
Abstract
We report three masked associative priming experiments with the lexical decision task that explore whether the initial activation flow of a visually presented word activates the semantic representations of that word's orthographic/phonological neighbours. The predictions of cascades and serial/modular models of lexical processing differ widely in this respect. Using a masked priming paradigm (stimulus onset asynchrony, SOA = 50 ms), words preceded by ortho-phonologically mediated associated “neighbours” (oveja–MIEL, the Spanish for sheep–HONEY; note that oveja is a phonological neighbour of abeja, the Spanish for bee) were recognized more rapidly than words preceded by an unrelated word prime (Experiments 1 and 3). Furthermore, the magnitude of the ortho-phonologically mediated priming effect (oveja–MIEL) was similar to the magnitude of the associative priming effect (abeja–MIEL). With visible primes and a 250-ms SOA, only the directly associated words showed a priming effect (Experiment 2). These findings pose some problems for a modular account and are more easily interpreted in terms of cascaded models.
Word identification not only involves the coactivation of the orthographic (or phonological) forms of the target … but also, these coactivated word forms activate their corresponding semantic representations in parallel. (Bowers, Davis, & Hanley, 2005, p. 138)
A widespread assumption in the literature on visual-word recognition is that a printed word activates not only its own memory representation in the “lexicon” but also memory representations of words that are orthographically/phonologically “close” to it. Therefore, a key issue for models of visual-word recognition is to determine whether this flow of activation follows a serial pathway based on modular processing, or whether it occurs in a cascaded fashion. Cascaded models that assume an interlevel flow of activation (e.g., Grossberg & Stone, 1986; Harm & Seidenberg, 2004; Plaut, McClelland, Seidenberg, & Patterson, 1996) maintain that coactivated forms at one level (e.g., orthographic) would result in units activated at other processing levels (e.g., phonological, semantic). In this way, these models predict that a given word's semantic links are activated even before this word has been recognized (i.e., before the system reaches equilibrium). In contrast, modular models assume that higher processing levels do not influence the time course of lower processing levels (e.g., see Forster, 1999, 2006). In other words, there is no way of evaluating the semantic properties of the candidates before a lexical entry has been accessed, as one basic tenet of these models is the form-first hypothesis (see Forster & Hector, 2002). Therefore, modular models assume that the correct lexical entry has to be selected among a word's competitors before any semantic information from that entry is accessed (see Boot & Pecher, in press, for a review).
To distinguish between these two approaches, one option is to use a semantic categorization task by manipulating the suitability of a given word (or nonword) with the correct category that is being tested (see Pecher, Zeelenberg, & Wagenmakers, 2005). This was the idea followed by Rodd (2004), who found that semantic representations began to be activated before there was a resolution at the level of form: the word leotards was harder to reject in a semantic categorization task (“Is it an animal?”) than cellar because the first resembled an exemplar, leopards, while the second resembled a nonexemplar, collar. These results converge with the findings reported by Forster and Hector (2002): In an animal categorization task, they found that the nonword turple was harder to discard than the nonword cishop because of the similarity of the first with turtle. Taken together, these data seem to support cascaded models, since the high (semantic) level information interfered with the form similarity manipulation. However, Forster and Hector (2002) suggested a modification of “form-first” modular models to account for those results: the links model (see also Forster, 2006). The links model states that there is an associative link between lexical entries and conceptual structures that might “serve as indicators of possible category membership, but do not provide detailed semantic information” (Forster & Hector, 2002, p. 1115). More specifically, Forster and Hector claimed that “each candidate is subjected to a very limited semantic test, sufficient to establish whether it is likely to be an animal name, but not much else” (p. 1112). Bear in mind that this approach still claims that a word has to be uniquely identified among the arrays of competitors in order to achieve the meaning (e.g., Forster & Shen, 1996; Sears, Lupker, & Hino, 1999); however, it allows certain taxonomical (category) links at the ortho-phonological level that might be activated before lexical selection has been completed (enough to enable category decisions such as living/nonliving but not enough for making decisions based on other features such as size; but see Boot & Pecher, in press).
