
Abstract
The purpose of the present study was to determine whether the action system plays a role in determining which stimulus characteristics capture attention by assessing the interference effects caused by dynamic discontinuities that have similar attention-capturing properties, but different action affordances. Results revealed that offset distractors caused significant interference when onset targets were presented in a keypress task, but did not increase reaction times when the same onset targets were presented in a goal-directed aiming task. In contrast, onset distractors caused interference when offset targets were presented in both keypress and aiming tasks. This pattern of results suggests that the attentional set, and thus the properties of stimuli that capture attention, is modulated by the interaction between stimulus and response expectations.
The majority of studies examining how peripheral events capture attention require participants to make some sort of keypress response (simple or choice) following the detection and/or identification of a target stimulus. These responses typically involve somewhat arbitrary spatial mappings between targets and responses; for example, the appearance of a target in the periphery might require the pressing of the spacebar. In contrast, the bulk of the actions we perform are goal-directed aiming movements that require a direct and accurate spatial coordinate system. The differences in the spatial requirements of keypress and aiming responses have motivated some researchers to question whether the processes of attention may be influenced by the context in which the target action is performed (Allport, 1987). Such considerations have led to the development of action-centred models of attention in which it is suggested that the manner in which attention is distributed is determined, in part, by the action that is to be performed (e.g., Tipper, Lortie, & Baylis, 1992). Two lines of evidence have supported action-centred models. First, several studies have revealed that distractor interference effects in goal-directed aiming tasks are mediated by the relative locations (e.g., Pratt & Abrams, 1994; Tipper et al., 1992) and action affordances (e.g., Bekkering & Neggers, 2002) of the target and distractor. These findings suggest that the goals of the task and the relative efficiency of responses afforded by competing stimuli influence the distribution of attention. Second, it has been demonstrated that the trajectories of aiming movements deviate towards or away from the location of nontarget stimuli depending on the timing and the salience of the stimuli (e.g., Tipper, Howard, & Houghton, 1999; Welsh & Elliott, 2004, 2005). As these trajectory deviations can only occur because of modifications in the representation (coding) of the executed movement, these data suggest that the attentional status of individual stimuli (i.e., attended versus inhibited) affects overall movement organization. Thus, there is considerable evidence that action planning affects attention, and, reciprocally, attention affects action planning.
The present experiment was designed to determine whether the to-be-performed action influences attentional capture by investigating how different types of distractors affect the time to organize keypress and aiming responses. To do so, we chose to examine onset and offset events because recent studies have revealed that these events have similar effects on attention when keypress responses are required. For example, Pratt and McAuliffe (2001) found that onset and offset cues facilitated response latencies to targets at cued locations at short stimulus onset asynchronies (SOAs) and inhibited responses for targets at cued locations at long SOAs when they were presented individually. The finding that both onset and offset cues effectively capture attention (see also Pratt & Arnott, 2008) is consistent with the notion that observers can adopt display-wide attentional sets that are broadly tuned for any localized luminance change (Gibson & Kelsey, 1998; Johnson, Hutchinson, & Neill, 2001). In other words, even though the change in luminance associated with onset and offset events are in the opposite direction, both events captured attention because they are localized luminance transients.
Although onset and offset events may be treated similarly by the attention system (although not identically, see Cole, Kentridge, Gellatly, & Heywood, 2003), these events may have different meanings for the action system when different types of response are required. Differences between onset and offset events might be irrelevant in keypress tasks because participants only need to identify and respond to the location of a luminance change. In contrast, onset and offset events might activate a different series of mechanisms when aiming responses are required because offset events are associated with the removal of an object from the action environment, and onset events are associated with appearance of a new object. As a result, onset events afford specific responses, whereas offset events afford either a nonspecific response or no response at all. If the action system can influence the attention system and mediate the capture of attention, then stimulus events that do not afford specific responses (i.e., offset events during aiming movements) may not capture attention. On the other hand, if the action system cannot influence the attention system in determining what does, and what does not, capture attention, then onset and offset stimuli should capture attention to the same degree in aiming and keypress tasks.
