
Abstract
Some recent evidence has favoured purely response-based implicit representation of sequences in serial reaction time tasks. Three experiments were conducted using serial reaction time tasks featuring four spatial stimuli mapped in categories to two responses. Deviant items from the expected sequence that required the expected response resulted in increased response latencies. The findings demonstrated a stimulus-specific form of representation that operates in the serial reaction time task. No evidence was found to suggest that the stimulus-specific learning was contingent on explicit knowledge of the sequence. Such stimulus-based learning would be congruent with a shortcut within an information-processing framework and, combined with other research findings, suggests that there are multiple loci for learning effects.
Sequencing is a fundamental human ability that is vital in almost every kind of skilled performance. As Fitts and Posner (1968, p .1) pointed out, “Skilled performance always involves an organized sequence of activities.” The sequencing of actions and information has been studied in a variety of different forms, but recently a productive experimental approach has developed from the work of Nissen and Bullemer (1987). Previous reviews of research on sequence learning suggest that one major question is exactly what kind of information do people learn (Clegg, DiGirolamo, & Keele, 1998; Goschke, 1998; Stadler & Roediger, 1998)? The serial reaction task (SRT) developed by Nissen and Bullemer (1987) involves both repeating patterns of stimuli and the corresponding sequences of responses. The benefits seen in reaction time and error rate in such paradigms could be associated with the sequencing of stimuli, the sequencing of responses, or the sequencing of some intermediate representations.
Several approaches have been adopted that can be used to determine what information is being represented in sequence learning (e.g., Buck-Gengler & Healy, 2001; Fendrich, Healy, & Bourne, 1991; Goschke, 1998; Howard, Mutter, & Howard, 1992; Mayr, 1996; Ziessler, 1994, 1998). One of the main techniques involves employing a transfer test, where performance is examined after changes in stimulus or response conditions (Cohen, Ivry, & Keele, 1990; Keele, Jennings, Jones, Caulton, & Cohen, 1995; Willingham, Nissen, & Bullemer, 1989).
Two recent studies using transfer of learning have provided evidence that strongly favoured response-based sequence representation in the SRT. Willingham (1999; Willingham, Wells, Farrell, & Stemwedel, 2000) demonstrated that implicit sequence learning could be based on response information alone. Positive transfer to a new series of stimuli that required the same sequence of responses was found, but no transfer was present when a previous series of stimuli required a new sequence of responses. Although there have been claims of a perceptual basis for sequence learning based on observation of sequences (e.g., Howard et al., 1992), more recent work suggests that such knowledge is probably dependent on explicit learning (Kelly & Burton, 2001; Willingham, 1999; see also Heyes & Foster, 2002).
A Shortcut Mechanism in Stimulus-Specific Learning
Two earlier debates about the locus of similarly elusive effects—repetition effects and practice effects—appear to mirror some of the apparently contradictory empirical findings that have been observed for representation in the SRT. Work in these other domains (e.g., Campbell & Proctor, 1993; Pashler & Baylis, 1991a, 1991b) may offer insight into the problems of determining the representation of sequences. Starting with the commonly held view that human information processing comprises a number of stages (Miller, 1988; Sanders, 1998; Sternberg, 1969), it is possible for learning to circumvent the processing within a stage (or stages) altogether thorough a bypass or shortcut mechanism. An early view of the mechanism for repetition effects was of such a bypass (Bertelson, 1963), with the idea that when a match occurred memory search to retrieve the correct response could be avoided.
It is certainly possible to construct a shortcut explanation of the basis for sequence learning in the SRT. The central notion within this type of account would be that, following learning, a type of stimulus more directly elicits the rapid execution of a response, bypassing intermediate stages of processing. This type of interpretation is compatible with the findings of Willingham et al. (1989) in the earliest attempt to determine the locus of learning in the SRT.
