
Abstract
In contextual cueing, the position of a search target is learned over repeated exposures to a visual display. The strength of this effect varies across stimulus types. For example, real-world scene contexts give rise to larger search benefits than contexts composed of letters or shapes. We investigated whether such differences in learning can be at least partially explained by the degree of semantic meaning associated with a context independently of the nature of the visual information available (which also varies across stimulus types). Chess boards served as the learning context as their meaningfulness depends on the observer's knowledge of the game. In Experiment 1, boards depicted actual game play, and search benefits for repeated boards were 4 times greater for experts than for novices. In Experiment 2, search benefits among experts were halved when less meaningful randomly generated boards were used. Thus, stimulus meaningfulness independently contributes to learning context–target associations.
Contextual cueing is a learning effect where repeated exposure to a specific arrangement of target and distractor items leads to a progressively more efficient search. In these learning paradigms, intermixed among novel displays, a subset of stimuli are consistently repeated where the positions of the target and other objects in the display are fixed. Over repetitions, this covariation is learned and used to guide visual attention, causing search times for repeated displays to progressively decrease at a faster rate than for novel displays (which often improve as the search task becomes more familiar). This effect has been demonstrated across a wide range of stimuli from arrays of letters (e.g., Chun & Jiang, 1998), to configurations of novel shapes (e.g., Chua & Chun, 2003; Chun & Jiang, 1999), to real-world scenes (e.g., Brockmole, Castelhano, & Henderson, 2006; Brockmole & Henderson, 2006a, 2006b) and has been described as one means by which humans are able to cognitively reduce the complexity of their visual environments and efficiently interact with the world despite rather strong limitations on attention and memory (Chun, 2003).
Although observers are able to learn associations between objects and their surrounding contexts, the speed with which context–target associations are learned and the magnitude of the learning benefit depend on the stimuli used as the search context. For example, with letter arrays contextual cueing develops over dozens of repetitions and tends to be less than 100 ms in magnitude (see, for example, Chun & Jiang, 1998). In contrast, when real-world scenes are used as few as four repetitions are needed to observe maximal learning benefits whereupon the magnitude of the learning effect has been observed to be more than 2,000 ms (Brockmole & Henderson, 2006b). These differences indicate that the strength of the contextual cueing effect is sensitive to the kind of information provided in different search contexts. Determining the relative importance of different types of information that are used to reference target position in these search paradigms is therefore a critical issue with regard to contextual cueing specifically and the development of behaviourally functional scene memory generally.
It is clear that contextual cueing involves learning associations between consistently mapped visual features as the extent of cueing depends on the ease with which repeating visual features can be associated with a specific target. For example, the visual similarity of, and spatial distance between, targets and context-defining distractors (Jiang & Chun, 2001; Olson & Chun, 2002), as well as the proportion of a visual display repeated across viewing encounters (Brockmole et al., 2006) all affect contextual cueing. Thus, a host of low-level visual differences across classes of stimuli might lead to differences in how targets are associated with visual information available in a display. However, it is also true that the types of stimuli that have been used in contextual cueing paradigms vary in the amount of semantic information they contain, with, for example, scenes providing much more semantic information than arrays of letters or shapes. Outside of contextual cueing paradigms, initial semantic representations of scene structure (Castelhano & Henderson, 2007) and semantic knowledge of the likely locations of objects (Eckstein, Drescher, & Shimozaki, 2006; Henderson, Weeks, & Hollingworth, 1999; Neider & Zelinsky, 2006; Torralba, Oliva, Castelhano, & Henderson, 2006) have been shown to increase search speed. 1 Thus, in addition to the extent and quality of the visual information available in a search display, abstract semantic information may also be important in contextual cueing.
In addition, recent research has investigated whether semantically inconsistent objects in scenes initially draw the eyes, with some results suggesting that they do (e.g., Becker, Pashler, & Lubin, 2007; Underwood & Foulsham, 2006) and some suggesting that they do not (e.g., De Graef, Christiaens, & d'Ydewalle, 1990; Gareze & Findlay, 2007; Henderson et al., 1999). Despite this controversy, what is not in dispute is the role of scene context in helping to direct the eyes to locations likely to contain the target of a current search based on the physical and semantic constraints imposed by scene layout.
