
Abstract
Three studies used a computer video game preparation to demonstrate latent inhibition in adult humans. In all studies participants fired torpedoes at a target spaceship by clicking the mouse. Conditioned stimuli (CSs) were presented in the form of coloured “sensors” at the bottom of the screen. Conditioning was conducted by pairing a sensor with an attack from the target spaceship. Participants learned to suppress their rate of mouse clicking in preparation for an attack. In Experiment 1 a total of 10 preexposures to the sensor CS, prior to conditioning, retarded acquisition of suppression. In Experiment 2 the effect of preexposure was shown to be context specific. Experiment 3 showed little generalization of the preexposure effect from one sensor CS to another. Experiment 3 also showed that preexposure did not make the sensor CS inhibitory. Comparisons with conditioned suppression procedures with animals and negative-priming procedures are briefly discussed.
Latent inhibition, also known as a “CS preexposure” effect, is said to occur when presentations of a putative conditioned stimulus (CS), prior to being paired with an unconditioned stimulus (US), retard acquisition of conditioned responding (e.g., Bouton & Rosas, 1997; Lubow, 1973). The effect has been important in the area of learning, arguably, in that the challenges it posed for the Rescorla–Wagner (Rescorla & Wagner, 1972) theory of conditioning helped to extend theories of learning into the area of attention (e.g., Lubow, 1989; Mackintosh, 1975; Pearce & Hall, 1980). Latent inhibition, or rather its near absence, has also been widely connected to schizophrenia (e.g., Braunstein-Bercovitz & Lubow, 1998; Lubow & Gewirtz, 1995) and perhaps holds promise as an empirical base from which to expand the application of learning theory towards the understanding of psychological dysfunction.
Many theories (e.g., Lubow, 1989; Mackintosh, 1975; Pearce & Hall, 1980) suggest that the acquisition deficit observed in latent inhibition is a true learning deficit caused by a reduction in the attention or associability of the CS during preexposure, perhaps due to learning about the absence of outcomes (Pearce & Hall, 1980) or by associating the CS with the context in which it occurs (e.g., McLaren & Mackintosh, 2000; Wagner, 1981). Others (e.g., Bouton, 1993; Kraemer & Roberts, 1984) have suggested that latent inhibition is better characterized as a performance deficit. In this view latent inhibition represents a failure to retrieve learning that was otherwise well established during conditioning.
Support for a retrieval view comes largely from studies on the contextual control of latent inhibition. One well-established property of latent inhibition is that it is reduced by a change of physical context between exposure and conditioning (e.g., Gray et al., 2001; Hall & Channel, 1985; Swartzentruber & Bouton, 1992; Westbrook, Jones, Bailey, & Harris, 2000). When the context of preexposure (e.g., “A” or “B”) is the same as that where conditioning and testing occur (i.e., “AAA”, where each letter represents the preexposure, conditioning, and test phases, respectively), latent inhibition is observed. When the context of preexposure is different from that of conditioning and testing (BAA), conditioned responding is acquired more rapidly, suggesting some loss of latent inhibition with a context change. Moreover, when the context is changed between conditioning and testing (AAB), conditioned responding is lost (i.e., latent inhibition may be recovered; see Westbrook et al., 2000, for a demonstration and review).
The debate regarding the mechanisms of latent inhibition is far from settled, and though of theoretical importance, the present paper remains neutral with respect to these issues. Rather, it focuses on demonstrating the phenomena in adult humans, which is important in that the effect has been viewed as difficult to achieve with humans. Indeed, there are few solid examples where participants receive simple presentations of a stimulus that result in the retarded acquisition of a response when those presentations are later followed by pairings with another stimulus. Additional clear demonstrations with a variety of preparations are important to establish the generality of the effect and to establish a variety of tools with which it can be investigated in humans. This point carries additional significance in that demonstrations of what has been called latent inhibition involving “masking tasks” have been challenged as perhaps demonstrating a different phenomenon, referred to as negative priming (see Graham & McLaren, 1998).
Very thorough and clear reviews of the literature with humans exist elsewhere (see Lubow, 1973; Lubow & Gewirtz, 1995; more recently, Escobar, Arcediano, & Miller, 2003), and only the most important points are reiterated here. From these reviews it is clear that the most reliable methods by which to obtain a latent inhibition effect in adult humans are tasks that require some sort of additional masking task during preexposure (but see Escobar et al., 2003). To justify this, some authors (e.g., Braunstein-Bercovitz, Hen, & Lubow, 2004) adhere to the theoretical position that latent inhibition is the result of reduced attention to the CS resulting in an impairment in its ability to enter into associations. They assume that latent inhibition requires initial attention to a stimulus that declines as a function of preexposure and that an experimental setting places demand characteristics on adult humans, causing their attention to to-be-conditioned stimuli to be continually maintained (see Graham & McLaren, 1998, for a similar discussion). According to this interpretation, demonstration of latent inhibition should require that the stimulus be presented in a manner whereby attention will not be maintained, and this is often assumed to occur with the use of an additional masking task.
In a masking procedure, presentations of the to-be-conditioned stimulus are typically embedded in some other task to which the participant is attending. For example, De la Casa and Lubow (2001) used the following protocol: During a preexposure phase participants were told to choose, over a series of trials, which of six letters on a screen was replaced by a black rectangle. A participant might view the letters A through to F arranged in a 2 × 3 grid with one of the letters covered by a black square. They were to pick quickly the letter on the keyboard that corresponded to the letter that was covered. The cue being exposed in this phase was a colour presented as a background on which the 2 × 3 grid of letters was presented. Thus, the colour in the background was being exposed, but it was also irrelevant to the task at hand. In the second phase, participants were instructed to press the letter that would be covered before it was covered. During this phase, coloured rectangles appeared, which were correlated with the letter that would be covered so that participants could learn to predict the covered letter based on the colour of the rectangle. After sufficient preexposure (30 trials in this study), it took participants longer to pick the letter associated with a preexposed colour than one associated with a colour that had not been preexposed.
The present studies demonstrate what is best characterized to be a latent inhibition effect in a video game procedure that may present a more naturalistic situation for participants and does not involve explicit masking. In a method similar to that developed by Arcediano, Ortega, and Matute (1996) a baseline of behaviour is obtained with participants playing a first-person video game (see Figure 1), and subsequent pairings of events lead to response suppression. In the game, a baseline of mouse-clicking is established by having participants fire torpedoes at a spaceship. On this baseline of mouse clicking, CSs in the form of coloured “sensors” appear, and USs are presented in the form of an inescapable attack by the onscreen spaceship. The attack damages the participant's spaceship for a particular amount of time. During this time the mouse is inoperable, mouse movements are not reflected on the screen, and clicking has no effect, producing an instant suppression of mouse clicking.