Why then might more recent research have found no stimulus-specific component to learning in SRT paradigms? Pashler and Baylis (1991a) showed that changes in the mapping of stimuli to responses disrupted the advantages of stimulus repetition that was based on a shortcut mechanism. This echoes the manipulations carried out by Willingham (1999) in examining the representation of sequence learning. Indeed transfer tasks by their very nature change the information-processing sequence from a particular stimulus to a particular response, but differ in where in the series of stages that change occurred. When the stages linked together by a shortcut remain intact following transfer then they could continue to offer a basis for stimulus-specific learning.
The current experiments examined whether, using a different type of assessment from a transfer test, evidence of any stimulus-specific information operating during the SRT could be found. Some studies (e.g., Clegg, 1998; Nattkemper & Prinz, 1997; Ziessler & Nattkemper, 2001) using variants of the SRT procedure have introduced deviant stimulus items that are either congruent or incongruent with the expected response in order to determine whether learning was contingent on the response or the stimulus. Instead of having participants respond to single items with individual keys, these experiments used each key to respond to multiple items. The intention of the current experiments was to look for sequence learning specific to, versus independent of, an individual stimulus through the use of deviant items that violated the expected sequential order.
Experiment 1
The first experiment was designed to test whether stimulus-specific learning was present in an SRT paradigm that provided a natural categorical mapping of pairs of stimuli to responses. Longer latencies to deviant items that required the same response as the expected item would indicate stimulus-specific learning.
Method
Stimuli and Apparatus
Stimuli were presented on the monitor of an Apple Macintosh computer using a modified version of the program MacLab (Costin, 1988). The stimuli consisted of a dot, 0.25 in. (0.64 cm) in diameter, and a horizontal line of 3 in. (7.6 cm) in length. The dot would appear in one of four vertically aligned positions, two above and two below the centre of the line, with potential positions separated by 1 in. (2.5 cm). Participants were seated approximately 25 in. (50 cm) from the computer screen.
Responses were made by pressing the “b” and “n” keys of a standard keyboard. The keys were labelled “above” and “below”, and the designation of key to response was counterbalanced across subjects. Participants practised pressing the keys in response to the words “above” or “below” appearing on the computer screen prior to the experiment. In the practice session each word occurred 10 times mixed in random order. Incorrect button presses were indicated with auditory feedback in the form of a beep. Subjects used their first two fingers (index and middle) to respond. The hand used to respond was counterbalanced across subjects, with half responding with their right and half with their left hand.
Procedure
As described above, subjects were first given an opportunity to practise using the keys required in the experiment. Subjects were then given on-screen instructions asking them to judge whether the dot appeared above or below the line and to press the corresponding key. After each response the dot disappeared (but the line remained on display) and was replaced with a new dot 200 ms later. No feedback was given as to either the correctness of the response or the response latency. The instructions asked subjects to respond “as quickly and accurately as possible”, and no mention of a repeating sequence was made. The trials were divided into 16 blocks of 180 responses, and subjects were allowed to take short breaks between blocks if they desired.
Following the end of the final block, subjects filled in a written questionnaire examining their perception of the experiment and in particular their awareness of a sequence. Subjects were asked questions including whether they knew of the presence of a repeating pattern of stimuli and whether they knew of a repeating pattern of responses, and they were encouraged to try to recall as much of the sequence as possible if they believed one existed in either or both cases. After producing written answers, subjects were verbally interrogated about the potential presence of a sequence, and they were encouraged to recall or guess even fragments of the sequence if they suspected one existed. No feedback was given as to the accuracy of recall.
Subjects
A total of 32 University of Oregon undergraduates (16 male and 16 female) participated in the study for either introductory psychology course credit or for payment. There was no overlap of participants between any of the experiments in this paper.