One way to test Forster and Hector's (2002) modular approach is to examine whether masked associative priming can be found from an ortho-phonological neighbour of the prime (e.g., leotards–SPOTS). Modular models, including the links model, would predict a null priming effect, since the relationship between leotards and spots does not rely on taxonomical shared features. In contrast, cascaded models would predict that the form-to-semantics links could be strong enough to produce an associative priming effect. Thus, the main goal of the present paper is to examine whether ortho-phonologically mediated associative links produce masked associative priming in lexical decision. Interestingly, a masked priming paradigm avoids the potential strategies that may occur in a single-presentation paradigm, such as the semantic categorization task employed by Rodd (2004) and Forster and Hector (2002). Furthermore, the semantic categorization task is replaced in this study by a “nonsemantic” task: lexical decision. Unlike semantic categorization, the lexical decision task does not have an inherent semantic component (or at least, not as strong as a semantic categorization task). Indeed, Rodd (2004) argued that the effects she obtained in the animal categorization task (and also those in the work by Forster and Hector, 2002) could have been due to a contextual priming effect, more than to a direct link between the form level and the semantic level. The reason behind Rodd's argument was that participants could have developed strategic preferences for animal exemplars (leopards) and extrapolated them to words that formally resemble animal exemplars (leotards). Using a masked priming technique avoids these strategic effects and hence represents a stronger test for distinguishing between cascaded and modular models.
While there is some evidence that supports modular models (see Guerrera, 2001; Hino, Lupker, & Pexman, 2002), most empirical evidence has been taken to support cascaded models. However, what we should note is that recent modifications of the form-first models (e.g., the links model) include some taxonomical interlevel semantic links—even though modularity is a basic tenet (see Forster, 2006). Thus, one key test for choosing between modular and cascaded models is to directly study nontaxonomical links between words, such as associative relationships. With a masked priming paradigm, Bourassa and Besner (1998) reported evidence in favour of a continuous flow of activation with nonword primes such as judpe (i.e., a nonword that is perceptually similar to judge) activating higher order representations that were associated to their corresponding base word (judpe–COURT; see also Perea & Lupker, 2003, for similar evidence with transposed-letter nonwords; e.g., jugde–JUDGE). However, one possible criticism of these results is that nonword stimuli do not have lexical representations (e.g., judpe), and the closer attractor for these stimuli would actually be the corresponding real word forms (e.g., judge; see Forster, 2006). A stronger test for cascaded models would be to use words that are phonological neighbours of another word. There is empirical evidence that shows phonologically mediated associative priming (tail activates STORY; e.g., Drieghe & Brysbaert, 2002; Lesch & Pollatsek, 1993; Lukatela & Turvey, 1994): These studies used homophonic and/or pseudohomophonic relationships between words (and nonwords), showing that a masked homophone of a given word (e.g., tail from tale) not only facilitates the recognition of that word (tale), but also activates its associates (STORY). This finding is at odds with a modular conception of lexical access and instead supports a cascaded view that allows some sort of semantic activation starting before the lexical entry has been selected from competitors.
In sum, the above-cited findings are consistent with cascaded models in which activation flows (early in processing) from phonology/orthography to semantics. Nonetheless, homophonic (and pseudohomophonic) relationships involve only a single change at the form/sound level, keeping the phonology constant (as in tail–tale) and just altering the orthographical representations of the words. In the present study, we examine the scope of phonological activation with a more extreme manipulation: We employed ortho-phonological neighbours (comparing male and tale, instead of tail and tale, and their influence on STORY). The phonological neighbourhood of a word is defined as the number of words that can be created by altering only one phoneme from a base word (e.g., gate, hate, and get are phonological neighbours). The phonological neighbourhood can be considered as an analogue to Coltheart's N (i.e., the number of “orthographic” neighbours; see Coltheart, Davelaar, Jonasson, & Besner, 1977). Clearly, in some transparent languages (such as Spanish), it is not easy to disentangle orthographic and phonological neighbours—most phonological neighbours are also orthographic neighbours. For this reason, we refer to the selected stimuli as ortho-phonological neighbours.