To determine whether the to-be-performed action affects what type of stimuli can capture attention, different groups of participants were required to make either aiming responses or choice keypresses to targets (onset or offset events) presented at one of four possible target locations. Importantly, each onset target appeared either alone or with a simultaneously presented offset distractor, and each offset target appeared either alone or with a simultaneously presented onset distractor. The rationale for using mismatched distractors and targets was that such combinations would provide the most sensitive test of any action-based influence on attentional capture. In addition, the mismatched distractors and targets also reduced the potential that the participants would confuse which event is the target (i.e., respond) and which event is the distractor (i.e., ignore).
Based on previous work regarding onset and offset cues with keypress responses (e.g., Pratt & McAuliffe, 2001), it was predicted that there should be no difference in the magnitude of the interference effects in the onset target/offset distractor and offset target/onset distractor pairings in the keypress task. No differences in the interference effects were predicted because such discrete responses do not require a continuous source of information to ensure accuracy. Thus, the action affordance of onset and offset stimuli should be similar for keypress responses. In contrast, because the action affordance of onset and offset stimuli for aiming responses differ, an asymmetrical pattern of interference was predicted for the aiming task. Specifically, because aiming movements require a continuous source of information to ensure that the response is completed accurately, an onset stimulus affords a specific response and action goal while an offset stimulus does not. Thus, onset stimuli should be much more effective at capturing attention than offset stimuli when an aiming task is required. If the requirements of the to-be-performed action influence attentional capture, then distractor interference effects should be observed in the offset target/onset distractor pairing, but not in the onset target/offset distractor pairing. However, if the to-be-performed action does not influence attentional capture, then the same magnitude of interference effects should be observed in offset target/onset distractor and onset target/offset distractor pairings.
Method
Participants
A total of 24 undergraduate students from the University of Toronto participated in the experiment for course credit. Each participant was tested individually in a single session that lasted 45–60 minutes. A total of 12 participants completed the aiming task, and a different 12 participants completed the keypress task. All had normal or corrected-to-normal vision, provided informed consent, and were naïve about the purpose of the experiment.
Apparatus and experimental stimuli
Participants sat at a table in front of a touchscreen monitor (ELO systems 21′ CRT). The top of the monitor was titled away from the participant such that the surface of the screen was at an angle of approximately 20° to the table top. All experimental stimuli were presented on the monitor. Participants who completed the keypress task made all responses on an English-language keyboard. Participants in this group were instructed to place their index and middle fingers of the left and right hands on the “X”, “Z”, “. > ”, and “/?” keys, respectively. Participants in the aiming task completed all responses on the touch screen. For this reason, a starting (“home”) position (an open green circle, 1.5 cm in diameter) was presented 20 cm below the centre of the line of placeholders (see Figure 1). The home position in the aiming task was the only between-task difference in the experimental stimuli.
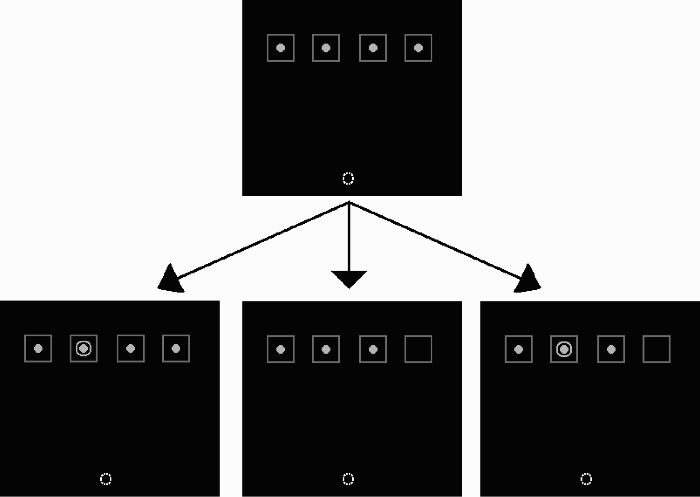
Diagrams of the experimental displays. The upper panel represents the initial display, and the row of lower panels represents the experimental displays. The lower left panel is an example of an onset-alone trial, the lower middle panel is an example of an offset-alone trial, and the lower right panel is an example of a trial in which both onset and offset event occur simultaneously. The dashed circle at the bottom of each panel represents the green circle that was the starting (“home”) location of the aiming movements during the aiming task. This green circle was not present during the keypress task. Note that the distances are not to scale.
