
Abstract
To obtain information on the mechanism of microbiologically influenced corrosion (MIC) under anaerobic conditions, metabolic processes and microbial community dynamics were analysed for MIC of carbon steel with lake-mud microbial consortium as inoculum. During one month of MIC experiment, the microbial consortium corroded carbon steel with sulphate reduction, methane and acetate production. The bacterial and archaeal community structure in the suspension culture and the crust formed on carbon steel were compared using high-throughput Illumina MiSeq sequencing and quantitative polymerase chain reaction. The data indicated that sulphate-reducing bacterium (Desulfovibrio genus), amino acid-degrading bacterium (Aminobacterium genus) and a hydrogenotrophic methanogen Methanofollis among Archaea increased in the crust. These microbes were estimated to collaboratively induced corrosion of carbon steel under anaerobic condition.
Introduction
Corrosion can cause serious economic damage to the steel infrastructures in the oil, gas and electric power industries. The estimated global annual cost of corrosion is 3-4% of the gross domestic product in industrialised countries [1]. Although the chemical reaction of metal with oxygen is the main cause of corrosion under aerobic conditions, it has been reported metals are quite vulnerable to corrosion induced by microbes under anaerobic conditions [2 4]. It was estimated that, among other processes, ∼10% of all corrosion damage is associated with microbiologically influenced corrosion (MIC) [5]. In large industrial facilities such as power plants, natural waters from a lake, river or sea are often used as a coolant in heat exchangers to remove heat in pipelines. Owing to moderate temperatures and diverse microbes in natural waters, pipelines used in cooling systems are susceptible to MIC. Sulphate-reducing bacteria (SRB) are considered to be primarily responsible for MIC. Corrosion by individual SRB strains has been investigated extensively, including various mechanisms of corrosion; e.g. consumption of hydrogen [6,7], production of corrosive sulphide chemicals [8] and acquisition of energy with iron as the electron donor [5]. In contrast, methanogens [9], acetogens [10] and nitrate-reducing bacteria [11] have been recently isolated growing on iron substrates. Moreover, culture-independent technologies revealed the composition of microbial communities in crusts collected from MIC-affected equipment [12–15]. These culture-dependent and -independent studies revealed that a diverse microbial group, rather than a single group, is commonly present at sites of rapidly progressing MIC, and implied that various types of microbes (SRB, acid-producing fermentative bacteria, acetogens and methanogens) and interactions between them might play a role in corrosion. However, key responsible microbes in crusts formed on metal surfaces and their impact on corrosion remain controversial because analysis about the dynamics of the microbial community during the corrosion process was limited [16].
Microbes accumulated on the surface of pipelines flowing the natural waters induce MIC in cooling systems. However, knowledge of microbial community dynamics during corrosion and the metabolic processes leading to MIC progression is limited, because during in situ experiments, it requires a long time for corrosion to occur, and detailed changes over time make reproducibility difficult. In the current study, corrosion tests with carbon steel, often used for pipelines and various infrastructures, were performed using previously enriched corrosive culture from a brackish lake. Metabolic and microbial profiles during the progression of corrosion were compared to reveal the microbes responsible for corrosion and their interaction in MIC of carbon steel under a blackish condition. Although corrosion in marine systems have been investigated, studies employing brackish conditions are limited [17,18]. This study makes important contributions in the understanding of corrosion in industrial facilities using brackish water, such as pipelines in a cooling system.
Materials and methods
Cultivation of iron-corroding enriched culture
The iron-corroding microbial community (H-1 enrichment), previously enriched from brackish lake mud with iron granules [19], was utilised in the current study. The enriched culture was maintained in serum bottles (50 m:l capacity) filled with 30 mL of a VM-I medium [20] containing 6 g L−1lactic acid and 5 g L−1Na2SO4, with the NaCl concentration modified to 10 g L−1, to mimic brackish conditions. The medium pH was adjusted to 7.5 with HCl, and the medium was autoclaved at 121°C for 15 min. After autoclaving, the pH of the medium was maintained around 7.0. Three grams of iron granules (1-2 mm in diameter, 99.98% purity, Tewksbury, MA, Alfa Aesar) were added to the medium. After sealing the serum bottles, the gaseous phase was repeatedly replaced with a N2/CO2 gas mixture at a 4:1 (v/v) ratio. The H-1 enrichment culture (1 mL) was inoculated into freshly prepared medium containing the iron granules, after brief homogenisation to detach microbes from the surface of the iron granules. All serum bottle cultures were incubated at 30°C without shaking. All chemicals were purchased from Fujifilm Wako Pure Chemical Corp. (Osaka, Japan).
Iron-corroding experiment with enriched culture
To analyse the iron-corroding activity and changes in the microbial community during H-1 enrichment culture, carbon steel coupons were used instead of iron granules. Carbon steel SS400 sheets (0.5 mm thick) were polished using #120 and #400 paper (MonotaRO Co. Ltd, Hyogo, Japan). The coupons were subsequently cleaned with acetone solution and stored in a vacuum desiccator for 1 day before use. A polished carbon steel sheet was cut into 8 × 8 mm coupons and their initial weight was determined. All coupons were autoclaved in a serum bottle under N2 atmosphere at 121°C for 15 min. A sterilised coupon was transferred to a 50 mL serum bottle filled with 30 mL of the modified VM-I medium. After sealing and gas replacement of the headspace to N2/CO2 gas (4:1, v/v), the bottles were inoculated with 1 mL of the H-1 enriched culture. Modified VM-I medium without carbon steel coupons was also prepared, as an iron-free control. In addition, another VM-I medium was prepared with coupon and without inoculation of the H-1 enriched culture for abiotic control experiments. Overall, 72 culture bottles were prepared for eight sampling days (days 0, 3, 5, 7, 10, 14, 21 and 28), in triplicate for each experiment.