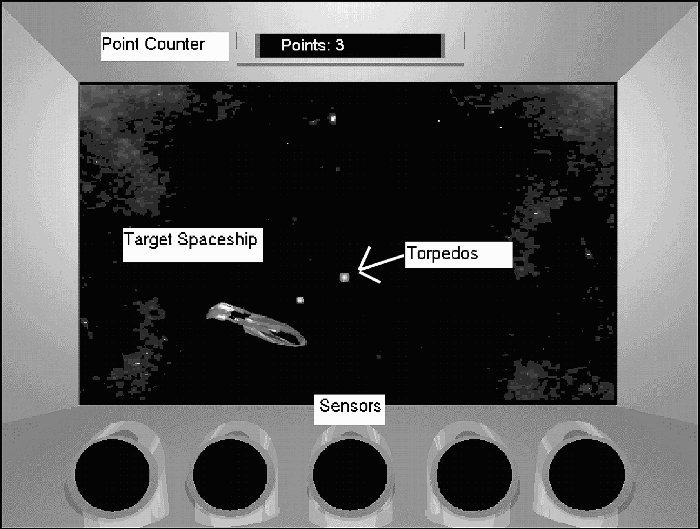
Video game screen where participants fire torpedoes at the target spaceship. Sensors are paired with attacks from the target spaceship. Participants learn to suppress their own torpedo firing to prepare for attacks. See text for details.
The cover story given at the beginning tells participants that an attack will drain their ship's power if they are not prepared for it. They are instructed that to prepare for an attack they need to conserve energy by suppressing their own rate of torpedo firing at the appropriate time. They are instructed that their sensors might help them, but they are not told exactly how the sensor will help, nor what a sensor will mean. The participants cannot escape an attack predicted by a sensor, but they can defensively modify their behaviour to better prepare for it. Through pairings of the sensor with attacks participants learn to expect an attack and to suppress their mouse clicking during the sensor CS to prepare for its effects. A latent inhibition manipulation simply involves presenting the sensor CS several times in the absence of any consequence.
In the first study presented here presentations of a sensor in the absence of an attack attenuated the rate at which suppression elicited by that sensor was acquired when it was later paired with an attack. In the second study, it was demonstrated that the preexposure effect observed in the first study was context specific, a finding that is consistent with a latent inhibition interpretation. When exposure to the sensor and subsequent pairings of it with an attack took place in different contexts, provided by different “space” backgrounds, suppression was acquired more rapidly. The final study replicated aspects of both studies and showed that the preexposure procedure did not give the CS conditioned-inhibitory properties and that the preexposure effect did not generalize substantially between stimuli.
Experiment 1
The first study manipulated preexposure to a sensor CS prior to conditioning. Two groups were trained to click the mouse and to fire torpedoes at the onscreen spaceship. For group red exposed, a red sensor was presented 10 times during this time, for 5 seconds on each presentation with an intertrial interval average of 10 seconds (CS offset to CS onset). No other events occurred during this phase. A control group, group context exposed, played the game for the same amount of time, but no sensors were presented. Without interruption or noticeable transition, conditioning began for both groups. Here the red sensor was presented and was paired with the attack US over the next 10 trials. A latent inhibition effect would appear as slower acquisition of suppression in group red exposed than in group context exposed.
Pilot work showed that simple conditioning occurred quickly with this method, often reaching an asymptote within six trials. Given the rapid conditioning that occurred, 10 trials were chosen for the preexposure phase, assuming that such preexposure would be sufficient to allow for the operation of any latent inhibition mechanism.
Method
Participants
Participants were 24 volunteers from a university in the southern United States. Participants were predominantly between the ages of 18 and 21 years, with approximately 65% being female. Some participants volunteered for course or extra credit at the discretion of individual instructors. In this and all subsequent studies, when credit for participation was involved alternative activities to earn the credit were available. All participants signed informed-consent forms, which informed them that the study involved factors that might affect learning. No further details (beyond the instructions below) and no hypotheses were presented.
Apparatus
The experiment used a video game running on IBM-compatible computers with a monitor resolution of 800 × 600 pixels and 24-bit colour. Computers were equipped with standard two-button mice.
In the game an image (see Figure 1) was presented on a 38.1-cm (15-in.) computer monitor, which represented a viewscreen inside of a spaceship through which stars and another spaceship could appear. The viewscreen was a rectangular window 618 pixels wide and 368 pixels tall (23.5 × 14 cm) centred from left to right and 3.5 cm below the top of the screen on a grey metallic background. A box appeared 1.3 cm below the top of the screen in which the word “Points” appeared in yellow. The box was 1 cm tall and 8 cm wide. At the bottom of the screen five black ovals appeared that were each 3.28 cm in diameter. The third was centred from left to right and was located 0.8 cm above the bottom of the screen. The other four ovals were spaced at intervals of approximately 2 cm to the left and right of the centre oval.
Coloured backgrounds were presented in the viewscreen, consisting of stars and nebulae. In the present studies two backgrounds were used. One background was a Hubble space telescope photograph of the Crab Nebula, which presented a predominately red background with red stars. Another background was a Hubble photograph of the Eagle 1 Nebula, which presented a primarily blue and yellow background. In Experiments 1 and 3, the Eagle 1 Nebula background was used. Both (counterbalanced) were used in Experiment 2.
On these backgrounds a spaceship, approximately 4 × 1.9 cm, was present, flying in a path randomly determined at run time. The spaceship was a three-dimensional rendering of a “Romulan Warbird” from the “Star Trek: The Next Generation” television series (trademark of Paramount Pictures). Every second the ship turned left, right, up, or down, or some combination of those directions. The direction of the turn was determined randomly, with the probability of a left or right turn being .5, and the probability of an up or down turn being independent of left and right and also occurring with a probability of .5. The degrees of each turn were determined randomly with a minimum of 9 and a maximum of 180 degrees. Turns occurred on the screen in a smooth angle-by-angle rotation with a turn of 180 degrees taking approximately 2.5 s to complete. When the spaceship went off the viewscreen it was programmed to turn around and fly back into view.
Conditioned stimuli were presented as the 5-s illumination of one or more (see Experiment 3) ovals at the bottom of the screen with the following red–blue–green (RGB) colour combinations: Red consisted of 100% red, zero green, and zero blue; green (used in Experiment 3) consisted of 100% green, zero red, and zero blue. All stimuli were presented in the centre (third) position unless otherwise noted.
