
Abstract
In Experiments 1 and 2 rats received uncorrelated presentations of a light conditioned stimulus (CS) and a food unconditioned stimulus (US) on each day of a preexposure phase. Control subjects received the same number of USs during the first half of preexposure and the same number of CSs during the second. Uncorrelated preexposure retarded inhibitory conditioning. Experiment 3 showed, however, that the different patterns of US preexposure experienced by the two groups could in itself influence the course of subsequent inhibitory conditioning. When this factor was equated by restricting the uncorrelated treatment to the first half of the pre-exposure phase (Experiment 2) or by extending the control treatment throughout the phase (Experiment 4) it was found that uncorrelated preexposure retarded excitatory conditioning, but facilitated inhibitory conditioning. This outcome challenges an interpretation in terms of the concept of learned irrelevance, which predicts that uncorrelated preexposure should retard both forms of conditioning.
Pavlovian conditioning is retarded in animals that have previously experienced uncorrelated presentations of the events to be used as the conditioned and unconditioned stimuli (the CS and US). Demonstrations of this learned irrelevance effect can be found in studies of pigeon autoshaping (Mackintosh, 1973), of conditioned suppression with rats (e.g., Baker, 1976; Bonardi & Hall, 1996), and in appetitive conditioning with rats (e.g., Baker & Mackintosh, 1977; Bennett, Maldonado, & Mackintosh, 1995; Bennett, Wills, Oakeshott, & Mackintosh, 2000). However, although the reality of the effect is well established, the mechanisms responsible are still a matter for debate. One possibility is that it reflects the operation of a special learning process—that during preexposure the animals are able to learn that the CS and the US are unrelated to each other, and that this retards acquisition when a correlation between these stimuli is established in a subsequent conditioning phase. (According to the specific account of learned irrelevance proposed by Mackintosh, 1975, this learning process is one that reduces the value of a US-specific associability parameter associated with the CS.) Before taking this theoretical step, however, it is appropriate to establish that the data said to require it cannot be explained in terms of simpler, known processes of associative learning.
First, it is necessary to demonstrate that the learned irrelevance effect cannot be explained in terms of the known effects of preexposure on the individual stimuli. Preexposure to a CS will retard subsequent conditioning—the well-known latent inhibition effect (e.g., Lubow, 1989). Similarly, conditioning is retarded after preexposure to the US, an effect known as the US-preexposure effect (e.g., Randich & LoLordo, 1979). Given these facts, it is not surprising that the procedure used in demonstrating learned irrelevance, which involves preexposure to both CS and US, should produce a more marked retardation of conditioning than that produced by exposure to either just the CS or just the US (the control procedures used by Baker & Mackintosh, 1977; Mackintosh, 1973). In order to demonstrate that uncorrelated preexposure has an effect over and above that produced by the summation of the latent inhibition and US-preexposure effects, it is necessary to make a comparison with a control condition in which subjects experience the same number of CSs and USs (but see Matzel, Schachtman, & Miller, 1988, for an alternative strategy). This was arranged in the experiments by Bennett et al. (1995, 2000) who gave control subjects preexposure to the same number of CSs and USs as those given in uncorrelated preexposure, but presented these events in separate sessions. More specifically, Bennett et al. (2000) gave rats in the uncorrelated condition one session per day over 10 days, each comprising CS and US presentations; rats in the control condition received only USs during the first 5 days of preexposure and only CSs during the last 5 days. Their finding, that the uncorrelated group learned less well than the control group in subsequent tests of conditioning, argues against any simple explanation of the learned irrelevance effect in terms of latent inhibition and the US-preexposure effect.
Second, it is necessary to show that the effects of uncorrelated preexposure cannot be explained in terms of associative changes that might be produced. However carefully it is arranged, it is possible that the schedule used during the preexposure phase of these experiments might establish associative links between the CS and the US. The fact that the CS does not evoke a conditioned response (at least after extensive training with the uncorrelated schedule, see Kremer, 1971) is evidence that the uncorrelated schedule does not generate excitatory conditioning (but see Rescorla, 2000). Inhibitory conditioning, however, would not be evident in the direct response evoked by the CS, and if the uncorrelated schedule were to establish the CS as an inhibitor, the retardation of subsequent excitatory conditioning could be explained without recourse to a concept of learned irrelevance. The best evidence against this interpretation comes from the demonstrations (provided, e.g., by Baker & Mackintosh, 1977, and by Bennett et al., 2000) that uncorrelated preexposure retards not only excitatory but also subsequent inhibitory conditioning.
This brief survey of alternative interpretations of the learned irrelevance effect makes apparent the central importance of the experiments reported by Bennett et al. (2000). As far as we are aware, this study is the only one to demonstrate a retardation of both excitatory and inhibitory conditioning and to make its comparison against a control condition that was matched to the experimental group in its experience of the CS and the US. Accordingly, the procedure used by Bennett et al. forms the focus of the present investigation. In our experiments we made use of an uncorrelated schedule identical to that described by Bennett et al. In Experiment 1 we attempted to replicate their central finding, looking for a retardation of inhibitory conditioning after preexposure to this schedule. Comparison was made with the control condition used by Bennett et al. in which subjects received a block of US sessions in the first half of preexposure and a block of CS sessions in the second half. Although this replication was successful, doubts about the adequacy of this comparison led us, in Experiment 2, to attempt to replicate the effects obtained in Experiment 1, but with an experimental group that, like the control group just described, experienced an interval of 5 days between its exposure to presentations of the US and the start of conditioning. In this experiment we assessed the effects of preexposure on both excitatory and inhibitory conditioning. The results of Experiment 2 indicated that the details of the schedule of US presentation might, in themselves, have important effects on the course of subsequent conditioning. This possibility was tested directly, and confirmed, in Experiment 3. In Experiment 4, therefore, we introduced a novel control condition in which the rats experienced separate CS-only and US-only sessions throughout the whole of the 10-day preexposure phase. (A preliminary report of Experiments 3 and 4 was presented by Bonardi & Ong, 2003.)
