
Abstract
The finding that in selective attention tasks responses to previously ignored stimuli are usually retarded is known as negative priming (NP). In previous studies it has been suggested that NP can depend on behavioural goals—that is, NP was observed only for task-relevant object dimensions. We extend these findings with two experiments demonstrating that stronger NP can be observed for task-relevant dimensions than for task-irrelevant dimensions (a) even if participants' tasks vary blockwise within an experiment and (b) if behavioural goals vary from trial to trial. These results suggest that selective NP is a much more flexible process than previously assumed.
The finding that in selective attention tasks responses to previously ignored stimuli are usually retarded is known as negative priming (NP; for reviews, see Fox, 1995; May, Kane, & Hasher, 1995; Neill & Valdes, 1996; Tipper, 2001). For example, if one is to ignore the word DOG on a prime display while reacting to another stimulus (e.g., the word HOUSE), responding to DOG in the following probe display is typically retarded compared to a control condition without stimulus repetition. Since its introduction by Dalrymple-Alford and Budayr (1966), Neill (1977), and Tipper (1985) the NP effect has been replicated and analysed in many studies with different materials, different subpopulations, and different tasks (see, e.g., Allport, Tipper, & Chmiel, 1985; Frings & Wentura, 2005); Neill & Valdes, 1992; Tipper, Brehaut, & Driver, 1990; Wentura, 1999; Wong, 2000).
In this article, we examine a specific NP effect, namely NP depending on behavioural goals. Allport et al. (1985) suggested that NP is a measure of “selection for action”—that is, goal-relevant information is differentiated from distractor information and selected for responding. Moreover, Tipper, Weaver, and Houghton (1994) found evidence that NP is selective depending on task-specific demands. They claimed that NP validly mimics the properties of cognitive control functions in real environments. These findings have been taken up by other researchers (e.g., DeHouwer, Rothermund, & Wentura, 2001; Read & Proctor, 2004; Wentura & Rothermund, 2003). However, in light of inconsistent results, Fox (1995) pointed out that the task specificity of NP could not be simply interpreted as clear evidence for selective NP, which is dependent on behavioural goals. Thus, the main question of this article is: Does selective NP indeed depend on behavioural goals, and, if so, how flexible is this process?
Evidence for Selective NP
NP has been found at the level of responses (e.g., Tipper, MacQueen, & Brehaut, 1988), at the level of physical features (e.g., DeSchepper & Treisman, 1991), and at the level of semantic representations (e.g., Allport et al., 1985). These findings have led to the concept that stimuli are represented multidimensionally, meaning that multiple object dimensions relating to the various properties of an object such as colour, identity, or size, are represented independently. In this regard, it is interesting to note that Tipper and colleagues (1994) reported data suggesting that inhibition of prime distractors is bound to the task-relevant object property and thus depends on the behavioural goals of participants. This finding is important because it is usually accepted that NP reflects a cognitive mechanism that organisms use to selectively control their behaviour in complex environments—and, of course, this mechanism should clearly depend on behavioural goals.
In Tipper et al.'s (1994) Experiment 1, participants had to respond to stimuli that varied in three dimensions (colour, location, identity). Colour served as a cue to discriminate between distractor and target stimuli (and cues were presented before the onset of prime and probe displays), whereas location was the dimension that participants had to respond to. Object identity, however, was completely irrelevant to the task. The results showed NP for the relevant dimension (i.e., object location). No NP effects emerged for the irrelevant dimension (i.e., object identity). In their Experiment 2, they replicated this finding with reversed assignment of object dimensions to task relevance—that is, participants had to respond to object identity whereas object location was completely irrelevant. Again, NP for the relevant dimension was found but no NP effects for the task-irrelevant dimension were observed. Tipper and colleagues concluded that NP indeed depends on behavioural goals. However, to make this claim even stronger, the behavioural goals should have been varied within participants.
