
Abstract
An interlimb practice paradigm was designed to determine the role that visual–spatial (Cartesian) and motor (joint angles, activation patterns) coordinates play in the coding and learning of complex movement sequences. Participants practised a 16-element movement sequence by moving a lever to sequentially presented targets with one limb on Day 1 and the contralateral limb on Day 2. Practice involved the same sequence with either the same visual–spatial or motor coordinates on the two days. A unilateral practice condition (control) was also tested where both coordinate systems were changed but the same limb was used. Retention tests were conducted on Day 3. Regardless of the order in which the limbs were used during practice, results indicated that keeping the visual–spatial coordinates the same during acquisition resulted in superior retention. This provides strong evidence that the visual–spatial code plays a dominant role in complex movement sequences, and this code is represented in an effector-independent manner.
Sequential behaviour is an important characteristic of human cognitive and motor activities in general and a fundamental part of our learned motor repertoire, ranging from everyday motor skills to complex sports skills. In the last three decades there has been an ever increasing interest in the study of sequential movements in an attempt to better understand the response characteristics observed in the production, learning, and transfer as well as the underlying representation and control processes involved in the production of simple and more complex movement sequences (e.g., Hikosaka et al., 1999; Kovacs, Muehlbauer, & Shea, in press; Povel & Collard, 1982; Rosenbaum, 1990; Shea & Wulf, 2005; Verwey & Clegg, 2005). This research demonstrates that a movement structure, defined by the way in which the elements are organized into subsequences, is initially stored in an abstract manner such that the entire sequence can be rescaled in time, force, or amplitude while maintaining effective control as long as the scaling changes are proportional across the entire sequence (e.g., Braden, Panzer, & Shea, 2008; Dean, Kovacs, & Shea, 2008; Muehlbauer, Panzer, & Shea, 2007; Wilde & Shea, 2006). Results have also demonstrated that participants are capable of effectively utilizing the movement structure developed using one set of effectors to produce the movement sequence using an unpractised set of effectors (e.g., Park & Shea, 2005) early in practice but that effector transfer becomes limited after extended practice (Park & Shea, 2003, 2005).
These findings are consistent with the theoretical proposal by Hikosaka et al. (1999), which argues that the learning of movement sequences involves both an effector-independent component and an effector-dependent component. A fundamental premise of the Hikosaka proposal is that sequence learning occurs simultaneously and independently at two levels. One level codes the movement sequence in visual–spatial coordinates (e.g., spatial locations of the end effectors and/or sequential target positions) and the other in motor coordinates (e.g., activation pattern of the agonist/antagonist muscle—joint angles, torques). According to this perspective the visual–spatial code develops relatively quickly (depending on the complexity of the task), is coded abstractly (effector independent), and is the dominant representation early in practice. However, the motor code, which develops more slowly, more effectively represents and integrates the movement dynamics and optimizes the capabilities of the specific effectors (effector dependent) used to produce the sequence and is the dominant representation late in practice. The result of extended practice is more effective and efficient, but more specific, responses.
The Hikosaka model, at least on the surface, also seems to have many similarities to theoretical perspectives that propose intrinsic and extrinsic coordinate or coding systems (e.g., Criscimagna-Hemminger, Donchin, Gazzaniga, & Shadmehr, 2003; Krakauer, Ghilardi, & Ghez, 1999). The intrinsic coordinates are thought to be represented in terms of an internal model of joint representations (Criscimagna-Hemminger et al., 2003), musculoskeletal forces and dynamics (Krakauer et al., 1999), and/or orientation of body segments relative to each other (Lange, Godde, & Braun, 2004). This type of coordinate system is thought to be effector dependent to the extent that biomechanical, neurological, and dynamic properties of the effectors used on the transfer test are dissimilar to those used during practice. The extrinsic coordinates are thought to reflect Cartesian coordinates of the task space with respect to the body and visual display. Thus, this type of coordinate system is thought to be effector independent when the extrinsic coordinates are reinstated during an effector transfer test even though intrinsic characteristics of the required transfer movement may have been altered.