Experiment 1
The aim of Experiment 1 was to replicate, with our procedures, the finding of Bennett et al. (2000) that preexposure to uncorrelated presentations of a CS and US will retard subsequent inhibitory conditioning, when comparison is made with a control group that has received the same number of CS and US presentations during preexposure. The design (see Table 1) was directly modelled on that of Experiment 1b of Bennett et al. (2000). All subjects (rats) received 10 days of preexposure. For group U–U (uncorrelated) each session contained 10 presentations of the CS (a light) and 10 presentations of the US (food), the schedule controlling these presentations being the same as that described by Bennett et al. (2000). Group US–CS (control) received US presentations over the first 5 days and CS presentations over the second 5 days. In order to match the number of events experienced, while maintaining the same average interevent interval, the session duration was doubled for these animals, allowing the presentation of 20 events (CSs or USs) in each session. In the test phase all rats learned a discrimination in which a new stimulus (a clicker) was reinforced, whereas a compound (light plus clicker) was not. If uncorrelated preexposure retards inhibitory learning we would expect the decline in responding to the compound to proceed less readily in group U–U than in group US–CS.
Daily treatment during the preexposure phase of each experiment
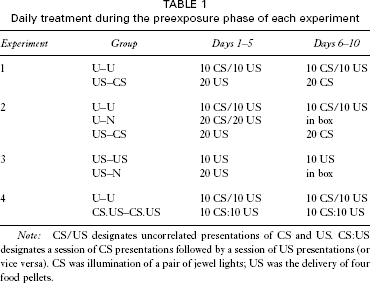
Note: CS/US designates uncorrelated presentations of CS and US. CS:US designates a session of CS presentations followed by a session of US presentations (or vice versa). CS was illumination of a pair of jewel lights; US was the delivery of four food pellets.
Method
Subjects
The subjects were 32 male hooded Lister rats with a mean ad lib weight of 485 g (range: 425–540 g). They had previously been used in a study of flavour aversion but were naive to all the stimuli and procedures used in the present experiment. They were housed in pairs in plastic tub cages with sawdust bedding. The colony room was lit from 8 a.m. to 8 p.m., and experimental procedures were conducted during the light portion of the cycle. Before the start of training the rats were reduced to 80% of their ad lib weights and were maintained at this level for the rest of the experiment by being fed a restricted amount of food at the end of each session.
Apparatus
Two sets of four standard Skinner boxes (supplied by Campden Instruments Ltd., Loughborough, UK) were used, each set being situated in a different room. Each box had three walls of sheet aluminium, a transparent plastic door as the fourth wall, a grid floor, and a white translucent plastic ceiling. One of the walls adjacent to the door contained a recessed food tray covered by a transparent plastic flap, 6 cm high × 5 cm wide, which was hinged to the top of the opening to the food tray. Pushing this flap inward from its vertical resting position allowed subjects to gain access to the food tray. An inward movement of the flap actuated a microswitch, and each closing of the switch was recorded as a single response. The flap automatically returned to its resting position when the subject removed its snout from the food tray. The sets of boxes in the two rooms differed only in that one set had black and white checked wallpaper on the wall that served as the door and on the wall immediately opposite; these sets of boxes were counterbalanced across the various groups in all experiments. Mixed-composition, 45-mg food pellets (Noyes, New Hampshire) could be delivered to the food tray. Retractable levers fitted alongside the food tray remained withdrawn throughout the experiment. Each box was housed in a sound- and light-attenuating shell. The light stimulus was provided by illuminating two 2.8-W jewel lights, situated on the front wall to the right and left of the food tray. The click stimulus, (10 Hz, 75 dB) was presented through a speaker mounted on the wall opposite the food tray. The boxes were controlled by a BBC microcomputer programmed in a version of BASIC.
Procedure
Magazine training. In the first 40-min session the animals were trained to retrieve pellets from the food tray; pellets were delivered according to a variable time (VT) 60-s schedule during this session. Any rat that did not all consume all the pellets delivered in this session received further sessions until it had done so.
Test excitor training. The rats next received excitatory conditioning with the stimulus (the clicker) that was to be used as the reinforced CS in the final test stage. This consisted of five 60-min sessions, each containing eight presentations of the 10-s clicker, followed immediately by the delivery of four food pellets over an interval of approximately 3 seconds; this constituted the US in all the reported experiments. The mean intertrial interval was variable with a mean duration of 6 min (range: 5–7 min). Responses (operations of the magazine flap) were recorded during the clicker and during the 10-s stimulus-free period (the pre-CS period) that preceded each trial.
Preexposure. The rats were then divided into two equal-sized groups, matched in terms of the responding they showed to the clicker in the previous stage. Group U–U received 10 daily sessions of uncorrelated presentations of a CS, the 10-s presentation of the two jewel lights, and the US. These events were scheduled in the manner described by Bennett et al. (2000) and may be described as follows. Each session contained 10 CS and 10 US presentations; the first CS presentation occurred 2 min after the start of the session, and thereafter CS presentations occurred after a variable intertrial interval (ITI) with a mean of 2 min (range 1–3 min). US deliveries were scheduled between CS presentations in the following manner. Each ITI was divided into ten equal time bins, 1–10; Bin 1 immediately followed the offset of the CS, and Bin 10 immediately preceded the 10-s pre-CS period that preceded each CS presentation. Within each session, one of the ten USs was scheduled to occur in Bin 1 of one of the ten ITIs, another US was scheduled to occur in Bin 2 of one of the ten ITIs, and so on, so that each of the set of bins 1–10 contained one US. This allocation was also constrained such that no more than two USs could occur in any one ITI, and the presentation of two USs in a single ITI could only occur once per session. This yielded sessions with either a single US in each of the ten ITIs, or sessions with a single US in eight of the ten ITIs, one ITI with two USs, and one ITI with none. This produced a rough alternation of CS and US presentations. In addition, as every session contained one US in one of the Bin 1s, and as Bin 1 immediately followed CS offset, and as the US was presented at the start of the bin, this yielded one CS–US pairing per session. The resulting sessions were of 25-min duration. Group US–CS received exactly the same schedule of US delivery as did group U–U and also exactly the same schedule of CS presentations. For these rats, however, CSs and USs were presented in separate blocks of sessions. For the first 5 days of this phase they received two consecutive daily sessions of preexposure to the US (effectively, therefore, a 50-min session containing 20 US presentations) and, on each of the second 5 days, two consecutive daily sessions of preexposure to the CS (effectively a 50-min session containing 20 CS presentations).
Conditioning. On the day after the last preexposure session all rats received a further session of training with the clicker, identical to those given in the first stage. Seven sessions of discrimination training followed. In these presentations of the clicker were reinforced, whereas presentations of a serial compound of the light immediately followed by the clicker were nonreinforced. The first three sessions consisted of six trials, two with the clicker and four with the compound. The ITI was variable with a mean of 6 min (range: 5–7 min), and the session duration was 40 min. Despite the fact that these parameters were copied from the study by Bennett et al. (2000), it became apparent that this regime was insufficient to maintain responding to the clicker. Accordingly a further session of reinforced training with the clicker alone (identical to those described above) was given after discrimination sessions 3 and 5; in addition, from test session 4 onward, the proportion of trials was changed such that there were four reinforced and two nonreinforced trials per session.
