
Abstract
When participants perform a sequence of different tasks, it is assumed that the engagement in one task leads to the inhibition of the previous task. This inhibition persists and impairs performance when participants switch back to this (still inhibited) task after only one intermediate trial. Previous task-switching studies on this issue have defined different tasks at the level of stimulus categorization. In our experiments we used different response modalities to define tasks. Participants always used the same stimulus categorization (e.g., categorize a digit as odd vs. even), but had to give a vocal, finger, or foot response (A, B, or C). Our results showed a higher reaction time and error rate in ABA sequences than in CBA sequences, indicating n − 2 repetition cost as a marker for persisting task inhibition. We assume that different response modalities can define a task and are inhibited in a “task switch” in the same way as stimulus categories are inhibited.
For humans to work efficiently, it is important to act according to the task that is relevant at the moment and to suppress all task-irrelevant information, competing intentions, and so on. The task-switching paradigm provides a possible way to examine processes of task inhibition (Monsell, 2003). This paradigm is widely used in experimental psychology because it is considered to be able to study the flexible adaptation to changing requirements and to provide insight into those cognitive control processes responsible for such adaptive behaviour.
One way to study task inhibition is the use of task-switching experiments with three tasks (Mayr & Keele, 2000; but see also Masson, Bub, Woodward, & Chan, 2003). The performance in a task is compared under two conditions: In an n − 2 task repetition, participants perform the same task as they have performed two trials before (i.e., task sequence ABA). In an n − 2 task switch, the task in trial n was not performed in any of the two preceding trials (i.e., task sequence CBA). It has been found that reaction time (RT) and error rate are increased in an n − 2 task repetition as compared to an n − 2 task switch (see, e.g., Arbuthnott & Frank, 2000; Mayr & Keele, 2000). To explain this effect, it is assumed that the engagement in a new task leads to the inhibition of the preceding task. The inhibition persists and impairs performance when switching back to this task, as in a task sequence ABA.
The first study in this context was the study of Mayr and Keele (2000). The authors found an n − 2 repetition cost (which they called “backward-inhibition” effect) by using an odd-item-out paradigm. Participants had to locate a deviant object (one out of four) according to different stimulus characteristics (colour, orientation, or movement). Other studies, taking up the experimental logic suggested by Mayr and Keele, also used different stimulus categorizations to define tasks. For example, in the study of Arbuthnott and Frank (2000), participants had to decide whether a letter was a vowel or a consonant, whether a digit was odd or even, or whether a sign was used primarily in text or mathematical context. Similar categorization tasks were used in studies by Arbuthnott and Woodward (2002) and by Hübner, Dreisbach, Haider, and Kluwe (2003). In the studies by Mayr (2001) and by Mayr and Kliegl (2003) participants had to decide about the form, the size, and the colour of a stimulus. Schuch and Koch (2003) used three different numerical judgements, and Dreher and Berman (2002) used three lexical decision tasks. To summarize, from all of theses studies it can be assumed that an n − 2 repetition cost occurs when participants switch back to a stimulus categorization that they have performed two trials before.
However, a study by Koch, Gade, and Philipp (2004) suggested that the n − 2 repetition cost occurs not only with different stimulus categorizations, but also with different manual response modes. In this study, participants switched among two choice-response tasks (numerical decisions; tasks A and B) and one simple-response task (unconditional double-press; task D; please note that the placeholder “D” is used instead of “C” as a abbreviation of “double-press”). The results showed that the performance in the simple-response task was worse when it had been performed in trial n − 2 (trial sequence DAD or DBD) than when it was after two choice-response tasks (trial sequence BAD or ABD). This data pattern suggested that an n − 2 repetition cost could be also found for different manual response modes (simple vs. choice response). “Task” inhibition, thus, also affects the mode of responding.
The aim of the present study was to extend the notion of n − 2 repetition cost even further. We were interested whether an n − 2 repetition cost can be demonstrated on the level of response modalities. In our daily life, flexibility is not only required and restricted to abstract “stimulus categorizations”. In contrast, many situations lead to a switch between different response modalities(e.g., talking and typing, or braking and changing gear). Yet, the modality in which a response is executed has been largely neglected so far in most task-switching studies. For example, in the study of Koch et al. (2004) the response mode (i.e., choice response vs. simple response) was manipulated but all responses were executed by key presses with the right or left index finger. Thus, the question arises whether the requirement to execute responses with different modalities (such as manual vs. vocal) may also lead to the finding of an n − 2 repetition cost. This question is important because the occurrence of a similar empirical phenomenon (e.g., the n − 2 repetition cost) could indicate that a task in task switching cannot only be defined by stimulus categorizations but also by other components that are relevant in a task set. The term “task set” (cf. Rogers & Monsell, 1995) is used to refer to all processes that are necessary to perform a specific task.