Design
For the two initial blocks of the experiment, the position of the dot followed a pseudorandom order. In the “random” blocks, the order of positions was not predictable, but in other respects the conditions matched the “sequence”. Individual items were associated to all others with a frequency that matched sequence blocks; no repetitions of any stimulus or triple response repetitions occurred; and within each section of 16 trials each of the four positions had to appear exactly four times (with the final 4 trials in each block featuring each item once). Following two random blocks, there were eight blocks in which the position of the dot followed a 12-item ambiguous sequence that repeated continuously. The sequence followed the pattern: 3, 1, 2, 4, 2, 3, 4, 1, 3, 2, 1, 4 (where “1” relates to the topmost potential dot location and “4” the lowest). After the 8 blocks in which the location of the dots followed this regular pattern, the 11th block returned to the random format used in the first 2 blocks. For the 12th block, the location again followed the repeating 12-item sequence. The random versus sequence nature of the first 12 blocks can be summarized as: R1 R2 S3 S4 S5 S6 S7 S8 S9 S10 R11 S12 (where R = random block, and S = sequence block).
After having these 12 blocks as an opportunity to learn the sequence (although not explicitly requested to do so), participants encountered a further 4 blocks that contained displacements of items within the sequence. Displacements always affected the dots closest to the line in the original sequence. Stimuli were displaced an equal distance either within the same category (with 2 becoming 1, or 3 becoming 4) or across to the other category (with 2 becoming 3, and vice versa). Displacements were “within category” (different stimulus/same response) or “between category” (different stimulus/different response). The deviations were only of one item within the sequence of 12, leaving the majority of the rest of the sequence intact. Displacements only occurred twice on every three cycles through the sequence (one same category, one other category). The position in the sequence affected by this switch was determined such that introducing the displacement did not create repetitions of items. Thus, in total, Blocks 13 to 16 contained 20 deviant items moved within the category and 20 deviant items moved between categories.
Measures
The median reaction time for correct responses was calculated for each subject in each block. For each type of element in the final four blocks (regular items and both classes of deviant items), the mean of the medians from each of the four blocks was calculated for each participant. Subjects were classified in terms of awareness of the sequence on the basis of the questionnaire responses that were collected at the end of the experiment (see below).
Results
The means of the median reaction times for block (1–12) revealed a significant effect of practice, F(11, 341) = 32.88, p < .05, MSE = 1,054. However, this includes general improvement at the task as well as sequence specific learning. General changes are not critical within these studies, and therefore although statistically significant these analyses are omitted from the reporting of subsequent experiments.
Further investigation looked in more detail at sequence learning by comparing performance in the random block (R11) to the average of the two surrounding sequence blocks (S10 and S12). There was a significant effect of sequence presence, t(31) = 2.67, p < .05 (see left panel of Figure 1), with faster mean reaction time for the sequence blocks (316 ms) than for the random block (329 ms). Thus learning specific to the sequence had occurred.
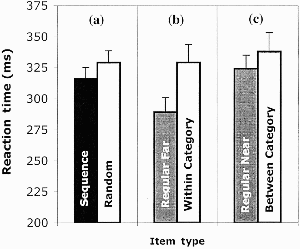
Experiment 1: Reaction time by item type for above/below judgements. (a) Sequence versus random; (b) within-category displacements versus regular near items; (c) between-category displacements versus regular far items (see text for details). Performance shown reflects mean of block median reaction times for correct responses, and error bars show standard errors.
Having shown that sequence learning occurred under the category response conditions, the most crucial analyses were conducted to examine the effects of displacing expected items in the final 4 blocks (Blocks 13 to 16). Looking only at the regular items within undisrupted runs through the sequence in these blocks, and breaking them down into “near” and “far” items, produced a significant effect of distance, t(31) = 6.17, p < .05. Far items (the top and bottom positions) were classified as “above” or “below” faster than those items near to the line (287 ms vs. 324 ms). Within-category displacements (different stimulus/same response) always moved items expected near to the line to the location far from the line. On the other hand, displacements between categories (different stimulus/different response) always produced a near item. This was done so all displacements involved equal distances and did not span other intervening positions (no displacements of expected far items were included because distance moved for within-versus between-category changes would then be different).