Data treatment. Responses (magazine entries) were recorded during CS presentations and pre-CS periods. Rates of response in the absence of the stimuli were low, so no correction was made for baseline levels of responding. In order to assess the inhibitory properties of the light, a suppression ratio was calculated of form a/(a + b), where a represents the score in the presence of the clicker when it was preceded by the light, and b the score in the presence of the clicker when presented alone. A score of .5 indicates equal levels of response on both clicker and light–clicker trials; formation of the discrimination is shown as a declining suppression ratio. On two occasions a rat failed to respond to the clicker during a trial block (see below), and in these cases the average of that rat's performance for the preceding and following trial blocks was substituted.
Results and Discussion
In the initial phase of training with the test excitor, all rats learned to respond during the clicker; the mean corrected response rates during the last session of this stage were 23.02 responses per min for group U–U and 20.44 for group US–CS. These scores did not differ reliably (F < 1). The corresponding pre-CS rates were 1.60 and 1.46 responses per minute, and these scores did not differ (F < 1). In these and all subsequent statistical analyses a significance level of p < .05 was adopted.
No data were recorded during the preexposure phase. Performance on the test discrimination is summarized in Figure 1, which shows the mean suppression score for each of the two groups over three blocks of sessions. The responses emitted by a given animal for each trial type were pooled over all trials in the block before the ratio was calculated. As noted above, after the first three test sessions, the proportion of reinforced to nonreinforced trials per session was altered. Accordingly, the data from the first three test sessions were pooled together to generate the score for Block 1; thereafter the data were pooled into two-session blocks. The two groups showed similar suppression on the first block, but by Block 2 group US–CS showed evidence of having learned the discrimination, displaying a substantially lower suppression ratio than did group U–U. This description was supported by the results of an analysis of variance (ANOVA) with group and blocks as the variables. There was no significant main effect of group, F(1, 30) = 2.13, but there was a significant effect of blocks, F(2, 60) = 21.19, and a significant Group ?Blocks interaction, F(2, 60) = 3.74. Exploration of this interaction with an analysis of simple main effects revealed that there was a difference between the two groups on Block 2, F(1, 87) = 8.61. This difference in the ratio scores was not a consequence of differences between the groups in their responding to the clicker or in their baseline response rates. Mean response rates during the reinforced clicker trials for all three test blocks are shown in Table 2. An ANOVA performed on these data with group and blocks as the variables revealed no significant main effects, largest F(2, 60) = 1.13; although there was a significant Group × Blocks interaction, F(2, 60) = 4.60, simple main effects analysis revealed that the groups did not differ on any of the test blocks, largest F(1, 44) = 2.43. The pre-CS response rates are also shown in the table; an ANOVA with group and blocks as factors revealed no significant effects or interactions, largest F(2, 60) = 2.38.
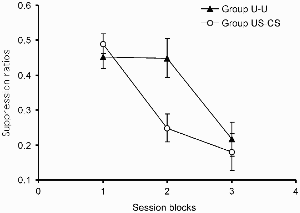
Experiment 1: Group mean suppression ratios in the test. Vertical bars show standard errors.
Experiment 1: Group mean response rates during test blocks
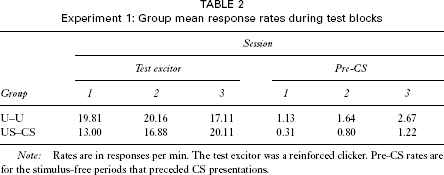
Note: Rates are in responses per min. The test excitor was a reinforced clicker. Pre-CS rates are for the stimulus-free periods that preceded CS presentations.
The results of this experiment are clear-cut. Acquisition of a discrimination in which a CS must acquire inhibitory properties is retarded in rats that have received preexposure involving uncorrelated presentations of that CS and the US, relative to a control group given preexposure involving a block of US presentations followed by a block of CS presentations. These results thus confirm those of Bennett et al. (2000, Exp. 1b) and may be said to demonstrate the operation of a learned irrelevance process.
Experiments 2A and 2B
When two groups differ in their rate of learning, the claim that one of them shows retardation rests on assumptions about the effects of the treatment given to the comparison group. There is no reason to suppose that the treatment given to the control group in Experiment 1 might have acted to facilitate learning in the test phase of that experiment, but it should be acknowledged that the groups compared in that experiment differed in a number of ways, in addition to the fact that group U–U received uncorrelated presentations of the CS and US in a way that group US–CS did not. For one thing, group US–CS received twice as much exposure to the training context as did group U–U; for another, group U–U received presentations of the US throughout the whole of the preexposure phase, whereas group US–CS experienced 5 days between the last US presentation of preexposure and the start of the test phase. The possible importance of these factors was (implicitly) acknowledged in the experiments carried out by Bennett et al. (1995) in their study of the effects of learned irrelevance training on subsequent excitatory conditioning. In these experiments they made use of an experimental condition in which the uncorrelated presentations of the CS and the US were restricted to the first 5 days of the preexposure period, the second 5 days consisting of either simple exposure to the apparatus or being spent in the home cage. They were able to demonstrate that this procedure produced a retardation of subsequent excitatory conditioning with respect to controls that received either a block of US-only sessions followed by a block of CS-only sessions or the reverse arrangement.
The present experiment continues the investigation of these factors. The design (see Table 1) included the uncorrelated and control conditions of Experiment 1, but in addition we included a further experimental group (labelled group U–N in the table), which, like the experimental group of Bennett et al. (1995), received the uncorrelated treatment in the first half of the preexposure phase and simple exposure to the context (N, nothing) in the second half. In Experiment 2a, these treatments were followed by an excitatory conditioning test phase. On the basis of previous results we might expect a retardation with respect to group US–CS both in group U–U (Bennett et al., 2000, Exp. 1a) and in group U–N (Bennett et al., 1995, Exp. 3). In Experiment 2b we made use of inhibitory conditioning in the test phase. The comparison between group U–U and group US–CS should allow replication of the effect obtained in Experiment 1 (and also by Bennett et al., 2000, Exp. 1b). The comparison between group U–N and this control condition has not been investigated previously for the inhibitory case, but if the treatment given to group U–N generates a learned irrelevance effect, a retardation of learning can be expected in this case too.