In the present study, we directly examined for the first time whether contextual cueing is affected by the degree of semantic meaning available in the search context. Because visual and semantic richness are confounded when one compares performance across different types of search displays (e.g., arrays of shapes versus scenes), in order to directly test the role of meaning in contextual cueing we needed to develop a new search paradigm in which we could manipulate the meaningfulness of the search display without varying any of the display's visual aspects. To accomplish this, we used configurations of playing pieces on chess boards as the search context. At any given moment during a game, the arrangement of pieces on a board bears on the outcome of the game and is thus meaningful. However, the meaningfulness of an arrangement of chess pieces to an individual observer is directly related to that observer's knowledge of the game of chess (Chase & Simon, 1973a, 1973b). For a novice player who understands very little about chess, the arrangement of pieces on a board is not particularly meaningful, and the chess board is analogous to an array of letters or shapes. For an experienced chess player, however, piece arrangements are meaningful and are more analogous to scenes. Thus, we examined the effect of stimulus meaning in contextual cueing by manipulating the knowledge of the observer rather than the properties of the stimuli.
In Experiment 1 we show that chess experts learned context–target associations over four times faster than did novices. In a second experiment we show that within the expert group, search benefits were halved when less meaningful randomly generated boards served as the search context compared to boards that depicted game play. In order to conduct both experiments with a very limited pool of highly skilled chess experts, however, two constraints were imposed on our methods. First, we were unable to address whether learning effects were implicit or explicit in nature, as is often done in contextual cueing research. The results of Experiment 2 would have been fatally contaminated had we included this assessment because participants would no longer have been naïve with respect to the experimental hypotheses. Second, we were not able to counterbalance the order in which experts completed each experiment. However, we took several precautions (discussed later) to ensure that any transfer effects across experiments were small compared with the effect of learning within each experiment. Despite these limitations, the results provide the first clear evidence that one's ability to extract meaning from visual displays serves a functional role in the development of memory for context–target associations and the efficiency with which visually guided tasks such as search are completed.
Experiment 1
Chess experts and novices searched for and identified a target letter embedded in illustrations of chess boards. These letters were used as search targets because their position could not be predicted a priori from the layout of the boards. Amid a series of filler trials on which a board was presented that had not been shown previously, a set of boards was consistently repeated. The search target was always located in the same position on these repeated boards. Therefore, for repeated boards the arrangement of chess pieces perfectly predicted target location (but not identity). The stimuli shown to experts and novices were identical, and so no differences in visual characteristics of the stimuli existed across the groups. Of interest was whether and to what extent contextual cueing under this situation is affected by level of expertise with the search context. If analysis of meaning plays a role in learning context–target associations, then experts should exhibit contextual cueing effects that develop faster and are larger in magnitude than those observed by novices.
Method
Participants
Two groups of participants were recruited. The novice group was composed of a sample of 10 undergraduate students at the University of Edinburgh (average age: 21 years). Responses on a postexperiment questionnaire (see Appendix) indicated that while these participants generally had a basic understanding of the rules of chess, they had little knowledge of chess strategy and infrequently played the game. The expert group was composed of 5 individuals rated among the top 100 chess players in Scotland (at the time of their participation, the average national rank of the sample was 65th; average Fédération Internationale des échecs, FIDE, rating of the sample was 2041) and were students or recent graduates of the University of Edinburgh (average age: 23 years).
Stimuli and apparatus
Stimuli consisted of 248 greyscale illustrations of chess boards containing 16–18 pieces reproduced from various books of game-play (see Figure 1 for an example). A total of 8 boards were randomly selected to be the repeated boards, which were presented multiple times throughout the experiment. The remaining 240 boards constituted the novel boards, which were each shown exactly once during the experiment. The boards subtended 433 pixels both horizontally and vertically on a 19′ cathode ray tube (CRT) with a display resolution of 800 × 600 pixels. A single grey “T” or “L” target letter subtending 5 pixels horizontally and vertically was digitally inserted into each board. The shade of this letter was adjusted so that its contrast against its background (a white or grey square) was equated. The letter was always placed in one of the corners of a square that contained a playing piece. Because participants began each trial looking at the centre of the board, the centre four squares never contained targets. Across the novel boards, the target was equally likely to appear in any eligible square, and “T” and “L” targets occurred with equal frequency within each square. For the repeated trials, eight target positions were randomly selected under the constraint that they were within a square containing a playing piece. Although the location of each target was held constant within each repeated board, the target's identity was randomly determined with each presentation.