A more recent study by DeHouwer and colleagues (2001) suggested that NP is even more selective than assumed by Tipper et al. (1994): Single irrelevant features of prime stimuli (e.g., their valence or their grammatical category) can lead to NP if these features are repeated in probe displays and then become task relevant. However, DeHouwer et al. (2001) used experimental procedures that differed considerably from a typical NP experiment. For example, prime and probe displays were composed of only one stimulus with different dimensions, which had to be separated in order to respond appropriately. Furthermore, different tasks were performed on prime and probe displays, respectively. Hence it is not clear whether their results can be generalized.
Recently, Read and Proctor (2004) provided further evidence for the claim that NP is dependent on behavioural goals. Using a spatial mapping design, they demonstrated that automatically activated incongruent location information of prime stimuli leads to increased reaction times if the previously incongruent location becomes the correct response to the subsequently presented probe target (see also Shiu & Kornblum, 1996). However, Read and Proctor (2004) did not manipulate the behavioural goal, and thus the claim that indeed behavioural goalsdetermine NP can not be verified by their results.
Moreover, some puzzling findings challenge a straightforward interpretation of selective NP. For example, the data pattern of Tipper et al. (1994) was somewhat more complex than our short summary suggests; probe targets that shared two or more stimulus dimensions with prime distractors yielded puzzling effects. For instance, if the prime distractor and probe target shared colour, identity, and location, positive rather than negative effects emerged 1 in their Experiments 1 and 2. In contrast, NP for this condition emerged in their Experiment 3, in which colour cues were presented simultaneously with the onset of prime and probe displays. Moreover, in a study of Neill, Valdes, and Terry (1992; see also Neill, Valdes, & Terry, 1995), selective NP was found for the location of stimuli although location was completely irrelevant to participants' task. These inconsistencies suggest caution in concluding that selective NP occurs at the level of the representation required for task performance (Fox, 1995). Overall, empirical evidence suggests that NP can be found at various levels of stimulus representations. However, the question of whether NP at various levels of stimulus representations is dependent on behavioural goals needs further clarification.
Note, in Experiments 1 and 2 of Tipper et al. (1994) colour cues were given before each prime and probe trial to indicate the colour of the target.
Overview
The main goal of this article is to analyse whether selective NP depends on behavioural goals. In Experiment 1, NP for task-relevant properties was analysed using stimuli varying in three properties (letter type, colour, word identity). The behavioural goal was manipulated within participants: In half of the trials, participants were to respond to word identity; in the remaining trials participants had to respond to colour. Stronger NP was expected for the task-relevant object dimension. Experiment 2 went even further: Behavioural goals changed randomly from trial to trial. Otherwise, Experiment 2 was a replication of Experiment 1. Again, stronger NP for the task-relevant object dimension was expected.
Experiment 1
Method
Participants
A total of 33 undergraduate students took part in the experiment for course credit. All of them had normal or corrected-to-normal vision. Their median age was 21 years (range from 19 to 30 years).
Material
The material set comprised the following eight German nouns with comparable word frequency (CELEX, Nijmwegen, Netherlands): Palme (palm), Perle (pearl), Pulver (powder), Dosis (dose rate), Donner (thunder), Teller (plate), Tenor (tenor), Tunnel (tunnel). Letters were 10 × 6 mm in size. The stimuli were presented at the centre of the screen and with interleaved letters. For half of the participants distractors were presented in uppercase letters whereas targets were presented in lowercase letters (for the other half of participants this assignment was the other way around). Additionally, stimuli were presented in eight different colours. The colours were the following standard colours of the system software Microsoft Windows: blue, green, red, yellow, brown, pink, olive, and turquoise.
Design
Essentially, the design comprised three within-subjects factors: First, priming condition for words (AR, IR, C); second, priming condition for colour (AR, IR, C); and finally participants' task (word vs. colour naming). These factors were varied orthogonally. There were two variations between participants—namely, letter type of stimuli (distractors in uppercase, targets in lowercase, and vice versa) and sequence of tasks (first block word task, second block colour task, and vice versa).