It is important to note that, in prior research, the activation of the associatively mediated word (tail–STORY or judpe–COURT) only occurs when the prime stimulus is presented under masked priming conditions, but not with longer exposures of the prime (see Bourassa & Besner, 1998; Drieghe & Brysbaert, 2002; Lesch & Pollatsek, 1993). These findings have usually been interpreted in terms of an activation–verification account (Paap, Newsome, McDonald, & Schvaneveldt, 1982; see also Grossberg & Stone, 1986): At long (conscious) exposures of the primes, the automatic activation that can be found under masked priming conditions disappears, and only the correct candidate may remain active, the reason being that the verification procedure of the target word has been completed.
In Experiment 1, we examined to what extent the ortho-phonological neighbours of a word facilitate access to the word's associative information in a lexical decision task with a masked priming procedure (stimulus onset asynchrony, SOA = 50 ms). Specifically, a given word (e.g., MIEL; the Spanish for honey) could be preceded by an associatively related prime (e.g., abeja; the Spanish for bee), by an ortho-phonological neighbour of the associatively related prime (e.g., oveja; the Spanish for sheep), or by an unrelated prime. Cascaded models would predict some facilitation not only from the associative pairs but also from the ortho-phonological neighbours (namely, the mediated associates). In contrast, if the associative links in lexical entries only report information of the category membership of a neighbour (as predicted by a form-first model such as the links model; see Forster, 2006; Forster & Hector, 2002), then no influence from the mediated associates would be expected. To anticipate the results, we found an advantage of ortho-phonologically mediated associations over the unrelated condition—as well as an associative priming effect. In Experiment 2 we used visible primes (and an SOA of 250 ms) to examine whether the effect of ortho-phonologically mediated associations vanishes when the prime words have been verified (and the nontarget competitors presumably have been discarded). To avoid any strategic effects that may occur with visible primes, we included a number of unrelated filler pairs, thus keeping the proportion of related pairs low. Finally, Experiment 3 was a replication of Experiment 1 using the same relatedness proportion as that in Experiment 2.
Experiment 1: Masked Primes
Method
Participants
A total of 69 students from the University of La Laguna took part in the experiment in exchange for course credit. All of them were native speakers of Spanish and had either normal or corrected-to-normal vision.
Materials
A total of 60 associatively related pairs (e.g., abeja–MIEL; the Spanish for bee–HONEY) were selected from the Spanish free-association norms (Fernández, Díez, Alonso, & Beato, 2004), with the first member of the pair used as a prime and the second as target. For the selected pairs, the mean associative strength (i.e., the first associative response to the prime) in these norms was 36%. Each target word could be preceded by: (a) an associatively related prime (e.g., abeja–MIEL); (b) a prime that shared with the associated prime all the phonemes except for one (e.g., oveja–MIEL, the Spanish for sheep–HONEY; note that in Spanish /b/ and /v/ correspond to the same phoneme, and therefore abeja and oveja only differ in a single phoneme); or by (c) a completely unrelated prime word (e.g., cloro–MIEL, the Spanish for chlorine–HONEY). The characteristics of the pairs used in the experiment are presented in Table 1. None of the words in the phonologically mediated and unrelated conditions was associatively (or semantically) related to its corresponding target word. (See the Appendix for a complete list of stimuli.) For the purposes of the lexical decision task, we created a set of 60 legal (pronounceable) target nonwords of similar length to the target words. A set of 60 Spanish words that were not used in any of the previous conditions was selected to prime these nonwords. Three lists of items were created, so that each target appeared only once in each list, but in a different priming condition (associatively related, ortho-phonologically mediated associated, or unrelated). Different participants were assigned to each list.