The US was presented in the form of an attack from the enemy spacecraft. A round green circle emerged from the rear of the spaceship and tracked to the centre of the viewscreen, growing larger with each step until it reached a size of approximately 5 cm where it exploded in a large fire-ball explosion. The time that elapsed from the emergence of the torpedo to the end of the explosion was less than 2 s and varied slightly as a function of the distance of the spaceship from the centre of the viewscreen. With the termination of the explosion, a message appeared on the screen, “Power at___percent. Controls Frozen for____seconds”, with the corresponding percentages and seconds appearing in the blanks. These numbers counted up to 100% and down to 0 s, second by second. The numbers that appeared in these blanks were determined by counting the number of times the participant clicked the mouse during the 5-s CS and dividing that number by the average number of times they clicked the mouse during the 5-s periods prior to all the CSs they had so far experienced, plus the number of clicks made in the CS—that is, CS clicks/(average pre + CS clicks). This modified suppression ratio was then multiplied by 120 to determine the number of seconds displayed in the message. If a participant clicked the mouse on average 10 times during the pre-CS period and clicked the mouse once during the CS, the ratio would compute to .09. Thus, the message appeared for 11 s. The number initially representing the percentage of power remaining was calculated by subtracting the modified suppression ratio from 1 and multiplying by 100. Each second, the seconds count was decremented by 1, and the power percentage was updated by the following equation: 98 – (seconds/120 * 100 + .5). When seconds turned to zero, the message disappeared, and the mouse became functional again. On trials where participants completely suppressed mouse clicking, power was indicated to be 100%, number of seconds of frozen controls was zero, and the message lasted for 1 s.
When participants fired torpedoes a small pink circle approximately 2 cm in diameter travelled from the centre top or bottom of the screen (depending on which was closest to the position of the mouse pointer) and travelled to the point on the screen to where the mouse was pointing at the time it was clicked. As it travelled, it decreased in size to approximately 1 cm to simulate the effect of distance. When a torpedo “tracked” the spaceship it flew to the point where the mouse was pointing at the time the mouse was clicked and then followed the spaceship until it impacted and exploded in a fire-ball explosion. Only the torpedo exploded. The spaceship was never destroyed, maintaining continuity across the game. Tracking lasted a variable amount of time depending on the distance of the torpedo to the spaceship, but never lasted more than 2 s.
Procedure
Participants were randomly assigned to groups red exposed or context exposed. There were initially 12 participants in each condition.
Participants from different experimental groups were run at the same time, and up to 10 participants could be run at once, although they were predominantly run in groups of 5 or fewer. When all participants had been seated, they were instructed to read silently the following instructions. Figure 1 was present at the top of the instructions. Items in bold below were presented in bold.
You are taking a “Space Academy” systems-proficiency test for promotion. You will be engaged in fictitious combat with an enemy spacecraft. Every time you shoot the enemy spacecraft with your Photon Torpedoes you will gain a point. When the enemy spacecraft shoots you, your ship's power will be drained and your controls will be frozen while the power is recharging. You will be unable to gain points during this time. Earning the most points in the least amount of time will earn a promotion to Captain. To earn the most points in the class you must learn how to use the sensors in your spaceship.
The more you fire, the more points you will earn.
Clicking the standard left mouse button will send power to your torpedo system.
When enough power is sent (usually one to five clicks) a torpedo will fire. The torpedo may “track” the enemy and hit, or it may miss altogether.
The enemy will attack you with its own homing torpedo.
An enemy homing torpedo will drain your power, freezing your controls until the system is recharged. You will be unable to gain points while your controls are frozen.
The time that your controls are frozen will make the game that much longer. (Remember, you want to earn the most points in the least amount of time!)
This part is very important. Over the course of the test, different colors will appear in your sensors. Particular colors or combinations might mean anything. Keep in mind also that they may mean nothing at all!! Pay close attention. Subtle changes may have important meanings.
Learning what sensors mean will help you avoid frozen controls. When you are about to be attacked, reduce your rate of firing. This reduction in firing will let you conserve power and avoid having your controls frozen for long periods of time. The more power conserved, the less damaged you may be.
Summary
Then, the attending researcher read the instructions aloud with the participants. Participants were instructed to place their right hand on the mouse and their left hand on the “s” key of the keyboard. The lights were turned out and participants were instructed to press the “s” key starting the program.
Baseline
Throughout the game participants clicked the left mouse button on a variable ratio (VR) 3 schedule where a random one in three clicks fired a torpedo and a random half of those torpedoes tracked the spaceship and exploded on it. When a torpedo exploded, a point was added to the point counter window at the top of the screen (the ratio between clicking and earning a point was, therefore, VR6). All participants were allowed to play for 60 s before any stimuli were presented.
Preexposure
Group red exposed then received trials where the middle sensor oval was illuminated with red, 10 times, for 5 s on each presentation. The time between presentations was randomly determined and averaged 10 s. The group receiving exposure to only the context simply played the game for the same amount of time without any CS presentations. A black sensor CS was programmed to appear in the sensor oval (which was already black) for 5 s, functionally producing no change in the display and ensuring that this phase was of identical length for the different groups.
Conditioning
Without any apparent transition, conditioning began. Both groups received 10 trials where the red CS appeared and was paired with an attack from the spaceship. The intertrial interval (ITI), US offset to CS onset, on these trials averaged 10 s.
Data analysis
Data consisted of the number of times the participant clicked the mouse during each 5-s CS and during the 5 s immediately before the CS for each trial during conditioning. These data were analysed separately with analysis of variance (ANOVA). Standard suppression ratios, CS clicks/(pre-CS + CS clicks), were also calculated and were analysed with ANOVA. Note that the modified suppression ratios used in the procedure were not used for data analysis. Pairwise comparisons were conducted with analysis of variance with error terms pooled from the overall analysis using the standard methods discussed by Howell (1987). Degrees of freedom were reduced using the Welch–Satterthwaite procedure to compensate for the pooling of potentially heterogeneous sources of variance. Two-tailed probabilities of results are reported for the reader, although a rejection criterion of p < .05 was adopted throughout. The data from Experiment 1 are presented in more detail than are those in the other studies, including separate analyses and presentations of the suppression ratio data, the responding in the CS, and data from the pre-CS period, to provide a full characterization of the data that resulted from this method. The remaining experiments contain analyses of only the suppression ratio data and the pre-CS data.
