
Abstract
Traditionally, movement kinematics are thought to reflect physical properties (e.g., position and time) of movement targets. However, targets may also evoke intentional goals like “to be in a certain position at a given time”. Therefore, kinematics may be viewed not as a reaction to stimuli, but rather as the means to attain intended goals. In the present study participants performed continuous reversal movements. It was first shown that kinematics towards temporal and spatial targets differ from kinematics away from those targets. Further, kinematics are different for movements to temporal (relatively short movement times, high and late peak velocity) and spatial (relatively long movement times, early peak velocity) targets (Experiments 1 and 2). In order to obtain evidence for the influence of goal representations on kinematics, combinations of temporal and spatial targets were investigated in Experiments 3 and 4. Specifically, the conditions were: spatial targets always present with varying temporal targets, temporal targets always present with varying spatial targets, and combined and separate spatial and temporal targets. Not only the physical features, but also how the targets were represented as movement goals, were important. Thus, movement kinematics do not simply reflect stimulus properties, but rather the representation of the intended goal.
To interact with the environment, a person often needs to direct movements towards certain positions at a given time, as, for example, when catching a ball. Thus, movements towards targets have to be specified according to the temporal dimension (“when” to be somewhere) and the spatial dimension (“where” to be). Traditionally, movement kinematics are thought to reflect physical properties (e.g., position and time) of those targets. However, there may be an alternative view to this. Targets may indeed not only represent stimuli in the environment to which a person “reacts” by using certain movement kinematics. Rather they may evoke intentional goals like “to be in a certain position at a given time”. Therefore, movements are not a reaction to stimuli, but are the means to attain intended goals (ideomotor principle, see James 1890/1981; Prinz, 1997). According to the ideomotor principle, intending and anticipating an action/movement effect is the first step in performing an action or movement. The aim of the present study is to provide evidence for this view on movement control, by showing that observed movement kinematics cannot always be explained by the combination of target stimuli alone. If target stimuli given in the environment are not sufficient to explain observed movement kinematics, it can be assumed that some kind of goal representation influences movement kinematics. In the following the term target is used to describe physical properties of experimental situations. The term goal is used to reflect participants’ representation of that target combined with the intention to be at the target.
A wide variety of studies indicate that movement targets/goals play an important role in determining the way movements are planned and/or executed (Aschersleben, Drewing, & Stenneken, 2002; Diedrichsen, Ivry, Hazeltine, Kennerley, & Cohen, 2003; Helsen, Elliott, Starkes, & Ricker, 2000; Ivry, Diedrichsen, Spencer, Hazeltine, & Semjen, 2004; Lackner & Dizio, 1994; Schiegg, Deubel, & Schneider, 2003; Semjen, 2002; Vos, Mates, & van Kruysbergen, 1995; Weigelt, Rieger, Mechsner, & Prinz, in press). In the following some of the results from those studies are described. Then the background for the questions of the present study is developed in more detail.
Several studies indicate the importance of spatial targets for movement organization. There is evidence that bimanual movements to similar targets are initiated faster than bimanual movements to dissimilar targets (Diedrichsen et al., 2003; Weigelt et al., in press). In addition, people recall limb positions better than limb movements (Smyth, 1984). Further, during preparation of a grasping movement, attention is mainly directed to those parts of an object that are to be grasped (Schiegg et al., 2003). In discrete aiming movements with predictable target locations, movement endpoints are continuously monitored (Lackner & Dizio, 1994). In addition, the eyes usually fixate the target location before a hand movement is initiated (Carnahan & Marteniuk, 1991, 1994; Helsen et al., 2000), regardless of the distance covered (Helsen, Starkes, & Buekers, 1997). The hand moves into central vision as it approaches the target; this is probably the reason why late visual information appears to be most important for limb control (Carlton, 1981; Chua & Elliott, 1993). There has been a considerable debate over which features are coded in sensory space and then transformed into corresponding parameters in motor space (Feldman, 1986; Imanaka & Abernethy, 1992; Khan, Hale, Garry, & Franks, 2002; Polit & Bizzi, 1987; Rosenbaum, Loukopoulos, Meulenbroek, Vaughan, & Engelbrecht, 1995). It is now recognized that participants code final positions in space when performing tasks in which distances have to be reproduced (Khan et al., 2002), although the theoretical accounts of the processes that lead to this effect differ. Endpoint coding could be related to neuromuscular factors as outlined in the equilibrium-point hypothesis (Feldman, 1986; Khan et al., 2002; Polit & Bizzi, 1987). However, it could also originate at a more central level, for example as abstract spatial codes in short-term memory (Imanaka & Abernethy, 1992) or stored postures in memory (Rosenbaum et al., 1995). In movements to spatial targets asymmetric velocity profiles are frequently observed (Hogan, 1984; Hogan & Flash, 1987). The asymmetry of velocity profiles increases when accuracy demands of the movement are high, and/or targets are small (Hogan & Flash, 1987; Langolf, Chaffin, & Foulke, 1976; MacKenzie, Marteniuk, Dugas, Liske, & Eickmeier, 1987; Soechting, 1984; for an overview, see Elliott, Helsen, & Chua, 2001).
Not only spatial targets but also temporal targets seem to play an important role for movement planning and execution. For instance, a large amount of research on tapping has shown that participants plan their movements in order to achieve temporal goals (e.g., Aschersleben et al., 2002; Drewing & Aschersleben, 2003; Vos et al., 1995). Theoretically, it is assumed that the internal clock does not trigger tapping movements, but provides target time points at which movements have to produce their meaningful effects—that is, contact with the response key (Billon, Semjen, & Stelmach, 1996). Temporal targets of a task can be described in a so-called “event structure account” (Ivry et al., 2004). Temporal constraints reflect general limitations in the ability to represent temporal relationships, as, for example, in bimanual coordination (Ivry et al., 2004). A similar phenomenon is described by the term “anchoring” (Jirsa, Fink, Foo, & Kelso, 2000). Experiments have shown that movements are more temporally and spatially consistent at points where the movement is linked with another event—for example, a metronome (Byblow, Carson, & Goodman, 1994; Kelso, Buchanan, & Wallace, 1991). There is less temporal variability at reversal points that coincide with a metronome beat than at other reversals. Anchored reversal points also generally display lower spatial variation than do unanchored reversal points within a frequency plateau, and they exhibit less of a shift in mean position when frequency is altered (Fink, Foo, Jirsa, & Kelso, 2000). Concerning kinematics, participants make movements of shorter duration towards temporal targets and longer movements in the return phase in tapping, regardless of whether the timing target is with the flexion or extension movement (Balasubramaniam, Wing, & Daffertshofer, 2004). Further, the interval preceding a forceful tap in a sequence of normal taps is shortened, and the one immediately following a forceful tap is lengthened (Billon et al., 1996). These studies involving temporal targets suggest that movements to salient or stressed events are performed faster than less salient or unstressed events.
Thus far, there is evidence in the literature that spatial and temporal targets play an important role in planning as well as overall organization of movements. However, most of the time comparisons have been made with different specifications of one target type (i.e., spatial targets of different difficulty, temporal targets that are stressed to different degrees). To the best of my knowledge, kinematics of target-directed movements have not been compared to kinematics of non-target-directed movements. One methodological problem is to make movements towards targets comparable to movements away from targets. For example, if one investigates discrete reaching movements towards spatial targets, one encounters several difficulties. Movements towards target locations are usually not comparable to those going away from the target (back to the starting location)—they differ in their time constraints, and in most studies they also involve different areas of workspace (there might, for example, be differences in moving away from the body compared with towards the body). In addition, differences in movement kinematics may also arise from the fact that a target-directed movement is followed by another movement (back to the starting position), whereas the movement towards the starting position is usually not followed by another movement (at least not immediately). In order to avoid some of the above-mentioned problems with discrete target-directed movements, reversal movements were chosen in the following experiments (in the horizontal plane—that is, movements along the medial-lateral axis).
The above-mentioned studies suggest that movements to spatial and temporal targets may differ in the way that they are controlled and consequently in their kinematics. However, such differences have sometimes been attributed to other factors. Carlton (1994) found that movements under temporal accuracy instructions were characterized by a single submovement, with peak velocity occurring at about 50% of the total movement time, whereas movements under time-minimization instructions tended to be made up of two submovements, with peak velocity occurring at about 40% of the total movement time. Carlton attributed the differences in the observed kinematics to the different type of instructions that he used. However, one can also argue that temporal accuracy instructions require the participants to attend to temporal targets, whereas the time-minimization task required the participants to attend to a spatial target. In fact, there is evidence that temporally and spatially constraint movements are controlled in different ways (Meyer, Abrams, Kornblum, Wright, & Smith, 1988; Zelaznik, Mone, McCabe, & Thaman, 1988). In particular, results indicate that movements with temporal precision requirements show a linear speed–accuracy relationship whereas movements with spatial precision requirements show a logarithmic speed–accuracy relationship (Fitts’ law). Most importantly, Zelaznik et al. (1988) showed that temporal precision requirements and not movement duration were the main factor influencing the speed–accuracy trade-off. One can therefore predict that movements towards temporal targets have a relatively late peak velocity, whereas movements towards spatial targets have a relatively early peak velocity.