Procedure
The experiment was conducted utilizing self-programmed software (using DirectX and Delphi) and a voicekey-box that was connected to the parallel port. All participants were tested individually. Participants started each trial by pressing the ENTER-key on the computer keyboard. Following the practice blocks, two blocks with 144 trials each were conducted. For half of the participants the task in the first block was to name the word identity of target stimuli, whereas for the other half of participants the task was to name the colour of target stimuli as quickly and as accurately as possible. Each trial consisted of a prime display followed by a probe display, each comprising one target and distractor stimulus. Targets and distractors were indicated by letter type (uppercase vs. lowercase; see Design). The position of the target (just below or just above the centre of the screen) was randomly varied. The distractor was always presented at the respective opposing position. After starting the trial a fixation marker (“+”) appeared for 300 ms before it was replaced by the prime display. The display was presented until the participant reacted and was then replaced by a blank screen, which lasted for 494 ms. After that, the probe display was presented (again, until the participant responded). Then, the participant started the next trial by pressing the ENTER-key.
The standard priming conditions of NP experiments (attended repetition (AR), ignored repetition (IR), and control (C) condition) were constructed independently for both colour and identity properties. In AR (colour) trials the prime target's colour subsequently became the probe target's colour; in AR (identity) trials the prime target (i.e., the word) became the probe target, independently of colour. In IR (colour) trials the prime distractor's colour was repeated as the probe target's colour; in IR (identity) trials the prime distractor (i.e., the word) was repeated as the probe target. In C trials the probe target's dimension (i.e., colour or identity, respectively) was different from the corresponding feature of prime distractor and target.
In each block there were 16 trials for each cell of the 3 (word: AR, IR, C) × 3 (colour: AR, IR, C) design. For half of the participants, the task was to name the word identity of targets in the first block and to name the colour of targets in the second block; for the remaining participants, the assignment was reversed. The sequence of priming conditions within each block was randomized. For each trial, initially four different words and colours were randomly drawn from the stimulus list and were assigned the role of prime target, prime distractor, probe target, and probe distractor. A particular stimulus dimension was never repeated in a subsequent trial and was not chosen again before all remaining stimuli of the list were selected. Then the appropriate replacements were made according to the priming condition of the trial. For example, an IR (word)/C (colour) trial was created by replacing the chosen prime distractor word with the probe target word while keeping the different colours as initially chosen.
Before both the word and the colour task block participants received 24 task-specific practice trials, respectively. To avoid differences in colour naming, participants were shown all colours with their labels before the colour task block. They were instructed to use those labels and were given as long as they needed to memorize the colour labels before the experiment. During the experiment, an experimenter coded participants' responses.
Results
Unless otherwise noted, all effects referred to as statistically significant throughout the text are associated with p-values of less than .05, two-tailed.
For inclusion in the analysis, the responses to both prime and probe displays had to be correct (9.6% of trials were discarded). Moreover, reaction times (RTs) below 200 ms and above 3,000 ms were excluded from the analysis. Overall, these criteria resulted in the exclusion of 16.3% of trials. The sequence of tasks, letter type, and position of stimuli did not interact with any of the following analyses; thus, these factors were discarded as well. Mean RTs and mean error rates as well as mean NP differences (i.e., mean RT for the C condition minus mean RT for the IR condition) are shown in Table 1.
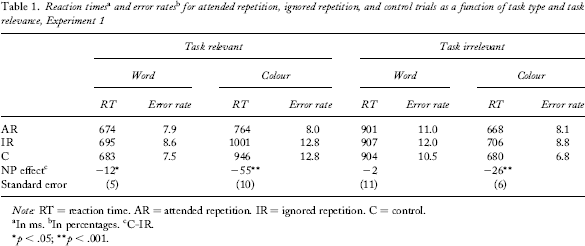
Note: RT = reaction time. AR = attended repetition. IR = ignored repetition. C = control.
In ms.
In percentages.
C-IR.
p < .05;
p < .001.
Selective NP
The critical analysis is a 2 (task relevance: relevant vs. irrelevant) × 2 (task type: word task vs. colour task) analysis of variance (ANOVA) with the four NP differences as dependent variables. The main effect of task relevance was significant, F(1, 32) = 5.22, MSE = 2,399, p < .05. On average, the NP effect was by M = 19 ms (SD = 49 ms) stronger for the task-relevant dimension. This indicates that NP is stronger for the goal-relevant dimension. Additionally, the main effect of task type was significant, F(1, 32) = 14.96, MSE = 2,457, p < .01, indicating that NP was stronger for the colour task. Furthermore, the interaction effect of task relevance and task type did not reach significance, F(1, 32) = 1.26, MSE = 2,370, p = .27, ns. As far as the error rates are concerned, a Task Relevance × Task Type ANOVA yielded no significant effects, all Fs < 0.82, all ps > .37, ns.