In this context, it can be discussed whether a task set is a unitary concept (see, e.g., Gilbert & Shallice, 2002) or whether a task set consists of different components (see, e.g., Kleinsorge & Heuer, 1999; Logan & Gordon, 2001; Meiran, 2000). As regards this question, the finding of the same empirical marker when switching among stimulus categorizations and when switching among response modalities favours the existence of different task set components. Further, such finding can answer whether switching among stimulus categorizations and switching among response modalities lead to similar cognitive “task representations” or whether “task representations” differ depending on the component that is switched in an experiment.
In the present study we were interested in whether response modalities have a functionally similar cognitive representation as stimulus categorizations. If this is true, we should find similar effects when switching among stimulus categorizations and when switching among response modalities. We expected to find an n − 2 repetition cost when switching among different response modalities. To test this expectation, we had participants perform the same numerical judgement in each trial while switching between vocal, finger, and foot responses. Finding n − 2 repetition cost on the level of response modalities would constitute an entirely new phenomenon related to the inhibition of response modalities, which would help specify the components of cognitive task representations. Specifically, it would indicate that the response modality, which has been largely neglected so far, plays a role in the cognitive representation of a task.
With our response modality-switching paradigm, we explored whether an n − 2 modality repetition cost follows the same principles as n − 2 repetition cost in the context of switching stimulus categorizations. We examined this hypothesized analogy with respect to two variables: preparation time and “decay” time.
Previous studies on n − 2 repetition cost used a task-cueing paradigm (Meiran, 1996). In this paradigm, a cue precedes an imperative stimulus to indicate the upcoming task. It has been found that prolonging the cueing interval, which can be used for task preparation, did not influence the size of the n − 2 repetition cost (see Hübner et al., 2003; Mayr & Keele, 2000; Schuch & Koch, 2003). It should be noted, though, that longer cueing intervals were associated with generally improved performance. In the present study, we tested whether we find the same data pattern with respect to response modalities.
With regard to decay time, it was found that an increased time interval between each trial reduced the n − 2 repetition cost (see Gade & Koch, in press; Mayr & Keele, 2000). In contrast to “active” preparation, an effect of a “passive” decay is assumed here (see Koch et al., 2004, for discussion). We tested whether this data pattern would show up also with respect to an n − 2 repetition of response modalities.
Additionally, we manipulated the type of task cue. For half of the participants, the modality of the next trial was cued by “direct” symbols of mouth, hand, and feet for vocal, finger, and foot responses (“direct-cue group”), whereas for the other half, the cues were abstract (square, triangle, and diamond; “abstract-cue group”). Previous studies, in which participants switched among stimulus categorizations, mostly used abstract cues. Because response modality switching has not been examined so far, we could not know how difficult it is to switch between response modalities. Therefore, we decided to use direct cues next to abstract cues. We assumed that the overall RT and error rate would be higher in the abstract-cue group, because participants have to translate the abstract cue into the modality (i.e., more difficult cue interpretation and retrieval of the modality). Additionally, a long preparation time might be more helpful with abstract cues because it allows for cue interpretation and modality retrieval to a larger extent (cf. Mayr & Kliegl, 2000, Exp. 3). Since both processes are mainly necessary with abstract cues, the preparatory-based performance improvement might be larger in the abstract-cue group. However, we had no theoretical reason to expect any influence of the type of cue on the n − 2 repetition cost. Finding no influence on the n − 2 repetition cost would support the idea that this inhibitory effect is not triggered by cue interpretation or “task” (i.e., modality) retrieval but by the selection of the correct response (see Schuch & Koch, 2003).
To increase the generality of the findings, we also varied the stimulus categorization (i.e., numerical judgement). That is, half of the participants performed a magnitude judgement (smaller/larger than 5), and the other half a parity judgement (odd/even). However, we did not expect any systematical effects of the numerical judgement on the n − 2 modality repetition cost.
In summary, in two experiments we examined the n − 2 repetition cost on the level of response modalities (i.e., vocal, finger, and foot responses). In Experiment 1, we investigated the n − 2 repetition cost and its dependency on the duration of preparation time. In Experiment 2, n − 2 repetition cost was analysed as a function of the time between the response and the next cue (response–cue interval, RCI, “decay” time).