In the first two experiments of the present study movements towards temporal (Experiment 1) and spatial (Experiment 2) targets were investigated with two aims: first, to see whether the different kinematic patterns observed in the above-mentioned studies for the respective types of movement can be replicated and, second, to establish a basis for investigating the influence of goal representations in the following experiments. In Experiment 3 and Experiment 4 combinations of temporal and spatial targets were investigated to see under which conditions spatial and temporal kinematic patterns are applied and to provide evidence for the influence of goal representations on movement kinematics.
EXPERIMENT 1
In Experiment 1, movements to temporal targets without spatial constraints were compared to movements to nontargets. Participants performed continuous reversal movements along the medial-lateral axis. The temporal target consisted of a tone that was presented to the participants at regular intervals. No spatial constraints were present; participants were allowed to choose the movement distance freely. Participants were instructed (in three different blocks) to be either somewhere on the left side when a tone occurred, or to be somewhere on the right side when a tone occurred, or to alternate between arriving somewhere on the right and left side when a tone occurred. Thus, movements to the temporal target (tone) and away from it had to be of the same distance (due to the use of continuous reversal movements), and the same temporal constraints applied to both parts of the reversal movement. By varying the side that had to be synchronized with the tone, the same areas of workspace apply to movements towards and movements away from targets. It was expected that movements to the temporal target have a relatively shorter duration and are performed relatively faster than movements away from it (see Balasubramaniam et al., 2004; Billon et al., 1996; Carlton, 1994).
Method
Participants
A total of 10 students (8 female, 2 male) from Munich universities took part in the experiment. All participants were right-handed according to the Edinburgh Inventory (Oldfield, 1971). The mean laterality quotient was 97 (SD = 5). Mean age was 21.9 (SD = 2.8) years. Participants were paid 10 euros for participation in the experiment. The experiment conformed to the British Psychological Society's (1993) guidelines for nonclinical experiments, and participants gave informed consent (this applies also to the following experiments).
Materials and apparatus
A 1,000 Hz tone, presented for 50 ms through the internal speakers, was used as a pacing signal. Movements were recorded with a Wacom Ultrapad A3 writing pad at a resolution of 500 pixels per cm and at a rate of 100 Hz. A cover screened the hand from view. Participants were to continuously make movements along the medial-lateral axis on the writing pad without pausing at the reversal points. Thereby they had to coordinate ending movements into one direction with the presentation of the pacing signal. Participants did not see their movement trace.
Three types of blocks were conducted, which differed in terms of the instructions (and frequency of the pacing signal). In the “tone-right” block participants were instructed to end movements somewhere on the right side, and in the “tone-left” block participants were instructed to end movements somewhere on the left side coincident with the occurrence of the tone. In the “tone-both-sides” block participants were instructed to alternate between arriving at the right and the left side when the tone occurred. Consequently, the interval between two successive pacing signals was only half of the intervals of tone-left and tone-right blocks for a given tempo condition. Five different tempi were used for the time available to conduct a complete reversal movement: 500, 750, 1,000, 1,250, and 1,500 ms. These times correspond to the interval between the presentations of the pacing signal in the tone-left and tone-right blocks. In the tone-both-sides block, the pacing signal was presented at the half of those times.
Procedure
The experiment took place in a dimly lit room. The writing pad was placed in front of the participants at a height that allowed for comfortable drawing. Participants performed the task with their right hand. Participants were instructed to continuously draw strokes along the medial-lateral axis and to reverse the direction without pausing. Thereby they had to coordinate ending movements into one direction with the presentation of the auditory signal.
Participants performed three blocks, the order of which was counterbalanced between participants. Each block started with a short demonstration phase, presenting an example rhythm and instructions. After the demonstration 20 experimental trials were conducted. The experimental trial series started with the slowest tempo, getting faster with each trial. In the sixth trial, the fastest tempo was presented (as in Block 5). In the succeeding trials the tempo became slower again. This procedure was repeated twice. Thus, four trials had to be performed in each tempo in each block. At the start of each trial a red box (0.5 × 0.5 cm) appeared at the starting position in the centre of the screen. As soon as participants entered the box the trial started, and the presentation of the pacing signal began. The duration of one trial was 40 s.
Data analysis
The first 10 s of each trial were always excluded from the analyses. In a first step, the position data were interpolated to yield a constant sample rate of 100 Hz because there was some variation in the sampling period of the writing pad (7–13 ms). In a second step, the kinematic data were smoothed using a nonlinear algorithm (Mottet, Bardy, & Athenes, 1994). In a third step, pen velocity was determined at each point in time. Stroke onset was defined as the point in time and space when participants were at the reversal points of the movements. This was also the endpoint of the previous stroke. These points were defined by the maximum displacement in the x-dimension. That is the most leftward and most rightward points of a movement, which were followed by two data samples indicating that the movement had reversed to the other direction. Strokes were excluded from analyses when (a) participants did not move continuously (the criterion was that a participant did not move more than 1 mm within the first 50 ms of a stroke—the strokes before and after the respective stroke were also excluded), (b) stroke duration was more than one third shorter or longer than the assigned stroke time for the corresponding condition, and (c) synchronization was off more than one third of the assigned stroke time. Using these criteria on average 256 (500-ms tempo), 199 (750-ms tempo), 167 (1,000-ms tempo), 121 (1,250-ms tempo), and 85 (1,500-ms tempo) strokes were available in each condition for each participant.
Because there were no differences in the pattern of performance in terms of whether the movement was conducted to the left or to the right within conditions, the data were collapsed over this factor, thus yielding 3 levels of the factor target: (a) movements towards the side where synchronization occurred with the tone—“temporal target”, from the blocks with the synchronization requirement on one side (tone-left and tone-right blocks); (b) movements to the other side—that is, away from the temporal target—“no target”, from the same blocks; (c) movements, where both sides of the reversal movement required synchronization—“two temporal targets”. A second factor was the time available to complete one reversal movement: tempo (500, 750, 1,000, 1,250, 1,500 ms). Data were subjected to repeated measurements analyses of variance (ANOVAs), and if significant effects were found further testing was done using contrasts.
Two sets of dependent variables were analysed, one to characterize the shape of the trajectory and one to characterize target variability. In order to describe the shape of the trajectory, proportional movement time (PMT, the time spent on one stroke relative to the time available for a whole reversal movement), peak velocity (PV, the maximum velocity value occurring during each stroke in cm/s), and proportional time to peak velocity (PTPV, the point in time in a stroke when peak velocity occurred, relative to the duration of the whole stroke) were analysed (see, for example, Helsen, Elliott, Starkes, & Ricker, 1998, for using proportional time as a dependent measure). To investigate target variability, reversal movement time variability (RMT_V, temporal variability) and endpoint variability (EP_V, spatial variability) were analysed. Reversal movement time is the time spent on a whole reversal movement—that is, two strokes. For example, in tone-left and tone-right blocks, the strokes to one side are supposed to be synchronized with the tones. Reversal movement time can then be calculated for the following conditions: from the point in time where a participant synchronizes the movement reversal with the tone to the next synchronization with the tone (“temporal target”), and the point in time where the movement at the no-tone side ends to the next time the participant is on the no-tone side (“no target”). In the blocks in which synchronization with the tone had to occur on both sides, the reference point was arrival on the same movement side. The variability of these times was used for RMT_V. Endpoint variability (EP_V) is the variability around the average position at which the strokes ended (only variability for the medial-lateral axis is reported). A graphical overview of the dependent variables using a position and a velocity profile is depicted in Figure 1.
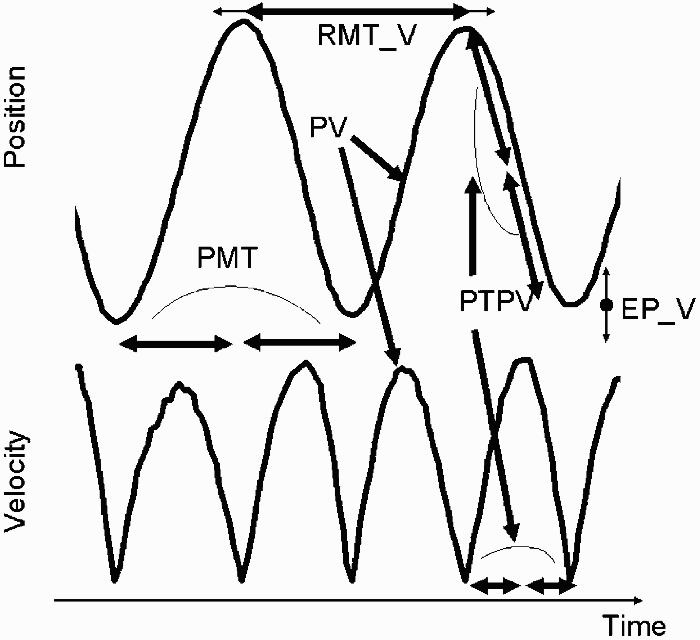
Graphical overview of the dependent variables using a position and a velocity profile. PMT = proportional movement time; PV = peak velocity; PTPV = proportional time to peak velocity; RMT_V = reversal movement time variability; EP_V = endpoint variability. In order to describe the shape of the trajectory, PMT, PV, and PTPV were analysed. PMT is the time spent on one stroke relative to the time available for a whole reversal movement, PV is the maximum velocity value occurring during each stroke, and PTPV is the point in time in a stroke when peak velocity occurred, relative to the duration of the whole stroke. To investigate target variability, RMT_V and EP_V were analysed. RMT_V is the variability of the time spent on a whole reversal movement (i.e., two strokes). EP_V is the variability around the average position at which the strokes ended (only variability for the medial-lateral dimension is reported).