Mean word frequency, length, and number of orthographic and phonological neighbours of the word–word pairs used in the experiments
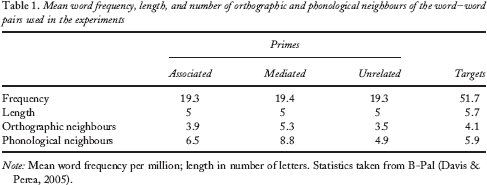
Note: Mean word frequency per million; length in number of letters. Statistics taken from B-Pal (Davis & Perea, 2005).
Procedure
The experiment was run individually in a quiet room. Presentation of the stimuli and recording of response times were controlled by PC-compatible computers. The experiment was run using DMDX (Forster & Forster, 2003). On each trial, a forward mask consisting of a row of hash marks (#s) was presented for 500 ms in the centre of the screen. Next, the prime was presented for 50 ms (3 cycles; each cycle corresponding to 16.6 ms on the CRT monitor). The prime was followed immediately by the presentation of the target stimulus in upper case. Reaction times were measured from target onset to the participant's response. All the strings were presented centred, in Courier New 12-point font coloured in white, on a black background. Participants were instructed to press the “M” button if the string formed an existing Spanish word and the “Z” button if the string was a nonword. They were not informed of the presence of lower-case items. Each participant received a different order of trials. None of the participants reported having seen the lower-case words when asked after the experiment. The whole experimental session lasted for about 6 minutes.
Results and Discussion
Incorrect responses (4% of the data) and responses beyond the 250–1,500-ms cut-off values (2.4% of the data) were excluded from the latency analyses. Mean response times and percentages of errors are presented in Table 2. Planned comparisons were conducted to assess the effect of associative priming (i.e., the difference between the unrelated condition and the associative condition) and the effect of phonologically mediated associative priming (i.e., the difference between the unrelated condition and the phonologically mediated associative condition). 1 List (List 1, List 2, List 3) was included as a dummy variable in the analyses to extract the variance due to the error associated with the lists.
We only report F ratios over participants. This is the appropriate analysis for testing the significance of the effects in a counterbalanced design, such as that used in the present study (see Clark, 1973; Raaijmakers, 2003; Raaijmakers, Schrijnemakers, & Gremmen, 1999). In any case, the p values corresponding to the F2 ratios in the present experiments essentially mimicked the reported p values of the F1 ratios.
Mean lexical decision times and percentage of errors for word targets in Experiments 1–3
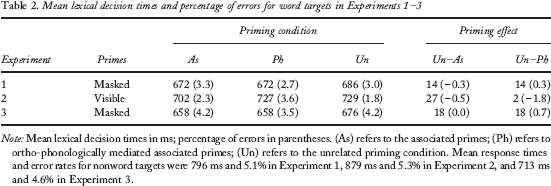
Note: Mean lexical decision times in ms; percentage of errors in parentheses. (As) refers to the associated primes; (Ph) refers to ortho-phonologically mediated associated primes; (Un) refers to the unrelated priming condition. Mean response times and error rates for nonword targets were 796 ms and 5.1% in Experiment 1, 879 ms and 5.3% in Experiment 2, and 713 ms and 4.6% in Experiment 3.
Words preceded by an associatively related prime were responded to 14 ms faster than the words preceded by an unrelated prime, F(1, 66) = 6.82, MSE = 982.57, p < .02, and, more important for the present purposes, words preceded by a phonologically mediated associated word were also responded to 14 ms more rapidly than words preceded by an unrelated prime, F(1, 66) = 8.92, MSE = 702.28, p < .01. Obviously, no differences were found when comparing the associated and the phonologically mediated associated priming conditions (less than 1 ms), F < 1.
The analyses of variance (ANOVAs) on the error rates did not show any significant differences (all Fs < 1).