On each sampling day, the serum bottles were opened and the carbon steel coupons were retrieved to observe their surface and determine their weight loss associated with corrosion. The surface of carbon steel was observed by a laser microscope (VK-X200, KEYENCE, Osaka, Japan). The carbon steel coupons were weighed after crusts formed on the coupon surface were removed using Clark's solution (the ratio of 1000 mL hydrochloric acid, 20 g Sb2O3 and 50 g SnCl2), according to ASTM G1 [21]. Weight loss associated with corrosion was calculated by comparing the initial weight to weights determined after incubation. The data are presented as the means and standard deviation for three coupons. After washing the carbon steel coupons in a Clark solution, the surface of the coupons was visualised using a scanning electron microscope (VE7800, KEYENCE, Osaka, Japan) with the beam voltage of 15 kV.
Chemical analysis of carbon steel corrosion
On each sampling day, H2, CO2 and CH4 contents in the headspace were determined using a gas chromatograph equipped with a thermal conductivity detector. Samples were loaded onto a Molsieve 5A column (GL sciences Inc., Japan) at 100°C using Ar as carrier gas (CP400; Varian Medical Systems, Palo Alto, CA, USA) [22]. H2S in the headspace was measured via H2S detector tubes (4L, 4LT, GASTEC CO, Kanagawa, Japan). The amount of organic acids (lactate, acetate, formate and propionate) in cultures was quantified using a high-performance liquid chromatographic system equipped with ultraviolet and infrared detectors (LachromeElite; Hitachi, Tokyo, Japan). Briefly, 10 μl of a culture filtered by Millex GP filter (SLGPX13NL, MilliporeSigma, Burlington, MA, USA)was loaded onto a GelPak column (GL-C610H-S; Hitachi, Tokyo, Japan) at 40°C and eluted at 0.5 mL min−1 using 0.1% H2PO4 [22]. Sulphate ion concentration in the culture medium was determined using an ion chromatograph (ICS-2000, Dionex, Sunnyvale, CA, USA) equipped with IonPac AS-11HC column (Thermo Fisher Scientific Inc., Waltham, MA, USA) at 30°C and samples were eluted at 1.4 mL min−1 using 27 mM KOH [19].
Molecular analysis of microbial communities
DNA extraction and quantification
A sample of the suspension culture (1.5 mL) was collected from the incubated cultures on all sampling days. In addition, on days 7, 14, 21 and 28, the cells in crusts formed on carbon steel coupons were washed with fresh medium and collected by scraping. Nothing was scraped from a surface of carbon steel coupon in the abiotic control experiment. DNA was extracted from both sets of samples using the Power Max soil isolation kit (MO Bio Laboratories, Carlsbad, CA, USA). The quantity and purity of the extracted DNA were assessed using a NanoDrop ND-1000 spectrophotometer (NanoDrop, Wilmington, DE, USA). The abundance of prokaryotic and methanogenic 16S rRNA genes was evaluated by quantitative polymerase chain reaction (qPCR), as described previously [23]. The standard 16S rRNA genes for prokaryotes and methanogenic archaea were prepared using the pGEM-T Easy vector system (Promega, Madison, WI, USA) after PCR of Escherichia coli IAM 1264 with the primer set B8F (5′-AGAGTTTGATCCTGGCTCAG-3′; E. coli positions 8-27) and U1492R (5′-GGYTACCTTGTTACGACTT-3′; E. coli positions 1492-1510) [24] and Methanosarcina barkeri JCM 10043 with the primer set A21F (5′-TCCGGTTGATCCYGSCRG-3′; E. coli positions 8-25) and U1492R [25]. The standard curves were constructed by dilution of these plasmids. Amplification efficiencies of PCR calculated from triplicate standard curves were 91.6 ± 7.6 and 85.1 ± 3.8, respectively. To calculate the total copy number per reaction, the copy number per area of carbon steel coupon, from which the DNA was extracted or per bulk volume, was multiplied by the total area of carbon steel coupon or the total suspension volume, respectively. Student's t-test was used to compare group means. No contamination was confirmed during the DNA extraction step and PCR procedure by PCR amplification of an abiotic control sample prepared in the same manner as the cultures.
Illumina MiSeq sequencing
Sequences of primers used for next-generation sequencing.

Processing of MiSeq sequencing data
The raw paired-end reads were merged and processed using QIIME (version 1.91) [27] to remove short sequences, singletons and noisy reads. Chimeric sequences were identified and removed using UCHIME [28,29]. The sequences were clustered into operational taxonomic units (OTUs) with a 97% similarity cut-off using UPARSE [30]. Taxonomic assignments were made using the USEARCH global alignment program [28] based upon sequence similarity in the Greengene database [31]. The obtained representative nucleotide sequences were deposited in the NCBI database under the accession numbers LC458583–LC458594.
Sanger sequencing of the 16S rRNA gene of methanogens
The archaeal 16S rRNA gene fragments were amplified using the primers Parch340R plus Parch519R [22] and DNA was extracted from suspension cultures and crusts from days 14 and 28. The PCR products ligated with pGEM-T easy vector (Promega, Tokyo, Japan) were used to transform E. coli JM109 (Takara Bio Inc., Tokyo, Japan). Plasmids were extracted and purified using a GenElute plasmid miniprep kit (Sigma, Tokyo, Japan). Sequencing reactions were carried out using the BigDye Terminator v3.1 cycler sequencing kit (Applied Biosystems, Waltham, MA, USA). Nucleotide sequencing was performed using an Applied Biosystems 3130xl Genetic Analyzer. All sequences were checked for chimeric artefacts using CHIMERA_CHECK (Ribosomal Database Project). The obtained nucleotide sequences were compared with those deposited in the GenBank/EMBL/DDBJ databases using nucleotide BLAST, and were deposited in the GenBank/EMBL/DDBJ database under the accession numbers LC379189–LC379192 andLC460266–LC460268.