Results
The means and standard deviations of the dependent variables for all the conditions are presented in Table 1.
Kinematic data for Experiment 1
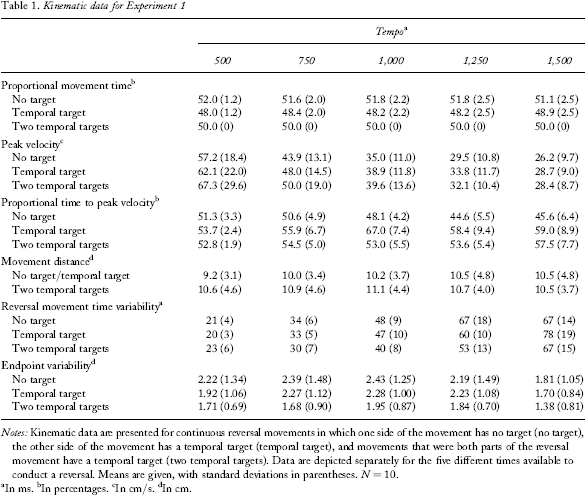
Notes: Kinematic data are presented for continuous reversal movements in which one side of the movement has no target (no target), the other side of the movement has a temporal target (temporal target), and movements that were both parts of the reversal movement have a temporal target (two temporal targets). Data are depicted separately for the five different times available to conduct a reversal. Means are given, with standard deviations in parentheses. N = 10.
In ms.
In percentages.
In cm/s.
dIn cm.
Shape of the trajectory
Proportional movement time (PMT)
The two-temporal-target level was excluded from this analysis, because it has by definition a mean of 50% and no variance in PMT. This is because data were collapsed over movements to the left and the right. Thus, a repeated measurements ANOVA with the factors target (temporal target, no-target) and tempo (500, 750, 1,000, 1,250, 1,500) was conducted. There was a significant main effect of target, F(1, 9) = 8.92, p =.02, indicating that in the temporal target condition participants had significantly lower PMT than in the no-target condition (48.4 vs. 51.6%). The Target × Tempo interaction was not significant.
Peak velocity (PV)
There was a significant main effect of tempo, F(4, 36) = 48.69, p <.0001, indicating that PV decreased at the slower tempi (59.6, 46, 36.9, 31.7, 27.5 cm/s from the fastest to the slowest tempo), no significant effect of target, and no interaction of Target × Tempo. However, when only comparing the temporal target and no-target conditions in one ANOVA, a significant effect of target was obtained, F(1, 9) = 15.72, p =.003, indicating that peak velocity was higher in the temporal target than in the no-target condition (42.3 vs. 38.4 cm/s).
Proportional time to peak velocity (PTPV)
There was no significant main effect of tempo, but a significant main effect of target, F(2, 18), p =.003, and a significant Target × Tempo interaction, F(8, 72) = 5.21, p <.0001. Movements to temporal targets had a higher PTPV (58.8%) than movements away from them (48%). PTPV for two-temporal-target movements levelled in between the other two kinds of movement. The interaction indicates that these effects were stronger at slower tempi. Apart from one difference between temporal target and two-target movements at the 1,500-ms tempo, all differences between the conditions were significant at the 1,000-, 1,250-, and 1,500-ms tempi (all p <.05).
Movement distance (MD)
There were no significant main effects or interactions. The average distance values ranged from 9.2 to 11.1 cm.
Target variability
Reversal movement time variability (RMT_V)
There was a significant main effect of tempo, F(4, 36) = 110.92, p <.001, indicating that RMT_V increased at slower tempi, a significant main effect of target, F(2, 18) = 3.61, p =.05, and a significant Target × Tempo interaction, F(8, 72) = 2.76, p =.01. At the medium tempi, RMT_V was higher in the no-target than in the two-target condition (significant for the 750-ms, 1,000-ms, and 1,250-ms tempi). None of the other effects was significant.
One may be concerned that results are influenced by the fact that participants performed the different conditions for the same amount of time rather than for the same number of movements. An analysis in which only the first 20 reversal movements after the 10-ms practice phase of each trial were included yielded, however, the same pattern of results as the analysis on all reversal movements: main effect of tempo, F(4, 36) = 122.43, p <.001; main effect of target, F(2, 18) = 3.58, p =.05; Target × Tempo interaction, F(8, 72) = 2.28, p =.03. Therefore, in the further experiments only the analysis on all data are reported.
Endpoint variability (EP_V)
There was a significant effect of tempo, F(4, 36) = 3.46, p <.02, but no effect of target and no Target × Tempo interaction. The only difference between the tempi that reached significance was between Tempo 1,000-ms and Tempo 1,500-ms, indicating that the former showed more variability than the latter (p =.01).
Discussion
In Experiment 1 it was investigated which movement kinematics participants exhibit when they move to a temporal target with no spatial constraints. Overall, the same target-directed kinematics were observed at all tempi—shorter movement times, higher peak velocity, and higher proportional time to peak velocity of movements towards temporal targets than of movements away from them. These results are in accordance with the observations that movements with temporal accuracy constraints show a relatively late peak velocity (e.g., Balasubramaniam et al., 2004; Carlton, 1994). The observed pattern is in the following called “temporal goal kinematics” to reflect that this is the movement pattern that participants choose to adopt in order to move to a temporal target. One may argue that the task used here does not only consist of temporal targets, but that there are also spatial constraints—being on the left or right side. However, no spatial targets are present, and no spatial precision is required. The present task does not have more spatial constraints than, for example, a tapping task, where participants move up and down or even tap on a table.
EXPERIMENT 2
In Experiment 2 the movement kinematics to purely spatial targets without temporal constraints in comparison to movements to nontargets were investigated. This time, it was expected that relatively long movement times and a relatively early peak velocity in movement to the spatial target in comparison to movements away from it would be observed (Carlton, 1994; Elliott et al., 2001; Helsen et al., 2000; Hogan & Flash, 1987; Langolf et al., 1976; MacKenzie et al., 1987; Soechting, 1984).
Method
Participants
A total of 10 students (8 female, 2 male) from Munich universities who had not taken part in the previous experiment participated. All participants were right-handed according to the Edinburgh Inventory (Oldfield, 1971). The mean laterality quotient was 87 (SD = 24). Mean age was 24.7 (SD = 2.9) years. Participants were paid 4 euros for participation in this experiment (they had the opportunity to take part in another unrelated experiment to earn more money).
Materials and apparatus
Stimuli were presented on a 17″ screen, with a resolution of 600 × 800 pixels. The vertical screen refresh rate was 75 Hz. Movements were again recorded with a Wacom Ultrapad A3 writing pad at a resolution of 500 pixels per cm and at a rate of 100 Hz.
The screen displayed, depending on conditions, one or two black boxes, one on the left and/or one on the right side. The centres of the boxes were separated by a distance of 10.7 cm. This distance on the screen resembled a distance of 7.3 cm on the writing pad. Size of the boxes was 0.8 cm in width and 1.6 cm in height. The index of difficulty (Fitts, 1954) was therefore 4.7 in the condition where there were boxes on both sides. Participants carried out movements along the medial-lateral axis between these boxes. The movement trace was displayed on the screen. Again, three types of blocks were conducted. In one block, there was a box on the right side, in another block, there was a box on the left side, and in a further block there was a box on both sides.
Procedure
Only differences to Experiment 1 are reported here. Behind the writing pad, a monitor was positioned slightly higher, and participants were seated approximately 60 cm away from the monitor. Participants were instructed to perform continuous reversal movements at a fast but not uncomfortable tempo. They were told that they should reverse the movements on the boxes. In case there was only a box on one side, they were supposed to reverse the movement somewhere on the opposite side of the screen.
A trial started when participants moved into the starting position in the centre of the screen. Trial duration was 60 s. Participants performed three trials within each block. The order of blocks was counterbalanced between participants. In order to get used to the task participants performed one trial of the condition in which there were boxes on both sides before the actual experimental blocks began. In this block, participants were instructed to perform the movements at a comfortable tempo.
Data analysis
Because there were no differences in the pattern of performance in terms of whether the movement was conducted to the left or to the right within conditions, the data were collapsed over this factor. ANOVAs were conducted with the factor target (no-target, spatial-target, two-spatial-target). On average, 310 strokes were available in each condition for each participant. To analyse the shape of the trajectory and target variability, the same dependent variables were used as those in Experiment 1.
Results
The means and standard deviations of the dependent variables for all the conditions are displayed in Table 2.