Each of these theoretical perspectives argue that two types of processing modules, representations, or coordinate systems depending on the theoretical perspective are developed independently, at different rates during practice, and limit effector transfer to different degrees. In addition, there is evidence that suggests that hemispheric specialization limits the development and/or the utilization of one or more codes depending on which limb is used during practice and subsequently used on a transfer test. The left hemisphere appears to play a dominant role in movement organization, selection (Rushworth, Krams, & Passingham, 2001; Schluter, Rushworth, Passingham, & Mills, 1998) and learning (Grafton, Hazeltine, & Ivry, 2002). The right hemisphere has been implicated in the development of spatial memory, learning, response selection (Schumacher, Elston, & D'Esposito, 2003), and specification of final position of movement (Sainburg & Wang, 2002). Additionally, left-hemisphere processing appears to be guided by motor representations, whereas right-hemisphere processing is driven by the visual–spatial environment (Goldberg, Podell, & Lovell, 1994). Thus, the transfer of the various representations may be asymmetric across limbs, and this asymmetry may change over practice as reliance on visual–spatial codes, for example, is reduced over practice, and greater reliance is placed on motor codes.
A number of experiments investigating effector transfer of movement sequences have used one set of effectors (e.g., right limb) for training and have tested transfer with an unpractised set of effectors (e.g., left limb) where the transfer test involved the same pattern of flexion and extension (e.g., mirror movements) as the previously practised sequence (e.g., Deroost, Zeeuws, & Soetens, 2006; Park & Shea, 2003, 2005). Other experiments have investigated effector transfer comparing performance in both mirror and visual–spatial (Cartesian) transfer conditions (e.g., Criscimagna-Hemminger et al., 2003, Kovacs et al., in press; Wang & Sainburg, 2003, 2005) Importantly, to our knowledge very few experiments, including those specifically testing the Hikosaka model (e.g., Bapi, Doya, & Harner, 2000), have investigated the influence of these codes on learning following interlimb (practising a task with left then right limbs or vice versa) practice (see Galea & Miall, 2006, for an exception). Some recent experiments have looked at bimanual practice (practice using both limbs at the same time) with transfer to single-limb performance (e.g., Berner & Hoffman, 2008; Burgess, Bareither, & Patton, 2007). This is quite different from practising a task with one limb and then another because bimanual tasks elicit coupling of the limbs while this would not be expected in intermanual practice. The primary question we address in the present experiment involves the determination of the role that visual–spatial and motor codes play in the learning of a dynamic movement sequence and the degree to which these codes can be utilized when training includes dominant and nondominant limb practice. More specifically, we propose, based on the Hikosaka model and our previous research, that interlimb practice of a complex movement sequence will result in enhanced learning when the same visual–spatial coordinates are available during practice with both limbs, even though the activation pattern of the muscles and pattern of joint angles are changed compared to when the same motor coordinates are available during interlimb practice or different motor and visual–spatial coordinates are available during unilateral practice.
These questions are important from both theoretical and practical standpoints because the Hikosaka model provides potential mechanisms to explain both the abstract and more specific codes that seem to develop over practice and the asymmetries that have been noted in some experiments using interlimb transfer paradigms. Further, the Hikosaka model seems like a good model to invest in because it is grounded not only in behaviour data but also by imaging data. In terms of the neural substrate, the Hikosaka et al. (1999) model proposed that intracortical serial connections between the association cortex, motor cortex, basal ganglia, and cerebellum develop. Visual–spatial processing is supported by circuits formed between the dorso-lateral prefrontal cortex (DLPFC; also see Robertson, Tormos, Maeda, & Pascual-Leone, 2001), anterior basal ganglia (head of the caudate), and posterior lobe of cerebellum, while motor processing is supported by circuits between the motor cortex, midposterior basal ganglia (putamen), and anterior lobe of cerebellum and dentate nucleus. The premotor area (PMA) and pre-supplementary motor areas (pre-SMA) provide the link between cortical areas. The imaging work suggests that the association cortex–anterior basal ganglia circuits are more active early in learning and seem to be available to consciousness and require attention. Explicit knowledge of a visual–spatial sequence can be utilized to guide the motor sequence. This has been termed as translation mechanism by Hikosaka, and the premotor area (PMA) seems to play a major role in this process. On the other hand, the circuits within the motor system appear to develop more slowly and at a more implicit level. Eventually, practice results in a shift from loops specific to visual–spatial coordinates processing to loops associated with motor coordinates processing after substantial practice. Additionally, given their independent nature, the visual–spatial and motor (especially) mechanisms might generate inconsistent/incongruent signals. In this case sequence production might rely on the visual–spatial mechanism and explicit knowledge and at the same time suppressing the motor mechanism during learning. This has been termed coordination by Hikosaka, and it is mediated by the presupplemental motor area (pre-SMA).