The initial display in each task consisted of four blue placeholder squares (3 × 3 cm) surrounding small, solid grey circles (1.5 cm in diameter) near the top of the screen. The placeholders were arranged in a horizontal line 3.5 cm apart. Offset and onset events consisted of the sudden disappearance of a small grey circle and the sudden appearance of a larger, open grey circle (2 cm in diameter) within a placeholder, respectively. The number and luminance of the pixels was equated for onset and offset events to ensure that the on-screen change in luminance of each event was identical, but in opposite directions. Because all stimuli were presented on a black background, an onset event was associated with a luminance increase, and an offset event was associated with a luminance decrease. Offset and onset events were either targets or distractors, depending on the block of trials. In the onset-target condition blocks, the onset event was the target, and the offset event was the distractor. In the offset-target condition blocks, the offset event was the target, and the onset event was the distractor. All combinations of onset and offset events (i.e., offset event alone, onset event alone, and onset event in one location and an offset event in another location) at each potential location were presented equally and randomly across all trials within a block, with the exception that onset and offset events were never presented at the same location within a trial.
Aiming task procedure
Each trial began with the initial display. Participants were instructed to initiate each trial by placing the index finger of their dominant (right) hand in the home circle. When the finger was in the home circle, the circle turned white, indicating that the trial was about to begin. The target and/or distractor event(s) were presented approximately 2 seconds later. Participants were told to move the finger as quickly and accurately as possible from the home position to the location (blue box placeholder) of the target while ignoring any distractor should one occur. They were to make no response on distractor-only trials. Once the participant successfully touched the screen inside one of the placeholders, the initial display was reset. The initial display remained until the participant returned the finger to the starting position, and it turned white again. Trials were self-paced to allow sufficient time for the participant to return the finger to the home position and to ensure that the finger of the participant was in the starting location prior to the onset of the target/distractor array. Trials on which only the distracting event occurred (catch trials) were reset after 1.5 s, and participants had to remove and then replace their finger in the starting position to initiate another trial.
There were 240 trials in each target condition, presented in blocks of 80 trials, for a total of 480 trials. Participants were given a break after each block. One half of the participants completed the onset-target trials before the offset-target condition, and the other half of the participants completed the offset-target condition first. The timing and location of the start and end of the response on each trial were recorded.
Keypress task procedure
The target and distractor events employed in the keypress task were identical to those used in the aiming task. Two seconds after the appearance of the initial display, the target and/or distractor event(s) occurred. Participants were asked to identify the location of the target by pressing the key that was associated with the target location as quickly as possible after the target event occurred. The far left box was related to the “Z” key (left-middle finger), the middle-left box was related to the “X” key (left-index finger), the middle-right box was related to the “. > ” key (right-index finger), and the far right box was related to the “/?” key (right-middle finger). At the beginning of each block, participants were reminded to ignore the distractor and to respond only to the location of the target. Thus, they were to restrain from responding if the distractor appeared alone. Once the response was recorded (or 1,000 ms past on distractor-alone trials), the initial display was reset. The next target/distractor condition was presented approximately 2s after the initial display was reset. Trials in the keypress task were experimenter-paced, unlike in the aiming task, because the fingers of the participant were always over the response position. Thus, no additional time was required to allow participants to return their fingers to the response locations.
Three blocks of 100 trials were completed in each target condition (600 total trials). An optional break was given between each block of 100 trials. Participants completed all 300 trials of one condition before beginning the other condition. One half of the participants completed the onset-target trials before the offset-target condition, and the other half of the participants completed the offset-target condition first. The timing and identification of the response on each trial were recorded.
Data reduction
Reaction time (RT) in the keypress task was defined as the time interval from the onset of the target until the initiation of key depression. RT in the aiming task was defined as the time interval from the onset of the target to the moment the finger lost contact with the screen, and movement time (MT) was defined as the interval from the moment the finger broke contact with the screen to the moment the finger contacted the screen inside one of the target areas.
Data for trials on which an incorrect button was pressed in the keypress task were removed from the data set. Similarly, data from trials in which the participant missed the target location in the aiming task were removed. Data for any trial on which RTs were less than 100 ms or greater than 1,000 ms, or when MTs in the aiming task were greater than 1,000 ms, were also removed. Overall, less than 1–7% of the trials per participant were removed from the aiming data, and 1–3% of trials per participant were removed from the keypress data.