Kinematic data for Experiment 2
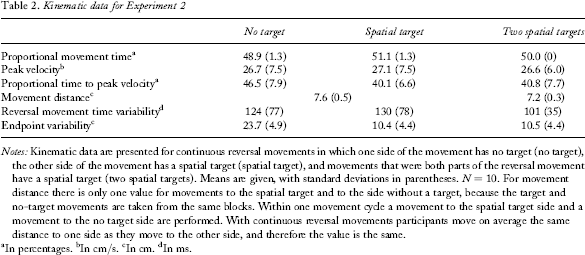
Notes: Kinematic data are presented for continuous reversal movements in which one side of the movement has no target (no target), the other side of the movement has a spatial target (spatial target), and movements that were both parts of the reversal movement have a spatial target (two spatial targets). Means are given, with standard deviations in parentheses. N = 10. For movement distance there is only one value for movements to the spatial target and to the side without a target, because the target and no-target movements are taken from the same blocks. Within one movement cycle a movement to the spatial target side and a movement to the no target side are performed. With continuous reversal movements participants move on average the same distance to one side as they move to the other side, and therefore the value is the same.
In percentages.
In cm/s.
In cm.
In ms.
Shape of the trajectory
Proportional movement time (PMT)
The two-spatial-target level was excluded from this analysis, because it had a mean of 50% and zero variance in PMT. Thus, a t test for dependent samples with the conditions spatial-target versus no-target was conducted. This comparison was significant, t(9) = 2.73, p =.02, showing that movements towards the spatial target had higher PMT than movements away from the spatial target. Absolute movement times (MTs) for one stroke were 522, 551, and 634 ms for the no-target, target, and two-target conditions, respectively.
Peak velocity (PV)
The ANOVA on PV did not result in a significant effect, F(2, 18) = 0.4.
Proportional time to peak velocity (PTPV)
The ANOVA on PTPV was significant, F(2, 18) = 15.31, p =.0001. The spatial-target condition and the two-spatial-target condition both had lower PTPV than the no-target condition (both p <.005). There were no significant differences between the spatial-target and two-spatial-target conditions.
Movement distance (MD)
Movements in the two-spatial-target condition were significantly shorter than movements with a spatial target on only one side, t(9) = 3.92, p =.004. However, this difference was not large (0.4 cm).
Target variability
Reversal movement time variability (RMT_V)
There were no significant differences between the conditions, F(2, 18) = 2.1.
Endpoint variability (EP_V)
There was a significant effect of target, F(2, 18) = 183.5, p <.0001. Spatial target and two-target movements had significantly less EP_V than no target movements (both p <.0001). There was no significant difference between spatial-target and two-spatial-target movements.
Discussion
In Experiment 2, movement kinematics to purely spatial targets were investigated. Movements towards spatial targets had higher movement time, earlier peak velocity, and less endpoint variability than movement away from spatial targets. The results are in accordance with several other studies in the literature in which spatially constrained movements were investigated (e.g., Carlton, 1994; Helsen et al., 1998; Helsen et al., 2000). It should further be noted that participants were not instructed to minimize movement time (as in Carlton, 1994), but were told to move at a fast but comfortable tempo. The fact that comparable results regarding the proportional time to peak velocity (e.g., 40% in Carlton, 1994; between 35.6 and 40.8% in Helsen et al., 1998; 40.1% in the present study) further speaks for a general kinematic pattern of movements to spatial targets, regardless of task instructions. Furthermore, as in Experiment 1, it should be emphasized that the effects were obtained by comparing movements to spatial target with movements to no targets. The observed pattern is in the following called “spatial goal kinematics” to reflect that this is the movement strategy that participants choose to adopt in order move to a spatial target.
Thus far, movements towards temporal targets (Experiment 1) and movements towards spatial targets (Experiment 2) were investigated. In Experiment 3 and Experiment 4 combinations of temporal and spatial targets are used to investigate the main issue of the present study: to provide evidence for the influence of goal representations on movement kinematics.
EXPERIMENT 3
In Experiment 3, participants were provided with spatial targets on both sides of the reversal movement. The temporal target was varied. The instructions emphasized that temporal accuracy was more important than spatial accuracy. There were three conditions: (a) movements to the spatial-temporal target; (b) movements to the spatial target (both from the same blocks); and (c) movements from the block with a spatial-temporal target on both sides.
Because movements to spatial and temporal targets seem to be controlled in different ways, it was expected that it would be difficult for participants to meet both targets equally well, especially at fast tempi. It was further assumed that movement kinematics would depend upon tempo. At fast tempi, it was assumed that participants would show movement kinematics that would resemble temporal goal kinematics towards the spatial-temporal target in comparison to the spatial target, because participants were instructed to give the temporal target priority.
Because there is less time pressure at the slower tempi, participants should be able to incorporate the spatial targets into their movement kinematics. However, there are several possibilities for how participants deal with the combination of spatial and temporal targets at slower tempi. First, it could be that participants show a mix of the spatial and temporal kinematic patterns towards the spatial-temporal target if they simply follow task demands. Thus, there should still be evidence of temporal movement kinematics. However, contrary to Experiment 1, in which proportional time to peak velocity occurred even later at slower tempi than at faster tempi, it was not expected that such an effect occurs in the present experiment, because this would counteract any attempts to incorporate spatial movement kinematics (“kinematics assimilation hypothesis”). Second, the temporal target aspect may become irrelevant or of minor importance for movement kinematics. There is enough time to meet both the temporal target aspects and the spatial target aspects. Participants should then show equal concern for the spatial targets on both sides of the reversal movement. Movements to the spatial-temporal target and movements to the spatial target should then not differ from each other in terms of their kinematics (“relevance for performance hypothesis”). Third, another possibility is that participants represent the side with the spatial-temporal target as the “goal-side” of the movement, because there are two targets on this side, a circumstance that could result in a stronger goal representation. The other side of the reversal movement has only one target (spatial target), which could result in a weaker goal representation; this side is therefore represented as the “no-goal-side”. Because slower movements allow for incorporating the spatial target into the movement kinematics, spatial goal kinematics should be shown towards the spatial-temporal target in comparison to the spatial target if goal representation plays a role (“goal representation hypothesis”).
Method
Participants
A total of 11 students (9 female, 2 male) from Munich universities who had not taken part in the previous experiments participated. All participants were right-handed according to the Edinburgh Inventory (Oldfield, 1971). The mean laterality quotient was 87 (SD = 17). Mean age was 22.6 (SD = 2.7) years. Participants were paid 10 euros for participation.
Materials and apparatus
Stimuli were as in the previous experiments (auditory stimuli as in Experiment 1, visual stimuli as in Experiment 2), and the general setup was also the same as before. Only the differences to the previous experiments are reported here.
Spatial targets were always present for both sides of the movement. Three types of blocks were conducted, which differed in terms of the instructions (and frequency of the pacing signal). In the tone-right block participants were instructed to be at the right box when the tone occurred, and in the tone-left block participants were instructed to be at the left box when the tone occurred. In the tone-both-sides block participants were instructed to alternate between the right and the left box when the tone occurred. Consequently, the interval between two successive pacing signals was only half of the intervals of the tone-left and tone-right blocks for a given tempo condition. Five different tempi were used for the time available to conduct a complete reversal movement: 500, 750, 1,000, 1,250, and 1,500 ms.
Procedure
Participants were instructed to continuously draw strokes between the two boxes displayed on the screen. They were to reverse the direction without pausing. Thereby they had to coordinate arrival on the boxes with the presentation of the auditory signal. The instructions emphasized that temporal accuracy was more important than spatial accuracy.
Data analysis
Data preparation was like that in the previous experiments. On average 290 (500-ms tempo), 209 (750-ms tempo), 184 (1,000-ms tempo), 148 (1,250-ms tempo), and 121 (1,500-ms tempo) strokes were available in each condition for each participant. Because there were no differences in the pattern of performance in terms of whether the movement was conducted to the left or to the right within conditions, data were collapsed over this factor, thus yielding three levels of the factor target: (a) movements towards the side where synchronization occurred with the tone, “STT” (spatial-temporal target), from the blocks with the synchronization requirement on one side (tone-left and tone-right); (b) movements to the other side—that is, away from the spatial-temporal target to the spatial only target—“ST” (spatial target) from the same blocks; and (c) movements where both sides of the reversal movement required synchronization (tone-both-sides block), “two-STT” (two spatial-temporal target). A second factor was the time available to complete one reversal movement: tempo (500, 750, 1,000, 1,250, 1,500 ms). Data were subjected to repeated measurements ANOVAs, and if significant effects were found further testing was done using contrasts. The same set of dependent variables as before was analysed (apart from movement distance, which was fixed by providing two spatial targets).
Results
The means and standard deviations of the different variables for all the conditions are presented in Table 3.
Kinematic data for Experiment 3
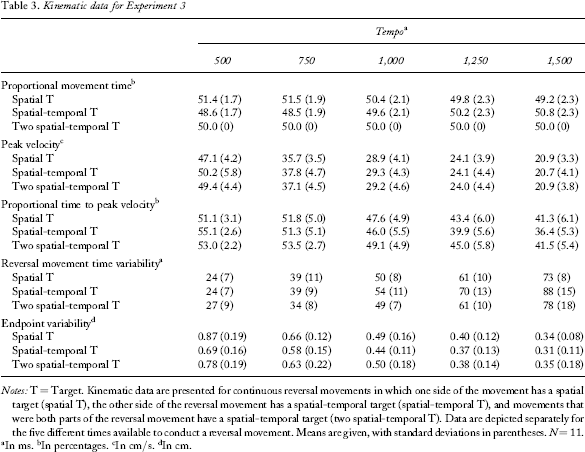
Notes: T = Target. Kinematic data are presented for continuous reversal movements in which one side of the movement has a spatial target (spatial T), the other side of the reversal movement has a spatial-temporal target (spatial-temporal T), and movements that were both parts of the reversal movement have a spatial-temporal target (two spatial-temporal T). Data are depicted separately for the five different times available to conduct a reversal movement. Means are given, with standard deviations in parentheses. N = 11.