Experiment 1
In Experiment 1, participants had to switch among three different response modalities. We expected to find an n − 2 modality repetition cost when comparing a modality sequence ABA (e.g., finger/foot/finger) and a modality sequence CBA (e.g., vocal/foot/finger). This finding would establish the basic response modality inhibition effect.
We included a manipulation of the cue–stimulus interval (CSI; 100 ms vs. 1,000 ms) to examine the influence of the preparation time on the n − 2 modality repetition cost. Additionally, we used two different types of cue (direct vs. abstract) and two different numerical judgements (magnitude vs. parity).
Method
Participants
A total of 32 participants (26 female and 6 male, mean age = 23.3 years) were evenly assigned to the four experimental groups (direct cues/magnitude, direct cues/parity, abstract cues/magnitude, and abstract cues/parity). They received 7 Euros for participation.
Stimuli and Tasks
Stimuli consisted of the digits 1–9, excluding 5. Participants had to decide whether a digit was smaller or larger than 5 (magnitude judgement), or whether it was odd or even (parity judgement). Each participant had to perform only one numerical judgement throughout the experiment.
Stimuli were presented one at a time in white in a frame at the centre of a black screen (15″ monitor) connected to an IBM-compatible PC. The digits were 1 cm high and approximately 0.5 cm wide. The viewing distance was 60 cm. For the direct-cue group, the frame was white and had the size of a square (width/height of 3.8 cm). Which modality to use was indicated by two pictures of a mouth (2.5 cm by 1.5 cm), a right and left hand (3.0 cm by 3.0 cm), or a right and left footprint (2.0 cm by 3.0 cm) shown on each side of the frame. In the abstract-cue group, the frame, which served as modality cue, was white and had the shape of a diamond (5.3 cm wide/high), indicating vocal responses, of a triangle (4.5 cm wide/4.0 cm high), indicating finger responses, or of a square (3.8 cm wide/high), indicating foot responses.
Vocal responses were given by saying “right” or “left”. Speech onset was recorded using a voice-key; “right” and “left” responses were coded by the experimenter with the right and left cursor keys. Finger responses were performed on an external keyboard with two response keys for the right and left index fingers. Response keys measured 1.2 cm by 1.7 cm and were separated by 3.8 cm. Foot responses were made on a separate external keyboard with two response keys (6.0 cm by 6.0 cm, separated by 23.5 cm) for the right foot and the left foot.
Procedure
The experiment was run in a single session of approximately 50 minutes. Instructions were both given on the monitor and orally. Instructions emphasized speed as well as accuracy. Participants were informed about the numerical judgement and the different response modalities. An instruction sheet concerning the stimulus category–response category mapping (e.g., odd–left) was placed in front of each participant throughout the experiment. The two possible mappings for each judgement were counterbalanced across participants.
Two practice blocks were run with 12 trials each. One practice block had a short CSI (100 ms); the other had a long CSI (1,000 ms). The experiment itself consisted of eight blocks of 96 trials each. Before each block, participants were informed about the CSI in the next block. Blocks with short and long CSI alternated within one experiment; the CSI duration in the first block was counterbalanced across participants.
A trial started with a black screen followed by a cue. After a variable preparation time (CSI), the imperative stimulus was presented in the middle of the cue frame. The interval between the response of the participant and the next imperative stimulus (response–stimulus interval, RSI) was 2.1 s after finger and foot responses but variable around 2 s after vocal responses. As the experimenter had to code vocal responses, the time between the response of a participant and the next cue (RCI) depended on the RT of the experimenter. We used an average of 300 ms to prolong the RCI in nonvocal trials. Thus, in finger and foot trials RSI was held constant at 2,100 ms with RCI being either 2,000 ms or 1,100 ms, and CSI being either 100 ms or 1,000 ms (i.e., RCI 2,000/CSI 100 vs. RCI 1,100/CSI 1,000). Vocal trials had approximately the same RCI and RSI.
The sequence of trials was controlled for an equal number of each response modality, stimulus category (odd vs. even, or smaller vs. larger), and modality sequence (n − 2 repetition vs. n − 2 switch). Immediate repetition of a response modality was excluded, because “task” repetitions were found to reduce the size of n − 2 repetition cost (Philipp & Koch, in press). Additionally, the immediate repetition of an imperative stimulus was excluded as well as immediate repetition of an imperative stimulus within one category.
Participants always received visual error feedback for 500 ms when they pressed the wrong key (German: “Falsche Taste”).