Results and discussion
Exclusion criterion
With suppression ratio data, participants sometimes had a pre-CS rate of zero on particular trials, making it difficult to interpret the corresponding suppression ratio, and these participants were eliminated from analyses. Inspection of their data showed that these participants were low responders on all trials. A selection criterion of an average rate of clicking during the pre-CS of at least one click per second across the preexposure phase was adopted. All participants responding, on average, less than one click per second were eliminated from analysis. The criteria of one click per second was adopted because with a 5-sec CS, measurement of suppression becomes very coarse with slower rates, and this criterion succeeded in eliminating all participants that had zero pre-CS rates on some trials.
There were initially 12 participants in each group. Application of the exclusion criterion led to elimination of 3 participants from group red exposed and 2 participants from group context exposed. Exclusion was not related to group, χ2 < 1.
Suppression ratio data
Figure 2 top shows the suppression ratio data during conditioning for groups red exposed and context exposed. An Exposure (red or context) × Trials ANOVA showed an effect of trials, F(9, 153) = 8.43, p < .001, and exposure, F(1, 17) = 19.26, p < .001, but the overall interaction was not reliable, F(9, 153) = 1.04, p = .41. Nevertheless, subsequent simple effect tests showed that the groups did not differ on the first trial, F(1, 89) = 1.02, p = .31, but did differ reliably on all other trials, Fs(1, 89) ≥ 5.38, ps ≤ .022, except Trial 9, F(1, 89) = 2.75, p = .1.
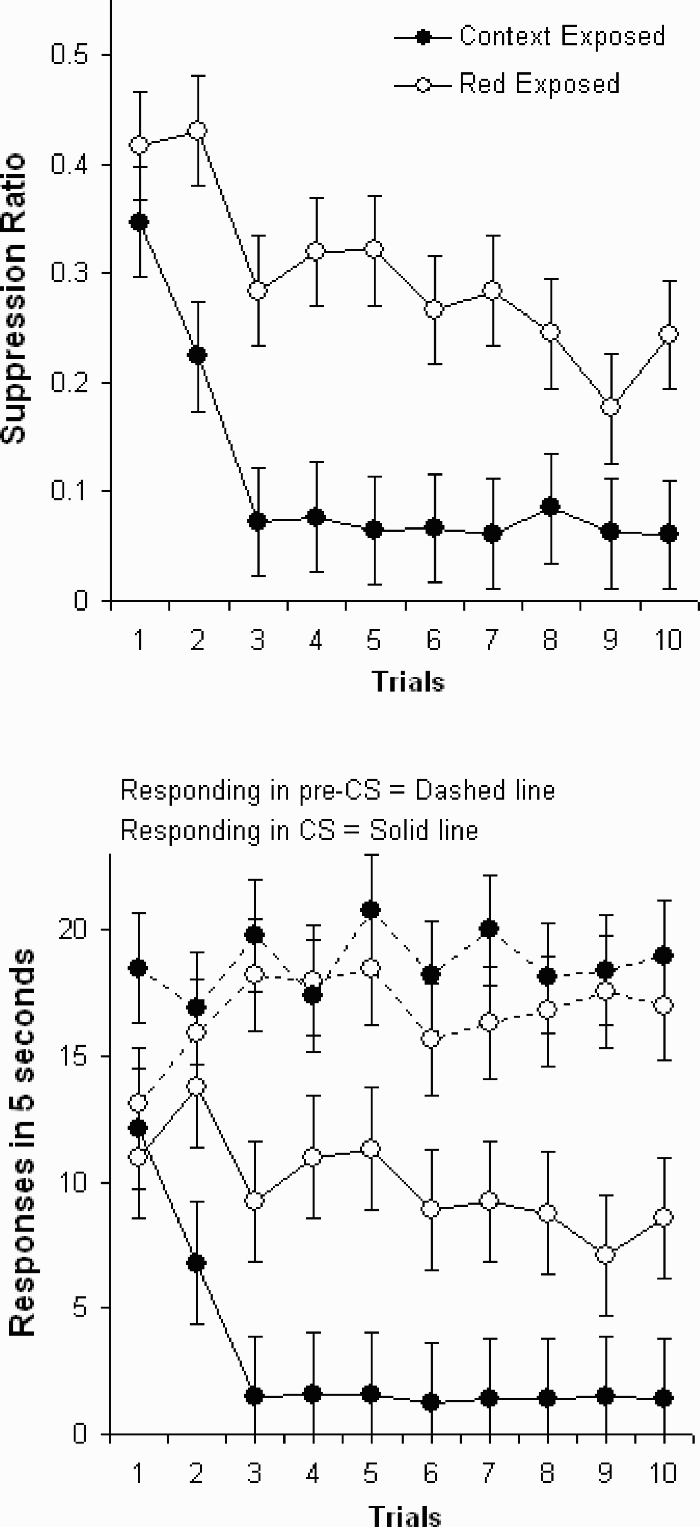
Data from the conditioning phase after participants received preexposure to the red sensor CS used in conditioning or the context alone in Experiment 1. The top panel shows suppression ratio data; the bottom panel shows the absolute responding during the sensor CS and the pre-CS. Error bars represent the standard error of the mean.
Some conditioning occurred between the first two trials in group context exposed, but not in group red exposed. The difference between Trials 1 and 2 was significant in group context exposed, t(89) = 3.15, p = .002, but not in group red exposed, t < 1. The difference between Trials 1 and 2 was larger in group context exposed than in group red exposed, F(1, 153) = 5.83, p = .017.
Responding in the CS
Responding in the CS and pre-CS during conditioning is shown in the bottom panel of Figure 2. An Exposure × Trials ANOVA of the responding in the CS showed an effect of trials, F(9, 153) = 5.45, p < .001, exposure, F(1, 17) = 7.67, p = .013, and an interaction, F(9, 153) = 2.08, p = .034. Simple effect tests (all Fs on 1 and 44 degrees of freedom) showed that the groups did not differ on the first trial, F < 1, but did differ on every other trial, Fs ≥ 4.85, ps ≤ .03, except Trial 9, F = 3.13, p = .08.
The difference between Trials 1 and 2 was significant in group context exposed, t(89) = 3.68, p < .001. The difference approached significance in group red exposed, t(89) = 1.82, p = .069, but reflected, if anything, an increase in responding. The difference between Trials 1 and 2 was larger in group context exposed than in group red exposed, F(1, 153) = 14.88, p < .001.
Responding in the pre-CS
An Exposure × Trials ANOVA showed an effect of trials, F(9, 153) = 2.27, p = .02. Pre-CS responding generally increased from an initial level of 15.95 to 18.05 across the 10 conditioning trials. There were no other reliable effects in the analysis, Fs ≤ 1.17, ps ≥ .31. Simple effect tests confirmed the overall analysis. The groups did not differ on any trials, Fs(1, 28) ≤ 3.21, ps ≥ .08 (Trial 1).