In ms.
In percentages.
In cm/s.
In cm.
Shape of the trajectory
Proportional movement time (PMT)
The two-STT condition was excluded from this analysis, because the mean of PMT is by definition 50% with zero variance. Thus, a repeated measurements ANOVA with the factors target (STT, ST) and tempo (500, 750, 1,000, 1,250, 1,500) was conducted. There was neither a significant main effect of target, nor one of tempo (which is this way by definition, because the sum of the percentages always equals 100% for each tempo). However, a significant Target × Tempo interaction, F(4, 40) = 4.24, p =.006, was observed. Comparisons revealed that at the two fastest tempi, movements towards the spatial-temporal target required significantly less PMT than movements to the spatial target (500 and 750 ms, both p <.05). No significant differences were obtained for the other tempi.
Peak velocity (PV)
There was a significant main effect of tempo, F(4, 40) = 274.92, p <.0001, indicating that PV decreased at the slower tempi. The main effect of target was not significant. Again, there was a significant Target × Tempo interaction, F(8, 80) = 2.48, p =.02. Comparisons showed that at the two fastest tempi movements towards spatial-temporal targets had a significantly higher PV than movements to spatial targets (500- and 750-ms tempi, both p <.05). PV in the two-STT condition and PV in the STT condition were not significantly different from each other. PV in the two-STT condition was higher than that in the ST condition for the 500-ms tempo (p =.03) and showed a tendency in the 750-ms tempo (p =.08). No significant differences were obtained at the other tempi.
Proportional time to peak velocity (PTPV)
There was a significant main effect of tempo, F(4, 40) = 44.45, p <.0001, indicating that PTPV decreased at the slower tempi (that is, at slower tempi peak velocity occurred relatively earlier within the stroke), a main effect of target, F(2, 20) = 4.14, p =.03, and a significant Target × Tempo interaction, F(8, 80) = 7.56, p <.0001. Comparisons showed that at the 500-ms tempo STT movements had a significantly higher PTPV than did ST movements (p =.01), but that at the 1,500-ms tempo STT movements had a significantly lower PTPV than did ST movements (p =.02). Two-STT movements had higher PTPV than ST movements and lower PTVP than the STT movements at the 500-ms tempo (both p <.05). Further, two-STT movements had higher PTPV than did STT movements at the 1,000-ms, 1,250-ms, and 1,500-ms tempi (all p <.05).
Target variability
Reversal movement time variability (RMT_V)
There was a significant main effect of tempo, F(4, 40) = 151.9, p <.0001, indicating that there was higher variability at slower tempi, a significant main effect of target, F(2, 20) = 8.35, p =.002, and a significant Target × Tempo interaction, F(8, 80) = 3.06, p =.005. At the slower tempi, RMT_V was higher in the STT than in the ST condition (1,000-ms, 1,250-ms, and 1,500-ms intertrial intervals, all p <.05). The two-STT condition showed higher RMT_V than did the other two conditions at the 500-ms tempo (both p <.05). In the slower tempo conditions RMT_V did not significantly differ between the two-STT and ST conditions, but was lower in the two-STT condition than in the STT condition at the 1,250-ms tempo (p =.02).
Endpoint variability (EP_V)
There was a significant effect of tempo, F(4, 40) = 129.14, p <.0001, indicating that there was lower variability at slower tempi. There was only a tendency for target, F(2, 20) = 2.91, p =.08, but a significant Target × Tempo interaction, F(8, 80) = 3.43, p =.002. The STT condition showed less EP_V than did the ST condition at all tempi—however, significantly only at the 500-ms and 1,250-ms tempi (both p <.05). The two-STT condition did not differ from the ST condition and showed significantly more EP_V than did the STT condition at the 500-ms and 1,000-ms tempi (both p <.05).
Discussion
As expected, movements towards spatial-temporal targets showed different kinematics than did movements to spatial targets. Most importantly, two different kinematic patterns for movements towards spatial-temporal targets were observed. In fast movements, participants showed temporal goal kinematics to the spatial-temporal goal in comparison to the spatial goal (relatively less movement time, higher and later peak velocity when moving toward the spatial-temporal target than when moving to the spatial target). In slow movements participants showed spatial goal kinematics towards the spatial-temporal target (relatively earlier peak velocity in movements towards the spatial-temporal target than in movements to the spatial target).
It is possible that these two different kinematic patterns solely depend on the tempo with which movements have to be performed. During fast movements there is time pressure; during slow movements there is not. Moving towards targets in a fast way might pose constraints (either cognitive and/or motor) on how a target-directed movement can be performed, which may not be present when moving towards targets slowly. However, in Experiment 1 with only temporal targets, no tempo-dependent differences in kinematics were observed; slow tempi showed the same kinematic pattern as fast tempi. The data in the literature also make it seem likely that the differences in kinematics that were observed are not due to tempo differences (Balasubramaniam et al., 2004; Billon et al., 1996; Carlton, 1994; Elliott et al., 2001; Hogan & Flash, 1987; Langolf et al., 1976; MacKenzie et al., 1987; Meyer et al., 1988; Soechting, 1984). In the study of Carlton (1994), for example, movements to spatial targets (time minimization instructions) were faster than movements to temporal targets (temporal accuracy instructions), but showed kinematic patterns that resemble those found here. (Even though the term “time minimization” seems to emphasize the temporal aspect of the task it has to be emphasized that there was actually no specific temporal target. That is, participants were supposed to be as fast as possible, but there was no given point in time at which they had to be in the target position.)
Rather than tempo differences, the observed kinematics reflect the type of target present and the goal participants have when performing a movement. When movements have to be executed fast, the primary goal may be, as instructed, to manage the synchronization task. This goal is temporal. With slow movements, however, participants focus on the spatial target as well. This interpretation matches the observation that spatial endpoint variability decreased with slower tempi. The data for slower movements are in accordance with what was termed “goal representation hypothesis” in the introduction to this experiment. Spatial targets were present on both sides of the reversal movement—therefore, one may wonder why the spatial target on the no-tone side should be less important. It is likely that the side at which synchronization has to occur is represented as the “goal side”, at which goal-directed kinematic patterns are applied. This is in accordance with the result that endpoint variability was smaller at the spatial-temporal target than at the spatial target side. It could be that more attention was paid to the task requirements at the side that was represented as the goal side.
Another interesting aspect in the data was that variability of reversal movement time was actually higher at the spatial-temporal target than at the spatial target positions at the slower tempi. This is contrary to what one might expect if participants optimize behaviour to meet the temporal target. There may be conflicts in the attempt to meet spatial and temporal goals at once, resulting in higher temporal variability when participants attend to the spatial target as well.
In the two-STT condition, peak velocity resembled the STT condition in fast movements. This seems plausible, because in fast movements the movements go to a temporal target in the two-STT as well as in the STT condition. Proportional time to peak velocity was higher in the two-STT than in the STT condition for slower movements. This result might be due to a stronger representation of temporal goal aspects in the two-STT conditions than in the STT condition, because the pacing signal occurred twice as often. However, it seems that spatial targets also became more important in the two-STT condition as movement tempo decreased. Peak velocity occurred earlier at slower tempi than at faster tempi, a result that is in contrast to Experiment 1 where proportional time to peak velocity was later at slower than at faster tempi.
Summing up, two different kinematic patterns for moving to the spatial-temporal target were observed. These two kinematic patterns are likely to be due to different priorities in temporal and spatial goal aspects of the task, modulated by tempo. Further, the specific pattern reflects that there is a cognitive representation of one side as the “goal” side.
EXPERIMENT 4
Experiment 4 was conducted to further investigate how the combination of spatial and temporal targets influences movement kinematics. The aim was to provide further evidence for the goal representation hypothesis that was put forward in Experiment 3. That is, it was intended to replicate a similar data pattern under different conditions. The experiments that were conducted so far have investigated movement kinematics under conditions with purely temporal targets (Experiment 1), purely spatial targets (Experiment 2), and spatial targets always present, and varying temporal targets (Experiment 3). Thus, in order to complete the picture, and to provide converging evidence for the obtained results, three further manipulations were conducted: (a) temporal targets on both sides of the reversal movement, spatial target on one of the sides (manipulation: two temporal); (b) spatial and temporal target on the same side of the reversal movement (manipulation: combined); and (c) spatial and temporal targets on opposite sides of the reversal movement (manipulation: separate).
The predictions for this experiment are straightforward and are based on the results obtained in the previous experiments. In the combined blocks, it can be expected that a similar pattern as that in Experiment 3 would be seen—different kinematic patterns depending on tempo. In fast tempi the temporal target should be more important; in slow tempi the spatial target should be more important. In the separate blocks (spatial and temporal targets on opposite sides), the movement kinematics should be the same in all tempo conditions, because the kinematic features are partly complementary for spatial and temporal targets. For the two temporal target blocks (spatial-temporal target on one side, temporal target on the other side) it was once more assumed that the kinematic patterns differ depending on tempo. Here it can be assumed that the side with the spatial-temporal target might be represented as the goal side. This manipulation could therefore provide further evidence for the goal representation hypothesis. Evidence for goal representation should, however, be observable at fast tempi this time and not at slow tempi as in Experiment 3. At fast tempi, participants should treat the side with the spatial-temporal target like they treat a temporal target, even though they were provided with temporal goals for both sides of the reversal movement. At slow tempi they should treat the spatial-temporal target like a spatial goal.