The results are straightforward: Associated word pairs and ortho-phonologically mediated associated word pairs produced exactly the same pattern of priming when compared to an unrelated, baseline condition. The percentage of pairs that showed a priming effect was 75% for the associative pairs and 72% for ortho-phonologically mediated associative pairs.
As indicated in the Introduction, most phonological neighbours in Spanish are also orthographic neighbours. To examine the relationship between semantics and phonology (rather than orthography), we conducted a post hoc analysis to examine the priming effect with phonological neighbours that are also orthographic neighbours (e.g., plano and piano) and phonological neighbours that are not orthographic neighbours (e.g., abeja and oveja). A total of 19 of the 60 pairs were exclusively phonologically related (i.e., they were not orthographic neighbours), whereas the rest were both orthographic and phonological neighbours. This post hoc analysis showed a facilitative effect of 18 ms for the exclusively phonologically mediated pairs, which was slightly higher than the priming effect obtained with the ortho-phonologically mediated pairs (an 11-ms effect). In sum, the observed priming effect does not seem to be restricted to mere orthographic relationships.
Experiment 2: Visible Primes
The results from Experiment 1 suggest that associatively related words are activated even before the phonological and orthographical encoding of a given word is completed. In Experiment 2, we examine whether a word's ortho-phonological neighbours activate an associatively related target word at a 250-ms SOA. An activation–verification account would predict that when the presented word is consciously processed, then the initially coactivated candidates would not be disruptive: Only the correct associative links would remain active, since the indirect or incorrect candidates would have been already verified and rejected (see Bourassa & Besner, 1998; Drieghe & Brysbaert, 2002; Lesch & Pollatsek, 1993).
Method
Participants
A total of 33 students from the University of La Laguna participated in the experiment for course credit. 2 None of them had taken part in Experiment 1.
The smaller number of participants in Experiment 2 (compared with Experiments 1 and 3) is due to the fact that associative priming can be easily obtained with visible primes. Instead, associative priming is smaller with masked primes, and hence, it is necessary to increase sample size (e.g., 132 participants in the masked priming experiments of Bourassa & Besner, 1998; or 96 in Lesch & Pollatsek, 1993).
Materials
The experimental trials were the same as those in Experiment 1. However, as the prime words were now visible, and to avoid any strategic/predictability effects, we included in the experiment 80 word–word pairs that shared no form/semantic relationship. We also added 80 pronounceable nonwords (primed by 80 unrelated new words) for the purpose of having 50% of word/nonword trials. Consequently, the percentage of associatively related pairs comprised less than 8% of the trials. As in Experiment 1, three lists were created, and each target appeared only once in each list, but each time in a different priming condition.
Procedure
The procedure was the same as that in Experiment 1, except that: (a) the mask (row of hash marks) was substituted by a fixation point (“ + ”), and (b) the time exposure of the prime was of 250 ms (15 cycles of 16.6 ms each). The experimental session lasted for around 13 minutes.
Results and Discussion
Incorrect responses (3.5% of the data) and response times beyond 250–1,500 ms (5% of the data) were excluded from the latency analyses. Mean response times and percentages of errors are summarized in Table 2. Planned comparisons were conducted to assess the effect of associative priming and the effect of phonologically mediated associative priming.
Words in the associated condition were responded to 27 ms faster than those in the unrelated condition, F(1, 30) = 12.94, MSE = 973.18, p < .01. In addition, phonologically mediated associated pairs showed very little benefit when compared to the unrelated condition (a negligible 2-ms effect), F(1, 30) = 0.12, MSE = 1,170.84. Indeed, the associative pairs showed faster response times than do the phonologically mediated associative pairs (a 25-ms difference), F(1, 30) = 9.30, MSE = 1,170.84, p < .01.
Comparisons of the error rates showed that words in the phonologically mediated associated condition were responded to less accurately than words in the unrelated condition, F(1, 30) = 5.41, MSE = 10.08, p < .03. The other comparisons were not significant (both ps > .15).