Design
Modality sequence (n − 2 repetition vs. n − 2 switch), modality (vocal vs. finger vs. foot), and CSI (100 ms vs. 1,000 ms) were within-subject independent variables. Type of cue (direct vs. abstract) was used as between-subject variable. RTs and error percentage were measured as dependent variables. Significance was tested at alpha = .05. As modality is a variable with three levels, we report ε-values when different from 1.0 and use the Huynh–Feldt test to report p-values based on corrected degrees of freedom. However, we still report noncorrected degrees of freedom.
At this point, we do not report any data concerning the between-subjects variable judgement (magnitude vs. parity) because we had no theoretical basis to expect any influence on the overall data pattern. Effects of the variable “judgement” are discussed briefly at the end of the report of Experiment 2.
Results and Discussion
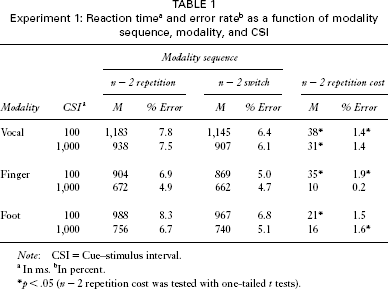
The first two trials of each block were discarded from analysis as were trials with RT above 2,500 ms or below 100 ms (2.4% of otherwise correct trials). For RT and error analysis only trials preceded by at least two other correct trials were included. The mean error rate was 6.6% (2.8% modality errors and 3.8% right/left errors), leading to an inclusion of 78.4% of the trials in the RT and 83.5% in the error analysis. RT and error data are shown in Table 1.
Note: CSI = Cue–stimulus interval.
In ms.
In percent.
p < .05 (n − 2 repetition cost was tested with one-tailed t tests).
To analyse the n − 2 repetition cost and its dependency on the preparation time and the type of cue, we computed four-way analyses of variance (ANOVAs) on RT data and error percentage with the within-subject independent variables modality sequence, modality, and CSI and the between-subjects variable type of cue. Both RT, F(1, 30) = 12.6, p = .001, and error analysis, F(1, 30) = 13.4, p = .001, yielded a significant effect of modality sequence. As predicted, an n − 2 modality repetition was slower (907 ms) and more error prone (7.0%) than was an n − 2 modality switch (882 ms; 5.7%), indicating an overall n − 2 modality repetition cost of 25 ms or 1.3%. Post hoc analyses of error rates, in which both types of error were analysed separately, showed that the effect of modality sequence was significant in modality errors, F(1, 30) = 13.2, p = .001, as well as in right/left errors, F(1, 30) = 4.9, p = .034.
In the RT analysis, the effect of CSI was significant, F(1, 30) = 192.2, p < .001, showing reduced RTs after a long CSI (779 ms) as compared to a short CSI (1,009 ms). Also, in the error data the effect of CSI, F(1, 30) = 5.3, p = .028, was significant, indicating that the overall error rate was reduced with a long CSI (from 6.8% to 5.8%). The interaction of modality sequence and CSI was not significant in RT or error analysis (Fs < 1.6). Thus, the n − 2 modality repetition cost was not significantly affected by the duration of the CSI, although a long preparation time had a substantial beneficial effect in RT and error data.
The RT analysis yielded a significant effect of modality, F(2, 60) = 104.7, p < .001, showing that RTs were higher in vocal trials (1,043 ms) than in foot trials (863 ms) or in finger trials (711 ms). However, modality had no influence on modality sequence (F < 1.3 for the interaction of modality and modality sequence). In the error analysis, modality was not significant, F(2, 60) = 2.9, p = .062, but showed a similar pattern as that for RT data: 6.9% errors in vocal trials, 6.7% errors in foot trials, and 5.3% errors in finger trials. Also, the interaction of modality and modality sequence was not significant (F < 1). Thus, the overall RT level and error rate were different between vocal, finger, and foot trials. Still, the n − 2 repetition cost was not affected by these differences and occurred in each response modality.
With regard to the between-subject variable type of cue, the RT analysis (but not error analysis; F < 1) revealed a significant effect of type of cue, F(1, 30) = 10.2, p = .003. Abstract cues increased the RT (985 ms) as compared to direct cues (804 ms). Additionally, there was a significant interaction of type of cue and CSI, F(1, 30) = 4.7, p = .038, indicating a larger overall RT reduction with long CSI in the abstract-cue condition (from 1,118 ms to 851 ms) than for direct cues (reduction from 901 ms to 707 ms). This interaction was found also in the error analysis, F(1, 30) = 5.2, p = .030, showing that a reduction of error rate with long CSI mainly occurred with abstract cues (reduction of 2.0% with abstract cues vs. 0.0% with direct cues). Thus, when comparing RT and error data with regard to an influence of the type of cue, we consistently found that a long preparation time was more beneficial with abstract cues—that is, when participants had to translate an abstract cue into the modality.