Given the distance effects, to answer the question of whether stimulus-specific learning had occurred a comparison of within category displacements to regular sequence items equivalent in distance from the line was conducted. These revealed a significant difference, t(31) = −4.83, p < .05 (see middle panel of Figure 1), with regular far from the line items showing significantly shorter latencies than far items produced by the within-category displacements. The effect of between-category displacements compared to regular near items was only of marginal significance, t(31) = − 1.94, p = .06 (see right panel of Figure 1).
Representation and Awareness
Learning in the absence of awareness is an issue that has drawn much discussion within the sequence-learning literature (e.g., Perruchet & Amorim, 1992; Shanks & Johnstone, 1998; Shanks & St. John, 1994; St. John & Shanks, 1997). In evaluating the representation of sequence learning, one major issue concerns whether the nature of the representation is contingent on awareness (see, for example, Willingham, 1999). Because the type of representation could conceivably differ, at this stage it seems prudent to attempt to assess whether representational codes vary for participants with and without awareness, whether or not the distinction ultimately proves itself to have utility.
Of the 32 subjects, 13 reported no knowledge of the sequence in terms of either locations (Positions 1, 2, 3, and 4) or response categories (“above” and “below”). These participants were considered less aware of a sequence (mean length of sequence recalled out of 12 items = 0.0). Of those showing at least some knowledge of the sequence, 11 participants gave three or four adjacent items correctly (mean length of sequence recalled = 3.7), and 8 participants were able to provide five or more contiguous items of the sequence (mean length of sequence recalled = 7.0). Although the participants producing the shorter correct series may have demonstrated some potential knowledge of the presence of a sequence, it was possible they simply guessed which elements followed consecutively. Any of the six possible combinations of three element sequences of above and below items (excluding triple repetitions of the same item that did not feature in the experiment) are present somewhere within the 12-item response sequence. Hence participants responding to the free recall of the sequence by guessing at least three items would generate correct sequences at least this long.
Further analyses were conducted to assess whether the findings of stimulus-specific learning found above were contingent on explicit knowledge of the sequence. An analysis of within-category deviant items using just the less aware participants showed the same pattern of results as the overall data, t(12) = − 2.97, p < .05 (regular far items = 309 ms vs. within-category displacements = 348 ms).
Finally, a correlation was conducted to explore whether there was any evidence of a relationship between the length of the sequence that participants could recall and the size of costs associated with within-category deviant items (on a score derived from within-category displacements minus regular far items). This analysis also produced no evidence to support the idea that stimulus-specific representation might be contingent on explicit knowledge (r = −.05, p > .05).
Discussion
The central question in the first experiment was whether a stimulus-based representation operates within the SRT paradigm. The results show such a representation was indeed involved since a change from the expected stimulus, even when not accompanied by a change of response, increased reaction time.
The stimulus-based learning in these data mirror Mayr's (1996) results that found spatial learning to be independent of the key response required. In this case, however, the distances between locations were far less extreme. The cost of a change of location observed was 28 ms for a visual angle between stimuli positions within a category of approximately 2.5°. This result compares to about a 62 ms advantage observed by Mayr (1996, Exp. 2) for stimuli with a visual angle of 27° between them.
The analyses do not rule out the possibility that awareness can change performance (see, e.g., Curran & Keele, 1993), especially given that some relatively small cell sizes were present and that there is no universal agreement on how to determine whether knowledge is truly implicit. Nonetheless, the positive results for just the less aware participants offer no strong indication that the type of representation operating critically depended on conscious knowledge in these circumstances.
One reservation in the interpretation of the results might be that some of the procedure in using a line on the screen to create categories for the judgements changed the task somewhat from that commonly employed in the SRT paradigm. Such a concern became the focus of a second experiment.