Left: Example stimulus from Experiment 1; the positions of the pieces are consistent with actual game-play. Right: Example stimulus from Experiment 2; a modified version of the left stimulus where the pieces and their positions were randomly selected save for the piece occupying the same square as the target. In both stimuli the target is a small grey T in the lower left corner of Row 1, Column 2 (occupied by a black knight; see inset for detail).
Design and procedure
Participants viewed 480 stimuli, divided into 30 blocks of 16 trials. Each block contained 8 novel trials and 8 repeated trials. At the beginning of each trial, observers were instructed to look at a dot in the centre of the display and to press a key when ready to view the chess board. The observers’ task was to search for and identify a target letter (T or L) embedded in the board. Upon identifying the target, observers pressed one of two response buttons. Search accuracy and response time were recorded. The trial was terminated if a response was not made within 20 s of scene onset. No information regarding the block structure or the repetition of boards was given to observers.
Results and Discussion
Trials were excluded from analysis if a response was not made (4% for novice group, 6% for expert group) or was incorrect (1% for each group). Novel trials were further trimmed by excluding those trials on which search times were greater than 2.5 standard deviations from the mean as calculated on a subject-by-subject basis (2% for each group). Repeated trials were not subjected to the standard deviation trim because the variance in search time was expected to vary across blocks, and, given only 8 trials per block, an accurate estimation of the variance within each cell was not possible (see Brockmole & Henderson, 2006b). To increase statistical power, we collapsed the 30 learning blocks into 6 epochs. Analyses focused on the effects of epoch and expertise on search time.
Table 1 summarizes search times for novel and repeated trials as a function of expertise and epoch. A 2 (expertise) × 6 (epoch) mixed-model analysis of variance (ANOVA) first considered only the novel trials as they establish the baseline search rate for each group across epochs. Search times for novel trials decreased over epochs, F(5, 65) = 4.02, p < .01, indicating that the search task became easier with practice. Experts, however, exhibited longer search times than novices, F(1, 13) = 15.5, p < .01, an effect that did not vary over epochs, F(5, 65) = 1.41, p = .23, demonstrating that this difference in baseline search rates was independent of the repetition of configurations and any incentive to memorize those configurations. Although a complete account of this potentially counter-intuitive effect is beyond the scope of this report, we note that at debriefing, experts reported that it was difficult for them to focus on the search task without also analysing the game situations depicted. This explanation for overall slower baseline search times among experts is consistent with previous experimental work suggesting that chess experts involuntarily and automatically process relationships among chess pieces (Reingold, Charness, Schultetus, & Stampe, 2001), although other explanations may also be possible.
Mean search time broken down by expertise, trial type, and epoch
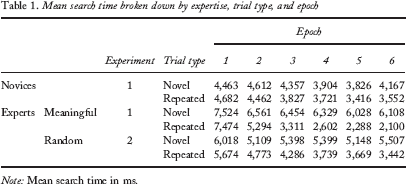
Note: Mean search time in ms.
In the context of the present study, because the groups differed in baseline search speed, for our remaining analyses, within each group we calculated a cueing effect based on a proportional measure where the difference in search time between the novel and repeated trials was divided by the search time observed for novel trials. So, for example, a cueing effect of .50 indicates that search times for repeated trials were half those observed on novel trials. Cueing effects are plotted in Figure 2, broken down by expertise and epoch. One-way repeated measures ANOVA was conducted within each group to demonstrate that the cueing effect increased across epochs for both groups: novice group, F(5, 45) = 2.84, p < .05; expert group, F(5, 20) = 17.0, p < .001. Having established that both novices and experts were sensitive to the covariation that existed between the arrangement of playing pieces on a chess board and the location of a target, a 2 (expertise) by 6 (epoch) mixed-model ANOVA compared the cueing effects for the novice and expert groups. As expected, a main effect of epoch was observed, F(5, 65) = 18.0, p < .001, as the magnitude of the cueing effect increased over epochs. Importantly, a main effect of expertise was also observed, F(1, 13) = 29.4, p < .001, as the magnitude of the cueing effect was, on average, greater for the expert group. The interaction of these factors was also reliable, F(5, 65) = 6.58, p < .001, as the cueing effect increased at a faster rate over epochs for the expert group. Linear regression analyses indicated that while the cueing effect for the novices increased by 3% per epoch, the rate for experts was 13% per epoch.