Results
Reaction times
Mean RTs for all target-alone and target-plus-distractor trials were calculated and were submitted to a 2 (task: keypress, aiming) by 2 (target: onset-target, offset-target) by 2 (distractor presence: target alone, target-plus-distractor) mixed analysis of variance (ANOVA) with task as a between-subjects factor and target and distractor presence as within-subjects factors. The results of this analysis revealed that RTs were shorter overall in the aiming (385 ms) than in the keypress (480 ms) task, F(1, 22) = 15.04, p < .001, MSE = 14,390.5, and were shorter for onset (409 ms) than for offset (455 ms) targets, F(1, 22) = 17.97, p < .001, MSE = 2,871.4. There was also an overall distractor interference effect with RTs for target-plus-distractor trials (437 ms) being longer than RTs for target alone trials (427 ms), F(1, 22) = 21.01, p < .001, MSE = 129.69. These main effects were mediated by significant two-way interactions between task and distractor presence, F(1, 22) = 13.38, p < .001, MSE = 129.69, and target and distractor presence, F(1, 22) = 8.27, p < .01, MSE = 158.33. Post hoc analysis of these two interactions using Tukey's HSD test revealed that distractor interference effects were (a) larger overall in offset target/onset distractor pairing than in the onset target/offset distractor pairing, and (b) present in the keypress task, but not present in the aiming task.
While distractor interference effects were not observed in the aiming task, it is possible that the magnitude of the overall differences between the mean RTs in the keypress and aiming tasks masked any condition-specific interference effects in the aiming task. As such, separate 2 (target: onset-target, offset-target) by 2 (distractor presence: target alone, target-plus-distractor) within-subjects ANOVAs were subsequently performed with the mean RTs from both keypress and aiming tasks. Consistent with the hypothesis that attentional capture is mediated by the to-be-performed action, the results of these analyses revealed different patterns of interference in the keypress and aiming tasks. Specifically, the analysis of RTs in the keypress task revealed a main effect for distractor presence, F(1, 11) = 46.51, p < .001, MSE = 97.8, but no interaction between target and distractor presence, F(1, 11) = 0.81, p > .38 (Figure 2b), whereas the analysis of RTs in the aiming task revealed no main effect for distractor presence, F(1, 11) < 1, but a significant interaction between target and distractor presence, F(1, 11) = 9.04, p < .05, MSE = 123.4 (Figure 2a). Subsequent simple-effect ANOVAs indicated that the RTs on target-plus-distractor trails were significantly (p < .05) longer than RTs on target-alone trials in all conditions except for the onset-target/offset distractor pairing in the aiming task, F(1, 11) = 2.68, p < .13, MSE = 136.3.
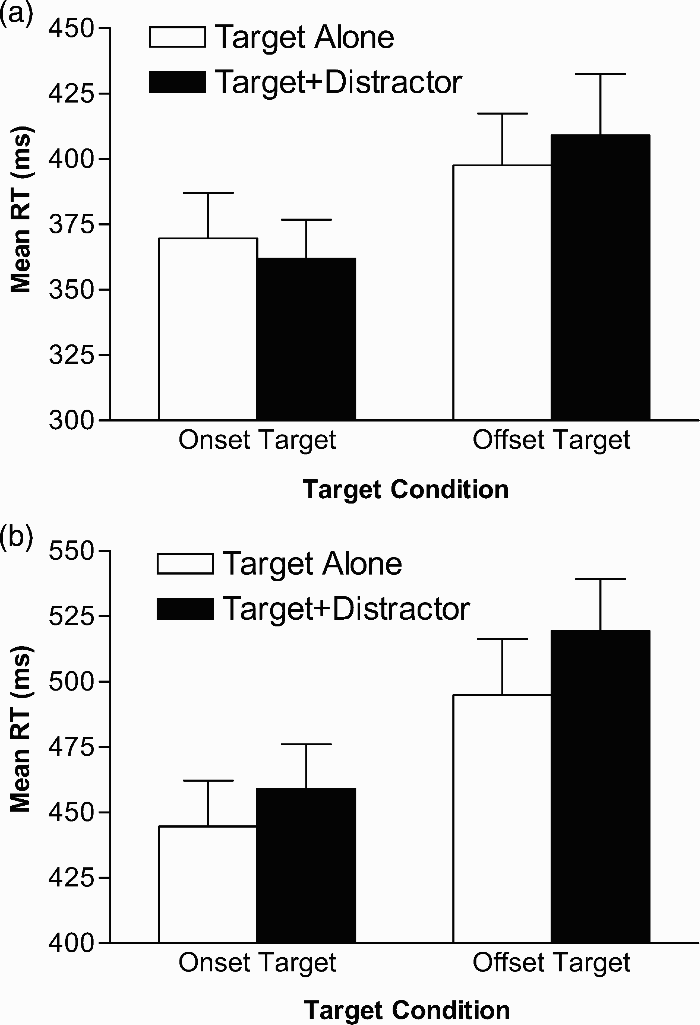
Mean reaction time (RT) in ms as a function of target condition and distractor condition for the aiming task (a) and the keypress task (b). Open bars represent values for trials on which the target was presented alone. Solid bars represent values for trials on which a distractor was presented simultaneous to target presentation. SEM bars are shown.