Results
Corrosion of carbon steel coupons by the H-1 enrichment culture
The H-1 enrichment culture was incubated for one month in the presence of carbon steel coupons. The time courses of carbon steel weight loss are shown in Figure 1. In the abiotic control, a continuous carbon steel weight loss was observed during incubation. The abiotic corrosion rate was calculated to be ca. 30μg-carbon steel day−1. In the H-1 enrichment culture, the weight loss of carbon steel gradually increased until day 10, similar to that in the abiotic control. After 10 days, the amount of weight loss of carbon steel in the H-1 enrichment culture was pronouncedly greater than that in the abiotic control. The biotic corrosion rate was calculated to be ca. 180 μg day−1 after 10-28 days. The MIC induced by H-1 enrichment culture proceeded six times more rapidly than that in the abiotic control after 10 days. A black layer was formed on the carbon steel surface in the H-1 enrichment culture after 10 days (Figure 2(a,b)), while less precipitate was observed in the abiotic control. Microscopic observation using laser microscopy revealed a small quantity of the attached product on the surface of carbon steel coupons after a 3-day immersion in the H-1 enrichment culture (Figure 2(c,e)). At 10 days, the amount of surface precipitate substantially increased with a layer of ca. 200 μm thickness heterogeneously covering the carbon steel coupons (Figure 2(d,f)). The increase in this thick layer and the weight loss of carbon steel after 10 days coincided. Many troughs were observed on the metal surface under the crust on day 28 by using scanning electron microscopy, but not in the abiotic control (Figure 2(g,h)). Hence, it was suggested that a relationship existed between the corrosion and crusts formation.
Time courses of the weight loss of carbon steel coupons with the H-1 enrichment culture (closed circles) and without the inoculum (closed squares). The data are presented as the means with standard deviations (SD) from triplicate experiments. Structure of carbon steel coupons after the corrosion experiment. (a) Carbon steel coupon before the experiment. (b) Carbon steel coupon after the corrosion experiment with the H-1 enrichment culture. (c–f) Laser microscopic analysis of the surface of a carbon steel coupon on day 3 (c,e) and 10 (d,f), with top view (c,d) and 3D view (e,f). Carbon steel coupon after incubation in the H-1 enrichment culture at 500-fold magnification (g) and one without inoculation at 200-fold magnification (h), after washing with Clark solution are shown.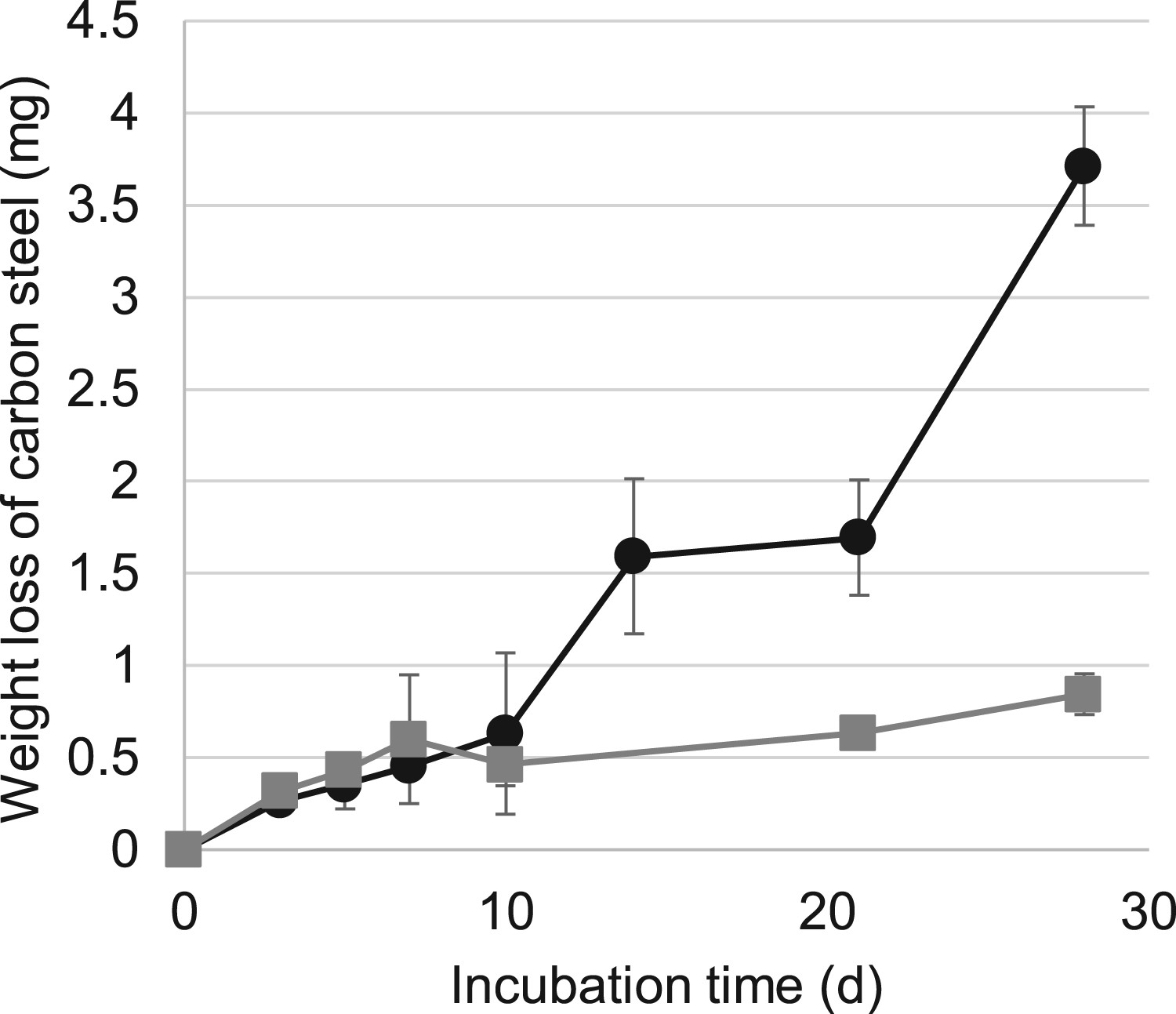