From a more practical standpoint this research has the potential to impact the design of practice and relearning therapy programmes designed to enhance performance of normal and neurologically impaired (e.g., stroke, Parkinson's patients) individuals. With a greater understanding of the mechanisms by which performers are able to transfer movements sequences and the limitations of transfer, new learning and relearning programmes can be constructed to optimize performance by concentrating on specific, rather than general, problems associated with learning and transfer. Further, by observing patterns of transfer, additional clues may emerge as to how these sequences are stored and represented in memory, compiled prior to execution, executed, and ultimately learned and transferred. In addition, the unique interlimb practice component of the proposed experiments has the potential to provide new and important clues as to the basis of the transfer asymmetries often noted in interlimb transfer experiments. Understanding the conditions that best facilitate transfer is extremely important in terms of the development and/or refinements of theoretical accounts of sequence production that consider not only the control and learning of a sequence, but also the mechanisms responsible for transfer and relearning (see Shea & Wulf, 2005).
The primary objective was to determine the extent to which the visual–spatial and the motor codes develop when practice is provided first with one limb and then with the other limb (interlimb practice). The results from previous research suggest that early in the learning process the visual–spatial code is stored in an abstract form that permits effector-independent performance. Therefore, we assumed that early in practice sequence learning would benefit from interlimb practice only when the visual–spatial coordinates are kept constant during practice with the two limbs. According to this viewpoint changing the visual–spatial coordinates while keeping the motor coordinates the same during transition from practice with one limb to practice with the contralateral limb should result in inferior performance. To test this hypothesis an interlimb practice paradigm was used. This paradigm allows the determination of which type of coding leads to the most effective retention and the conditions under which interlimb practice leads to the most effective learning. In this design, as in many learning paradigms, performance on the retention test where all participants are tested under the same conditions after a 24-hr delay is the best measure of learning. Performances during practice, where participants are tested under different conditions, may be influenced by temporary effects associated with the specific practice conditions.
Method
Participants
College students (N = 48; age 18 to 35 years) volunteered to participated in the experiment. The participants had no prior experience with the experimental task and were not aware of the specific purpose of the study. All participants were right-hand dominant as determined by the Edinburgh Handedness Inventory (Oldfield, 1971) prior to the experiment. Local ethics committee approved the protocol. Informed consent was obtained prior to participation in the experiment.
Apparatus
The apparatus consisted of a horizontal lever and computer/monitor (see Figure 1). The axle of the lever, which rotated freely in ball-bearing supports, allowed the lever to move in the horizontal plane over the table surface. Near the distal end of the lever, a vertical handle was attached. The position of the handle could be adjusted so that when the participant rested their forearm on the lever, their elbow aligned over the axis of rotation, they could comfortably grasp the handle (palm vertical). The horizontal movement of the lever was monitored (700 Hz) by a potentiometer. A pointer attached to the end of the lever extended so that it could be positioned within the projected targets on the monitor. The potentiometer data were used to provide lever position information to the participant; the data were stored for later analysis. The targets and total movement time were projected on the monitor.