Experiment 2
Experiment 1 employed a vertical alignment of stimuli positions; however, the standard arrangement in SRT research has been to use a series of horizontal positions. Additionally the presence of the line on the screen in the earlier experiments might have biased a visual search to preferentially examine locations on one side of the line first, uniquely giving the appearance of a within-category advantage for above/below judgements. To address these issues a horizontal arrangement of stimuli was employed, requiring a new type of judgement.
Method
Stimuli and Apparatus
Stimuli and apparatus were similar to those used in Experiment 1 but with the four positions laid out horizontally rather than vertically. Each of the four potential locations was indicated by a square box, 0.75 in. (1.9 cm) wide, with each box 0.25 in. (0.64 cm) from its neighbour(s). A dot (0.25 in. in diameter) appeared in one of the four locations until a response was made, and following a 200-ms delay the next dot would appear in the new location. Responses were again made by pressing the “b” and “n” keys; however, in this experiment the keys were labelled with stickers as “left” and “right”, respectively. Stimuli were presented on the monitor of an Apple Macintosh computer using PsyScope software (Cohen, MacWhinney, Flatt, & Provost, 1993).
Procedure
The procedure was similar to that in Experiment 1. While the structure and nature of the blocks remained the same, the length of the blocks was shortened to 120 trials (from 180). The same sequence was employed with sequence positions left to right corresponding to the top to bottom arrangement present previously. All participants responded with their dominant hand. The other change was that during the instruction phase subjects were shown the four possible positions for the dot and were told to classify a dot falling in either of the two possible leftmost positions as “left” and in either of the two rightmost positions as “right”. Unlike in Experiment 1, no on-screen line was used to delineate the two categories.
Subjects
A total of 38 Colorado State University undergraduates, 15 males and 23 females, participated in the study for introductory psychology course credit.
Results
As in Experiment 1, sequence learning was investigated by comparing performance in the random block (R11) to the average performance in the two surrounding sequence blocks (S10 and S12). There was a significant effect of sequence presence, t(37) = 4.13, p < .05 (see left panel of Figure 2), with shorter response latencies in the sequence blocks (368 ms) than in the random block (407 ms).
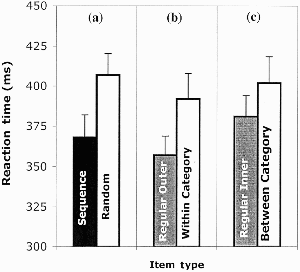
Experiment 2: Reaction time by item type for left/right judgements. (a) Sequence versus random; (b) within-category displacements versus regular near items; (c) between-category displacements versus regular far items (see text for details). Performance shown reflects mean of block median reaction times for correct responses, and error bars show standard errors.
The critical analyses once again concerned the effects of displacing expected items in the final 4 blocks (Blocks 13 to 16). A significant effect of distance was present, t(37) = 3.93, p < .05. Outside items (the leftmost and rightmost positions) were classified as “left” or “right” faster than those items near to the centre (357 ms vs. 381 ms). Stimulus-specific learning was therefore assessed through a comparison of within-category displacements to regular sequence items equivalent in distance from the line. These revealed a significant difference, t(37) = −3.76, p < .05 (see middle panel of Figure 2) with regular outer items showing significantly shorter latencies than outer items produced by the within-category displacements. The effect of between-category displacements compared to regular inner items was also statistically significant, t(37) = −2.03, p = .05 (see right panel of Figure 2).
Representation and Awareness
Of the 38 subjects, 17 reported no awareness of the presence of a sequence (mean length of sequence recalled of the 12 items = 0.0), 8 participants were able to report or guess only 3 or 4 items from the sequence (mean number recalled = 3.5), and 13 were aware of the sequence and able to recall 5 or more items (mean recall = 6.5 items).
To examine whether stimulus-specific learning was contingent on explicit knowledge, a separate analysis was conducted that was confined to just the less aware participants. As in the overall data, there was a significant cost associated with a change of stimulus, t(16) = −3.85, p < .05, with responses to regular outer item positions (359 ms) showing significantly shorter latencies than those to outer items produced by the within-category displacements (406 ms).