Changes of contents of metabolite in the course of the corrosion experiment
The time courses of changes of the major metabolite concentrations are shown in Figure 3. Sulphate ion contents in the H-1 enrichment culture gradually decreased from 35.4 to 29.1 mM until 14 days and became stable after 14 days (Figure 3(a)). H2S production by sulphate reduction increased from 3 to 10 days (200.8 µM in headspace) and slightly decreased after 10 days (Figure 3(d)). The colour of the culture turned to black, concurrently with the generation of H2S, due to the formation of FeS. In the abiotic control, sulphate ion contents remained roughly constant during the experiment. The contents of lactate in the H-1 enrichment culture decreased immediately and were completely consumed by day 7. Concurrently with a decrease in lactate, ∼40 mM acetate was accumulated on day 7 in the H-1 enrichment culture, and remained constant between days 7 and 14. After 14 days, acetate contents increased again, to ∼80 mM. In the abiotic control, the lactate and acetate contents changed slightly. Contents of CO2 and CH4 were constant in the abiotic control during the experiment, while that of H2 was slightly increased (Figure 3(c,d)). The CO2 and H2levels increased in the H-1 enrichment culture immediately after the cultivation began and peaked on day 7, after which they gradually decreased. Generation of CO2 and H2 accompanied the reduction in lactate and sulphate contents until day 7. After 7 days, simultaneously with the reduction in H2 and CO2 contents, CH4 production began.
Time courses of metabolite level changes with the H-1 enrichment culture (closed circles) or in media without inoculation (closed squares) for (a) sulphate, (b) lactate (solid line) and acetate (broken line). The produced gas levels are shown for (c) CO2, (d) H2 (solid line), CH4 (broken line) and H2S (dot line). The data are presented as the means with standard deviations (SD) from triplicate experiments.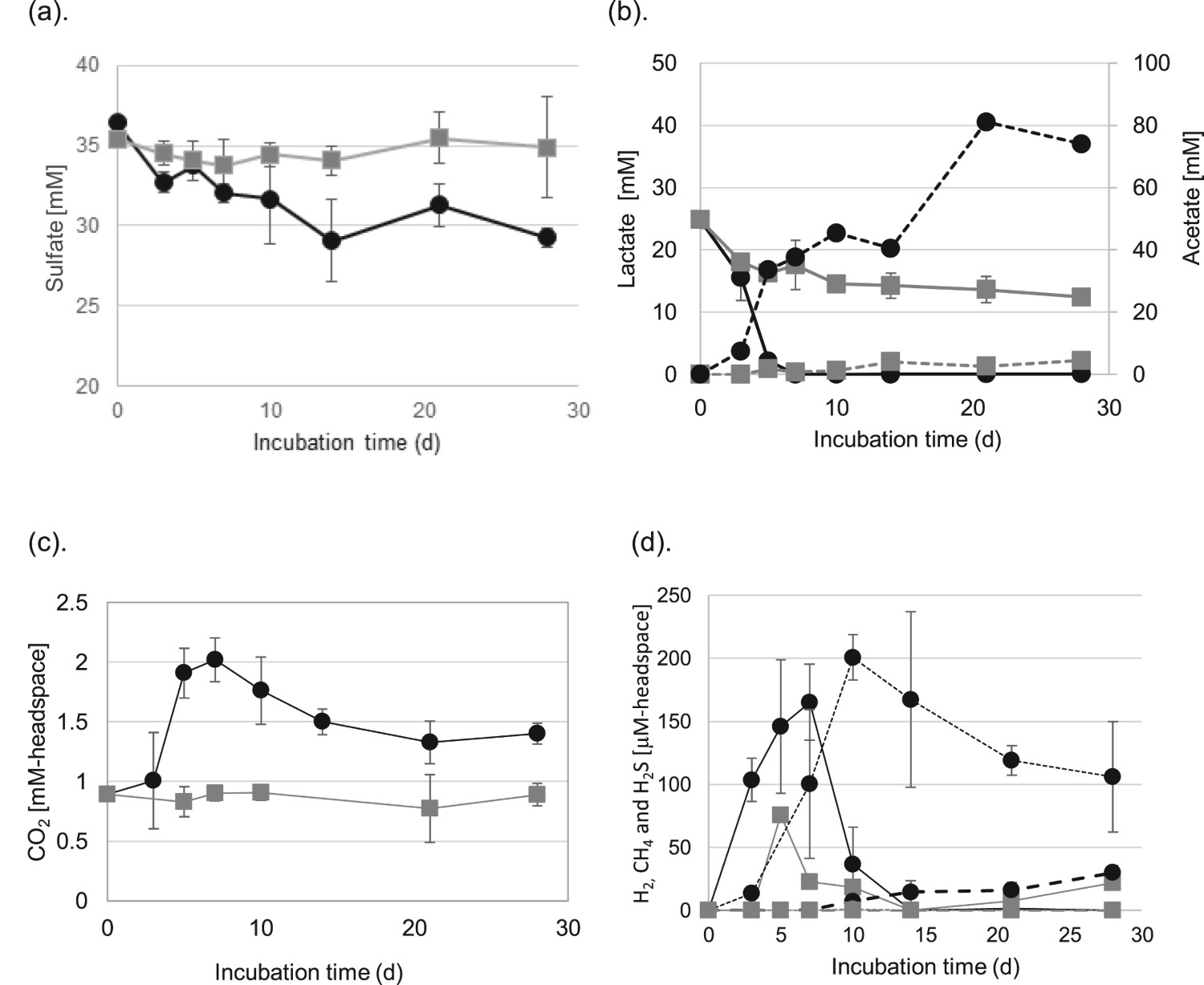
Microbial community in suspension cultures and crusts formed on carbon steel