Analysis of both the responding in the CS and the suppression ratio data supports the same conclusions. After 10 preexposures to the CS used in conditioning, acquisition of suppression was considerably slower than that after exposure to the context.
Experiment 2
The second experiment was designed to assess whether the latent inhibition effect demonstrated in the first study would be affected by a context change. The study employed a 2 × 2 between-subjects factorial design manipulating latent inhibition by whether or not the stimulus was preexposed and manipulating whether conditioning took place in the same or different context from preexposure. It was expected that acquisition of suppression would be retarded following CS preexposure when preexposure and conditioning took place in the same context and less so when they took place in different contexts.
The contexts were provided by different space backgrounds visible on the screen. A context switch was initiated by a message box appearing on the screen that informed the participant that they, their sensors, and the enemy spacecraft were being transported to another galaxy for further testing. It was made explicit to the participant that they were still using the same ship and sensors and that they were engaged with the same enemy spacecraft. These instructions were designed to minimize the possibility that the participants simply perceived the sensors and spaceship as different stimuli. In the absence of these instructions participants might have believed the sensor to belong to a different set, or the enemy spacecraft to be a different ship, and have been learning something about novel stimuli as opposed to learning about stimuli that they had encountered before.
In the study, participants began by playing the game for one minute in context different to ensure familiarity with that context, followed by a preexposure phase in context same. In the preexposure phase, half of the participants received 10 exposures to the CS to be used in conditioning while the other half simply played the game without exposures for the same amount of time. Conditioning was conducted with half of the participants in each preexposure condition in context same and the other half in context different.
Method
Participants
Participants consisted of 44 students from a university in the southern United States and 40 students from a university in the south of Spain. The characteristics of the participants and the experimenter interactions with them were otherwise the same as those in the previous study.
Apparatus
The same apparatus as that used in the previous experiment was used here. All words present in the instructions and game were presented in Spanish for those participating in Spain.
Procedure
All participants within each country were randomly assigned to conditions exposed same, exposed different, novel same, or novel different. All participants completed informed consent forms and were given the instructions in the same way as in the previous experiments.
Participants played the game for 60 s with no stimulus presentations in context different (either the Crab Nebula or Eagle 1 Nebula background, counterbalanced). A context switch was then made with the appearance of a standard Microsoft Windows message box in the middle of the screen. The text, “Please attend to this important message”, was displayed on the title bar of the message box, and the text in the message box read: “You, your sensors, and the enemy are being transported to another galaxy for further testing. Press OK to proceed.” When the participant pressed the “OK” button, another message box was displayed. The text, “Press OK now for immediate transport”, was displayed in the title bar, and the text in the message box read: “Remember, you, your sensors, and the enemy are being transported to another galaxy.” When the participant pressed the “OK” button the screen flickered, and the current background (e.g., Crab Nebula) was replaced with the alternative (e.g., Eagle 1 Nebula).
The portion of the screen upon which the sensor was presented did not change; only the image seen through the part of the screen corresponding to the viewscreen changed. The sensor was sufficiently far from the viewscreen (see again, Figure 1) so that its colour was not affected by changes in the viewscreen background. There was no perceptible change in the appearance of the sensor between the two different backgrounds present in the viewscreen—an issue of some importance (see Gray et al., 2001, for discussion).
After the initial exposure to context different, participants in the groups receiving preexposure received 10 exposures to the red sensor CS in the same manner as did participants in group red exposed in the previous study, in context same. Participants that received no exposure played the game for the same amount of time as did the participants in the context exposed conditions in the previous study, again in context same. After the preexposure phase, a context switch was initiated for the participants that would receive conditioning in a context different from that used for preexposure. No transition occurred for participants receiving conditioning in the same context as preexposure. Conditioning was conducted exactly as in the previous experiment.
Results and discussion
The data from Experiment 2 are shown in Figure 3, which is arranged to show the effect of context in each of the conditions where the CS was preexposed (left-hand display) or not (right-hand display).
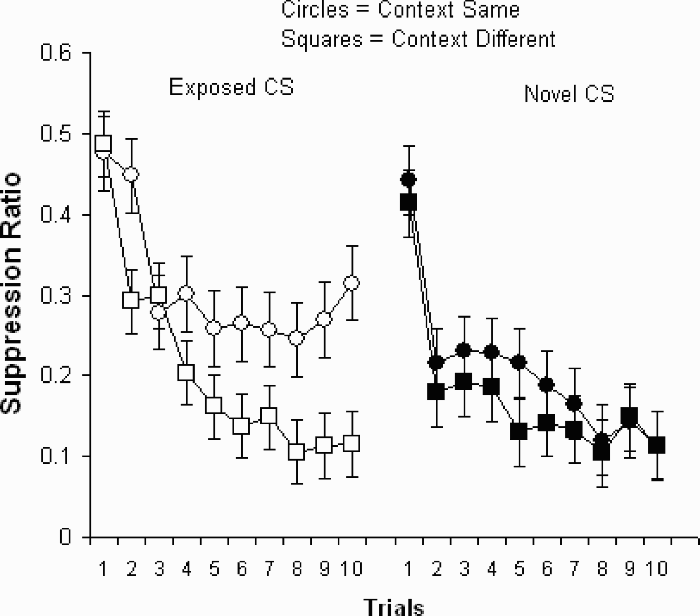
Data from Experiment 2. Participants were preexposed, or not, to a red sensor CS and subsequently conditioned in either the same or different context as preexposure. Left-hand points show the effect of a context shift on the preexposed sensor CS, and the right-hand points show the effect of a context shift on the novel sensor CSs. Error bars represent the standard error of the mean.
The same screening criterion as that used in the previous study (pre-CS < 1 per second) was used here, resulting in the exclusion of 17 of the participants. The majority of the excluded participants were Spanish (13 of the 17), and exclusion was not independent of the country of origin, χ2(1) = 7.91, p = .005. The larger portion of Spanish participants excluded due to low response rates may be due to a misinterpretation of the instructions for some participants. Observation during participation was consistent with the idea that many participants believed that the mouse pointer (in the shape of a plus-shaped crosshair) had to be over the onscreen spaceship (a difficult task) before clicking. Despite the higher number eliminated, exclusion was not related to group assignment within either the Spanish participants, χ2(3) = 1.25, p = .74, or the American participants, χ2(3) = 4.95, p = .18. Combined, there was still no relationship between group membership and selection, χ2(3) = 2.27, p = .52.
