain. For example, several lines of converging evidence suggest that the PFC and amygdala play an important role in emotional experience and behavior (for a review, see Davidson & Irwin, 1999). The presence of inhibitory projections from orbital and medial regions of PFC to GABA (gamma-amino butyric acid) neurons in the amygdala (Amaral, Price, Pitkanen, & Carmichael, 1992), a subcortical region thought to be associated with negative emotion, supports the notion that PFC plays an important regulatory role in negative affect.
Morgan, Romanski, and LeDoux (1993) found that lesions to medial PFC in rats prolonged the maintenance of a conditioned aversive response (but see Gewirtz, Falls, & Davis, 1997), further implicating PFC as an inhibitor of amygdala activity. In research with humans, Abercrombie et al. (1996) found that for control subjects, but not depressives, glucose metabolism in the left medial and lateral PFC was inversely associated with glucose metabolism bilaterally in the amygdala; several treatment studies have shown that left frontal activation often increases following remission of depressive symptoms (e.g., Baxter et al., 1992; Bench, Frackowiak, & Dolan, 1995). These findings further suggest that the left PFC in particular may act to inhibit the amygdala (see Davidson, Putnam, & Larson, 2000, for further elaboration of this proposal). Inhibition of the amygdala by left PFC may be one of the neural mechanisms underlying both automatic and voluntary emotion regulation. Such inhibition likely occurs both tonically and phasically. Tonic inhibition of the amygdala may be reflected in studies in which subjects with higher left prefrontal activation report more trait positive affect and less trait negative affect, and display greater reactivity to positive stimuli (e.g., Tomarken, Davidson, & Henriques, 1990; Tomarken, Davidson, Wheeler, & Doss, 1992), than individuals with lower levels of left prefrontal activation. In contrast, individuals with high levels of negative affect or depression may have relatively unregulated amygdala activation, in addition to relative right-sided resting PFC activation (for a review, see Davidson, Jackson, & Kalin, 2000).
Of particular interest is the role of prefrontally mediated mechanisms of automatic emotion regulation in resilience to major life stressors. Although the results of the current study were obtained during the relatively rapid presentation of unpleasant stimuli in a carefully controlled laboratory environment, we hypothesize that the neural mechanisms at work in this study may well be the same as those that help to impart resilience to the negative effects of longer-term, idiographic stressors. It is our hope that the current research will help to catalyze research activity examining resilience in the face of affective challenge. Positive affect, well-being, and both automatic and voluntary emotion regulation are all likely determinants of resilience to stressful life events. There is strong evidence suggesting that the left PFC plays a primary role in facilitating positive reactivity, in addition to inhibiting negative reactivity (via subcortical structures), processes that are not mutually exclusive (Davidson & Sutton, 1995). In fact, some data suggest that one function of positive affect may be to “undo” the effects of previous negative affect (Fredrickson & Levenson, 1998), thus suggesting an additional regulatory role for the left PFC. The ability to maintain and enhance positive affect, even to the point of causing cognitive distortion of actual events (e.g., Taylor & Brown, 1988), is undoubtedly an important protective factor against mood and anxiety disorders. It will be important for future studies to examine the regulation of positive emotions, in addition to the voluntary and automatic regulation of negative affect.
SUMMARY
In the current study, we found that baseline frontal pole and fronto-central EEG asymmetry measures in the alpha band were significantly inversely correlated with startle-magnitude difference scores after picture offset, but not during picture presentation. Greater relative left-sided anterior activation at baseline was associated with lower startle magnitude after the offset of negative stimuli, whereas this resting anterior activation was not significantly associated with emotional reactivity during picture viewing. These findings suggest that baseline left prefrontal activation is associated with relatively fast recovery following an aversive event, supporting the idea of a frontally mediated mechanism underlying one parameter of affective style: duration of emotional response following offset of an emotion-eliciting stimulus. This experimental paradigm provides one example of an empirical parsing of the related and interactive constructs of initial emotional reactivity and subsequent automatic regulation of that emotion.
Footnotes
1. We conceptualize as “automatic” those regulatory processes that are spontaneously exhibited in the absence of any specific instructions to regulate emotion. In the current study, participants were not asked if they had attempted to regulate their emotional responses to unpleasant stimuli. Although it is possible that some participants did make such voluntary attempts, such attempts seem far less likely in this case than in paradigms (e.g., Jackson, Malmstadt, Larson, & Davidson, 2000) in which participants are specifically instructed to regulate their emotional responses.
2. Affectively pleasant pictures were presented in order to establish the presence of valence-dependent emotion-modulated startle. Predictions were made only for the association between frontal EEG asymmetry and negative reactivity. Indeed, post hoc analyses revealed no significant correlations between positive reactivity and frontal asymmetry.
Acknowledgements
The research reported in this article was supported by National Institute of Mental Health Grants MH43454, P50-MH52354, and P50-MH61083. D.C.J., J.B.N., and H.L.U. were supported by T32-MH18931 (R.J.D., director). The authors wish to thank Brian Ostafin for his help in collecting and processing the data. A subset of these data was presented at the 41st Annual Meeting of the Society for Psychophysiological Research in Montreal, Quebec, Canada, October 2001.