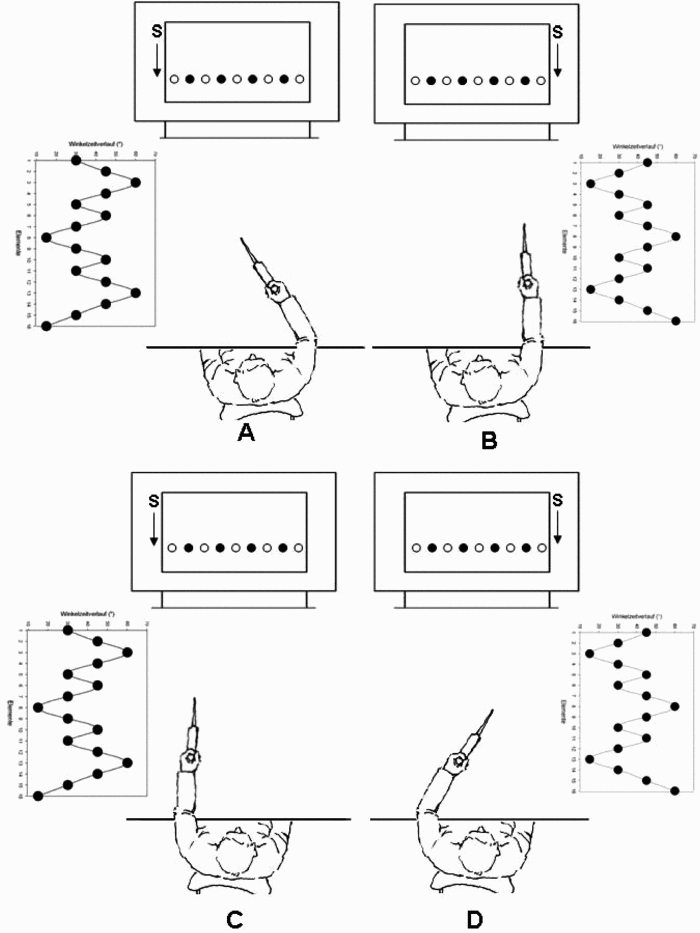
Illustration of the lever and acquisition/test conditions (A to D) and examples of the movement pattern (A to D). The start position (S) and targets (filled circles) used during sequence production are also shown. The unfilled targets were also projected onto the table top but were not illuminated during acquisition or testing. Note that during acquisition and retention only one circle was illuminated (filled) at a time.
Procedure
Upon entering the testing room, participants were seated on a height-adjustable chair facing the monitor. The chair was adjusted so that participant's lower arm was at approximately an 80-degree angle to the upper arm at the starting position. Instructions were given, informing participants of how to perform the task. To begin each block participants were told to move the lever to the starting position (designated as “S” in Figure 1). When the start position was achieved the outline of nine circles (targets) were projected on the monitor. The diameter of the targets represented approximately 1.5 degrees of elbow extension (or flexion) with the centres of the four targets used in the sequence representing lever positions of 13.34, 26.68, 40.02, and 53.36 degrees (13.34 increments) from the start position. For convenience, the targets used in the sequence were labelled 1, 2, 3, and 4 from the start position. The display of the outlines of the targets indicated that the block was about to begin. After a random foreperiod (2–5 s in 0.5-s intervals) a “start” tone was presented, and the first target was illuminated (outline was filled in). Participants were instructed to move the lever from one illuminated target to the next illuminated target as quickly and as smoothly as possible. Upon crossing the edge of the target the illumination was “turned off”, and the next target was immediately illuminated until the sequence was completed. If the participant missed a target, the target remained illuminated until the participant returned the lever (pointer) to the target position. A 16-element sequence (targets: 2, 3, 4, 3, 2, 3, 2, 1, 2, 3, 2, 3, 4, 3, 2, and 1) was repeated 10 times in each of 16 acquisition blocks on each of two days (32 blocks total). After hitting the last target, a stop tone was presented, and the display of the targets was removed. A rest interval of 30 s was provided after each block. Participants were not informed that the targets were presented in a repeated order.
Prior to entering the testing room participants were randomly assigned to one of six practice conditions defined by the ways in which the two practice days were organized and the limb that they used on Day 1 (left start or right start). Figure 1 illustrates the four practice conditions used to construct practice conditions. In one practice condition participants were required to sequentially move to target positions that required the same pattern of flexion and extension on each day but with different limbs (Figures 1A and 1D or 1D and 1A). This condition was termed “mirror” because the pattern of lever movements and the goal pattern displayed for the left and right limb movements were mirror images. In Hikosaka et al. (1999) terms this was considered a motor-congruent condition because the motor coordinates (pattern of muscle activation of the homologous muscles) were the same across the two days of practice. In the second condition participants were required to move to the same sequentially presented target positions on each day of practice but with different limbs. In this condition different patterns of flexion and extension were required but the same visual–spatial pattern of movement required across the two days was the same (Figures 1A and 1C or 1D and 1B). This condition was termed the “nonmirror” condition because the visual–spatial target positions across practice with the two limbs were the same. In Hikosaka et al. (1999) terms this was considered a visual–spatial congruent condition because the same visual–spatial coordinates were required. In the third condition, termed neither, the motor and visual–spatial coordinates were changed across practice days, but the same limb was used (Figures 1A and 1B or 1D and 1C). The neither condition was used as a reference from which to compare the relative effectiveness of the mirror and nonmirror practice conditions. The various practice conditions are depicted in Figure 1. Note that conditions were designed so that the limb used during the first practice session was counterbalanced. Practice days were separated by 24 hours. Note also that the same sequence of targets was used on each practice day and on all tests—only the limb and/or the designated start position was changed.