The suppression ratio data from conditioning were analysed with an initial Trials × Latent Inhibition (LI; exposed CS or novel CS) × Context (same or different from preexposure) by Country ANOVA. No effects involving Country were reliable in this analysis nor in the same analysis applied to pre-CS responding, Fs ≤ 1.81, ps ≥ .08. The country variable was subsequently ignored. There was an effect of trials, F(9, 558) = 27.05, p < .001, LI, F(1, 62) = 4.82, p = .03, and context, F(1, 62) = 4.25, p = .043. The overall LI × Context interaction was not significant, F(1, 62) = 1.78, p = .187. The Trials × LI × Context interaction approached the typical level of significance, F(9, 558) = 1.78, p = .081. No other effects were reliable, ps > .153.
Because of our a priori interests in both the LI × Context and the LI × Context × Trials three-way interaction (which approached the .05 level of reliability) we conducted analyses to fully assess a potential interaction by investigating the effect of LI within each of the same and different contexts and the effect of context on both the exposed and the novel CSs. Both sets of analyses supported the same conclusions: Preexposure was most effective when the preexposure and conditioning contexts were the same, and a change of context only affected the rate of conditioning to the preexposed CS. For brevity, only the analyses of the effect of context are reported below.
Effect of context
Effect of context on conditioning of a preexposed CS
A Trials × Context ANOVA on responding to the preexposed CS (left side of Figure 3) showed effects of trials, F(9, 558) = 15.54, p < .001, context, F(1, 62) = 5.67, p = .03, and a Trials × Context interaction, F(9, 558) = 2.06, p = .03. Pairwise comparisons (all Fs on 1 and 247 degrees of freedom) showed differences on Trials 2 (F = 5.94, p = .015), 6 (F = 3.91, p = .049), 8 (F = 4.74, p = .03), 9 (F = 5.88, p = .015), and 10 (F = 9.59, p = .002). There were no other differences, Fs ≤ 2.83, ps ≥ .094.
Effect of context on conditioning of a novel CS
A Trials × Context ANOVA on responding to the novel CS (right set of points on the left side of Figure 3) showed an effect of trials, F(9, 558) = 15.12, p < .001. The effect of context and the interaction were not reliable, Fs < 1. Pairwise comparisons corroborated this analysis showing no differences on any trial, Fs(1, 247) ≤ 2, ps ≥ .158.
Pre-CS response rates
Responding during the pre-CS averaged 15.65 (SD = 6.05). Parallel analyses of pre-CS responding to those outlined above produced no reliable effects, ps ≥ .08.
Experiment 2 replicated the effect of preexposure seen in Experiment 1. Exposure to the sensor CS prior to pairings with an attack reduced the rate at which participants learned to suppress their torpedo firing when preexposure and conditioning took place in the same context. When preexposure and conditioning took place in different contexts no measurable latent inhibition effect was observed. The context change increased the rate of acquisition to the preexposed CS but did not affect the rate of acquisition to the CS that was not preexposed.
Experiment 3
The third study assessed the extent to which the effect of preexposure generalized to another stimulus and tested the reasonable assumption that the present procedure might make the preexposed CS a conditioned inhibitor, as did Escobar et al. (2003). Although the US had not been experienced at the time of preexposure, it is possible that the instructions generated an expectation of being attacked, which did not occur in the preexposure phase. Such a condition could make the preexposed sensor an inhibitor for attack.
There were five groups in this study, and the design is shown in Table 1. The first three groups were designed to replicate and extend Experiment 1 with full counterbalancing between red and green sensor CSs as Sensors A and B. In one group Sensor A was preexposed and conditioned. In another, Sensor B was preexposed while Sensor A was conditioned. In the third condition, only the context was exposed, and conditioning took place with Sensor A. These groups should allow for any generalization of the preexposure effect to be observed.
Design of Experiment 3
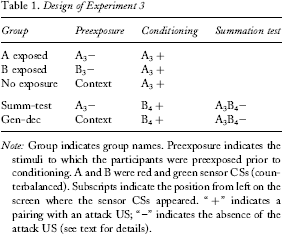
Note: Group indicates group names. Preexposure indicates the stimuli to which the participants were preexposed prior to conditioning. A and B were red and green sensor CSs (counterbalanced). Subscripts indicate the position from left on the screen where the sensor CSs appeared. “ + ” indicates a pairing with an attack US; “–” indicates the absence of the attack US (see text for details).
Two other groups were run to assess the possibility that the stimuli used in preexposure could become inhibitory. In group summ-test, participants received preexposure to A located in the third sensor position as was done in the other groups and previous studies. Conditioning was conducted with B located in the fourth sensor position from the left. Four summation tests were conducted in extinction at the end of the conditioning phase where the sensor used in conditioning was illuminated in the fourth position, and the preexposed sensor was illuminated in the third position. If the preexposed sensor has inhibitory properties, then there should be less suppression to the compound in this group than in a corresponding control. The control condition, group gen-dec, was a group in which no sensor was preexposed, and conditioning was conducted with B in the fourth position. Thus, Sensor A's presence on the summation test was novel, and its influence would represent only the effect of a generalization decrement.
These last two groups also allow us to further assess the degree to which the effects of stimulus preexposure generalize between the stimuli used here. Recall that in the first group described above preexposure and conditioning were with the same colour sensor. Retarded acquisition of suppression was expected. With the second condition, preexposure and conditioning were with different coloured sensors, and retarded acquisition of suppression was expected only to the extent that generalization between these stimuli occurs. In group summ-test the difference between the stimulus of preexposure and conditioning was further enhanced. Preexposure was with one colour in one position, and conditioning was with a different colour located in a different position, thus the stimuli varied by more than one dimension. The positions of the sensors were not counterbalanced, so as to allow a completely untarnished view of the impact of changing position, but, as seen later, analysis of the results shows that this is likely to not be an issue.
Method
Participants
Participants were college students from a college in the south-east United States with the same characteristics as those in the previous studies, and all interactions with the attending researcher were likewise the same.
Apparatus
The same apparatus as that used in the previous studies was used here.
Procedure
Preexposure
Participants were randomly assigned to five conditions. In groups A exposed, B exposed, and summ-test, participants received 10 preexposures to either the red or the green (counterbalanced) sensor CS in the third position, as in the previous studies. Participants in groups no exposure and gen-dec simply played the game for the same amount of time with no stimulus exposures.