The RT analysis additionally revealed a significant interaction of modality and type of cue, F(2, 60) = 13.7, p < .001. The data pattern shows that the RT benefit of direct cues was larger in vocal trials (290 ms) than in foot trials (154 ms) or finger trials (99 ms). No other interaction in RT analysis was significant (Fs < 2.1). For error data, the interaction of modality sequence, CSI, and type of cue was significant, F(1, 30) = 5.0, p = .034. Descriptively, a long preparatory interval reduced the error rate in both n − 2 repeat and n − 2 switch trials in the abstract-cue group. With direct cues, a long preparatory interval reduced the RT in n − 2 switch trials but not in n − 2 repeat trials. However, this effect only occurred in vocal trials, leading also to a significant four-way interaction of modality sequence, modality, CSI, and type of cue, F(2, 60) = 3.4, p = .040, ε = .984. However, we believe that this is not a systematic effect. No other interaction in the error analysis was significant (Fs < 1.4).
In summary, modality repetition from trial n − 2 to trial n increased both RT and error rate. This increase resulted in an n − 2 modality repetition cost, which can be compared to the n − 2 repetition cost found in other studies (e.g., Arbuthnott & Frank, 2000; Koch et al., 2004; Mayr & Keele, 2000). The n − 2 repetition cost occurred for each response modality (i.e., vocal, finger, and foot responses) and was observed in modality errors as well as in right/left errors. As in other experiments, the n − 2 repetition cost was not influenced by the duration of the preparation interval (Hübner et al., 2003; Mayr & Keele, 2000; Schuch & Koch, 2003).
The type of cue did not affect this general data pattern. The only effect that was found consistently in RT and error data was a larger benefit of a long preparation time with abstract cues. Abstract cues, which need to be translated into the correct modality information, showed a larger CSI effect (i.e., a better performance after a long preparation time) than did direct cues, in which such a translation is not necessary. Therefore, we assume participants could use the long preparation time to encode the cue, translate it into the correct modality, and retrieve this modality. Importantly, the need for the cue translation did not affect the n − 2 modality repetition cost. Thus, we conclude that the n − 2 modality repetition cost can be found with both direct and abstract cues.
The average n − 2 repetition cost of 25 ms in Experiment 1 is relatively small compared to experiments in which participants switched between different stimulus categorizations. For example, Mayr and Keele (2000, Exps. 1a and 1b) found an n − 2 repetition cost of 35 ms and 31 ms, and Arbuthnott and Frank (2000) even found an n − 2 repetition cost of 98 ms. One reason for the small size of the effect that we observed might be that the two RCIs we used were quite long (2,000 ms or 1,100 ms). In contrast to the CSI, the duration of RCI was found to influence the n − 2 repetition cost (Gade & Koch, in press; Koch et al., 2004; Mayr & Keele, 2000). Therefore, the long time interval between each trial might have reduced the n − 2 repetition cost in general. We focus on this question more specifically in Experiment 2, in which we replicate Experiment 1 but reduce the duration of the RCI and additionally manipulate the duration of RCI instead of the CSI.
Experiment 2
Experiment 1 showed that the duration of the preparation interval did not influence the size of the n − 2 modality repetition cost. In contrast, the duration of the RCI was shown to affect the size of the n − 2 repetition cost (Gade & Koch, in press; Koch et al., 2004; Mayr & Keele, 2000). Therefore, in Experiment 2 we further explored whether this difference could be found also when switching between response modalities. We were interested whether the duration of the RCI would affect the size of the n − 2 modality repetition cost.
To study the influence of the RCI we kept the preparation time short (100 ms) in Experiment 2 but manipulated the duration of the time between the response of participants and the next cue. As we found rather small effects in Experiment 1, we also decided to use shorter RCI intervals in general (500 ms and 1,500 ms). Thus, the RSI could vary between 600 ms (RCI 500/CSI 100) and 1,600 ms (RCI 1500/CSI 100), as compared to the long and constant RSI of about 2,000 ms in Experiment 1. We expected to find substantial n − 2 modality repetition cost in the short RCI condition and a reduction of this cost with long RCI.
Again we varied the type of cue and the numerical judgement between participants, but did not expect these variations to affect the general pattern of results.
Method
Participants
A total of 32 new participants (20 female and 12 male, mean age = 26.7 years) were tested in exchange for course requirements. They were evenly assigned to the different experimental conditions (direct vs. abstract cues, and magnitude vs. parity).