A correlation was conducted in order to examine the relationship between the length of the sequence that participants recalled and the size of costs associated with within-category deviant items (within-category displacements minus regular far items). This analysis produced no evidence to support the idea that stimulus-specific representation was related to explicit knowledge (r = −.09, p > .05).
Discussion
These findings match with those from Experiment 1 and suggest a stimulus-specific component to learning because a change from the expected stimulus, even when not accompanied by a change of response, increased reaction time. Beyond replicating the results of Experiment 1, the modified stimulus layout also rules out several potential explanations that might relate to the paradigm changes used in the earlier experiment. Stimulus-based learning does not depend on the presence of the line to delineate the categories; it can be found both with and without markers for potential stimulus-positions available; nor does it result from a contrast between horizontal and vertical alignments. Moreover, the same pattern of results was found once again for only those participants who subsequently demonstrated no major awareness of the sequence.
Experiment 3
One notion that has been proposed is that task properties such as the stimulus–response mapping may be critical to the nature of performance in SRT paradigms (Clegg, 1998; Hoffman, Sebald, & Stöcker, 2001; see also Grafton, Salidis, & Willingham, 2001, for an illustration of this in another form of implicit learning). For example, more difficult mappings of stimuli to responses stress response-related elements of information processing and might tend to focus learning on that type of information. Thus an alternative account of the absence of stimulus-specific learning in the experiments of Willingham and colleagues (Willingham, 1999; Willingham, Wells, Farrell, & Stemwedel, 2000) would be that response-based representations develop under more incompatible response formats.
The same stimuli and responses as those employed in Experiment 2 were utilized but with a different mapping of keys to locations. Identical “left” versus “right” judgements to those in Experiment 2 were required, but with responses made using incompatible key response formats (e.g., “left” responses made with the right key of the pair).
Method
Stimuli and Apparatus
Stimuli and apparatus were similar to those used in Experiment 2. Responses were again made by pressing the “b” and “n” keys; however, in this experiment the keys were labelled with stickers as “right” and “left”, respectively, producing an incompatible mapping of the stimuli to the responses. Items appearing on the left side of the screen required a response with the right key, whereas items appearing on the right side of the screen required a response with the left key.
Procedure
The procedure was identical to that in Experiment 2.
Subjects
A total of 34 Colorado State University undergraduates (10 males and 24 females) participated in the study for introductory psychology course credit.
Results
A total of 2 participants who made 45% or greater errors in the final four blocks were excluded from the analyses for inattentive performance.
The first analysis once again confirmed sequence learning by comparing performance in the random block (R11) to the average performance in the two surrounding sequence blocks (S10 and S12) There was a significant effect of sequence presence, t(31) = 5.30, p < .05 (see left panel of Figure 3), with shorter response latencies within the sequence blocks (429 ms) than within the random block (490 ms).
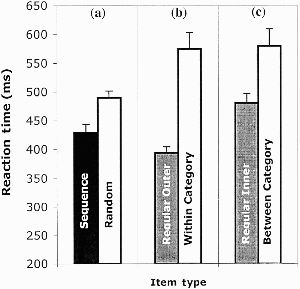
Experiment 3: Reaction time by item type for left/right judgements with incompatible responses. (a) Sequence versus random; (b) within-category displacements versus regular near items; (c) between-category displacements versus regular far items (see text for details). Performance shown reflects mean of block median reaction times for correct responses, and error bars show standard errors.
The central analyses were again focused on the impact of displacing expected items. As in both previous experiments, a significant effect of distance was found, t(31) = 7.72, p < .05. Outside items (the leftmost and rightmost positions) were classified as “left” or “right” faster than those items near to the centre (394 ms vs. 481 ms). Stimulus-specific learning was therefore assessed through a comparison of within-category displacements to regular sequence items equivalent in distance from the line. These revealed a significant difference, t(31) = −6.60, p < .05 (see middle panel of Figure 3) with regular outer from the line items showing shorter latencies than outer items produced by the within-category displacements. The effect of between-category displacements compared to regular inner items also produced a significant difference, t(31) = −4.48, p < .05 (see right panel of Figure 3).