Method
Subjects and Apparatus
The rats for Experiment 2a were 48 male hooded Lister rats with a mean ad lib. weight of 438 g (range: 385–550 g). They had been subjects in an experiment on flavour aversion learning, but were naïve to the stimuli and procedures employed here. The experiment was performed in two identical replications of 24 rats (n = 8 for each group). Experiment 2b used a further 48 male hooded Lister rats with a mean ad lib weight of 411 g (range 320–500 g). The experiment was again conducted in two replications, each of 24 rats (n = 8 in each group). The rats in one replication were completely naïve; those in the other had taken part in a study on flavour aversion learning, but were naïve to the stimuli and procedures used here. The rats were housed and maintained as in Experiment 1. The apparatus was the same as that used in Experiment 1.
Procedure
The magazine training and preexposure treatments for groups U–U and US–CS of Experiment 2a were the same as those described for these groups in Experiment 1; no training with the clicker was given in this experiment. Group U–N received two consecutive 25-min sessions of uncorrelated pre-exposure, each containing 10 presentations of the CS and 10 of the US, on each of the first 5 days of the preexposure phase; apart from their different distribution across the preexposure phase, these sessions were identical to those experienced by group U–U. For the second 5 days they were placed in the apparatus for the same period of time (i.e., 50 min on each day) but no stimulus presentations occurred. The test phase consisted of 10 sessions each containing 10 trials in which the light was presented, immediately followed by the US. These sessions were identical to those from the preexposure phase, except for the fact that each US presentation was programmed to occur at the offset of the CS, so that all trials were CS–US pairings. These trials were scheduled with the same variable ITI (with a mean of 2 min) as that used for CS presentations in the preexposure phase; two consecutive daily sessions were given.
The same three preexposure conditions were used in Experiment 2b. The procedure differed from that of Experiment 2a only in that a phase of training with the test excitor (the clicker) intervened between initial magazine training and the preexposure phase and between the preexposure and the test phase, and in the nature of the test phase. In an attempt to ensure that responding to the clicker was firmly established, the amount of initial test excitor training was increased from that used in Experiment 1: There were six 50-min sessions each comprising 20 reinforced presentations of the clicker, separated by a variable ITI of mean duration 2 min (range 1–3 min). Two further sessions of this type were delivered immediately prior to the test. As in Experiment 1, the test phase consisted of discrimination between the clicker and the light–clicker compound. There were eight daily 25-min sessions, each comprising seven reinforced clicker trials and three nonreinforced compound trials. The variable ITI was the same as that used in the preexposure phase. In all other respects the procedure was the same as that described for Experiment 1.
Results and Discussion
Acquisition of responding to the light in the test phase of Experiment 2a is shown in Figure 2. In order to attenuate the effects of individual differences in levels of responsiveness, we made use of a ratio measure: the number of responses to the CS during each session divided by the number in the absence of the CS. As the figure shows, this score increased with training in all three groups, but did so more readily in group US–CS than in either of the other two groups, which did not differ from one another. An ANOVA performed on these data with group and sessions as the variables revealed no main effect of group, F < 1, but there was a significant main effect of session, F(9, 405) = 25.41, and a significant Group × Session interaction, F(18, 405) = 2.25. A simple main effects analysis revealed that there was a difference among the groups on Sessions 8 and 10, Fs(2, 144) = 5.72 and 3.38, respectively. Tukey's test revealed that on Session 8 both group U–N and group U–U differed from group US–CS; on Session 10, group U–N differed from group US–CS. Groups U–N and U–U did not differ on either of these sessions. These differences were not a consequence of differences in baseline responding; Table 3 shows group mean response rates in the absence of the CS for each of the test sessions. An ANOVA performed on these data with group and sessions as the variables revealed only a main effect of sessions, F(9, 405) = 11.1; there was no main effect of group (F < 1), and no Group × Session interaction, F(18, 405) = 1.04.
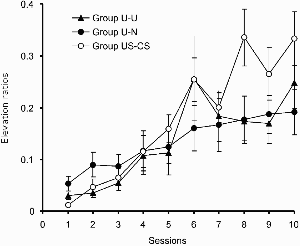
Experiment 2a: Group mean elevation ratios for responding during the light in the test. Vertical bars show standard errors.
Experiment 2a: Group mean response rates in the absence of the CS during the test sessions
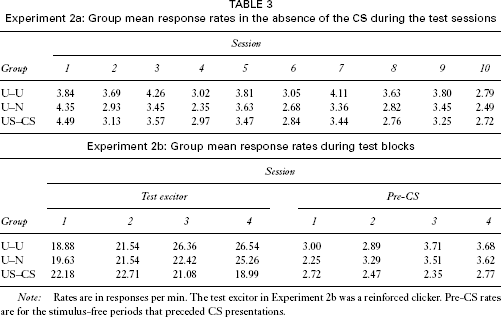
Note: Rates are in responses per min. The test excitor in Experiment 2b was a reinforced clicker. Pre-CS rates are for the stimulus-free periods that preceded CS presentations.
These results replicate the findings reported by Bennett et al. (1995, 2000). Uncorrelated preexposure to CS and US retarded subsequent excitatory conditioning with respect to a control group that had received a block of US presentations in the first half of the preexposure phase and a block of CS presentations in the second half of the phase. This retardation was evident both in rats that had received uncorrelated presentations throughout preexposure (group U–U) and in rats that had received a block of event-free sessions in the second half of the preexposure phase (group U–N). The results of Experiment 2a thus appear to dispel any concerns over the effects of the exact details of the scheduling of event presentations during the course of preexposure, at least for the case of excitatory conditioning. The result of Experiment 2b, however, complicate the picture.
In the first phase of Experiment 2b, all rats learned to respond during the clicker. The mean response rates during the last session of this stage were 25.44 responses per min for group U–U, 26.14 responses per min for group U–N, and 27.84 responses per min for group US–CS. An ANOVA revealed that these scores did not differ, F < 1. The corresponding pre-CS response rates were 2.68, 2.98, and 2.64 responses per min for groups U–U, U–N, and US–CS, respectively, and these scores also did not differ, F < 1.