The amounts of prokaryotic and methanogenic cells from the H-1 enrichment with and without carbon steel coupon were calculated by quantitative PCR. The abundance of prokaryotes was similar in suspensions of the two experiments collected on any sampling day (0.1-3.2 × 109 copies mL−1) (Figure 4). The amounts of methanogen were also similar in suspension samples of the two experiments until day 21 (0.5-6.9 × 106 copies mL−1), while that without a carbon steel coupon decreased more rather than those with a coupon on day 28 (p < 0.01). Large amounts of prokaryotes (0.9-2.9 × 109 copies cm−2) were contained on the crust attached to carbon steel surface after day 7 (Figure 4). The cell density of methanogen was low on day 7 (9.5 × 103 copies cm−2) and increased ∼100 times by day 21 (3.4 × 106 copies cm−2, p<0.005). The high concentration of microbial cells in the crust likely impacted microbial corrosion.
Cell density of prokaryotes and methanogens in corrosion tests. Time-course of cell density changes of prokaryotes (closed circles: with carbon steel; closed squares: without carbon steel) and methanogens (open circles: with carbon steel; open squares: without carbon steel) in suspension culture. Cell densities of prokaryotes (closed triangles with dashed line) and methanogens (open triangles with dashed line) in crust formed on carbon steel on days 7, 14, 21 and 28. The data are presented as averages from triplicate experiments.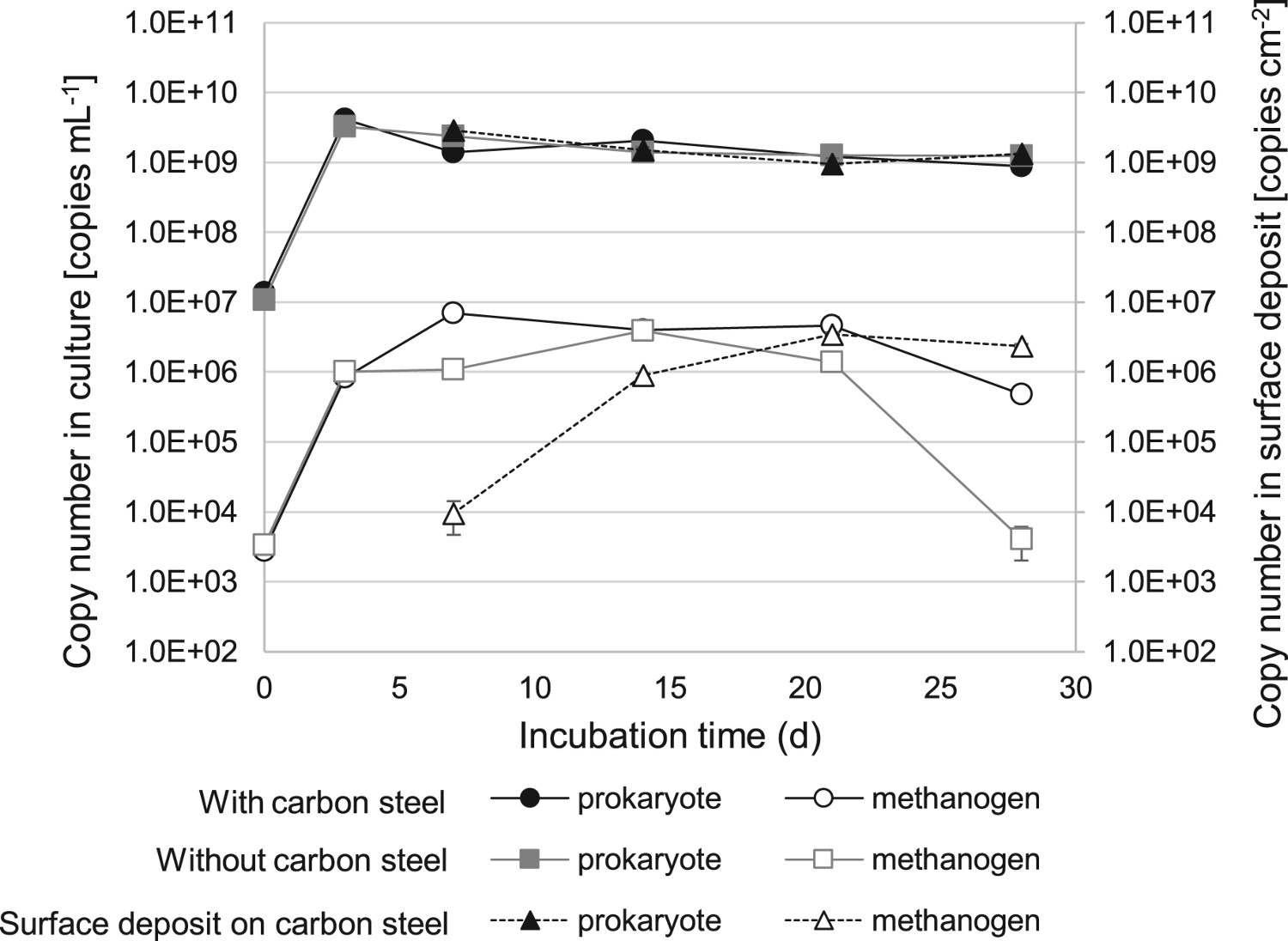
The 16S rRNA sequences V4 regions of prokaryotic and archaeal communities in suspension culture and in crusts were identified by high-throughput Illumina sequencing and Sanger sequencing method. At the genus level, Aminobacterium, Desulfovibrio, Clostridium, Sedimentibacter and Bacteroides were the core bacterial genera in all samples (Figure 5(a)). Microbial diversities in suspension and crust samples were similar during incubation time, respectively. However, different bacterial species dominated in the suspension and crust samples. In the majority of suspension samples in the presence of carbon steel, multiple unidentified genera belonging to class Clostridia and that to classes other than Clostridia in the phylum Firmicutes were accounting for 44.4-18.0% and 38.2-59.4%, respectively, in total. Sedimentibacter was the second-highest represented group, accounting for 7.3-19.9% of all genera. Bacterial communities in suspension in the absence of a carbon steel coupon showed similar diversity with those in the presence of carbon steel, except that the growth of Sedimentibacter was delayed (Figure 6). Although bacterial community in suspension with no carbon steel in day 28 showed much different about Clostridia than any others, the influence by the absence of carbon steel on Clostridia community is still unclear.