Method
Participants
A total of 10 students (7 female, 3 male) from Munich universities who had not taken part in the previous experiments participated. All participants were right-handed according to the Edinburgh Inventory (Oldfield, 1971). The mean laterality quotient was 88 (SD = 17). Mean age was 25.5 (SD = 5) years. Participants were paid 14 euros for participation.
Materials and apparatus
Only differences to the previous experiments are reported here. The screen displayed one black box, the centre of which was, depending on the condition, either 5.35 cm to the left or 5.35 to the right of the centre of the screen. Participants carried out movements along the medial-lateral axis and were required to end one stroke of the reversal movement on the box. The trace of the movement was displayed on the screen.
Procedure
Six blocks were conducted, which differed according to the instructions regarding the temporal target and the position of the spatial target (as well as the frequency of the pacing signal). Within the blocks, tempo was varied between trials. In three blocks the box was displayed on the right side. In one of them participants were instructed to be at the box when the tone occurred (combined). In another block participants were instructed to be on the left side when the tone occurred (that is, opposite to the box, separate). In the third block participants were instructed to alternate between arriving on the right and on the left side, synchronizing each tone with a stroke (two temporal targets). Consequently, the interval between two successive pacing signals was only half of that of the intervals of the first two blocks mentioned. In three other blocks the box was displayed on the left side of the screen. Participants were again instructed to be on the right (separate), on the left (combined), or alternately on both sides (two temporal target) when the tone occurred.
At the start of each trial a red box (0.5 × 0.5 cm) appeared as the starting position on the side opposite to the box. Participants were instructed to cover approximately the distance between the box and the starting position in their movements. Participants performed six blocks (as described above); the order of them was counterbalanced between participants. Within the blocks, the same tempi as those in Experiment 1 and Experiment 3 were used.
Data analysis
On average 322 (500-ms tempo), 255 (750-ms tempo), 188 (1,000-ms tempo), 153 (1,250-ms tempo), and 120 (1,500-ms tempo) strokes were available in each condition for each participant. Data were collapsed over movements to the left and to the right, because there were no differences in the pattern of results, thus providing six target conditions: (a) combined blocks, movements towards the temporal and spatial target (spatial-temporal target); (b) combined blocks, movements away from the temporal and spatial target (no target); (c) separate blocks, movements towards the temporal and away from the spatial target (temporal target); (d) separate blocks, movements away from the temporal and towards the spatial target (spatial target); (e) blocks with the temporal target on both sides of the reversal movement, to the side with the spatial target (spatial-temporal target); (f) blocks with the temporal target on both sides of the reversal movement, away from the spatial target (temporal target). Data were subjected to repeated measurements ANOVAs, and in the case of significant effects further testing was done using contrasts.
Statistical analysis was conducted separately for (a) combined blocks, (b) separate blocks, and (c) blocks with the temporal target on both sides. For each of those manipulations ANOVAs with the factors target (combined: spatial-temporal target vs. no target; separate: spatial target vs. temporal target; two temporal: spatial-temporal targets vs. temporal target), and tempo (500, 750, 1,000, 1,250, 1,500 ms) were conducted. The same two sets of dependent variables as those in the previous experiments were analysed to characterize target-related movement kinematics (i.e., shape of the trajectory) and target variability.
Results
The means and standard deviations of the dependent variables for all the conditions are displayed in Table 4.
Kinematic data for Experiment 4
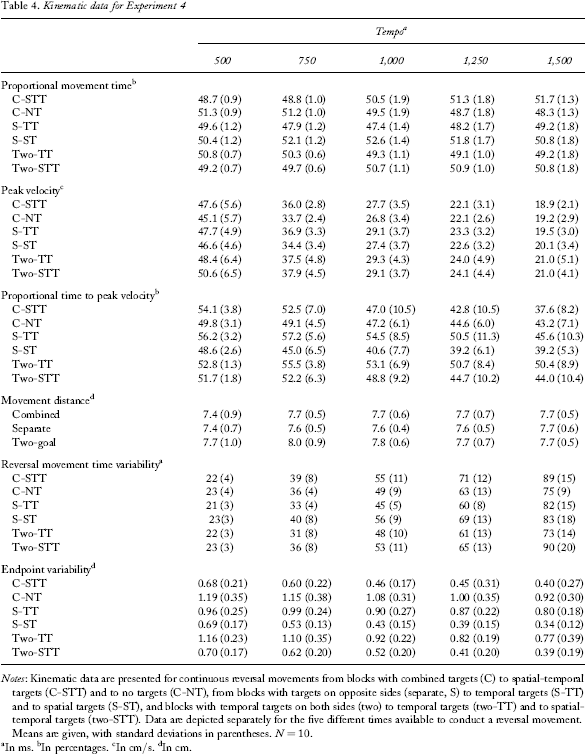
Notes: Kinematic data are presented for continuous reversal movements from blocks with combined targets (C) to spatial-temporal targets (C-STT) and to no targets (C-NT), from blocks with targets on opposite sides (separate, S) to temporal targets (S-TT) and to spatial targets (S-ST), and blocks with temporal targets on both sides (two) to temporal targets (two-TT) and to spatial-temporal targets (two-STT). Data are depicted separately for the five different times available to conduct a reversal movement. Means are given, with standard deviations in parentheses. N = 10.
In ms.
In percentages.
In cm/s.
dIn cm.
Combined blocks (spatial-temporal target vs. no target)
Shape of the trajectory—proportional movement time (PMT)
None of the main effects was significant. However, there was a significant Target × Tempo interaction, F(4, 26) = 16.78, p <.0001. Comparisons revealed that at the two fastest tempi, movements towards spatial-temporal targets required significantly less PMT than movements away from spatial-temporal targets (500-ms and 750-ms tempo, both p <.05), but that at the two slowest tempi the reverse was the case (1,250-ms and 1,500-ms tempo, both p <.05).
Shape of the trajectory—peak velocity (PV)
There was a significant main effect of tempo, F(4, 36) = 304.11, p <.0001, indicating that PV decreased at the slower tempi. There was also a significant main effect of target, F(1, 9) = 5.88, p =.04, and a significant Target × Tempo interaction, F(4, 36) = 4.33, p =.006. Movements towards spatial-temporal targets had higher PV at the fast tempi (500 ms: p =.03, 750 ms: p =.01), but there were no differences at the slow tempi.
Shape of the trajectory—proportional time to peak velocity (PTPV)
There was a significant main effect of tempo, F(4, 36) = 31.43, p <.0001, indicating that PTPV decreased at the slower tempi. There was no significant main effect of target, but a significant Target × Tempo interaction, F(4, 36) = 5.24, p =.002. The interaction can be explained by a higher PTPV in movements towards spatial-temporal targets at the fast tempi and a lower PTPV at slow tempi than that for movements away from targets. However, only at the fastest tempo was a statistical tendency between movements towards and away from spatial-temporal targets observed (500 ms: p =.07).
Target variability—reversal movement time variability (RMT_V)
There was a significant main effect of tempo, F(4, 36) = 165.55, p <.0001, indicating that RMT_V was higher with slower tempi. There was a tendency for target, F(1, 9) = 3.89, p =.08, and a significant Target × Tempo interaction, F(4, 36) = 3.05, p =.03. The interaction is due to higher RMT_V in movements towards spatial-temporal targets than in movements away from spatial-temporal targets at slower tempi (significant at the 1,500-ms tempo, p =.02).
Target variability—endpoint variability (EP_V)
There was a significant main effect of tempo, F(4, 36) = 6.63, p =.0004, indicating that EP_V decreased with slower tempi, and a significant main effect of target, F(1, 9) = 36.54, p =.0002, indicating that movements to the spatial-temporal target showed less endpoint variability than did movements to the no-target side. There was no significant Target × Tempo interaction.
Separate blocks (spatial target vs. temporal target)
Shape of the trajectory—proportional movement time (PMT)
There was no main effect of tempo, but a significant main effect of target, F(1, 9) = 17.35, p =.002, indicating that movements towards temporal targets (48.5%) had a lower PMT than did movements towards spatial targets (51.5%). There was also a significant interaction of Target × Tempo, F(4, 36) = 7.83, p =.0001. Comparisons showed that this was due to the fact that the differences between movements towards temporal targets and spatial targets were smaller at the 500- and 1,500-ms tempi than at the other tempi (both p <.05).
Shape of the trajectory—peak velocity (PV)
There was again a significant main effect of tempo, F(4, 36) = 311.56, p <.0001, indicating that PV decreased at the slower tempi. There was no significant main effect of target, but a significant Target × Tempo interaction, F(4, 36) = 4.7, p =.004. Movements towards temporal targets displayed a significant higher PV than did movements towards spatial targets at the medium tempi (750 and 1,000 ms, both p <.05).