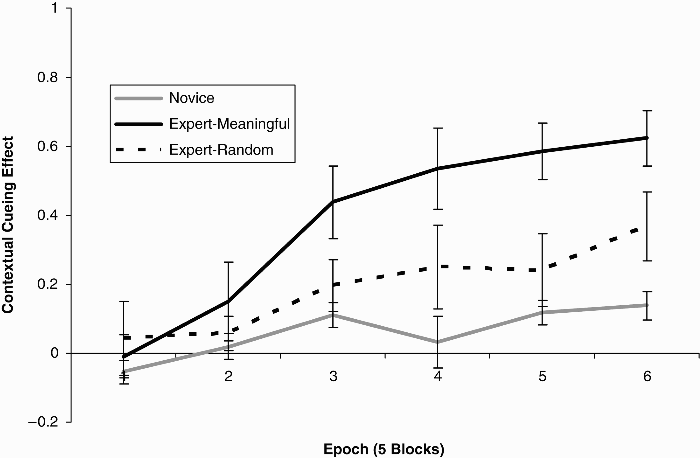
Results of Experiments 1 and 2: Average cueing effects (with standard errors) for the expert and novice groups broken down by epoch. The expert group is further broken down by the meaningfulness of the board layouts (i.e., real game positions or random positions).
In addition to using a proportional measure of cueing in order to control for baseline search rates between the novice and expert groups, we obtained converging evidence that the groups differed in their rates of learning despite their baseline search differences by extracting subsamples of 3 novices and 3 experts that resulted in equivalent baseline search times (experts: 5,501 ms; novices: 5,293 ms). A 2 (expertise) × 6 (epoch) mixed-model ANOVA demonstrated that the cueing effect increased over epochs, F(5, 20) = 8.93, p < .001, and that this increase differed across the novice and expert subgroups, F(5, 20) = 5.24, p < .01. For novices, the cueing effect increased from –.051 to + .137 (cf. –.055 to + .138 for the entire novice sample). For experts, the cueing effect increased from –.063 to + .554 (cf. –.010 to + .623 for the entire expert sample).
These results of Experiment 1 suggest that expertise in a given search context affects the rate at which context–target associations are learned and the magnitude of the resulting learning benefit. Given exactly the same stimuli, chess experts learned the association between a target's location and a predictive context (defined by an arrangement of chess pieces) over four times faster and to a benefit over four times greater than that observed for chess novices. Furthermore, experts’ greater contextual cueing benefits more than compensated for overall slower baseline search rates as their search speed for repeated trials was faster than that for novices from Epoch 3 onward (see Table 1). In this experiment, the arrangement of playing pieces on each board illustrated moments in game play that are more meaningful to experts than to novices, suggesting that stimulus meaning plays an independent role in contextual cueing. In Experiment 2, we further examine the role of meaning by presenting chess experts with boards illustrating random arrangements of pieces, thereby reducing their meaningfulness.
Experiment 2
One's ability to learn context–target associations depends on his or her ability to recognize previously encountered contexts. Previous investigations of memory for chess boards have demonstrated that recall and recognition advantages observed for experts are reduced, and sometimes eliminated, when piece positioning is randomized (e.g., Chase & Simon, 1973a, 1973b; Gobet & Clarkson, 2004; Gobet et al., 2001; Gobet & Simon, 1996; Gobet & Waters, 2003; Goldin, 1979; Huffman, Matthews, & Gagne, 2001; Saariluoma, 1994; Schneider, Gruber, Gold, & Opwis, 1993). Poorer memory for random board layouts among chess experts has been linked to a reduced ability to extract meaningful patterns from the stimuli, resulting in a reduced ability to efficiently encode the array of playing pieces (Chase & Simon, 1973a, 1973b) or to use long-term memory for previously encountered patterns (DeGroot, 1965; Gobet et al., 2001).