Microbial diversity in suspension cultures and in the attached fractions on carbon steel. (a) Relative abundance of the bacterial 16S rRNA gene sequences in suspension cultures (sampling at days 7, 10, 14, 21 and 28) and crusts on carbon steel (sampling at days 14, 21 and 28) presented at the phylum level. (b) Dominant methanogenic species in suspension cultures and crusts. *Bacteria belonging to Firmicutes, Clostridia other than Clostridium, but unidentified at genus level. **Bacteria belonging to Firmicutes other than Clostridia, but unidentified at class level. Microbial diversity in culture suspensions with (+) and without carbon steel coupon (−). Relative abundance of the bacterial 16S rRNA gene sequences in culture suspensions (sampling at days 7, 10, 14, 21 and 28) presented at the phylum level. *Bacteria belonging to Firmicutes, Clostridia other than Clostridium, but unidentified at genus level. **Bacteria belonging to Firmicutes other than Clostridia, but unidentified at class level.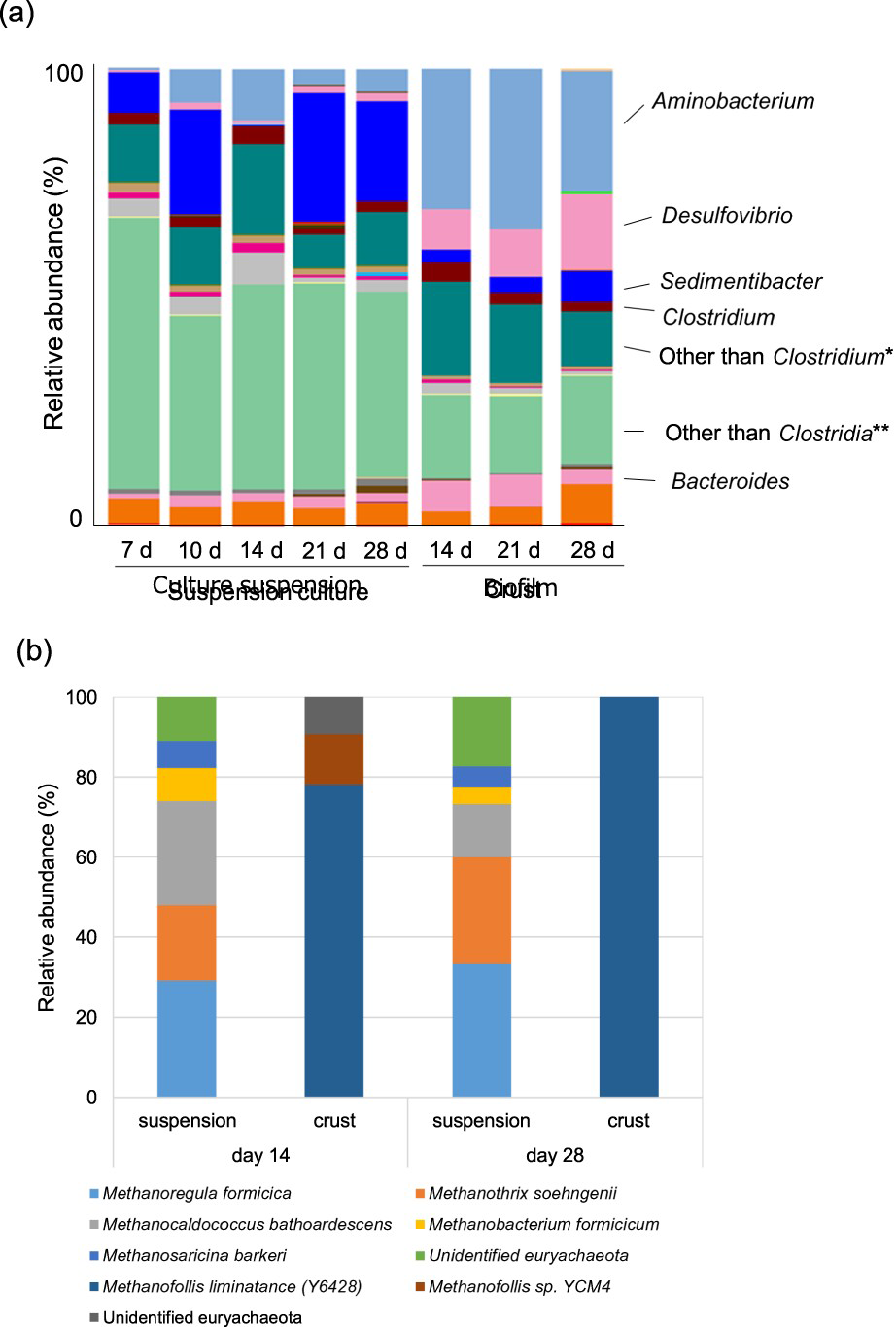
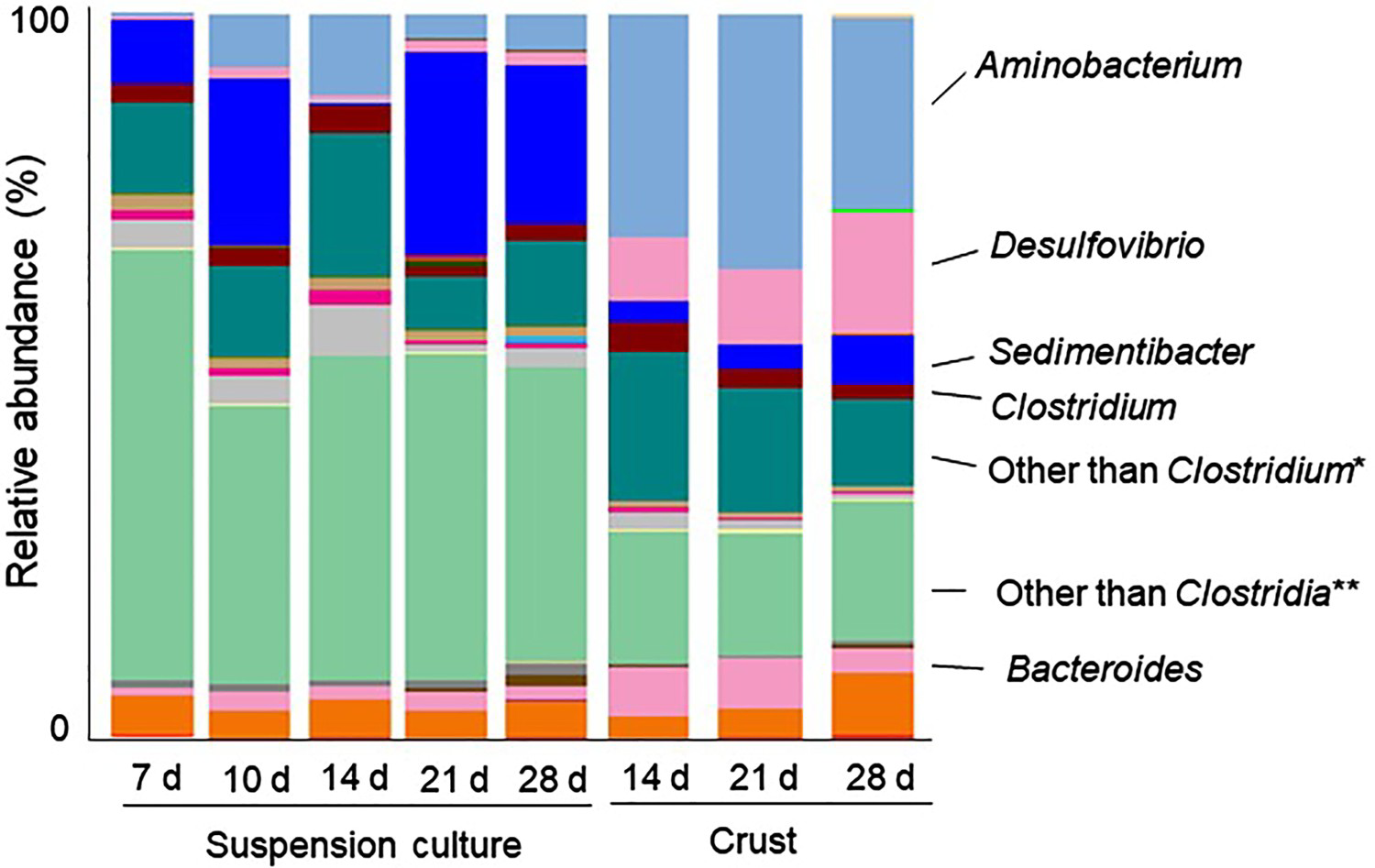
In crust samples, Aminobacterium was dominant, accounting for 26.3-35.2% of all genera, which was significantly higher than 0.5-11.1% in suspension culture. Moreover, the relative abundance of Desulfovibrio (Proteobacteria) in crusts was clearly higher than in suspension (8.7-16.9% of all genera in crusts, but less than 1% in suspensions). Desulfovibrio is commonly found in corrosion products developed on the surface of ship hulls, heat exchangers, wastewater pipelines, and oil pipeline and petroleum reservoirs, resulting in MIC [32–33].
An archaeal community change was not observed according to time-course after corrosion started (Figure 5(b)). Based on the Sanger analysis of 96 clones, the community from suspension samples contained three dominant methanogenic species: the hydrogenotrophic methanogen Methanoregula sp., Methanocaldococcus sp. and acetoclastic methanogen Methanothrix sp. Two minor species, related to Methanobacterium formicicum and Methanosarcina barkeri were detected in suspension cultures. In contrast, the crust sample community was dominated by a sole hydrogenotrophic methanogenic group of Methanofollis with 99% identity to M. liminatans (Y16428) at 28 days, while the minor group with 99% identity to Methanofollis sp. YCM4 was detected at 14 days (Figure 5(b)).