Representation and Awareness
Of the 32 subjects included in the analyses, 12 reported no awareness of the presence of a sequence (mean length of sequence recalled of the 12 items = 0.0), 6 were able to report or guess only 3 or 4 items from the sequence (mean number recalled = 3.8), and 14 were aware of the sequence and able to recall 5 or more items (mean recall = 7.1 items).
To examine whether stimulus-specific learning was contingent on explicit knowledge, a separate analysis was conducted that was confined to just the least aware participants. As in the overall data, there was a significant cost associated with a change of stimulus, t(11) = −4.12, p < .05, with regular outer items (400 ms) showing significantly shorter latencies than outer items produced by the within-category displacements (625 ms).
A correlation conducted on the length of the sequence that participants recalled and the size of costs associated with within-category deviant items produced no evidence to support the idea that stimulus-specific representation was contingent on explicit knowledge (r = −.25, p > .05).
Discussion
Participants displayed performance to the deviant stimuli moving within the category consistent with the operation of a stimulus-based representation. As in both the previous experiments when unexpected items occurred that required the expected response an increase in latency was observed. This would contradict an explanation of stimulus-specific learning purely in terms of the stimulus–response mappings in the task (see Clegg, 1998; Hoffmann, Sebald, & Stöcker, 2001). The type of representation does not seem to be determined strictly on the basis of task characteristics such as stimulus–response mapping, and thus the absence of stimulus-based learning from the studies conducted by Willingham (Willingham, 1999; Willingham et al., 2000) cannot be accounted for by any differences on that dimension between those experiments and the previous ones presented in this paper.
General Discussion
The results from all three experiments clearly converge on stimulus-based representation operating within the SRT paradigm. All the findings showed that deviant items to the previously learned sequence that required the same response as the expected item resulted in increased reaction times. Although there is no way to rule out the possibility that some awareness exists (see Merikle, 1994), and therefore explicit knowledge could have been the basis for stimulus-based learning, the current experiments offer no data to support such an idea.
Multiple Mechanisms for Learning
The stimulus-specific learning effects found offer an extension to findings from transfer tests (Willingham, 1999; Willingham et al., 2000). While work by Willingham and colleagues would be congruent with the idea that there is only a stimulus-independent dimension to implicit learning in the SRT, these new data implicate at least one other potential mechanism for learning. Overall then the pattern of results across various studies would be consistent with the idea that the representation is not always composed of information entirely of one type (see also Goschke, 1998). The idea that multiple pathways for speed-up might exist mirrors the findings for repetition and practice effects (Campbell & Proctor, 1993; Pashler & Baylis, 1991a, 1991b).
The results do not address the question of whether different representations of learning occur within a single system or are associated with separate systems. A great deal of research has attempted to evaluate whether multiple systems exist and if so to characterize their nature (Keele, Ivry, Mayr, Hazletine, & Heuer, 2003). Evidence of apparent parallel learning of stimulus- and response-based sequences (e.g., Goschke, 1998; Mayr, 1996) certainly seems to implicate separate systems but, as work in the field of memory has shown, separating system changes from process changes is remarkably tricky (see, e.g., Parkin, 1999).
A Shortcut Account of Stimulus-Based Learning
The presence of stimulus-specific learning in the current experiments, while it was apparently absent from other studies, would be consistent with the notion that stimulus-based learning takes the form of a shortcut in information processing. Such an explanation can account for the otherwise contradictory findings from previous research using transfer tests without a need to suggest that contamination of implicit performance by explicit knowledge underlies the difference (Willingham et al., 2000).