To broaden the picture, we additionally analysed mean RTs separately for the IR and C conditions of the relevant and irrelevant dimension, respectively.
NP Effects for the Task-Relevant Dimension
A 2 (priming condition: IR, C) × 2 (relevant task: word vs. colour) ANOVA of mean RTs yielded significant main effects of priming condition, F(1, 32) = 31.23, MSE = 1,206, p < .001, as well as task condition F(1, 32) = 387.41, MSE = 6,919, p < .001, meaning that (a) a significant NP effect emerged and that (b) responses were slower if colour was the relevant dimension. Additionally, the interaction effect of task and priming conditions was significant, F(1, 32) = 15.24, MSE = 996, p < .001. However, the interaction is of the ordinal type because for both tasks a significant NP effect emerged: On average, participants reacted M = 12 ms (SD = 30 ms) slower in IR word trials than in C word trials (independent of colour), t(32) = 2.36, p < .05. For the colour-naming task a comparable pattern emerged. On IR colour trials participants reacted M = 55 ms (SD = 59 ms) slower on average than they did on C colour trials (independent of word identity), t(32) = 5.36, p < .001.
NP Effects for the Task-Irrelevant Dimension
A 2 (priming condition: IR, C) × 2 (irrelevant task: word vs. colour) ANOVA of mean RTs yielded, besides a significant main effect of task type, F(1, 32) = 274.17, MSE = 5,438, p < .001 (which again reflects higher RTs for colour-naming trials), a significant main effect for priming condition, F(1, 32) = 5.60, MSE = 1,206, p < .05. This effect is, however, further qualified by an interaction effect that just missed the conventional criterion of significance, F(1, 32) = 3.31, MSE = 1,418, p = .08. As expected, no NP emerged for the feature word identity if the task goal was to name the colour of stimuli, M = 2 ms (SD = 64 ms), t(32) = 0.21, p = .83. For the feature colour, we found NP even if the goal was to name the word, M = 26 ms (SD = 35 ms), t(32) = 4.34, p < .001. However, in accordance with our hypothesis, this effect was significantly smaller than NP for colour if the goal was to name the colour, t(32) = 2.66, p = .01.
Attended Repetition Effects
For the sake of completeness, we also analysed AR effects. For word trials in the word-naming task a positive priming effect of M = 9 ms (SD = 31 ms) for AR trials emerged that just missed the conventional criterion of significance, t(32) = 1.62, p = .06 (one-tailed). For colour AR trials in the colour task, a positive priming effect of M = 182 ms (SD = 88 ms) emerged that was significantly different from zero, t(32) = 11.83, p < .001. 2 Note that pure repetition (i.e., exactly the same stimulus—the same word in the same colour—was repeated in the AR condition) yielded significant benefits if compared to word control trials (i.e., the word target but not the colour changed from prime to probe), M = 25 ms (SD = 40 ms), t(32) = 3.51, p = .001, as well as if compared to colour control trials, M = 119 ms (SD = 85 ms), t(32) = 8.08, p < .001.
It might be argued that task relevance should moderate the AR effects as well. Note, however, that a moderation of AR by task relevance cannot be safely interpreted because of a confound. Consider, for example, AR effects for words: If word-naming is the task, participants have to repeat their response in the AR condition (e.g., to name “Palm” in prime and probe trial), but they have to alter their response in the C condition (e.g., to name “Thunder” followed by “Palm”). On the other hand, if colour naming is the task, participants have to either repeat or alter their colour response from prime to probe in both AR and C conditions. Note, for NP effects this confound does not exist. Nevertheless, we analysed whether task relevance influenced AR effects. In a 2 (task relevance: irrelevant vs. relevant) × 2 (task type: colour vs. word) ANOVA with AR benefits (control minus AR) as the dependent variable, both main effects as well as the interaction were significant, F(1, 32) = 72.61, MSE = 3,405, p < .001, for task relevance, F(1, 32) = 52.14, MSE = 5,356, p < .001, for task type, and F(1, 32) = 97.09, MSE = 2,229, p < .001 for the interaction. The interaction was of the ordinal type—that is, AR benefits for colour were bigger when colour naming was the task than when word naming was the task (182 ms vs. 14 ms), t(32) = 11.64, p < .001. Numerically but not significantly, an analogue data pattern emerged for words—that is, AR benefits for words were greater when word naming was the task than when colour naming was the task (9 ms vs. 3 ms), t(32) = 0.48, p = .63, ns.