Shape of the trajectory—proportional time to peak velocity (PTPV)
There was a significant main effect of tempo, F(4, 36) = 21.03, p <.0001, indicating that PTPV decreased at the slower tempi. There was also a significant main effect of target, F(1, 9) = 9.74, p =.01, indicating that movements towards temporal targets showed higher PTPV than did movements towards spatial targets (52.8% and 42.5%, respectively). This difference was modified by a Target × Tempo interaction, F(4, 36) = 4.6, p =.004. Comparisons showed that the difference was significantly higher at the 750-ms, 1,000-ms, and 1,500-ms tempi than at the 500-ms and 1,500-ms tempi (all p <.05).
Target variability—reversal movement time variability (RMT_V)
There was a significant main effect of tempo, F(4, 36) = 133.72, p <.0001, indicating once more that RMT_V increased at the slower tempi. There was also a significant main effect of target, F(1, 9) = 6.91, p =.03, indicating that movements towards temporal targets showed less RMT_V than did movements towards spatial targets. This difference was modified by a Target × Tempo interaction, F(4, 36) = 3.31, p =.02. Comparisons showed that the differences between movements towards temporal and towards spatial targets were significant for all except the slowest tempo (all p <.05).
Target variability—endpoint variability (EP_V)
There was a significant main effect of tempo, F(4, 36) = 15.02, p <.0001, indicating once more that EP_V decreased at slower tempi. There was also a significant main effect of target, F(1, 9) = 71.15, p <.001, indicating that movements towards spatial targets showed less EP_V than did movements towards temporal targets. There was a tendency for a Target × Tempo interaction, F(4, 36) = 2.22, p =.09, indicating that differences between conditions in EP_V increase at slower tempi.
Two temporal target blocks (spatial-temporal target vs. temporal target)
Shape of the trajectory—proportional movement time (PMT)
None of the main effects was significant, but there was a significant Target × Tempo interaction, F(4, 36) = 5.61, p =.001. Comparisons showed that at the fastest tempo, movements towards spatial-temporal targets had lower PMT than did movements towards temporal targets (p =.003), but at the slower tempi, movements towards spatial-temporal targets showed a tendency for higher PMT (1,000 ms: p =.08) or significantly higher PMT (1,250 ms: p =.02) than did movements towards temporal targets.
Shape of the trajectory—peak velocity (PV)
There was a significant main effect of tempo, F(4, 36) = 109.2, p <.0001, indicating that PV decreased at the slower tempi. There was also a significant main effect of target, F(1, 9) = 4.95, p =.05, and a significant Target × Tempo interaction, F(4, 36) = 8.59, p <.0001. Movements towards a spatial-temporal target showed higher PV than did movements towards a temporal target at the fastest tempo (500 ms, p =.0004).
Shape of the trajectory—proportional time to peak velocity (PTPV)
There was a significant main effect of tempo, F(4, 36) = 4.13, p <.007, indicating that PTPV decreased at the slower tempi. There was also a significant main effect of target, F(1, 9) = 19.01, p =.002, indicating that PTPV was higher in movements to the temporal target (52.5%) than in movements to the spatial-temporal target (48.3%). This effect was modified by a significant Target × Tempo interaction, F(4, 36) = 4.91, p <.003, which indicated that the difference between temporal and spatial-temporal target increased with slower tempi (it was also not significant at the 500-ms tempo).
Target variability—reversal movement time variability (RMT_V)
There was a significant main effect of tempo, F(4, 36) = 102.08, p <.0001, indicating that there was higher variability at the slower tempi. There was also a significant main effect of target, F(1, 9) = 17.44, p =.002, indicating that RMT_V was higher in movements to the spatial-temporal target than in movements to the temporal target. This effect was modified by a significant Target × Tempo interaction, F(4, 36) = 4.8, p <.003, which indicated that the difference between temporal and spatial-temporal target increased with slower tempi (it was not significant at the 500-ms tempo and showed only a tendency at the 750-ms tempo).
Target variability—endpoint variability (EP_V)
There was a significant main effect of tempo, F(4, 36) = 102.08, p <.0001, indicating that EP_V decreased with slower tempi. There was also a significant main effect of target, F(1, 9) = 17.44, p =.002, indicating that EP_V was lower when there was a spatial-temporal target (0.53 cm) than when there was a temporal target (0.95 cm). There was no Target × Tempo interaction.
All blocks—movement distance (MD).
An ANOVA on MD was conducted with the factors type of block (combined, separate, two temporal targets) and tempo (500 ms, 750 ms, 1,000 ms, 1,250 ms, 1,500 ms). Neither the two main effects nor the interaction became significant. Values ranged from 7.37 cm to 8.04 cm. Thus, the instruction to cover the approximate distance between the starting position and the box seems to have been successful.
Discussion
In Experiment 4 combined spatial and temporal targets on one side (combined blocks), temporal and spatial targets on opposite sides (separate blocks), and the influence of a spatial target when a temporal target is always present (two temporal target blocks) were investigated. Overall, the results are consistent with those of the previous experiments and are in favour of two different kinematic patterns that depend on whether participants focus on a temporal and/or a spatial goal. They further show that goal representations have an impact on movement kinematics.
The data from the combined blocks showed different kinematic patterns depending on tempo—fast movements were performed using temporal goal kinematics, slow movements using spatial goal kinematics. Those results replicate the conditions of Experiment 3, in which both sides of the reversal movements consisted of a spatial target, but only one a temporal target. The data from the separate blocks showed that movements towards temporal targets showed temporal goal kinematics (relatively shorter movement time, higher and later peak velocity, less reversal movement time variability, higher endpoint variability than movements to spatial targets) and movements to spatial targets showed spatial goal kinematics. The target variability data indicate that participants tried to optimize the current goal.
The results of the two temporal target blocks (spatial-temporal target vs. temporal target) are the most interesting with respect to the goal representation hypothesis. Different kinematic patterns were observed at fast and slow tempi. At fast tempi temporal goal kinematics were observed towards the spatial-temporal target (relatively shorter movement time and higher peak velocity than for movements towards the temporal target). At slow tempi spatial goal kinematics were observed in movements towards the spatial-temporal target (relatively longer movement times and earlier peak velocity than for movements towards the temporal target). To explain the effects at fast tempi, it is necessary to recruit the goal representation hypothesis again. In case participants apply temporal goal kinematics, proportional movement time should be the same for movements to the temporal and the spatial-temporal target (relevance for performance hypothesis). If participants behave to the physical features of the spatial and temporal targets at once, the kinematics towards the spatial-temporal target should be a mix of spatial and temporal goal kinematics (kinematics assimilation hypothesis). However, participants showed temporal goal kinematics when moving to the side at which an additional spatial target was present. This effect is a complementary to the results of Experiment 3 where spatial goal kinematics were observed when participants moved to the side on which the temporal target was presented in addition to the spatial target. It seems that in both cases, the effect is due to a representation of the side with the combined goal as the goal side (goal representation hypothesis).
GENERAL DISCUSSION
The main goal of the present study was to investigate whether movement kinematics are to be viewed as a reaction to target stimuli (position, time), or rather the means to attain intended goals (to be in a certain position at a given time). In order to provide a basis for this question, it was first shown that movement kinematics towards temporal and spatial targets are different from the movement kinematics away from those targets. Further, kinematics are different for movements to temporal and spatial targets (Experiments 1 and 2). To obtain evidence for the influence of goal representations on kinematics, combinations of temporal and spatial targets were investigated in Experiments 3 and 4 using the following conditions: spatial targets always present and varying temporal targets (Experiment 3); temporal targets always present, with varying spatial targets (Experiment 4); and combined and separate spatial and temporal targets (Experiment 4).
The question of whether movements towards targets differ in their kinematics from movement away from targets can be answered unequivocally with “yes”. Movements to a specific target were always different from movements away from the target. The specific kinematic pattern depended on the type of target: temporal and/or spatial. Table 5 gives an overview of the main results of the four experiments and also illustrates the two different kinematic patterns used to move towards temporal (kinematic pattern “T”) and spatial (kinematic pattern “S”) targets. The kinematic pattern used to move towards temporal targets consists of relatively short movement times, high peak velocity, and high proportional time to peak velocity—that is, peak velocity occurs relatively late. Comparatively little time of the movement is therefore spent in the target area. The kinematic pattern used to move towards spatial targets consists of relatively long movement times towards the target and relatively little proportional time to peak velocity. A relatively long period of the movement is therefore spent in the target area. Consequently, movement endpoint variability is lower. The findings replicate previous findings (e.g., Meyer et al., 1988; Zelaznik et al., 1988; see also below). Previously, kinematic differences have sometimes been attributed to differences in methodological procedure (Carlton, 1994). In the present study it was shown that they depend on the type of movement target. It was ruled out that the two kinematic patterns are caused by different movement tempi and have nothing to do with spatial and temporal targets. Simple tempo differences could not explain why participants chose to perform all tempo conditions in Experiment 1 using one kinematic pattern, but did not do so in Experiment 3. Tempo differences could also not explain the different kinematic patterns in combined and two-temporal-target blocks of Experiment 4.