The results of the present experiment are clear-cut. Only the associative condition (but not the ortho-phonologically mediated condition) showed a benefit relative to the unrelated condition. Thus, with a prime exposure of 250 ms, when the ortho-phonological encoding of the prime has (presumably) been successfully completed, the form-based mediations exert no facilitative influence, and only the correct candidate remains active after the verification processes. What is more, if we look at the error data, mediated primes exerted some small inhibition on the recognition of the target word, which is in line with previous evidence showing that at long prime exposures, orthographic effects become inhibitory rather that facilitative (see Napps & Fowler, 1987).
Experiment 2: Masked Primes + Low Relatedness Proportion
Experiment 1 showed a significant masked priming effect for both associated word pairs (abeja–MIEL) and ortho-phonologically mediated, associated word pairs (oveja–MIEL). In Experiment 2, with visible primes and a 250-ms SOA, we only found a priming effect for directly associated word pairs. Leaving aside the SOA, one potential difference between these two experiments was that Experiment 2 used a number of filler pairs so that the proportion of related pairs was kept low. In a recent study, Bodner and Masson (2003) found that relatedness proportion could modulate the magnitude of the masked associative priming effects: Effects were greater when the proportion of associative pairs was high (but see Perea & Rosa, 2002). The goal of Experiment 3 is to reexamine the findings from Experiment 1 with the same materials as those in Experiment 2 (i.e., keeping a low proportion of related pairs). 3
We thank Ken Forster and two anonymous reviewers for suggesting this experiment.
Method
Participants
A total of 51 students from the University of La Laguna took part in this experiment. They were paid 5€ for their participation. None of them had participated in Experiments 1–2.
Materials
The materials were the same as those in Experiment 2.
Procedure
The procedure was the same as that in Experiment 1.
Results and Discussion
Incorrect responses (less than 4% of the data) and response times above 1,500 ms or below 250 ms (2.1% of the data) were excluded from the latency analyses. Mean response times and percentages of errors are presented in Table 2. As in Experiments 1–2, planned comparisons were conducted to assess the effect of associative priming and the effect of ortho-phonologically mediated associative priming.
Words preceded by an associatively related prime were responded to 18 ms faster than words preceded by an unrelated prime, F(1, 48) = 10.14, MSE = 791.48, p < .01, and words preceded by an ortho-phonologically mediated associated prime were also read 18 ms faster than words primed by a completely unrelated word, F(1, 48) = 4.34, MSE = 1,963.61, p < .05. There were no signs of a difference between the associatively related and the ortho-phonologically mediated conditions (less than 1 ms), F < 1.
The analyses on the error data did not reveal any significant effects (all ps > .25).
The results are again straightforward. When briefly presented and masked, prime words associatively related to the target word (e.g., abeja–MIEL) exerted the same facilitation in the target recognition as ortho-phonologically mediated associated prime words did (e.g., oveja–MIEL). Thus, we successfully replicated the findings from Experiment 1, this time using a low proportion of related pairs. As reported by Perea and Rosa (2002), the magnitude of masked associative priming was not altered by the proportion of associatively related pairs—that is, the observed masked priming effects appear to be automatic in nature.
General Discussion
The main findings of the present lexical decision experiments can be summarized as follows: (a) masked associative priming occurs not only with directly associated words (e.g., abeja–MIEL; bee–HONEY), but also with ortho-phonologically mediated words (e.g., oveja–MIEL; sheep–HONEY); (b) this pattern of data is not modulated by the proportion of associatively related pairs in the list; and (c) when the primes are presented for 250 ms, the associative priming effect is robust for the directly associated prime words, while it vanishes for the ortho-phonologically mediated prime words. Taken together, the present findings have clear implications for models of visual word recognition.