Discussion
The results were clear cut: NP depends on behavioural goals—that is, NP is clearly stronger for task-relevant dimensions. The results confirm previous findings, which had already suggested that NP depends on behavioural goals. However, the data presented here extend previous findings because in our experiment behavioural goals varied within an experiment. This corroborates the claim that indeed behavioural goals determine the level of representation at which NP is found. However, for further validation of the claim that NP captures some “real-world” selection process flexibly adapting to the environment, evidence would be more convincing if behavioural goals were varied randomly from trial to trial. We pursued this strategy with Experiment 2.
It should be noted that NP for colour was significantly different from zero even if colour was irrelevant to the task. We return to this fact in the General Discussion. However, the crucial result that NP is dependent on behavioural goals is not challenged because NP for colour naming was significantly stronger if colour was the relevant task. Nevertheless, this colour-specific NP pattern demonstrates that selective NP is not an all-or-nothing mechanism. Moreover, this finding is in line with previously observed selective NP for irrelevant object properties (e.g., Neill et al., 1992; Tipper et al., 1994).
Experiment 2
Method
Participants
A total of 26 undergraduate students took part in the experiment. All of them were German native speakers and had normal or corrected-to-normal vision. Their median age was 20 years (range from 19 to 25 years); they received 4€ for participation.
Material, Design, and Procedure
The material, design, and procedure were exactly the same as those in Experiment 1 with the following exceptions. A total of 216 trials were conducted. Within this set-up, word task trials and colour task trials were randomly intermixed. For each prime display a black frame with variable line width was presented around the prime stimuli that indicated whether participants should react to word identity (i.e., the frame was 10-pt bold) or to colour (i.e., the frame was 1-pt thin). The assigned task applied to both prime and probe displays (i.e., the task varied only between trials and never within a trial). However, the frame was only shown in the prime display. Moreover, the frame was shown directly after pressing the ENTER-key; hence it was presented 300 ms before the prime stimulus.
This task was much more difficult for participants; hence, they worked through 20 word task trials (all with bold frames) and 20 colour task trials (all with thin frames) and finally received 92 practice trials (in which the task varied randomly from trial to trial), which were administered before the experimental trials started.
Results
For inclusion in the analysis the responses to both prime and probe displays had to be correct (8% of trials were discarded). Moreover, RTs below 200 ms and above 3,000 ms were excluded from analysis. Overall, these criteria resulted in the exclusion of 14% of trials. The factors letter type and position of stimuli did not interact with any of the following analyses and hence were discarded. Mean RTs and mean error rates as well as mean NP differences are shown in Table 2.
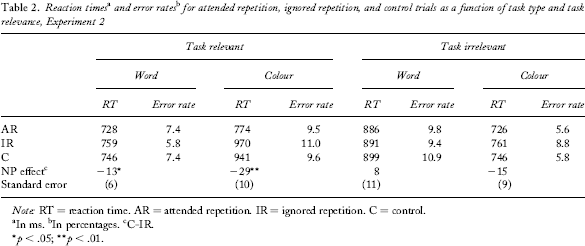
Note: RT = reaction time. AR = attended repetition. IR = ignored repetition. C = control.
In ms.
In percentages.
C-IR.
p < .05;
p < .01.