Responding to the clicker was well maintained during the test phase. The group mean response rates during reinforced clicker trials in each of the two-session blocks of the test are shown in Table 3. An ANOVA with group and blocks as the variables revealed no significant effects or interactions, largest F(6, 135) = 2.14. The pre-CS response rates on these trials are also shown in Table 3, and these also did not differ, an observation supported by the results of an ANOVA with group and blocks as the variables, which revealed no significant effects or interactions, Fs < 1.
As in Experiment 1, discrimination performance on the test was assessed by means of a suppression ratio. Group mean ratio scores for successive two-session blocks of this phase are shown in Figure 3. On one occasion a rat made no responses during the click in either session of a block, and the average of that subject's ratio scores for the preceding and following blocks was substituted. There were two notable features of these results. First, a comparison of group U–U and group US–CS revealed that we succeeded in replicating the observation of Bennett et al. (2000) and of Experiment 1b, that inhibitory learning appears to be retarded after uncorrelated preexposure: Animals in group U–U showed higher suppression ratios, and hence weaker inhibition, than those in group US–CS. The second, more surprising, observation was that this effect was reversed when the uncorrelated CS and US presentations were confined to the first half of the preexposure stage—animals in group U–N showed lower suppression ratios, and hence greater inhibition, than those in group US–CS.
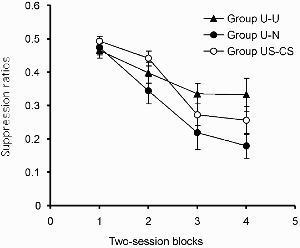
Experiment 2b: Group mean suppression ratios in the test. Vertical bars show standard errors.
This description was largely supported by the results of an ANOVA with group and blocks as the variables. This revealed significant main effects of group, F(2, 45) = 4.04, and of block, F(3, 135) = 30.89, but no interaction between these two factors, F(6, 135) = 1.63. Exploration of the group effect using the Newman–Keuls test revealed a significant difference between group U–N and group US–CS, although the difference between group U–U and group US–CS fell short of significance. Our failure to find a significant difference in this latter comparison may have been a consequence of the procedural changes that we made in this experiment in order to maintain the excitatory strength of the clicker during the test (compared with Experiment 1 we gave more pretraining to the clicker and used a higher proportion of reinforced to nonreinforced trials during the test). However, the direction of the difference was the same as that obtained in Experiment 1 (and also by Bennett et al., 2000); thus, given that the balance of evidence suggests the existence of this effect, we feel justified in concluding that inhibitory learning is retarded when the comparison is made between group US–CS and group U–U.
The results for group U–N were quite unexpected. In this case there was no sign of a retardation of learning. When the treatment given to rats in the uncorrelated group was modified so that they, like those in the control group, experienced all their USs in the first half of the preexposure phase, the pattern of results was reversed such that the light acquired inhibitory properties faster after uncorrelated preexposure. In other words, when the groups were matched in this way, the uncorrelated schedule retarded subsequent excitatory conditioning (Experiment 2a) but facilitated inhibitory learning (Experiment 2b).
The results for group U–U (a retardation of both excitatory and inhibitory conditioning) are thus consistent with the suggestion that the preexposure treatment generates a learned irrelevance effect. Those for group U–N, on the other hand, are what might be expected if the uncorrelated schedule resulted in inhibitory learning during preexposure. The remaining experiments were designed to help resolve this apparent contradiction.
Experiments 3A and 3B
In order to develop an explanation for the puzzling pattern of results generated by Experiment 2, it is necessary to explain why the U–U and U–N treatments should produce different effects and should do so only in the case in which the preexposed CS was trained as an inhibitor in the test phase. There are many procedural differences between these two groups, but perhaps the most salient (from the rats’ point of view) is that group U–N experienced a food-free interval between the exposure phase and the test phase, whereas group U–U experienced food presentations throughout preexposure. It is not obvious why this difference should be of consequence but, as a first step, we thought it worthwhile to investigate the matter directly. In the present experiment, therefore, we gave two groups of rats the same preexposure as that experienced by groups U–U and U–N of the previous experiments, except that no CSs were delivered during the preexposure phase. That is, rats in group US–US (see Table 1) received food presentations throughout the preexposure phase (paralleling group U–U), whereas rats in group US–N received all their food presentations in the first half of the preexposure phase (paralleling group U–N), merely being placed in the apparatus, in the absence of scheduled events, in the second half of the phase. In Experiment 3a this preexposure was followed by excitatory conditioning with the light as the CS; in Experiment 3b the light was trained as an inhibitor. The question of interest was whether these different schedules of US-preexposure would have different effects on subsequent conditioning and, more specifically, whether the effect on the inhibitory and excitatory procedures would differ.
Method
Subjects and Apparatus
The subjects for Experiment 3a were 16 naïve male hooded Lister rats with a mean ad lib weight of 314 g (range: 285–330 g). A further 16 rats (mean ad lib weight 268 g, range 260–285 g) were used in Experiment 3b. They were reduced to 85% of their ad lib weights before the start of the experiment. They were housed and maintained exactly as in Experiment 1. The apparatus was a set of four Campden Instruments Skinner boxes similar to those used in Experiment 1.
Procedure
All rats received an initial session of magazine training. Those in Experiment 3a then went on directly to the preexposure phase. Those in Experiment 3b received five sessions of training with the test excitor prior to the start of preexposure. Each session contained six clicker–food pairings; the mean ITI was variable with a mean duration of 6 min (range: 5–7 min). In each experiment the rats were divided into two equal-sized groups, US–US and US–N, for the preexposure phase. The pre-exposure schedule was the same as that used for groups U–U and U–N of the previous experiment, except that no CS presentations were delivered.
In the test phase, rats in Experiment 3a received 12 daily sessions of excitatory conditioning with the light as the CS. The procedure was the same as that described for Experiment 2a. On the day after the last preexposure session, rats in Experiment 3b received a further session of training with the clicker, identical to those administered earlier. The test sessions each consisted of six trials: four reinforced clicker trials and two trials on which the clicker was preceded by the light and nonreinforced. The mean ITI was variable with a mean duration of 6 min (range: 5–7 min), and the whole session lasted 40 min. As the rats had received no preexposure to the light that was to become the conditioned inhibitor, acquisition was considerably faster than that in the previous two experiments, and only three sessions were required in this phase.