In Experiment 2, the chess experts engaged in the same search task, but under conditions where the stimuli consisted of randomly generated board layouts. To the degree that contextual cueing is affected by meaning, learning effects should be slower developing and of lesser magnitude compared to those observed in Experiment 1.
Method
Participants
The experts that participated in Experiment 1 also participated in Experiment 2. In order to minimize any potential carry-over effects, substantial time elapsed between each experiment (M = 40 days), and participants were not debriefed about the purpose of either experiment until the completion of Experiment 2. Post hoc analyses (reported below) indeed indicate that with these precautions the effect of transfer across experiments was small compared with the effect of learning within each experiment.
Stimuli and apparatus
Stimuli consisted of 248 chess boards. Each board was paired with a board used in Experiment 1 such that (a) the target was located in the same position, (b) the target square was occupied by the same playing piece, and (c) the boards contained the same number of pieces (see Figure 1). The remaining playing pieces included on each board were randomly selected (without replacement) from the set of 31 possible remaining pieces that could appear on the chess board at the start of the game, and the positions of these randomly selected playing pieces on the board were randomly determined. All other aspects of the stimuli were the same as those in Experiment 1.
Design and procedure
The design and procedure were the same as those in Experiment 1.
Results and Discussion
As in Experiment 1, trials were excluded from analysis if a response was not made (4%) or was incorrect (1%), and novel trials were further trimmed to exclude search times greater than 2.5 standard deviations from the mean calculated on a subject-by-subject basis (2%). To increase statistical power, blocks were again collapsed into 6 epochs. Analyses contrasted search performance in Experiment 2 (random boards) to that in Experiment 1 (meaningful boards).
Table 1 summarizes search times for novel and repeated trials as a function of meaningfulness and epoch. Preliminary analyses conducted on the novel trials indicated that search times for random boards were faster than those for meaningful boards, F(1, 4) = 13.6, p < .05, although this effect did not vary over epochs, F(5, 20) < 1. A marginal decrease in search time over epochs was observed, F(5, 20) = 2.33, p = .08, again indicating that the search task probably became easier with practice. Because the experiments differed in baseline search speed, as in Experiment 1, our remaining analyses focused on the cueing effect within each condition.
A 2 (experiment) by 6 (epoch) repeated measures ANOVA compared the cueing effects for the meaningful and random boards (see Figure 2). As expected, a main effect of epoch was observed, F(5, 20) = 7.75, p < .001, as the magnitude of the cueing effect increased over epochs. Importantly, a main effect of board type was also observed, F(1, 4) = 40.6, p = .003, as the magnitude of the cueing effect was, on average, smaller for the random boards. The interaction of these factors was also reliable, F(5, 20) = 5.02, p = .004, as the cueing effect increased at a slower rate over epochs for the random boards. Whereas linear regression analyses indicated that while the cueing effect for the meaningful boards increased by 13% ms per epoch (see Experiment 1), the rate for the random boards was 6% per epoch.
Despite our attempts to minimize possible carry-over effects across experiments, it is nevertheless true that the baseline search rate in Experiment 2 was reduced compared to Experiment 1. Although it is not clear why the baseline search rate decreased across experiments (e.g., practice with the search task or the reduction of distracting semantic information are both possible candidates), one could argue that reduced cueing effects are correlated with reduced baseline search rates. While we cannot rule out the possibility of any degree of correlation, the data do not reasonably support the claim that this correlation is perfect: Although the baseline search rate was reduced by 16% in Experiment 2 relative to Experiment 1, the contextual cueing effect was reduced 51% across the experiments. Thus, whatever the cause of the decrease in the baseline rate of search, it did not have an equal effect on the baseline search rate and the contextual cueing effect.