Selective NP
As in Experiment 1, the critical analysis is a 2 (task relevance: relevant vs. irrelevant) × 2 (task type: word task vs. colour task) ANOVA with the four NP differences as dependent variables. The main effect of task relevance was significant, F(1, 25) = 4.10, MSE = 1,904, p = .05, thereby replicating the finding that NP is stronger for the task-relevant dimension. On average, the NP effect was by M = 17 ms (SD = 44 ms) stronger for the task-relevant dimension. Additionally, the main effect of task type was significant, F(1, 25) = 4.37, MSE = 2,233, p < .05, (i.e., NP was increased for colour naming), while the interaction effect of task relevance and task type did not reach significance, F(1, 25) = 0.17, MSE = 2,765, p = .69, ns.
With regard to error rates, a 2 (task relevance) × 2 (task type) ANOVA only yielded a significant main effect of task type, F(1, 25) = 8.33, MSE = 5.66, p < .01 (i.e., more errors in the colour-naming task), both other Fs < 0.48, both other ps > .50, ns.
NP Effects for the Task-Relevant Dimension
A 2 (priming condition: IR, C) × 2 (relevant task: word vs. colour) ANOVA again yielded a task effect, F(1, 25) = 172.95, MSE = 6,205, p < .001 (i.e., RTs were longer for colour naming). Most importantly, as expected, we found a significant NP effect, F(1, 25) = 12.89, MSE = 890, p = .001. The interaction of task and priming conditions was not significant, F(1, 25) = 1.40, MSE = 1,063, p = .25, ns. For both tasks a significant NP effect emerged: For word naming, participants on average reacted M = 13 ms (SD = 33 ms) slower in IR trials than in C trials (independent of colour), t(25) = 2.10, p < .05. For the colour-naming task, participants on average reacted M = 29 ms (SD = 53 ms) slower in IR trials than in C trials (independent of word identity), t(25) = 2.73, p < .01.
NP Effects for the Task-Irrelevant Dimension
Apart from the significant main effect of task type, F(1, 25) = 128.85, MSE = 4,331, p < .001, a 2 (priming condition: IR, C) × 2 (relevant task: word vs. colour) ANOVA yielded no main effect of priming condition and no interaction effect, F(1, 25) = 0.27, MSE = 1,324, p = .61, ns, and F(1, 25) = 2.52, MSE = 1,436, p = .13, ns. For colour naming, word identity had no influence: Numerically, there was a positive difference (instead of a NP effect) for word trials in the colour task, M = 8 ms (SD = 60 ms), t(25) = 0.69, p = .50, ns. For word-naming trials, colour numerically yielded a NP effect, M = 15 ms (SD = 43 ms). However, this effect was not significant, t(25) = 1.81, p = .08, ns.
Attended Repetition Effects
For reasons of comparison to Experiment 1, we again computed AR benefit effects (see above). For word trials in the word-naming task a positive priming effect for AR trials of M = 18 ms (SD = 37 ms) emerged that was significantly different from zero, t(25) = 2.53, p < .05. For colour AR trials in the colour task, a positive priming effect of M = 167 ms (SD = 79 ms) emerged that was significantly different from zero, t(25) = 10.76, p < .001. 3
Again we analysed whether task relevance influenced AR effects. In a 2 (task relevance: irrelevant vs. relevant) × 2 (task type: colour vs. word) ANOVA with AR benefits (control minus AR) as the dependent variable, both main effects as well as the interaction were significant, F(1, 25) = 55.94, MSE = 2,515, p < .001, for task relevance, F(1, 25) = 64.02, MSE = 2,302, p < .001, for task type, and F(1, 25) = 54.19, MSE = 2,606, p < .001 for the interaction. AR benefits for colour were greater when colour naming was the task than when word naming was the task (167 ms vs. 20 ms), t(25) = 9.74, p < .001. AR benefits for words were not moderated by task relevance (18 ms vs. 18 ms), t(25) = 0.01, p = .99, ns. Note, however, that the AR benefit effects could not be safely interpreted due to a confound in the control condition (see above).
Additionally, we computed pure repetition priming effects (see Experiment 1). Words in the same colour yielded a repetition benefit of M = 30 ms (SD = 51 ms) that was significantly different from zero, t(25) = 3.00, p < .01. Repetition priming for colour was again stronger, with M = 111 ms (SD = 57 ms), and was also significantly different from zero, t(25) = 10.02, p < .001.