Results and Discussion
For Experiment 3a, ratios were calculated as in Experiment 2a, and the resulting data are shown in Figure 4. Both groups acquired conditioned responding over the course of the test session: For much of the test there was little evidence of a difference between them; although at the very end there appeared to be a slight advantage for group US–US, this difference was not statistically reliable (and appeared to be largely due to one rat in group US–N responding at very high rates during the CS). An ANOVA with group and session as the variables revealed a significant main effect of session, F(11, 154) = 13.36, but no effect of group, or Group × Session interaction (Fs < 1). The mean rates of responding outside the CS periods during the test sessions are shown in Table 4. These did not differ between the two groups; an ANOVA with group and sessions as the variables revealed no significant effects or interactions, largest F(1,14) = 3.54. There is no evidence from these results that the different schedules of US preexposure experienced by the two groups influenced the course of subsequent excitatory conditioning.
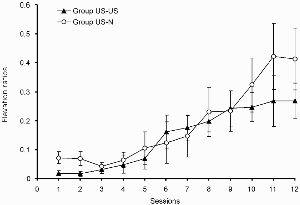
Experiment 3a: Group mean elevation ratios for responding during the light in the test. Vertical bars show standard errors.
Experiment 3a: Group mean response rates in the absence of the CS during the test sessions
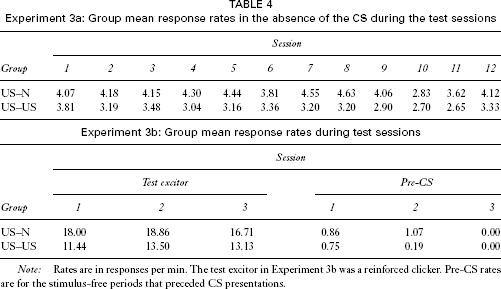
Note: Rates are in responses per min. The test excitor in Experiment 3b was a reinforced clicker. Pre-CS rates are for the stimulus-free periods that preceded CS presentations.
In the first phase of Experiment 3b, all rats learned to respond during the clicker. The mean corrected response rates during the last session of this stage were 20.43 response per min for group US–N and 15.50 for group US–US. These scores did not differ significantly, F(1, 13) = 1.37. The corresponding pre-CS rates were 0.14 and 0.75 responses per min, respectively, and these scores did not differ, F(1, 13) = 2.81.
Suppression ratios were calculated for each test session, as in Experiment 2b. One rat failed to respond to the clicker on any of the test sessions and so was omitted from the study: In addition one rat did not respond to the clicker during the first test session; as it was therefore not possible to take the average of the ratio scores from the preceding and following sessions, as in previous experiments, on this occasion the group mean ratio was substituted for that rat's score. An ANOVA with group and session as the variables was performed on the response rates on reinforced clicker trials from all three test sessions (see Table 4). This analysis revealed no significant main effects or interactions, largest F(1, 13) = 1.73. The pre-CS response rates are also shown in the table, and these did not differ; an ANOVA with group and sessions as factors revealed no significant effects or interactions, largest F(2, 26) = 1.31.
The suppression ratio scores shown in Figure 5 indicate that group US–N acquired the discrimination more rapidly than did group US–US, a conclusion that was supported by the results of an ANOVA with group and sessions as the variables. This revealed no significant main effect of group, F(1, 13) = 1.92, but there was a significant effect of session, F(2, 26) = 4.59, and a significant Group × Session interaction, F(2, 26) = 3.50. Exploration of this interaction with an analysis of simple main effects revealed that there was a difference between the two groups on Session 2, F(1, 36) = 4.79. In addition, there was an effect of sessions in group US–N, F(2, 26) = 8.63, but not in group US–US, F < 1.
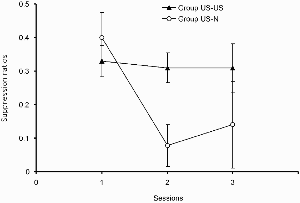
Experiment 3b: Group mean suppression ratios in the test. Vertical bars show standard errors.
The results of Experiment 3b show that preexposure in which the US is presented throughout the preexposure phase will retard subsequent inhibitory learning (compared to the procedure in which US presentations were concentrated in the first half of the phase). We can only speculate as to why this should be so. One possibility is that this treatment results in strong context conditioning, which summates with responding on compound trials, reducing the apparent inhibitory effect. But such an account would predict that an effect should be observed on trials on which the test excitor was presented alone and also on the simple excitatory conditioning test trials of Experiment 3a, where no differences were observed. Perhaps it is the case that responding on trials on which both an excitor and inhibitor are presented will be more unstable, being determined by two opposing influences, than that on trials in which an excitor is presented alone—this might make responding on compound trials more susceptible to the effects of differences in contextual associative strength.
However, whatever their explanation, the findings of the present experiment require us to reconsider the interpretation of the results presented so far. We have shown (Experiment 2a) that uncorrelated preexposure retards subsequent excitatory conditioning, whether this preexposure involves the presentation of USs throughout the preexposure phase (group U–U) or is concentrated in the first half of the phase (group U–N). This result is consistent with the notion of learned irrelevance. To rule out other interpretations, such as the possibility that the preexposure schedule renders the CS inhibitory, it is necessary to show that subsequent inhibitory conditioning is also retarded. This result is found, however, only if the animals given uncorrelated preexposure differ from the controls in experiencing USs throughout the preexposure phase (group U–U of Experiments 1 and 2b). If the two groups are equated in this respect (group U–N of Experiment 2b), then uncorrelated preexposure produces the opposite effect, accelerated inhibitory learning—what would be expected if uncorrelated preexposure made the CS inhibitory. The finding of the present experiment, that inhibitory learning is retarded in rats that have experienced US presentations throughout the preexposure phase, suggests the following interpretation of this, otherwise puzzling, pattern of results. First, experience of the uncorrelated schedule used in these experiments renders the CS inhibitory, thus slowing subsequent excitatory conditioning (Experiment 2a) and facilitating inhibitory conditioning (group U–N of Experiment 2b). Second, experiencing USs throughout the preexposure phase retards inhibitory learning, an effect that acts to mask the inhibitory effect of the uncorrelated schedule, producing the effect obtained in group U–U of Experiments 1 and 2a.