On Day 3 two retention tests (see Table 1), which required participants to produce the sequence under the same conditions as those that they experienced during practice on Day 1 (Retention 1) and Day 2 (Retention 2), were conducted. The order of the test was counterbalanced. The primary movement-dependent measure was element duration, which was computed as the elapsed time from “hitting” (crossing the target boundary) the currently illuminated target to “hitting” the next illuminated target. Retention Test 1, which involved the same set-up as participants experienced on Day 1 practice, was considered the primary measure of learning because this test was identical for all participants that started with the right hand/arm (Figure 1, Condition A) and for all participants that started with the left hand/arm (Figure 1, Condition D). Retention Test 2 measured performance in the same conditions as those that were experienced on Day 2, but this condition differed across groups.
Design of experiment
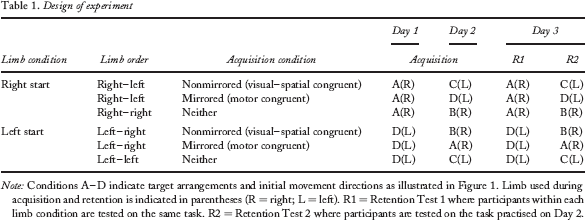
Note: Conditions A–D indicate target arrangements and initial movement directions as illustrated in Figure 1. Limb used during acquisition and retention is indicated in parentheses (R = right; L = left). R1 = Retention Test 1 where participants within each limb condition are tested on the same task. R2 = Retention Test 2 where participants are tested on the task practised on Day 2.
Results
Acquisition
Mean element duration for the three acquisition conditions over the three days is provided in Figure 2. For the two acquisition days separate 3 (acquisition condition: mirror, nonmirror, neither) × 2 (limb condition: left start or right start) × 16 (block: 1–16) analyses of variance (ANOVAs) with repeated measures on block were calculated. The acquisition analysis for Day 1 indicated main effects of limb condition, F(1, 42) = 14.64, p < .01, η2 = .26, and block, F(15, 630) = 139.37, p < .01, η2 = .77. Participants using the left limb on Day 1 performed more poorly than participants using their right limb. Duncan's new multiple range test on block indicated that element duration decreased throughout acquisition. The analysis failed to indicate a main effect of acquisition condition, F(2, 42) = 2.08, p > .05, η2 = .09. However, the Limb Order × Block interaction, F(15, 630) = 2.96, p < .01, η2 = .06, was significant. Simple main effects analysis failed to indicate performance differences between participants that started with their right and left limbs on Blocks 1–4 but participants using their right limb performed the task more quickly on Blocks 5–16 than did participants using their left limb. Because it was important to verify that there were no acquisition condition differences at the end of Day 1 practice we conducted an additional one-way ANOVA on acquisition condition using only performance on Day 1, Block 16. The analysis again failed to indicate a main effect of acquisition condition, F(2, 42) = 2.33, p > .05, η2 = .09.
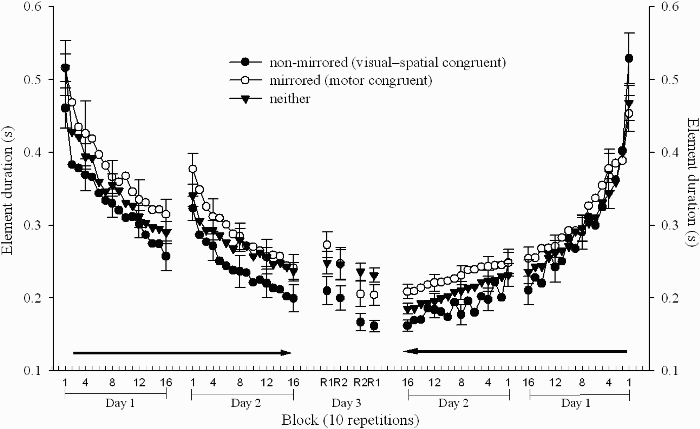
Mean element duration during acquisition (Blocks 1–16) on Day 1 and Day 2 for the groups that began practice with their left limb (left panel) and groups that began practice with their right limb (right panel). Note that the left panel displays the acquisition data for the nondominant to dominant limb order groups from left to right while the right panel displays the data for the dominant to nondominant limb order groups from right to left. Retention results (Day 3) are displayed in the centre panel for all groups.