Overall, the results of the RT analyses support the hypothesis that attentional capture is mediated by the to-be-performed action because onset distractors interfered with the processing of responses to offset targets in both keypress and aiming tasks, but offset distractors only interfered with the processing of responses to onset targets when a keypress response was required (Figure 3). In other words, the results indicate that both onset and offset stimulus events capture attention when an arbitrary, but spatially related, choice keypress was required (see also Folk, Remington, & Johnston, 1992; Folk, Remington, & Wright, 1994; Gibson & Kelsey, 1998). In contrast, a distracting stimulus that did not afford a specific aiming response (i.e., offset distractor) did not capture attention when aiming responses were required.
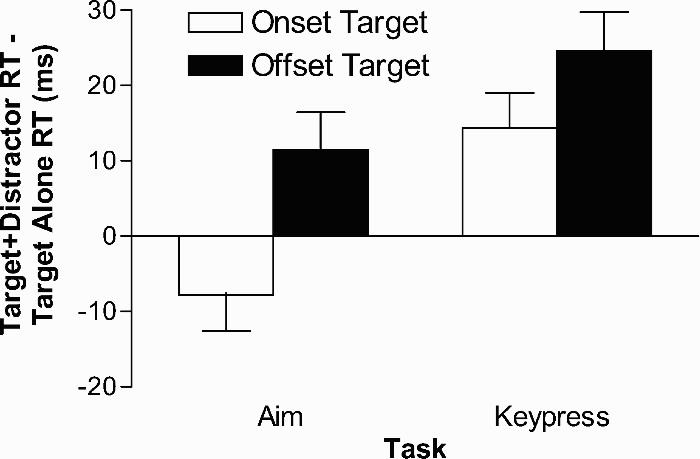
Mean distractor interference effects (target-plus-distractor RTs minus target alone RTs) in ms as a function of task (aim/Experiment 1, keypress/Experiment 2) and target condition. SEMs of the difference scores are shown.
Movement time
While the RT results revealed an absence of interference in the onset target/offset distractor pairing in the aiming task, it is possible that participants were strategically trading speed for accuracy in this condition and may have shifted interference effects in this condition to MT. If this was the case, then a disproportionately large interference effect would be observed in MT in the onset target/offset distractor pairing. To test this possibility, mean MTs in the aiming task were submitted to a 2 (target: onset-target, offset-target) by 2 (distractor presence: target-alone, target-plus-distractor) repeated measures ANOVA. The analysis of MTs only revealed a main effect for distractor presence, which indicated that the distractor slowed response execution overall, F(1, 11) = 19.02, p < .005, MSE = 50.9. The interaction between target and distractor presence was not significant, F(1, 11) < 1 (mean MTs: onset target-alone = 295 ms; onset target/offset distractor = 302 ms; offset target-alone = 292 ms; offset target/onset distractor = 303 ms). Thus, the condition-specific interference effect found in RT was not the result of strategic differences where participants initiated their responses as quickly as possible based on the first environmental change on onset-target trials and then relied on time-consuming feedback-based processes to correct the movement to the actual target location.
Errors
There were no trials on which a participant in the aiming task completed a response to a distractor location. Thus, a between-task comparison on response errors was not possible. However, a 2 (task: keypress, aiming) by 2 (target: onset-target, offset-target) mixed ANOVA was performed on the number of responses initiated on catch trials. This analysis revealed that there were no significant differences in the number of movements initiated on catch trials in the onset- and offset-target conditions, F(1, 11) < 1, and no significant interaction between task and target, F(1, 11) < 1. This error analysis adds further support to the notion that the condition-specific interference effect found in RT was not the result of a trade-off between speed and accuracy.