Discussion
The data of Experiment 2 confirm findings of Experiment 1: Significantly stronger NP effects emerged for task-relevant dimensions than for task-irrelevant dimensions. Even if behavioural goals varied randomly from trial to trial, NP depended on these goals.
In this experiment, we found no significant NP for the task-irrelevant conditions. However, since the irrelevant NP effect for colour failed to reach significance in Experiment 2, we cannot fairly reject the hypothesis that NP never emerges in task-irrelevant conditions. We return to this issue in the General Discussion.
General Discussion
Experiments 1 and 2 yielded clear evidence that selective NP is tied to behavioural goals. As outlined before, it is assumed that stimuli are represented at various levels of representations relating to different object properties. Our data showed strong evidence that behavioural goals determine which object properties are selectively ignored. Moreover, the selective ignoring of distractor information is much more flexible than previously assumed. In Experiment 1, goals were manipulated between blocks (but within participants), and NP for the task-relevant dimension was significantly stronger than NP for the task-irrelevant dimension. Even if behavioural goals changed randomly from trial to trial as in Experiment 2, NP for task-relevant dimensions was stronger than NP for task-irrelevant dimensions. Hence, the data reported here strongly suggest that the selection processes that are measured in NP paradigms are highly flexible and can adapt to task demands that are changing from trial to trial. This flexibility seems essential for controlled behaviour in complex environments and strengthens the claim that “real-world” cognitive processes are measured in NP paradigms. In the remainder of the General Discussion we discuss our data with regard to inhibition and memory accounts of NP.
Selective NP with Regard to Inhibition and Memory Accounts
Our results can be explained by the inhibition account and are in line with the arguments put forward by Tipper et al. (1994). In particular, the model by Houghton and Tipper (1994) fits the data well (although with a small change to the model, see Tipper et al., 1994). This model (Houghton & Tipper, 1994; Houghton, Tipper, Weaver, & Shore, 1996) assumes that stimulus representations are written into an object field and that an internal match–mismatch process, which is controlled by a supervisory system, enhances the representation of target information while suppressing representation of distractor information. Thus, the status of the distractor representation receives an activation level below baseline. If the distractor becomes the target in the next trial, reaction time is retarded because the representation needs more activation for excitation. As Tipper and colleagues (1994) have suggested, the model could be improved if one assumes that object representations are weighted by task relevance. If, for example, colour is the task-relevant object property, the colour representation of stimuli would receive more excitatory feedback than would other properties in the case of target stimuli and would receive more inhibitory feedback in the case of distractor stimuli. Hence the model could straightforwardly predict selective NP with selective inhibition processes. This model even offers an explanation for the fact that we found NP for the task-irrelevant colour: If relevance changes the weights of attributes, a corresponding change of NP effects would be predicted. It would not be predicted, however, that NP should never appear for irrelevant dimensions because of other causes. For instance, specific dimensions might be more affected than others by the inhibition process. That is exactly what we have found: For both dimensions—i.e., words as well as colours—we observed a moderation of NP by changing the relevance of the dimension. In this regard it poses no problem for the model that colour leads to bigger NP than words and that NP for colour is found even if it is irrelevant.
Selective NP can also be explained by the episodic retrieval theory (Neill, 1997), at least if one assumes that memory traces of stimuli also constitute multiple representations of the stimuli. If so, then the “do-not-respond tag” would be added to the task-relevant part of the memory trace of a stimulus, and, thus, selective NP could be explained by a selective episodic retrieval account (Read & Proctor, 2004). For example, if the task is to respond to colour, only the colour part of the stimulus representation would be encoded with a “do-not-respond tag”. Interference from retrieved response information with required responses would only occur for responses to colour. Thus, in principle, episodic retrieval can also explain our data, although this approach seems a little less suited to explain the NP effect for colour if colour was irrelevant.
Overall, both Experiment 1 and Experiment 2 demonstrated highly flexible NP for task-relevant object properties. This flexibility has not been demonstrated so far in other studies. It emphasizes the importance of the NP paradigm as a valid measurement of “real-world” cognitive control processes.