Conditioning
Following preexposure, participants in group A exposed received conditioning as in the previous studies where the sensor used in preexposure was paired with an attack over 10 trials. Participants in group B exposed received conditioning with the sensor not used in preexposure. Participants in group sum-test and gen-dec received pairings of the colour not preexposed with the attack, but the colour was presented in the fourth sensor oval from the left instead of the third.
Summation test
Following conditioning, participants in groups sum-test and gen-dec received four trials where the sensor used in conditioning was illuminated, and the sensor colour not used in conditioning was illuminated in the third position. For participants in group sum-test, this was the sensor presentation used in preexposure. For the participants in group gen-dec, the sensor presentation was novel.
Data analysis
Data were analysed as in the previous study. In this study, data from the preexposure phase were also collected and analysed.
Results and discussion
Data are shown in Figure 4. The points on the left represent conditioning of A in groups A exposed, B exposed, and no exposure. The points in the middle represent conditioning in groups summ-test and gen-dec. Group summ-test received exposure to A and conditioning with B in a different position. Group gen-dec received conditioning with B in the fourth position without exposure to A. The points on the right represent suppression on the summation test where both Stimulus A and Stimulus B were present, in groups summ-test and gen-dec.
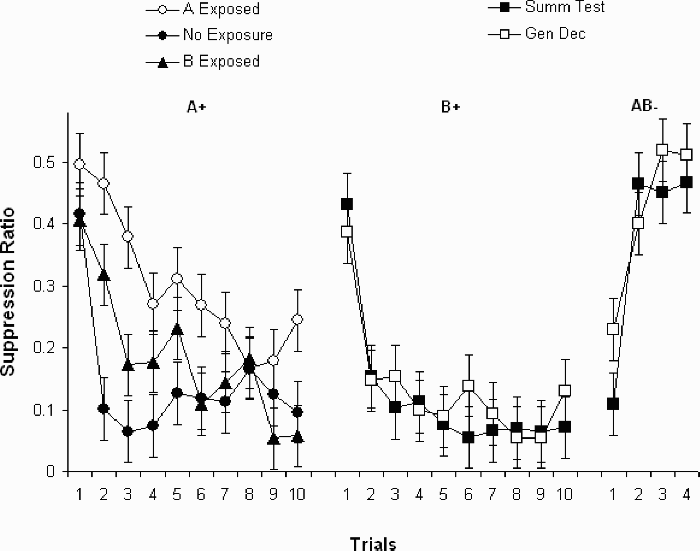
Participants received preexposure, or not, to Sensor A or B in the third sensor position (see Figure 1) prior to conditioning with A in the third position (left-hand points) or B in the fourth position (middle points). Participants conditioned with B received a summation test (final four right-hand points) where A was presented in the third position in compound with B in the fourth (see text for details). Error bars represent the standard error of the mean.
There were 8 participants in each condition except for groups A exposed, B exposed, and no exposure, which had 9. Half of the participants in each group received conditioning with the green CS and the other half with the red CS, except in groups A exposed and no exposure, in which 5 participants received conditioning with the red CS, and group B exposed, in which 5 participants received conditioning with the green CS. The same selection criterion as that used in the previous studies was applied here, resulting in the exclusion of 2 participants in the no exposure condition who received conditioning with the green CS. Exclusion was not related to group membership, χ2 < 1.
Preexposure
A Group × Trials analysis of the suppression ratios during preexposure showed no effects involving trials. Neither the effect of trials nor its interaction with group were significant, Fs < 1. The effect of group was not reliable, F(4, 36) = 1.45, p = .24. Responding during trials where a sensor CS was preexposed (groups A exposed, B exposed, summ-test) did not differ from responding during the same period of time where no CS was present (groups no exposure and gen-dec). Presenting the sensor CS neither augmented, nor unconditionally suppressed, pre-CS responding at any point in the preexposure phase. Suppression ratios averaged .48 during this phase. The same analysis of pre-CS responding showed only an effect of trials, F(9, 324) = 3.85, p < .001, but no other effects, Fs ≤ 1.27. Pre-CS responding averaged 14.14 (SD = 7.03) at the beginning of preexposure and averaged 17.88 (SD = 5.12) at the end.
Conditioning
Conditioning occurred most slowly in group A exposed, followed by group B exposed, and then the other conditions, which did not differ from each other. A Group × Trials × Counterbalance group (conditioning with red or green) ANOVA of the 10 conditioning trials showed an effect of trials, F(9, 279) = 28.17, p < .001, group, F(4, 31) = 5.88, p = .001, and a Group × Trials interaction, F(36, 279) = 1.63, p = .016. There was no effect of counterbalance group, F < 1, and it did not interact in any way with the other factors, Fs ≤ 1.21, ps ≥ .44. The counterbalance variable was subsequently ignored.
None of the groups differed from each other on the first conditioning trial, Fs < 2.45, ps > .11. Among the groups that received no preexposure (no exposure and gen-dec) simple effect tests on the remaining trials (all Fs on 1 and 153 degrees of freedom) showed no differences, all Fs < 1. Groups receiving no preexposure, followed by conditioning with the CS in either the third or fourth position, were indistinguishable.
Group A exposed showed the slowest conditioning and differed from group no exposure on Trials 2 through to 6, and 10, Fs ≥ 4.52, ps ≤ .03. Group A exposed differed from group B exposed on Trials 2, 3, 6, and 10, Fs > 4.43, ps ≤ .037. Group A exposed differed from group sum-test, which was preexposed to one colour in the third position and conditioned with another in the fourth position, on Trials 2 through to 7, and 10, Fs ≥ 6.17, ps ≤ .01. Exposure to the sensor CS used in conditioning attenuated acquisition of suppression more so than exposure to another sensor CS in either the same or a different position.
Group B exposed differed from group no exposure on Trial 2, F = 9.19, p = .003, but not on any other trial, Fs ≤ 2.25, ps ≤ .14. Group B exposed also differed from group sum-test on Trials 2 and 5, F ≥ 4.74, p < .03, but not on any other trial, Fs ≤ 2.51, ps ≥ .11. Within these two groups different colours were used for preexposure and conditioning, but between these two groups the same colours were used in preexposure and conditioning, with conditioning taking place in different positions between the groups. Finally, group gen-dec did not differ from group summ-test on any trial, Fs ≤ 1.36, ps ≥ .24. Exposure to a sensor CS different from that used in conditioning attenuated acquisition of suppression to a small degree, evident on one trial only (Trial 2). When both the colour and the position of the sensor was changed, no attenuated response suppression was observed.