Discussion
There is clear behavioural (Adam & Pratt, 2004; Tipper et al., 1992; Welsh & Elliott, 2004) and neurophysiological (Rizzolatti, Riggio, & Sheliga, 1994) evidence that attention and action systems are intricately linked. These observations have formed the basis of action-centred models of selective attention through which it is proposed that the processes of attention influence the actions that we perform and, reciprocally, that the action system can influence the distribution of attention. The purpose of the present study was to determine whether the action system influences which events will, or will not, involuntarily capture attention. Participants were asked to respond to an offset or an onset target in the presence of a distracting onset or offset distractor (respectively) by either completing a goal-directed aiming movement to the location of the target or pressing a spatially related key. Although both of these response modes required participants to identify target from nontarget stimuli and choose a target response from a series of nontarget responses (i.e., neither task required a simple detection response), the nature and amount of visual information that was required for the successful completion of keypress and aiming responses was quite different. The main determinants of success for a keypress response are the abilities to detect the correct location of the abrupt change and then correctly link that location to an arbitrary, but spatially compatible, response. Thus, for keypress responses, a continuous source of target information is not required, and, as a result, any dynamic change in the environment (onset or offset events) should afford such a response. In contrast, in order to successfully complete an aiming movement, the action system requires both the detection of the change and a continual source of information so that an accurate spatial representation of the target location is available for precise response programming and feedback-based control (Heath, Westwood, & Binsted, 2004). As such, when aiming responses are required, the attention system should be most sensitive to dynamic changes in which an intact object is present throughout response organization and execution (onset, but not offset events).
Consistent with the hypotheses that dynamic discontinuities have different action affordances in different response modes and that the action system can influence the capture of attention, a nontarget offset event only captured attention and subsequently interfered with the processing of a response to a target onset event when a keypress response was required. Importantly, the offset event did not produce interference when an aiming movement was required. Thus, it appears that both the expected characteristics of the target stimulus (Folk et al., 1992; Gibson & Kelsey, 1998) and the requirements of the action interact to form the basis of the attentional set that influences the distribution of attention and the stimulus properties that capture attention (Welsh & Elliott, 2004). The intricate and reciprocal links between attention and action systems, which are supported by recent neurophysiological evidence of functional upstream connections from primary motor cortex to parietal cortex in humans (e.g., Chouinard, van der Werf, Leonard, & Paus, 2003; Fox et al., 1997), should not be unexpected if one considers that one of the main purposes of the attention system is to provide detailed information on a subset of environmental stimuli for the performance of goal-directed actions (Allport, 1987). Thus, our potential for action should be driven by attentional capture, and the manner in which we attend to objects in the environment should be driven by the actions we are intending to perform.
While the present results add to the growing evidence that the to-be-performed action influences attentional capture, it is possible that the absence of an interference effect in onset target/offset distractor condition in the aiming task was caused by relative differences in the efficiency of response programming to onset and onset stimuli. Because RTs to onset stimuli were shorter than RTs to offset stimuli in the aiming task, the response to the onset target may have been organized and released before the nontarget response to the offset distractor reached a level of activation of sufficient amplitude to interfere with response programming to the onset target. If the S–R compatibility or response processing efficiency account of the interference effects is correct, however, then one would also expect an absence of interference in the onset target/offset distractor condition in the keypress task because RTs to onset targets were also shorter than RTs to offset targets (note that the task by target interaction in RT was not significant; F < 1). In other words, if RTs to onset and offset stimuli provide an indication of response programming efficiency and subsequent interference, then one would also expect an absence of interference in the onset target/offset distractor condition in the keypress task. As the magnitude of the interference effects in the onset target and offset target conditions in the keypress task were significant and not different from each other, it is unlikely that the relative efficiency of response programming was the underlying mechanism.
Overall, the results of the present study have two important and related implications. First, it seems that a reconsideration of the factors that influence the attentional set (Folk et al., 1992; Gibson & Kelsey, 1998) is warranted because there is converging evidence that the goals of the action system can also influence the direction of visual search by modulating the priority given to stimuli that possess varying action-based properties (see also Bekkering & Neggers, 2002). Second, these results highlight the need to carefully consider response mode when designing studies of visual selective attention because the type of response appears to be a factor that determines which stimuli capture attention and, as a result, what information will be drawn from the visual field.