Experiments 4A and 4B
Whatever the merits of the hypothesis just outlined, the results of the previous experiment demonstrate that it is necessary, in assessing the effects of the uncorrelated schedule, to make comparison with a control group that experiences the same general schedule of US presentation. That is, the U–U group of the previous experiments should be compared with a control that also receives food presentations on each day of the preexposure phase. In this experiment, therefore, we introduced a novel control treatment that received the same number of CS and US presentations as group U–U, and which experienced these events throughout preeexposure, but did not experience uncorrelated presentations of CS and US in the same session. Animals in this group (labelled group CS.US–CS.US) received two sessions on each of the 10 days of preexposure, one comprising CS presentations and the other US presentations (see Table 1). Excitatory conditioning was tested in Experiment 4a and inhibitory conditioning in Experiment 4b. The comparison of these two groups should allow assessment of the hypotheses derived from the previous experiments. If the uncorrelated preexposure given to group U–U generates a learned irrelevance effect, then learning should be retarded both in Experiment 4a and in Experiment 4b. However, if the uncorrelated procedure produces inhibitory learning, then we can expect to find a retardation of excitatory learning in Experiment 4a, but a facilitation of inhibitory learning in Experiment 4b.
Method
Subjects and Apparatus
The subjects in Experiment 4a were 32 male hooded Lister rats, with a mean ad lib weight of 387 (range: 350–425 g). A further 32 rats from the same stock (mean ad lib weight of 520 g; range 420–660 g) were used in Experiment 4b. All rats had previously taken part in a study on flavour aversion learning, but were naïve to the stimuli and procedures used in the present experiments. They were housed and maintained exactly as in Experiment 3, and the apparatus was that used in Experiment 1.
Procedure
After magazine training, all received five sessions of excitatory conditioning with the clicker, exactly like those administered in Experiment 1.
The preexposure given to group U–U was the same as that described for the equivalent groups of previous experiments. The control group (group CS.US–CS.US) received separate sessions of CS preexposure and US preexposure, as did subjects in the US–CS groups of the previous experiments, but the distribution of these sessions across the training phase differed. Specifically, there was one CS session and one US session on each day of the preexposure phase, one session immediately following the other. Across days these were scheduled in a double alternation sequence (i.e., a CS session followed by US session on day n; US then CS on day n + 1; US then CS on day n + 2; CS then US on day n + 3; and so on).
The rats in Experiment 4a then received six conditioning sessions of excitatory conditioning with the light, following the procedures described for Experiment 2a. Those in Experiment 4b received a single reminder session with the clicker, identical to those given in the pretraining phase, followed by eight sessions of discrimination training between the reinforced clicker and the nonreinforced light–clicker compound. The detailed procedures were those described for the test sessions of Experiment 3b. Details not specified above were the same as those reported for previous experiments.
Results and Discussion
In the first phase all rats learned to respond during the clicker in the first stage of training. The mean response rates during the last session of this stage in Experiment 4a were 22.88 responses per min for group U–U and 25.00 for group CS.US–CS.US. These scores did not differ (F < 1). The rates of pre-CS responding for this session were 0.66 and 1.60 responses per min for groups U–U and CS.US–CS.US, respectively, and these too did not differ reliably, F(1, 29) = 1.98. In Experiment 4b the mean rates during the last session of clicker training were 24.29 responses per min for group U–U and 27.05 responses per min for group CS.US–CS.US (F < 1). The rates of pre-CS responding on this session were 0.71 and 0.80 responses per minute for groups U–U and CS.US–CS.US, respectively (F < 1).
Elevation ratios for the excitatory conditioning test of Experiment 4a are shown in Figure 6. (One subject, in group CS.US–CS.US, stopped collecting any food pellets in the test stage, and the data produced by this subject were omitted from the analyses that follow.) Both groups showed learning over the course of the test, and this learning was less rapid in group U–U than in group CS.US–CS.US. An ANOVA with group and sessions as the variables revealed no main effect of group, F(1, 29) = 1.43, but there was significant main effect of session, F(5, 145) = 29.32, and a significant Group × Session interaction, F(5, 145) = 2.71. Exploration of this interaction with a simple main effects analysis revealed that the groups differed significantly on Session 6, F(1, 97) = 8.01. The groups did not differ in their baseline response rates; the rates of responding outside the CS periods are shown in Table 5. An ANOVA performed on these data with group and session as the variables revealed no significant effects or interactions, largest F(1, 29) = 1.33.
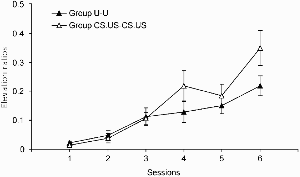
Experiment 4a: Group mean elevation ratios for responding during the light in the test. Vertical bars show standard errors.
Experiment 4a: Group mean response rates in the absence of the CS during the test sessions
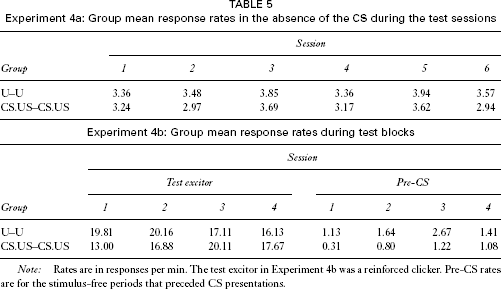
Note: Rates are in responses per min. The test excitor in Experiment 4b was a reinforced clicker. Pre-CS rates are for the stimulus-free periods that preceded CS presentations.
The data for the test phase of Experiment 4b were reduced to two-session blocks as in Experiment 2b. The group mean response rates during reinforced clicker trials in each of the blocks are shown in Table 5. An ANOVA with group and blocks as factors revealed no significant effects or interactions, largest F(3, 90) = 2.16. The pre-CS response rates on these trials are also shown in Table 5, and these did not differ, an ANOVA again revealing no significant effects or interactions, largest F(3, 90) = 2.02.
The results of principal interest, the ratio scores for the test discrimination, are shown in Figure 7. One rat failed to respond during the clicker in the last block, but since this rat had produced a ratio of zero on the previous two blocks (suggesting complete suppression had been achieved), a score of zero was assigned for this final block. Over the course of testing both groups came to show suppression of responding to the clicker when it was accompanied by the light, but acquisition of this suppression was more rapid in group U–U than in group CS.US–CS.US. This description of the data was supported by the results of an ANOVA with group and block as the variables. This revealed no effect of group, F(1, 30) = 3.42, but a significant effect of block, F(3, 90) = 16.11, and a significant Group × Block interaction, F(3, 90) = 3.83. Exploration of this interaction with a simple main effects analysis revealed that the groups differed significantly on Block 2, F(1, 105) = 12.14.
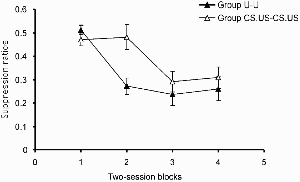
Experiment 4b: Group mean suppression ratios in the test. Vertical bars show standard errors.