Stimuli, Tasks, Procedure, and Design
Stimuli and numerical judgements were identical to those of Experiment 1. The only difference in comparison to Experiment 1 relates to the timing of each trial. CSI was short (100 ms) in each trial, but RCI alternated block-wise between 500 ms and 1,500 ms (including 300 ms for vocal response coding).
Thus, in Experiment 2 modality sequence (n − 2 repetition vs. n − 2 switch), modality (vocal vs. finger vs. foot), and RCI (500 ms vs. 1,500 ms) were within-subject independent variables. Type of cue (direct vs. abstract) was a between-subject variable. Effects of the between-subject variable judgement (magnitude vs. parity) are reported at the end of Experiment 2. RTs and error percentage were measured as dependent variables.
Results and Discussion
The first two trials of each block were discarded from analysis. The outlier criterion was applied to all trials in which the RT exceeded 2,500 ms or was less than 100 ms (3.0% for otherwise correct trials). For both RT and error analysis only correct trials preceded by at least two other correct trials were included. The mean error rate was 8.4% (2.9% modality errors and 5.5% right/left errors). Thus, 74.3% of all trials were included in the RT analysis and 80.4% in the error analysis.
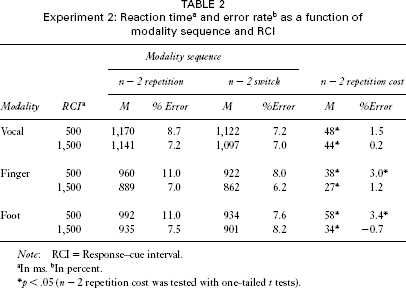
Data were analysed as in Experiment 1. We conducted four-way ANOVAs on RT and error data with the within-subject independent variables modality sequence, modality, and RCI, and with the between-subjects variable type of cue. The analyses revealed a significant effect of modality sequence in RT data, F(1, 30) = 40.3, p < .001, and in the error data, F(1, 30) = 8.2, p = .008. For an n − 2 modality repetition both RT (1,015 ms) and error rate (8.7%) were higher than for an n − 2 modality switch (973 ms, 7.4%; see Table 2). That is, the n − 2 modality repetition cost was found again and, with regard to RT data, was higher than in Experiment 1 (42 ms vs. 25 ms). As in Experiment 1, we conducted post hoc analyses on both types of error separately. Whereas the effect of modality sequence was significant for modality errors, F(1, 30) = 11.1, p = .002, it was not significant for right/left errors (F < 1).
Note: RSI = Response–cue interval.
In ms.
In percent.
p < .05 (n − 2 repetition cost was tested with one-tailed t tests).
The main effect of RCI was significant in both RT, F(1, 30) = 12.5, p = .001, and error data, F(1, 30) = 14.6, p = .001. With a longer time between response and cue the performance was better (971 ms, 7.2%) than with a short time (1,017 ms, 8.9%). The interaction between modality sequence and RCI—that is, the reduction of the n − 2 modality repetition cost (49 ms with short and 36 ms with long RCI)—was not significant in RT data (F < 1.1). However, in the error analysis the interaction of modality sequence and RCI was significant, F(1, 30) = 6.7, p = .015, showing a reduced n − 2 repetition cost with long RCI (0.1%) as compared to short RCI (2.6%). Thus, as in Experiment 1 we found an n − 2 modality repetition cost. The duration of the RCI had a general effect on performance and, at least in the error data, reduced the n − 2 repetition cost. This data pattern is comparable to that found in studies on the effect of switching among stimulus categorizations.
The RT analysis revealed a significant effect of modality, F(2, 60) = 112.9, p < .001, ε = .960. Again, vocal trials resulted in a higher RT (1,132 ms) than did foot trials (951 ms) or finger trials (908 ms). The interaction of modality and modality sequence was not significant (F < 1), indicating that the n − 2 repetition cost was found in each modality. However, there was a significant interaction of modality and RCI, F(2, 60) = 3.7, p = .032. The data pattern shows that the RT reduction with a long RCI is most pronounced in finger trials (65 ms), followed by foot trials (45 ms) and vocal trials (27 ms). In the error data, modality was not significant (F < 1) and did not interact with any other variable (Fs < 1.9).