Overview of the results of Experiments 1–4
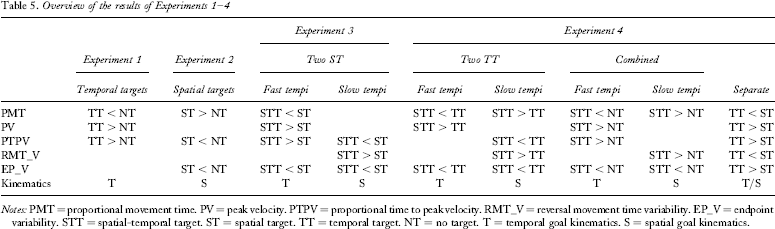
Notes: PMT = proportional movement time. PV = peak velocity. PTPV = proportional time to peak velocity. RMT_V = reversal movement time variability. EP_V = endpoint variability. STT = spatial-temporal target. ST = spatial target. TT = temporal target. NT = no target. T = temporal goal kinematics. S = spatial goal kinematics.
Are the temporal and spatial goal kinematics complementary to each other? The separate blocks of Experiment 4 showed that the kinematic patterns can appear to be complementary when a spatial target is on one side and a temporal target on the other. However, the separate blocks in this experiment were the only instance in all experiments in which movements towards spatial targets had lower peak velocity than did other movements. In all other experimental conditions, in which presumably spatial goal kinematics were used, peak velocity did not differ between the movement towards spatial targets and the movement away from them. This was the case when spatial targets were compared to no targets (Experiment 2), but also when spatial-temporal targets at a slow tempo were compared to spatial targets (Experiment 3), no targets, or temporal targets (both Experiment 4). When movements were conducted using temporal goal kinematics, peak velocity was higher or tended to be higher than that for the other movement, regardless of whether temporal targets were compared to no targets (Experiment 1), or whether spatial-temporal targets (at fast tempi) were compared to spatial targets (Experiment 3), no targets, or temporal targets (both Experiment 4). It can thus be assumed that the effect in the separate blocks of Experiment 4 (that is, lower peak velocity when participants moved towards spatial than towards temporal targets) can be attributed to temporal goal kinematics used when moving towards the temporal target. Thus, though one could argue that the temporal and spatial goal kinematic patterns are complementary regarding proportional movement time and proportional time to peak velocity, this does not hold for peak velocity. Therefore, the two kinematic patterns are not simply the opposite of each other. They probably reflect the kinematic features necessary to be able to meet the respective goals.
The features of both kinematic patterns are consistent with several results in the literature. There are several reports indicating that movements towards spatial targets result in asymmetric velocity profiles (profiles skewed to the right; i.e., a greater proportional time is spent after peak velocity; e.g., Chua & Elliott, 1993; Elliott, Lyons, & Dyson, 1997). When vision of movement target and movement trajectory is available, participants spend a greater proportion of their overall movement times after peak velocity than when vision is occluded. This extra time seems to be used for a feedback-based reduction of aiming errors (Chua & Elliott, 1993). Presumably, participants attempt to get the limb to the target area as quickly as possible so that they have more time to use visual feedback to achieve the final target position. Participants then use corrective submovements to improve final endpoint accuracy while reducing endpoint variability (Elliott et al., 2001; Novak, Miller, & Houk, 2000). The existence of submovements may account for the observed asymmetry of the velocity profile (Crossman & Goodeve, 1983; Novak, Miller, & Houk, 2002). The asymmetry of velocity profiles increases when accuracy demands of the movement are high, and/or targets are small (Elliott et al., 2001; Helsen et al., 1998; Hogan & Flash, 1987; Langolf et al., 1976; MacKenzie et al., 1987; Soechting, 1984). These observations agree with the findings for spatial goal kinematics in the present study. Participants were more accurate when using spatial goal kinematics, and they displayed skewed velocity profiles.
The observed kinematic pattern when participants move towards temporal targets can be compared to studies on kinematics in finger tapping. Participants make movements of shorter duration towards temporal targets and slower movements in the return phase in tapping, regardless of whether the timing target is the flexion or extension movement (Balasubramaniam et al., 2004). Further, intervals preceding forceful taps are shortened, and intervals following them are lengthened (Billon et al., 1996; Semjen & Garcia-Colera, 1986). It could be argued that forceful taps in these studies are represented as major goals, resulting in a more pronounced temporal goal kinematics for the forceful taps, whereas the other taps are considered minor goals. The shortened interval preceding and the lengthened interval following the forceful tap thus seem to be in accordance with the observation of shortened movement times in movements towards temporal targets than in movements to other/no targets. A similar effect is also found when participants have to produce force pulses—larger forces speed up the implementation of the movement (Keele, Ivry, & Pokorny, 1987). Furthermore, in drummers, the duration of the interval after an accented stroke is lengthened (Dahl, 2000). One question in this context is why there is a systematic shortening before and lengthening after the movement towards the temporal goal. One could argue that the shortening–lengthening pattern is intended to enhance the perceptual effects of the goals. However, it could also be the case that participants are systematically biased in the estimation of the time intervals (see Billon et al., 1996, for a discussion of this effect in tapping).
The effects of tempo observed on the variability measures are consistent with the literature: With movements to spatial targets, spatial variability is inversely related to movement time; the variable error in movements to temporal targets is, however, positively related to movement time (Schmidt, Zelaznik, Hawkins, Frank, & Quinn, 1979).
The results of the present study also point to limitations of the cognitive-motor system in meeting both spatial and temporal goals at the same time. Interestingly, with movements towards spatial-temporal targets at slow tempi (use of spatial goal kinematics), temporal performance showed higher variability at the spatial-temporal target not only in comparison to temporal targets (Experiment 4)—an effect that does not seem surprising—but also in comparison to spatial targets (Experiment 3) and no targets (Experiment 4). This was the case even though instructions prioritized temporal targets.
So far, the discussion mainly dealt with the effects of the structural aspects of the tasks (bottom-up influences)—that is, the influence of physical characteristics of targets (spatial, temporal) on performance. However, several aspects in the results suggest that movement kinematics were not only determined by the physically present target features. Instead, combinations of the target features seemed to result in specific goal representations (top-down influences, goal representation hypothesis), which also influenced movement kinematics. One may argue that the result of the combined condition in Experiment 4 (temporal or spatial kinematics depending on tempo) is already evidence for cognitive influences on the movement characteristics. It could be that participants evaluate their ability to meet the task requirements in a given situation and choose their movement kinematics accordingly. However, the results of Experiment 3 and the two temporal target manipulation of Experiment 4 support more unequivocally the goal representation hypothesis. In Experiment 3, at slower tempi, participants showed spatial goal kinematics towards the spatial-temporal target. However, the obtained effect cannot be explained by the presence of the additional spatial target alone—spatial targets were present at both sides of the reversal movement. A similar effect was observed in Experiment 4. Here, temporal goal kinematics were observed for movements towards a spatial-temporal target in comparison to movements towards a temporal target at fast tempi. It seems likely that these effects are observed because one side of the reversal movement is represented as the goal side. Representing certain movement sides as goal sides seems to have a top-down influence on goal-directed kinematic patterns. One mediating mechanism might be attention, which may not be distributed equally between both sides of the reversal movements. Rather, attention may be allocated to a higher degree to the side that is represented as the goal side.
Many classical theories of motor control, dealing with the effect of targets on movement kinematics (e.g., anchoring, e.g., Byblow et al., 1994) can be described as sensory-motor theories. According to these theories, actions or movements are reactions to properties of the current stimulus situation. Sensory-motor accounts would have to predict that movement kinematics can be explained by the combination of target stimuli alone. Thus, sensory-motor accounts are not sufficient to explain the observed data pattern.
The view presented here differs. Movements are viewed not as a reaction to stimuli, but as the means to attain intended goals (ideomotor principle; see James, 1890/1981; Prinz, 1997). The way in which goal representations can influence movement kinematics is by the process of anticipation. Goal representations evoke the anticipation of the desired goal state of the motor system. Movement kinematics are then chosen to achieve this desired goal state. In the present experiments the experience of arriving at a certain position at a certain time represents an action effect. The anticipation of this effect activates the corresponding motor commands. A similar theoretical framework is represented in the concept of internal models (forward and inverse, e.g., Wolpert, Ghahramani, & Flanagan, 2001). A forward model predicts the sensory consequences of a movement; an inverse model predicts motor commands from desired consequences or action effects. Thus, one can argue that the respective motor commands for goal-directed actions are chosen using an inverse model.
To sum up, the data are in support of an ideomotor rather than a sensory-motor view on movement control. Thus, existing models of movement control will ultimately have to include goal representations to fully explain observed movement characteristics.
CONCLUSION
The main goal of the present study was to investigate whether movement kinematics can be viewed as a reaction to target stimuli (position, time), or rather the means to attain intended goals (to be in a certain position at a given time). In order to provide a basis for this question, it was first shown that movement kinematics towards temporal and spatial targets differ from kinematics away from those targets. Further, kinematics are different for movements to temporal (relatively short movement times, high and late peak velocity) and spatial (relatively long movement times, early peak velocity) targets (Experiments 1 and 2). To obtain evidence for the influence of goal representations on kinematics, combinations of temporal and spatial targets were investigated in Experiments 3 and 4. Not only the presence of the physical features of the targets, but also the way in which targets are represented, is important for determining the observed movement kinematics. Results are in support of an ideomotor rather than a sensory-motor view on movement control. Existing models of movement control will ultimately have to include goal representations of participants to fully explain observed movement characteristics. Further research has to investigate whether the present findings on continuous movements generalize to movements in discrete aiming situations.