Analysis of the Day 2 data indicated main effects of acquisition condition, F(2, 42) = 3.48, p < .05, η2 = .14, limb condition, F(1, 42) = 13.25, p < .01, η2 = .25, and block, F(15, 630) = 74.61, η2 = .64, p < .01. Participants in the nonmirror acquisition condition performed significantly better than the participants in the mirror condition, and participants that started with the left limb performed more poorly (even though they were now performing with the right limb) than participants that started with their right limb. Duncan's new multiple range test on block indicated that element duration decreased across blocks until Block 7, after which no further decrease was found. In addition, the Limb Condition × Block interaction, F(15, 630) = 2.08, p < .01, η2 = .05, was significant Simple main effects analysis indicated significantly higher element durations for participants in the left start limb condition than for participants in the right start limb condition on all blocks on Day 2. However, the differences were larger at the beginning of practice than at the end of Day 2 practice.
Transition from Day 1 to Day 2
An additional 3 (acquisition condition: mirror, nonmirror, neither) × 2 (limb condition: left start or right start) × 2 (Day 1, Block 16; Day 2, Block 1) ANOVA with repeated measures on block was conducted. This analysis was conducted to determine whether limb order or acquisition condition influenced the transition to the new limb. The analysis indicated main effects of limb condition, F(1, 42) = 23.38, p < .01, η2 = .36, acquisition condition, F(1, 42) = 4.21, p < .05, η2 = .17, and block, F(1, 42) = 79.91, p > .05, η2 = .67. While performance decreased from the end of Day 1 practice to the beginning of Day 2 practice, the increase was smaller for participants in the right start limb condition than for the participants in the left start limb condition. In addition, the transition for the nonmirror groups was less abrupt than that for the mirror groups. All interactions were not significant.
Retention
For Day 3, a 3 (acquisition condition: mirror, nonmirror, neither) × 2 (limb condition: left start or right start) × 2 (test: 1 and 2) ANOVA with repeated measures on test was conducted. The analysis indicated main effects of acquisition condition, F(2, 42) = 7.75, p < .01, η2 = .27, and limb condition, F(1, 42) = 9.60, p < .01, η2 = .29, but not test, F(1, 42) = 1.50, p > .05, η2 = .04. Duncan's multiple range tests on acquisition condition indicated significantly faster retention performance for the nonmirror condition than the motor and neither condition, which did not differ from each other. Participants that initially practised (Day 1) with the right limb produced the sequence more quickly (shorter element durations) than participants that first practised with the left limb.
In addition, to verify that the acquisition condition differences observed on the retention tests were not due to differences at the end of Day 1 practice, an additional 3 (acquisition condition: mirror, nonmirror, neither) × 2 (limb condition: left start or right start) × 2 (test: 1 and 2) analysis of covariance (ANCOVA) with repeated measures on test retention analysis was conducted using performance on Day 1, Block 16 as a covariate. The analysis again indicated a main effect of acquisition condition, F(2, 42) = 6.72, p < .01, η2 = .25, with the least squares post hoc procedure indicating differences between the mirror and nonmirror acquisition conditions. However, as might be expected using performance at the end of Day 1 as a covariate, the main effect of limb, which was significant in the analysis without the covariate, was not significant. All other main and interactive effects were also not significant.