With regard to the type of cue, only the interaction of modality and type of cue was significant, F(2, 60) = 3.5, p = .038, in RT data. The data pattern indicates that the RT benefit of a direct cue was larger in vocal trials (171 ms) than in foot trials (153 ms) and in finger trials (89 ms). The main effect of type of cue itself was not significant, F(1, 30) = 2.9, p = .096. No other effect or interaction was significant in RT data (Fs < 2.5). In the error analysis, the effect of type of cue was significant, F(1, 30) = 4.7, p = .038, indicating fewer errors with direct cues (6.5%) than with abstract cues (9.6%). No other interaction was significant (Fs < 1.4). In general, we observed no influence of type of cue on the n − 2 modality repetition cost or the RCI effect.
In summary, we found substantial n − 2 modality repetition cost in Experiment 2. The RCI manipulation led to a significant reduction of this cost in error data, but the same numerical data trend was not significant in RT data. In comparison to Experiment 1, the n − 2 repetition cost was increased in RT (42 ms vs. 25 ms) but not in error data (1.3% ms vs. 1.3%). This increase in RT data may have been caused by a general reduction of the RCI duration from 1,550 ms in Experiment 1 to 1,000 ms in Experiment 2. Again, the n − 2 modality repetition cost appeared robust in the face of a manipulation of the type of cue.
The Influence of Judgement
To test for any effects of the manipulation of the judgement (magnitude vs. parity) between subjects, we calculated five-way ANOVAs for both RT and error percentage in Experiments 1 and 2. Modality sequence, modality, CSI/RCI, type of cue, and judgement were used as independent variables. Importantly, neither RT nor error data in Experiment 1 or in Experiment 2 indicated any systematic influence of the judgement on the size of the n − 2 repetition cost. Yet, there have been some general effects of judgement. In Experiment 1, the analysis of RT data revealed a significant effect of judgement, F(1, 28) = 6.4, p = .017. RTs for the parity judgement (961 ms) were higher than those for the magnitude judgement (827 ms). As regards error data in Experiment 1, the interaction of judgement and modality was significant, F(2, 56) = 5.8, p = .005. Whereas participants made more errors in vocal trials (8.1%) than in foot (6.2%) and in finger trials (4.4%) when performing a magnitude judgement, the error rate was higher in foot trials (7.3%) than in finger (6.3%) and vocal trials (5.7%) when participants performed the parity judgement. Additionally, the interaction of judgement, modality, and modality sequence was significant, F(2, 56) = 4.1, p = .022. Participants who performed the magnitude judgement demonstrated the highest n − 2 repetition cost in finger trials (1.4%) followed by vocal trials (1.0%) and foot trials (−0.2%). In contrast, with the parity judgement the n − 2 repetition cost was larger in foot trials (3.4%) than in vocal trials (1.8%) and finger trials (0.7%). No other main effect or interaction was significant in the RT and the error analysis of Experiment 1 (Fs < 3.1). In Experiment 2, for RT data, the interaction of type of cue and judgement was significant, F(1, 28) = 4.3, p = .048. With direct cues, the participants performing the parity judgement were faster (918 ms) than participants performing the magnitude judgement (932 ms), whereas the pattern was reversed with abstract cues (1,207 ms in parity and 918 ms in magnitude). For error data, the interaction of type of cue and judgement was not significant (Fs < 1). However, the interaction of judgement, type of cue, and modality was significant, F(2, 56) = 3.9, p = .026. The data pattern shows that participants on the abstract cue/parity group produced more errors than did participants in the other groups in finger (11.3% in the abstract cue/parity group vs. 5.6–7.9% in the other groups) and foot trials (12.4% vs. 5.2–9.9%), but that this difference could not be found in vocal trials (7.2% vs. 4.3–9.6%). No other effect or interaction was significant in the RT and the error analysis of Experiment 2 (Fs < 3.9). Taken together, the data pattern associated with the numerical judgement does not seem very systematic, so we ignore this variable in the following discussion.
General Discussion
In both experiments, we showed an n − 2 repetition cost when switching among three response modalities. The performance in a modality sequence ABA (e.g., finger/foot/finger) was worse than the performance in a modality sequence CBA (e.g., vocal/foot/finger). This n − 2 modality repetition cost was not influenced by the duration of the preparation interval (Experiment 1). The duration of the time between each trial (i.e., RCI or decay time) affected the n − 2 repetition cost in error data but not in RT data (although there was the same trend; Experiment 2). Similarly, the n − 2 modality repetition cost was higher in Experiment 2 (42 ms) than in Experiment 1 (25 ms)—probably because the RCI was about 500 ms shorter in Experiment 2 than in Experiment 1.