When specific stimuli are mapped to an entirely different category of responses or when the stimuli themselves are changed then any learning based on a shortcut mechanism (unless that shortcut led directly from one stimulus to the next stimulus) would be disrupted. Using these types of manipulation in transfer tests (for example, changing the mapping of spatial stimuli to responses, or switching stimuli from letters to spatial locations), Willingham (1999) observed only stimulus-independent learning. In contrast, Keele and colleagues (Keele et al., 1995) changed response execution from a manual to a vocal format. In this manipulation, intermediate information-processing stages from the specific stimulus through to selection of the response category would have been preserved. Under these circumstances some transfer of learning occurred.
Where in the series of information-processing stages might the locus of the stimulus-based advantage lie? In the context of practice effects, Pashler and Baylis (1991b) pointed out that when using familiar types of stimuli and simple motor responses, processes such as perception or the motor execution have comparatively little scope for speed-up. In contrast the links between the stimulus identity and the required response (response selection) are novel to the experimental situation, and thus learning associated with these stages offers larger potential benefits for performance.
The partial transfer found by Keele (Keele et al., 1995) might suggest that, as with repetition effects (Pashler & Baylis, 1991a), shortcuts through the response selection stages link to both the general category of the response (e.g., “left”) and the specific location of the response key (e.g., the left key). The absence of effector-specific learning (Cohen et al., 1990; Grafton, Hazeltine, & Ivry, 1998, 2002; Keele et al., 1995) suggests that any shortcut does not link to the specific motor action required (e.g., move the index finger of the right hand), although this may change when extended practice occurs (Verwey & Clegg, in press).
Is Spatial Learning Based on a Response Sequence?
Nattkemper and Prinz (1997) in a study using deviant items produced no evidence of stimulus-based representation of sequences. The contrast of the current findings with those from Nattkemper and Prinz who did not employ spatial stimuli, but paired letters with keys, raises the question of whether stimulus-based learning is centred upon learning of eye movements to the stimulus locations and hence is actually a response-based system. Robertson, Tormos, Maeda, and Pascual-Leone (2001) suggest there are different neural systems involved for learning spatial sequences. In addition, there is also at least some evidence showing that implicit learning of eye movements can occur. Using a manual tracking task with a repeating pattern, Grafton, Salidis, and Willingham (2001) demonstrated that some brain activation changes for the repeating pattern could be associated to oculomotor areas. Whether or not such brain systems underlie learning of the spatial locations in the SRT task remains a question for future research. Mayr's (1996) findings of spatial learning, independent of the response have been interpreted as being based upon orienting from either eye movements or attention (see also Stadler, 1989, for a similar finding in another form of implicit learning, and Lambert, 2003, for some discussion of implicit learning and visual orienting). In Mayr's experiments the large distances between the locations employed may encourage such an explanation; however, in these experiments the stimuli occurred relatively short distances apart. Moreover, if learning were purely based upon eye movements, why would these provide an advantage in the current experiments using spatial stimuli but not others (Clegg, 1998; Willingham, 1999; Willingham et al., 2000)?
Once again work on repetition effects might shed light on this issue of why work such as Nattkemper and Prinz's (1997) with deviant items showed no stimulus-based effects. Campbell and Proctor (1993) demonstrated the potential importance of whether or not stimulus and response sets could be categorized. In the current experiments, the spatial stimuli mapped naturally to existing categories (above and below, left and right). On the other hand, Nattkemper and Prinz (1997) paired letters to responses in an arbitrary fashion (see also Ziessler, 1994). For example, they assigned the letters “V” & “H” together to a response made with the middle finger key on the left hand. Thus the presence or absence of the stimulus-based learning might not be driven by the spatial nature of stimuli but rather by whether stimulus sets reflect categories or not. As discussed above, some authors have proposed that the nature of representation in the SRT might change with task factors (Clegg, 1998; Hoffmann et al., 2001). While the current experiments suggest that stimulus–response compatibility alone does not appear to change the locus of learning, it may be that aspects of the task such as whether stimulus and response sets exist could influence the type of representation.