The same analysis as that applied to the suppression ratio data was applied to the pre-CS response rates. That analysis showed only an effect of trials, F(9, 279) = 2.16, p = .025, as pre-CS response rates increased from an initial mean of 16.18 (SD = 5.12) at the start of conditioning to 18.90 (SD = 6.90) on the final trial. There were no other reliable effects, Fs ≤ 1.94, ps ≥ .127.
Summation test
A Group (sum-test vs. gen-dec) × Trials ANOVA was applied to the four summation test trials. There was an effect of trials, F(3, 42) = 27.33, p < .001, but no effect of group, F(1, 14) = 1.89, p = .29, and no interaction, F(3, 42) = 1.79, p = .16. The trend for suppression in group gen-dec to be less than that in sum-test observable on the first trial was only a trend; it was not significant, F(1, 51) = 3.60, p = .064. The same analysis of pre-CS responding showed no significant effects, all Fs ≤ 2.34, ps ≥ .15. Pre-CS response rates averaged 19.67 (SD = 5.64) in this phase.
Experiment 3 replicated the basic effects shown throughout this paper. Preexposure to the CS used in conditioning retarded acquisition of suppression more so than did exposure to a different-coloured CS in the same position (group B exposed). Exposure to a different-colour CS produced a small attenuation effect (group B exposed) when compared to a no-exposure control or to exposure to a different-colour CS conditioned in a different position (group summ-test). The latter condition was not different from no-exposure controls (groups no exposure and gen-dec).
The results produced no evidence that the preexposure treatment caused the stimuli to become inhibitory, which is consistent with a latent inhibition interpretation of these results (Reiss & Wagner, 1972; Rescorla, 1971). When the preexposed sensor was compounded with the one used in excitatory conditioning, the resulting suppression was no different to the suppression that would be expected on the basis of generalization decrement. Of course, the lack of a difference is a null result and thus should be viewed with caution. Given the null result, it is worth noting that the lack of an effect was observed across a range of responding, so any possible effect was not likely to be occluded by floor or ceiling effects. The resulting trend evident on Trial 1, although not statistically reliable at the traditional .05 level of significance, was in the opposite direction to a result favouring an interpretation in terms of conditioned inhibition.
General discussion
The present studies were conducted to assess whether preexposure to stimuli prior to conditioning would result in a latent inhibition effect in adult human participants in a simple video game method. Beginning with Experiment 1, and across all three studies, preexposure to a stimulus that was subsequently used in conditioning produced retarded acquisition of responding. The effect was clearly evident on a trial-by-trial basis, reflecting that there was indeed a reduced rate of acquisition.
Experiments 2 and 3 confirmed that the preexposed stimulus had the additional properties of a latent inhibitor. In Experiment 2 the effect was shown to be context specific (e.g., Escobar et al., 2003; Hall & Channel, 1985; Killcross & Dickinson, 1996), as would be expected on the basis of latent inhibition. Experiment 3 showed that the treatment did not endow the stimulus with measurable conditioned inhibitory properties. The preexposed CS did not affect responding elicited by another CS paired with the same US. Experiment 3 also showed that that there was little generalization of the preexposure effect between similar sensor CSs.
It has been suggested that many demonstrations of latent inhibition in human participants, those using masking tasks, might perhaps be demonstrations of negative priming (Graham & McLaren, 1998). Thus, a consideration of that possibility for the present procedures and results is warranted.
In a negative priming task participants are typically presented with a complex stimulus (e.g., De la Casa & Lubow, 2001; Leboe, Leboe, & Milliken, 2003; Tipper, Weaver, Cameron, Brehaut, & Baestedo, 1991) and are required to attend (respond) to one part of the stimulus while ignoring the other. Subsequently, their ability to respond to the once-ignored stimulus is attenuated. The characteristics of negative priming are thus similar to latent inhibition. The characteristics differ in that negative priming is said to last over long intervals, even months in some cases (DeSchepper & Treisman, 1996), while latent inhibition has been shown to dissipate with the passage of time (e.g., Aguado, Symonds, & Hall, 1994; Rosas & Bouton, 1997; but see De la Casa & Lubow, 2002). Negative priming is said to generalize well to other stimuli, even dissimilar stimuli that belong to the same conceptual category (e.g., Graham & McLaren, 1998; Tipper, 1985). Latent inhibition, on the other hand, generalizes to a much more limited degree (Lubow, 1973) as was observed in Experiment 3.
Simple procedural issues make it unlikely that the results of the present study were due to negative priming. Masking was involved, arguably, only to the same extent that it is present in animal preparations (see Gray et al., 2001, for similar reasoning). The procedure was designed to be directly analogous to a latent inhibition procedure where CSs are preexposed and conditioned while rats are lever pressing for food or water (e.g., Rodríguez & Alonso, 2004). In such paradigms, rats are engaged in reinforced lever pressing when presentations of a CS occur. Pairings of the CS with an aversive US, such as shock, lead to the CS eliciting defensive responses, which reduces the rate of lever pressing during the CS. In the present procedure it was necessary that participants be engaged in a task (mouse clicking) during both preexposure and conditioning when the CSs appeared, but they were explicitly instructed to attend to these CSs. Ignoring the stimuli was not necessary for optimal performance of the baseline task as is typically the case with negative priming. This point is further reinforced by the analysis of the preexposure phase in Experiment 3. Presentations of the stimuli in preexposure did not impact responding on the baseline task.
The observation that the presentation of the sensor CSs did not influence baseline responding, either initially or at the end of preexposure, also bears on another possible interpretation of the effect observed here. As the sensors were 5 s in duration, and most participants could click at a rate of three clicks per second, five torpedos could easily be launched during this time. Half of those would result in earning points, so the preexposure procedure led to the coincidental pairings of the stimulus with earning points, though not at a rate above that in the absence of the stimuli. Nevertheless, suppression of mouse clicking in conditioning could be construed as a form of counterconditioning in the groups where the CS was preexposed. However, this interpretation is unlikely. If coincidental pairings of the CS with points scored did lead to conditioning, then an increase in the baseline response rate should have occurred, at least during the later stages of the preexposure phase when the CS was presented. The thorough analysis of preexposure in Experiment 3 showed that this was not the case. This issue has also been raised with respect to the conditioned suppression technique with animals (Joseph, 1995) and has been found to be an ineffective explanation (Killcross, Dickinson, & Robbins, 1995).
While these studies do not provide additional information that helps to differentiate between explanatory mechanisms of the effects of preexposure, the present method is one in which it is possible to easily and robustly demonstrate a preexposure effect with the known properties of latent inhibition. The method is sufficiently different from negative priming tasks as to be an unlikely demonstration of such an effect, and cannot be easily attributed to conditioned inhibition or coincidental counterconditioning during preexposure.