Our experiments replicated the well-known n − 2 task repetition cost on the level of response modalities. What was known for switching among different stimulus categorization tasks (e.g., Arbuthnott & Frank, 2000; Mayr & Keele, 2000) cannot only be found for different response modes (Koch et al., 2004) but also for different response modalities. Thus, the notion of n − 2 repetition cost can be generalized from input variations (i.e., stimulus categorizations) to output variations (i.e., response modalities).
The comparability of the n − 2 repetition cost when switching among response modalities and when switching among stimulus categorizations supports the idea that a “task” in task switching cannot only be defined by different stimulus categorizations but also by different response modalities. The definition of a task (or task set; Rogers & Monsell, 1995) may thus be more general than implicitly assumed in previous studies. A task cannot be defined only by an input manipulation, but also by the use of different effectors (and presumably also by other manipulations, see, e.g., Hahn, Andersen, & Kramer, 2003, for a manipulation of different manual response sets). Thus, it can be argued that a task set is not a unitary concept but consists of different components and that each of these components (e.g., stimulus categorizations or response modalities) is capable of defining a task. Additionally, tasks that are defined by different components seem to result in a similar cognitive task representation.
In previous task-switching studies in which participants switched among three tasks, a task was defined by stimulus categorizations and, thus, by the mapping of stimulus categories to response categories (see, e.g., Arbuthnott & Frank, 2000; Mayr, 2001; Schuch & Koch, 2003). When we speculate about the way response modalities can define a task, we see two different possibilities. On the one hand, it might be that a task indeed is defined by the mapping of stimulus categories and response categories. The influence of response modalities, in such a view, might be due to a binding of response categories and response modalities. That is, each mapping of a stimulus category to a response category is modality specific. For example, the category “odd” is not mapped to the abstract concept “left” per se, but to “respond with left index finger” or “say left”. This way, different tasks are defined by modality-specific stimulus–response rules.
On the other hand, it is possible that what we know for the mapping of stimulus categories and response categories also is true for a mapping of response categories and response modalities. This way, we introduce a second “mapping”. Different tasks, then, would be defined by the mapping of the response categories “left” and “right” to different response modalities (e.g., left-finger vs. left-foot vs. left-vocal). In the same way as the mapping between stimulus categories and response categories can be changed, the mapping between response categories and response modalities can be changed. The observed effects may be independent of which mapping was changed. Presently, the data do not allow us to decide between these two possibilities.
However, each of the two described possibilities indicates that theories and models of task switching should not only focus on explaining how we adapt to demands for changing input transformations. Rather, these theories and models should be able to explain what happens when we perform the same input categorization but switch among different output variables (e.g., different response modalities). Our data suggest that it is likely that the same processes can account for both manipulations—our experiments at least seem to indicate that the cognitive representation is functionally similar, irrespective of what defines a task. Yet, even if the same processes and the same models can account for both manipulations, it is important to take response modalities into account when trying to explain task-switching effects.
Our experiments do not only stress the role of response modalities. The findings concerning the type of cue also point to the relevance of response-related processes in general. The type of cue (and thus the difficulty of cue encoding and modality retrieval) only seemed to influence the benefit of a long preparation time. With abstract cues, a long CSI reduced RT and error rate more than with direct cues. Thus, the additional translation from the abstract symbol to the response modality might (at least partially) have taken place in the preparation time. Importantly, however, the type of cue did not influence the n − 2 modality repetition cost. Thus, we conclude that the difficulty of the cue-to-modality translation (i.e., the retrieval of the modality) is not responsible for the size of the n − 2 repetition cost. Although the retrieval of “task rules” was found to have an influence on task switching in experiments with two tasks (Mayr & Kliegl, 2000), in the present study, the n − 2 repetition cost was not affected by the retrieval difficulty of the response modalities. This finding is consistent with the idea that the n − 2 repetition cost is related to the selection of the specific response (cf. Schuch & Koch, 2003).
In the present study, we showed that the notion of “task inhibition” in the context of the task-switching paradigm also accounts for response modalities. As a consequence, we would assume that response modalities can also define tasks in task-switching experiments with only two tasks. It would, therefore, be important to extend our findings by showing that in different situations (like in two-task experiments) switching between different response modalities produces the same effects as does switching between different stimulus categorizations. Finding the same effects with both manipulations would extend the generality of task-switching effects and would increase the need to account for response-related task-set components in theories of task switching.
Extending our findings to different situations like task-switching experiments with two tasks or dual-task situations would also extend the possible practical implications. This way, we might also gather a greater understanding for situations in which we have to perform different tasks with different response modalities, like in driving a car. Thus, we might also be able to enhance the ecological validity of research on task switching.