Discussion
The primary purpose of the present experiment was to determine the role of visual–spatial and motor coordinates in the production and learning of a complex (16-element dynamic arm movement task) movement sequence and to determine the degree to which the codes based on these coordinates can be trained across limbs. We hypothesized that practice with two different coordinate systems, even if the practice involved the same limb (neither acquisition condition) or practice involved the same pattern of effector activation and joint angles (mirror acquisition condition), would not be as effective as interlimb practice when the same visual–spatial coordinates were used throughout practice (nonmirror acquisition condition). At first glance this appears counterintuitive because different patterns of muscle activation and joint angles were required across the two days of practice in the nonmirror acquisition condition, but this hypothesis is consistent with recent effector transfer data indicating enhanced transfer when visual–spatial codes were reinstated on the effector transfer tests (Kovacs et al., in press) after 1, 4, and 12 days of practice. A secondary purpose was to determine whether performance during acquisition and retention was similar regardless of the order in which the limbs were practised. This is important because research has demonstrated that some aspects of performance transfer better from dominant to nondominant limb (e.g., Criscimagna-Hemminger et al., 2003; Lange et al., 2004) while others appear to transfer better from nondominant to dominant limb (e.g., Lange et al., 2004).
We viewed Retention Test 1 as the primary test of learning because the test conditions were identical for each group in the respective limb condition (Condition A for the right to left groups and Condition D for the left to right groups). This was also the condition that each of the groups practised on Day 1(see Table 1). Retention Test 2 involved conditions that each group experienced on Day 2 but the testing conditions were different across groups. The results of the retention analysis, however, indicated enhanced retention on both retention tests for the groups whose two days of practice schedule involved the same visual–spatial coordinates relative to groups where the same motor coordinates were maintained over the two days or practice and the group that practised with the same limb but both the visual–spatial and motor coordinates were changed across practice.
Interlimb practice: Sequence coding
The primary purpose of the present experiment was to determine the role that visual–spatial and motor coordinates play in the coding and learning of complex movement sequences. The present data, which involved dominant arm practice followed by nondominant arm practice and nondominant arm practice followed by dominant arm practice, demonstrated retention advantages for groups provided interlimb practice with the same visual–spatial coordinates (nonmirror condition) compared to groups where the motor coordinates were the same (mirror condition) or the same limb was used but the coordinate system was changed (neither condition). These results are consistent with our initial hypotheses, the Hikosaka model, which proposed that visual–spatial codes play a dominant effector-independent role early in practice, and our recent coding and effector transfer findings following unilateral practice where performance on effector transfer tests were performed better when the visual–spatial coordinates were reinstated than when the motor (mirror) coordinates were reinstated (Kovacs et al., in press). Additionally, the present results extend the findings of van Mier and Petersen (2006), Grafton et al. (2002), and Willingham, Wells, Farrell, and Stemwedel (2000) who have demonstrated for a wide variety of tasks (e.g., pattern drawing, serial reaction time) enhanced effector transfer performance when visual–spatial cues are reinstated following unilateral practice.
We believe, however, that this is the first time that practice with one limb and then with the contralateral limb (interlimb practice) has been shown to enhance retention and transfer performance when the same visual–spatial coordinates (nonmirror) are reinstated relative to when they are changed (mirror and neither conditions). This is especially noteworthy because different patterns of muscle activation and joint angles were required across the two days of practice. Indeed, maintaining the same muscle activation patterns and joint angles while using different limbs across the two days of practice resulted in inferior retention of both practised tasks. Further, we suggest that inconsistencies in the way coordinates are changed from practice to effector transfer tests in past experiments may have added to the inconsistencies found in the effector transfer data following unilateral practice reported in the literature.
Interestingly, the Hikosaka et al. (1999) model, which proposed that relatively early in practice movement sequences are coded in visual–spatial coordinates (also termed extrinsic/Cartesian codes/coordinates), and empirical findings testing this model (e.g., Bapi et al., 2000; Bapi, Miyapuram, Graydon, & Doya, 2006), which suggest that visual–spatial codes are represented in a relatively abstract, effector-independent manner, are similar in some respects to a number of other theoretical perspectives. Each of these theoretical perspectives argues that two types of processing modules, representations, or coordinate systems are developed independently, at different rates during practice, and limit effector transfer to different degrees. The Hikosaka model is similar in many ways to theoretical perspectives that propose intrinsic and extrinsic coordinate or coding systems (e.g., Criscimagna-Hemminger et al., 2003; Krakauer et al., 1999). The intrinsic coordinates are thought to be represented in terms of an internal model of joint representations (Criscimagna-Hemminger et al., 2003), musculoskeletal forces and dynamics (Krakauer et al., 1999), and/or orientation of body segments relative to each other (Lange et al., 2004). The intrinsic coordinate system is thought to be effector dependent to the extent that biomechanical, neurological, and dynamic properties of the effectors used on the transfer test are dissimilar to those used during practice. The extrinsic coordinates are thought to reflect Cartesian coordinates of the task space with respect to the body and visual display. In each perspective the extrinsic coordinate system is thought to be effector independent when the extrinsic coordinates are reinstated during an effector transfer test even though intrinsic characteristics of the required transfer movement may have been altered.