Discussion
The corrosion amount of carbon steel increased after crust formation at day 10 in the H-1 enrichment culture. The relative abundance of Aminobacterium, Desulfovibrio and Methanofollisin crust formed on carbon steel was specifically higher than that in the planktonic community after day 10. Close contact of bacterial cells and metal is an important factor affecting the corrosion rate [20,34]. These increased microbes in crust along with corrosion were estimated to contribute to the construction of the corrosive microbial community on carbon steel surface, and exerted a corrosive effect. We hypothesised the roles of these microbes on corrosion based on metabolite analysis. Fast-growing bacteria belonging to Firmicutes reduced sulphate using lactate as a carbon source in bulk medium. This reaction simultaneously produced metabolites of lactate (acetate, CO2, H2) and hydrogen sulphide inducing initial formation of a black layer surmised to be iron sulphides on carbon steel coupons until day 10. Firmicutes includes many sulphate-reducing bacteria group [35]. Detected H2S in the headspace (Fg3d) was lower than consumed sulphate ion by these microbes (Figure 3(a)). This is because measurement in headspace did not include soluble sulphides and consumed sulphide for the formation of iron sulphide. Although sulphate reduction and lactate degradation started by planktonic microbes from first few days, corrosion induction in H-1 enrichment culture was estimated to be delayed ∼10 days until corrosive bacteria increased and constructed community in black thick crust including iron sulphide on carbon steel. Acetate production was dominant process after day 10, while sulphate reduction and methanogenesis gradually increased. An increase in acetate and methane after day 7 indicated the decrease in CO2 and H2 in H-1 enrichment culture after day 7 was due to acetogenesis and methanogenesis, which produces acetate and methane using CO2 and H2 as substrate. In addition, acetate was estimated to be produced from a degradation of other organic component than lactate in VM-I medium (e.g. yeast extract and tryptone). Although microbial species affiliated with acetogen were not found in the microbial community in culture or in the crust on carbon steel, Aminobacterium can produce acetate as a product of amino acid degradation as shown in papers by Baena et al. [36] and Hamdi et al. [37].
The impact of SRB for carbon steel corrosion by H-1 enrichment cultures was shown by our previous research conducting a corrosion test under conditions inhibitory for each microbe [19]. Removal of sulphate ions from the medium reduced corrosive activity of the H-1 enrichment culture by 81.7% compared to the modified VM-I medium. Some have suggested that covering metal surfaces with corrosion crusts (such as iron sulphides) protects against further corrosion by limiting the accessibility of microbes and direct reaction of dissolved sulphide with carbon steel [38,39]; however, crust formation was followed by higher corrosion amount in the current study. Corrosive SRBs, Desulfovibrio, have an ability to induce corrosion even in a thick layer of corrosion products by the uptake of electrons from iron through semiconductive corrosion products such as iron sulphides, after corrosion products covered on iron surfaces [5,38]. In the current study, Desulfovibrio dominated in crusts but not in the cultures after all of the lactate was consumed, there are possibility that Desulfovbrio used other organic compounds than lactate and carbon steel coupon as an electron source in the crust. Therefore, it is essential to analyse corrosion influence by Desulfovibrio in the crust using the electrochemical method.
In addition to Desulfovibrio, Aminobacterium was suspected of involvement in MIC because of its high relative abundance. Acetate production after lactate was consumed out was estimated to be based on other organic carbon including mainly amino acids in a medium such as yeast extract and tryptone. It has been reported that Aminobacterium degrades a wider range of amino acids in the presence of hydrogenotrophic methanogen Methanobacterium formicicum than in its absence, and the accumulation of acetate and CH4 was increased compared to that in a pure culture of Aminobacterium [36,37]. In contrast, it has been reported that the presence of acetate shortened the lag-time for Methanofollis growth, while acetate was not required for growth [40]. Acetate produced by Aminobacterium might enhance the predominance of Methanofollis among hydrogenotrophic methanogens in the crust. It was estimated that Aminobacterium and Methanofollis influenced each other in the crust on carbon steel. Acids produced by microbes increase the corrosion rate by decreasing the local pH and removing the oxide layer from the metal surface [41,42]. The corrosion rate of carbon steel in an acetate-containing medium is related to the concentration of acetic acid. The distribution between acetic acid and acetate depends on pH. In the current study, pH of the medium dropped from 7.0 to 5.5-6.0 after day 28. Under those conditions, suspension at day 28 contained acetate at high concentration (80 mM), while a small fraction (<10%) of the acetate will persist as the protonated acid [43]. Therefore, it was estimated that acetate produced by Aminobacterium sp. influenced on carbon steel corrosion, while relative contribution among the other corrosion mechanisms is necessary to be clarified in future studies.
Conclusion
Changes of metabolites and microbial communities in H-1 enrichment culture with the progress of carbon steel corrosion were analysed over time in the blackish environment under anaerobic condition. These analyses revealed that thick black crust surmised to include iron sulphide was formed on carbon steel and significant corrosion was induced after day 10. Next-generation sequencing and qPCR clarified that a distinct microbial community from that in culture medium was constructed on carbon steel. Moreover, these analyses in the current study suggest that Desulfovibrio sp., Aminobacterium sp. and Methanofollis sp. locally enriched on carbon steel, and their involvement to carbon steel corrosion. However, the roles and interaction of these microbes in carbon steel corrosion are still unclear. Future studies should delineate the corrosion-associated mechanism based on interactions between these microbes in crusts, which could allow for avoidance of MIC on pipelines under brackish environments.