Experiments 1 and 3 replicate previous findings in the literature on masked associative priming, showing a fast activation of the directly associated candidate of a given word (e.g., nurse facilitates the recognition of DOCTOR compared with the unrelated word butter; e.g., Bodner & Masson, 2003; Perea & Gotor, 1997). Furthermore, these effects appear to be automatic in nature: The size of the associative priming effect was similar independently of the proportion of associatively related words (see Pecher, Zeelenberg, & Raaijmakers, 2002; Perea & Rosa, 2002, for converging evidence; but see Bodner & Masson, 2003). But the novel result from Experiments 1 and 3 is that this priming effect occurs not only with directly associated primes (abeja–MIEL), but also with an ortho-phonological neighbour of the associated prime that shared no semantic relationship with the target word (oveja–MIEL). This finding strongly suggests that, at a very early stage of lexical access, both primes (abeja and oveja) produce a similar activation at a higher processing level (i.e., semantic level) when compared to the unrelated condition. That is, the flow of activation seems to be equally distributed so that all candidates initially activated by a given word activate their corresponding associated words. These results add support for cascaded models, that (as stated by Bowers, 2002, p. 422) assume that “incorrect generalizations tend to be produced (e.g., activating the semantics of CHAIR when the input is CHAID)”. Importantly, the present results represent a step forward with respect to this assumption in two ways. First, it should be noted that these “incorrect generalizations” have been typically tested in tasks where the semantic component is an inherent element in the task (e.g., living/nonliving decisions or size decisions; see Boot & Pecher, in press). In the present experiments, we found a parallel pattern in lexical decision—this suggests that the underlying processes may be common to normal silent reading. Secondly, as stated in the Introduction, the strongest attractor for a nonword stimulus like CHAID would be the base word CHAIR, and hence the closest semantic representation for the input stimulus would be the semantics of the closest real word. Thus, even though there is some evidence from lexical decision experiments in which the semantics of a nonpresented item are activated (e.g., Bourassa & Besner, 1998, or Perea & Lupker, 2003, using nonword primes; e.g., judpe–JUDGE or jugde–JUDGE), the present study represents a stronger test for this effect, since the mediated primes were always words and therefore had their own lexical/semantic representation. Furthermore, by employing a more extreme manipulation than with homophones/pseudohomophones (note that abeja and oveja only share 3 of 5 letters and 4 of 5 phonemes, unlike homophonic pairs like tail–tale in tale–STORY), we have shown that at early stages of visual encoding, not only phonologically equal words (homophones) activate each other's semantic representations, but also ortho-phonological neighbours. Thus, the present findings can be taken as strong evidence in favour of cascaded models (e.g., Grossberg & Stone, 1986; Harm & Seidenberg, 2004; Plaut et al., 1996).
What are the implications of the present findings for modular models? Forster and Hector (2002) hypothesized a form-first modular framework (e.g., the entry-opening model; see Forster, 1999), which could be modified to account for our findings in the semantic categorization task (and also for the results in Rodd's work, 2004). Their modification was based on the acceptance of a category membership link between lexical entries and conceptual structures (the Links model; see Forster, 2006, for further evidence). Such a link would be strong enough to activate a neighbour's taxonomical categories, “but not much else” (Forster & Hector, 2002, p. 1112). We believe that the present data challenge this explanation, since we found strong noncategorical associations in a lexical decision task. As indicated above, in a modular model (e.g., Forster & Hector, 2002), lexical items would be related by their form and by the co-occurrence in language with no interactivity with the semantic information. In order to account for the present findings, a modular model would need to assume that information about the patterns of co-occurrence (hence, associative relationships between words) of lexical items may occur at the form level (see Forster, 2006). However, this modification—which would be akin to cascaded models—is not really in the spirit of a modular model. Furthermore, the presence of neighbourhood congruency effects on semantic categorization tasks involving living decisions and size decisions (Boot & Pecher, in press) would still be unaccounted for. Future research should be directed to test whether purely semantic relationships can produce similar results in a nonsemantic task (e.g., will pug prime wild boar the same as pig would in a lexical decision task?).