Thus, we would predict that this pattern of retention would not be found when simpler movement sequences were used. We make this prediction based on the notion that motor codes are developed with less practice when the movement pattern is less complicated (see Kovacs, Han, & Shea, 2008). Indeed, motor codes have been shown to be the dominant code for a relatively simple task (two elements) even after a single practice session. However, when complicated movement sequences like those used in the present experiment have been tested, visual–spatial codes have been shown to remain dominant even after 12 days of practice (Kovacs et al., in press).
Interlimb practice: Transfer symmetry
A secondary purpose of the present experiments was to determine whether the performance and learning advantage observed when the same visual–spatial coordinates were used in interlimb practice was influenced by the order in which the arms were used during practice. Studies from Sainburg and Wang (2002) as well as Wang and Sainburg (2006) indicate asymmetry in interlimb transfer depending on whether the dominant or the nondominant arm is used during practice. This has been referred to as the “dominant limb advantage” because intrinsic (motor) codes are thought to transfer better from the dominant to nondominant limbs than vice versa. On the other hand, the Hikosaka model, as well as theoretical perspectives by Keele, Jennings, Jones, Caulton, and Cohen (1995), Verwey (1994), and Grafton et al. (2002) suggest that the spatial (extrinsic) codes are stored in relatively abstract effector-independent ways leading to transfer symmetry. Thus, we predicted transfer symmetry when movement sequences are practised with both limbs. We did find, irrespective of the order of the arms used for practice, pattern symmetry in the transfer results. That is, our results showed learning advantages for the nonmirror condition where visual–spatial coordinates were congruent across practice for both the dominant limb first acquisition conditions and nondominant limb first acquisition conditions.
However, it should be noted that the main effect of limb order was also significant. Participants that started with the dominant (right) limb performed better toward the end of Day 2 practice, even though performance on Day 2 was with the nondominant limb, than did the groups that received nondominant limb practice on Day 1 and performed with their dominant limb on Day 2. In addition, the transition from one limb to the other during acquisition (Figure 2, right) was less abrupt for participants going from the right to left limb (Figure 2, right) than vice versa (Figure 2, left). Most importantly, participants whose initial practice was with the dominant limb performed significantly better on the retention tests than did participants that started with the nondominant limb regardless of the coding scheme. These findings suggest a general dominant limb advantage not specifically related to the various coding schemes. This is different from the dominant limb advantage noted in numerous experiments (e.g., Criscimagna-Hemminger et al., 2003; Kovacs et al., in press) where motor codes appear to be asymmetrically transferred. Sainburg and Kalakanis (2000), for example, argue that there are different neural control mechanisms for the dominant and nondominant limb movements with asymmetric hemispheric access to these mechanisms. This suggests that patterns of transfer asymmetries noted by Sainburg and colleagues may only arise when motor codes are the dominant form of coding and that these asymmetries may not surface for complicated multielement movement sequences until after very extensive practice when the motor code begins to play a more dominant role in movement production.
Summary
Learning of a complex movement sequence was enhanced when the visual–spatial coordinates for the target sequence were unchanged during interlimb practice. This finding is consistent with both the notion that the visual–spatial code is represented in an effector-independent manner and the notion that visual–spatial coding plays a dominant role early in learning. The findings are also consistent with recent research that shows enhanced effector transfer performance on visual–spatial transfer tests following unilateral practice. The present experiment, for the first time, demonstrates that the way interlimb practice is constructed influences learning of the movement sequence. These findings are important not only for theoretical reasons but for practical reasons related to training protocols designed to increment learning when one limb is constrained because of injury, disease, and/or special circumstance where either limb may be required to perform the sequence of movements.