The results of Experiment 2 provide further evidence that the meaningfulness of a given search context affects the rate at which context–target associations are learned and the magnitude of the learning benefit. While experts were able to learn the association between an arbitrarily located target and an array of randomly selected and positioned playing pieces, the rate of learning and the resulting learning benefit were approximately halved compared to a situation where board layouts reflected actual game-play.
General Discussion
Expertise has been demonstrated to affect performance on a wide range of visually based tasks. Chess experts are better able to recall previously studied board layouts than are novices (Chase & Simon, 1973a, 1973b). Change blindness is alleviated among football experts when changes are made to football-related images (Werner & Thies, 2000). Inattentional blindness is reduced among basketball experts when unexpected objects enter into view while watching basketball-related events (Memmert, 2006). Tennis and cricket experts are better able to anticipate the movement of balls following serves and pitches (e.g., Goulet, Bard, & Fleury, 1989; Land & McLeod, 2000). Hockey experts fixate tactically critical areas more rapidly when making defensive strategy decisions in real time (Martell & Vickers, 2004). Gymnastics experts make fewer and longer fixations when searching for performance errors (Moreno, Reina, Luis, & Sabido, 2002). Thus, expertise allows an individual to more efficiently guide attention to task-relevant visual information and improve performance on visually guided tasks. One general mechanism that may underlie these expertise effects is an enhanced ability to use semantic information over and above strictly visual information to predict the locations of a display's task-relevant content. Thus, building on this prior work, we examined this possibility by contrasting the ability of chess experts and novices to learn and mentally represent associations between a search target and its surrounding context in order to assess the degree to which semantic information is used in these learning and memory processes.
A variety of visual factors including the similarity and spatial proximity of search targets and consistently associated contextual elements affect the ease with which observers can learn once unknown target positions (i.e., contextual cueing). In two experiments, we investigated whether the degree of semantic meaning portrayed by a visual display also affects the rate at which context–target associations can be learned. In order to examine semantic effects on contextual cueing while controlling visual input, we devised an experimental paradigm where novice and expert chess players searched for target letters embedded in illustrations of chess boards. These letter targets were used because their positions in the display could not be predicted a priori by the search context. In Experiment 1, boards depicted actual game play and search benefits for repeated boards were four times greater for experts than for novices. In Experiment 2, search benefits among experts were halved when less meaningful randomly generated boards served as the learning context. Thus, one's ability to extract meaningful relationships among the component objects in a display contributes to his or her ability to use the context created by those objects to locate a search target independently of the quality and nature of the provided visual input.
Why does semantic information contribute to contextual cueing? While a complete accounting of the mechanisms involved in contextual cueing falls outside the scope of this report, here we briefly discuss three possible explanations consistent with the current results and other related literatures. First, semantically rich displays may give rise to inherent expectations concerning the arrangement of elements. As a result, the association of target positions and semantic information may enable faster orienting of attention to the target than in a situation in which the target can only be associated with visual context. Second, semantic information may help reduce the visual similarity (i.e., confusability) of visual contexts. For example, even subtle visual differences in the arrangement or identities of objects in a display may drastically alter the display's semantic identity. Associating target locations with semantic context may therefore enable faster memory encoding and more reliable memory retrieval. Finally, it is possible that the nature of the memory representations involved in learning visual and semantic context differ. That is, the repetition of visual contexts might be encoded implicitly, whereas semantic contexts might be encoded explicitly, resulting in awareness of the repetition and thus more robust learning when semantic information is available.
Regardless of the root cause, the present study indicates that the use of simple arrays of letters and shapes has led to only a partial characterization of how memory for visual contexts develops and how it can in turn be used to guide behaviour. The effects observed in this report indicate that the manner in which consistently arranged information guides attention to known search targets can be driven by different memory mechanisms involved in the encoding and retention of complex displays. In addition, the results of this study provide at least a partial explanation for the differing strength of contextual cueing effects observed across a wide range of visual displays. Whereas arrays of letters and shapes tend to produce search benefits of the order of a couple of hundred milliseconds, real-world scenes generate search benefits of the order of a couple of thousand milliseconds (Brockmole & Henderson, 2006b). Based on the current findings, this difference seems to be, at least in part, a reflection of the degree of semantic meaning contained in those displays.