The results of Experiment 4a demonstrate that the uncorrelated preexposure procedure used by Bennett et al. (2000) will produce a retardation of excitatory conditioning even when the control group, like the experimental group, has experienced USs throughout the pre-exposure phase. Although this outcome in itself is consistent with the notion that the uncorrelated procedure generates a learned irrelevance effect, the results of Experiment 4b disconfirm this interpretation. The learned irrelevance notion predicts that uncorrelated preexposure should also retard inhibitory conditioning, but Experiment 4b produced quite the reverse effect—a facilitation of inhibitory learning in group U–U.
Although we have argued that the control procedure used in the present experiment is superior in some respects to that used in earlier experiments, we should acknowledge that other (unwanted) differences between the groups remain. Although the groups were matched in that they both received USs on each day of the preexposure phase, they were not matched in their total exposure to the context: Group CS.US–CS.US received twice as many sessions as group U–U. In order to equate context exposure, it would have been necessary to give rats in group U–U a session in the context without USs on every day of preexposure, or to halve the length of sessions in group CS.US–CS.US—either of these procedures would have constituted a substantial departure from the procedures used by Bennett et al. (2000). We think it unlikely, however, that this difference could be responsible for the results of this experiment—in Experiments 1 and 2b we also observed enhanced inhibitory conditioning after uncorrelated preexposure in group U–U, even though this group received the same total exposure to the context as did its control group, group US–CS. Taken together, these results support the conclusion that it is the uncorrelated preexposure treatment, rather than differences in exposure to the context, that are responsible for the enhancement in inhibitory conditioning that we observed in Experiment 4b.
General Discussion
In the experiments reported here we have successfully replicated the finding of Bennett et al. (2000) that preexposure to uncorrelated presentations of a CS and a US will retard subsequent conditioning in which this CS is trained as an inhibitor (Experiments 1 and 2b). This finding is important, as it is central to the claim that such preexposure generates a learned irrelevance effect. There are many demonstrations that uncorrelated preexposure can retard excitatory conditioning (as was observed in Experiments 2a and 4a of the present report), but it is when this outcome is coupled with a similar retardation of inhibitory learning that an explanation in terms of learned irrelevance seems to be required.
Other aspects of our results, however, cast doubt such an explanation. Experiment 3 compared groups given preexposure to just the US, either throughout the whole preexposure phase or concentrated in just the first half. These treatments had no differential effects on subsequent excitatory conditioning, but we found, in Experiment 3b, that inhibitory conditioning proceeded less rapidly in the subjects that had experienced the US throughout. Now in some studies of the uncorrelated procedure presentations of the US (and CS) have been given just in the first half of the phase (the U–N groups of our experiments); in others they have been given throughout (the U–U groups). It is clearly important, given the results of our Experiment 3, that each of these groups should be compared against its appropriate control (which we take to be one that has experienced the same schedule of US presentation during preexposure). When this is done the results are clear. In each case the uncorrelated group performs less well in excitatory conditioning, but is superior in inhibitory conditioning.
The learned irrelevance hypothesis cannot explain this pattern of results, but how is it to be explained? It is what would be expected if the uncorrelated treatment left the CS with some degree of associative inhibition, but our usual assumption about the uncorrelated schedule is that, after extensive training, it will leave the CS associatively neutral. This is not to say, however, that no associative learning goes on during exposure to an uncorrelated schedule. It has long been known that chance pairings of the CS and US can establish excitation in the early stages of such training (Kremer, 1971), a result that has recently been confirmed for an appetitive training procedure, like that used here, by Rescorla (2000). Rescorla went on to argue that further experience of the uncorrelated schedule engages an inhibitory learning process that acts to counteract the initial excitatory learning. There was no suggestion, however, that this training would establish the CS as a net inhibitor—although after extensive training the CS will control both excitatory and inhibitory associations, these will match, leaving the CS associatively neutral.
Whatever is true of Rescorla's (2000) experiments, the results of those reported here suggest that we might need to reconsider the consensus view that the uncorrelated treatment results in a CS with no net associative strength. Our results are best accommodated by the assumption that the CS possessed some measure of inhibitory strength at the start of the conditioning phase. How might this have arisen? A close examination of the details of the schedule used in these experiments reveals no sign of a negative correlation between the CS and the US—indeed, quite the reverse. In our uncorrelated schedule (which was modelled directly on that described by Bennett et al., 2000), the animals received 10 of the 100 US presentations immediately following the CS. As the total duration of CS during preexposure was 16.67 min (100 × 10-s presentations), the probability of a US presentation occurring per min of CS presentation was 10/16.67 = .60. The total duration of the sessions was 250 min, making the overall probability of food .40 (100/250). According to this analysis, therefore, CS presentations heralded an increase in the probability of food. If we hold to the view that the uncorrelated schedule produces inhibitory learning, then it follows that we cannot rely on the overall correlation alone to predict what type of learning will occur about the CS; some other factor must play a role. Perhaps the critical factor is that the way the CSs and USs were scheduled in these experiments: The constraint that a US should occur during each ITI, resulted in an almost completely reliable alternation between CSs and USs. If we assume that animals are particularly sensitive to this sort of predictability, this may explain why its effects were sufficient to override the fact that CS and US were sometimes paired. How this suggestion might be incorporated within our formal theories of association formation remains to be specified. It should also be noted that this inhibitory effect must have been substantial, given it was observed even in Experiment 4, in which the control condition was one in which the control group experienced some sessions with only CS presentations, and others with only US presentations. This procedure in itself has been shown to produce “between sessions” inhibitory learning under some circumstances (e.g., Baker, 1977). Although the conditions in which Baker observed this effect were rather different from those observed here (Baker employed a conditioned suppression procedure, whereas the present experiments all used appetitive reinforcement), if a similar effect were to be found with our parameters, it would suggest that the inhibition produced by the “uncorrelated” schedule was even stronger than that produced by the between-sessions treatment.
Whatever the merits of this analysis, the results reported here have important implications for the concept of learned irrelevance. This concept has received its strongest support from the demonstration (Bennett et al., 2000) that exposure to the uncorrelated schedule can retard not only subsequent excitatory and but also subsequent inhibitory conditioning. Our experiments have shown that, when the comparison is made with what may be considered a more appropriate control group than that used by Bennett et al., inhibitory conditioning is facilitated not retarded. This result, along with the uncontested finding of retarded excitatory conditioning, is best explained by the suggestion that the uncorrelated schedule used in these studies is capable of generating inhibition.