One finding that deserves some comment is that the magnitude of the priming effect was similar for associative pairs and for ortho-phonologically mediated associative pairs. On a priori grounds, one would have expected to obtain a greater priming effect for the directly associated pairs. There are two possible explanations for this, which we now examine. First, when a word is so briefly presented (namely, for 50 ms) and masked, there is a high degree of noise in the letter detection system, and rather than activating uniquely the correct lexical entry (e.g., oveja), other formally similar entries are also highly activated (e.g., abeja; see Carreiras, Duñabeitia, & Perea, 2007; Perea, Duñabeitia, & Carreiras, in press). To test this possibility, we conducted a masked identity priming experiment with the targets used in Experiment 1. The results showed a greater priming effect for the identity primes (50 ms, abeja–ABEJA) than for the ortho-phonological neighbours (24 ms; oveja–ABEJA) relative to the unrelated condition. Thus, at least at a form/word level, abeja activates ABEJA more than oveja does, and this poses some problems for the explanation based on a noisy recognition system. Secondly, the lack of a difference in terms of associative priming between abeja–MIEL and oveja–MIEL may be due to strong (and, to some degree, similar) cascaded coactivation of the associative/semantic representations of a word and its orthographic/phonological neighbours. This is in line with the conception of a distributed cascaded-like associative activation that produces similar semantic activation levels for all the initially preactivated word forms. This finding is consistent with the presence of similar priming effects for word–word pairs (judge–TRIAL; 14 ms) and nonword–word pairs (jugde–TRIAL; 12 ms) found by Perea and Lupker (2003). Likewise, Perea, Duñabeitia, and Carreiras (2007) found that, for highly fluent bilinguals, masked associative priming was remarkably similar within languages and between languages (21 vs. 18 ms across two experiments)—on a priori grounds one would have predicted a stronger associative priming effect within a language. Clearly, differences between differences in masked priming experiments are bound to be small, and one might require a huge experiment to obtain a reliable difference. (Note that masked associative priming effects are usually small even for strong associates.) Thus, one might argue that the fact that the magnitude of the priming effect for associative pairs does not differ significantly from ortho-phonologically mediated associative pairs is probably a question of experimental power. Although the experimental power of the present set of experiments was high, it may not be high enough to capture these very small differences. More research is needed to clarify the source of this finding.
What is the role of prime exposure duration? In Experiment 2, we found that the prime exposure duration was decisive for the ortho-phonologically mediated associative relationship to remain active. With a 250-ms SOA, only the associative related pairs showed a benefit relative to the unrelated condition. This is consistent with prior studies with homophones and pseudohomophones that show that, when the primes were visible, there is no longer an effect from phonologically mediated associative pairs (e.g., Drieghe & Brysbaert, 2002; Lesch & Pollatsek, 1993; Lukatela & Turvey, 1994; see also Bourassa & Besner, 1998, for a purely orthographic manipulation using nonword stimuli). That is, the candidates that have been rejected “are no longer active in the word recognition system because a failed verification inhibits them” (Bourassa & Besner, 1998, p. 63). The present study extends this pattern of data to ortho-phonological neighbours in a transparent orthography (Spanish). Thus, these findings are consistent with the view that cascaded activation is mainly an automatic and ballistic rapid stream that occurs at the initial stages of lexical access, and that conscious processing of a word restricts the activation to only the correct candidate by following a verification procedure.
In summary, the present paper provides strong support for the idea in the quotation that opened this paper (see Bowers et al., 2005): There is spreading activation that flows throughout the semantic level even before a given lexical entry has been fully activated. Later in processing, when the correct candidate is selected, only its semantic links remain active, as predicted by an activation–verification framework (e.g., see Grossberg & Stone, 1986). The present findings support cascaded models rather than serial/modular models and also shed some light on how lexical access is achieved.