
Abstract
The Simon effect has most often been investigated with key-press responses and eye fixation. In the present study, we asked how the type of eye movement and the type of manual response affect response selection in a Simon task. We investigated three eye movement instructions (spontaneous, saccade, and fixation) while participants performed goal-directed (i.e., reaching) or symbolic (i.e., finger-lift) responses. Initially, no oculomotor constraints were imposed, and a Simon effect was present for both response types. Next, eye movements were constrained. Participants had to either make a saccade toward the stimulus or maintain gaze fixed in the screen centre. While a congruency effect was always observed in reaching responses, it disappeared in finger-lift responses. We suggest that the redirection of saccades from the stimulus to the correct response location in noncorresponding trials contributes to the Simon effect. Because of eye–hand coupling, this occurred in a mandatory manner with reaching responses but not with finger-lift responses. Thus, the Simon effect with key-presses disappears when participants do what they typically do—look at the stimulus.
The influence of stimulus–response (S–R) compatibility on behaviour has most often been investigated with symbolic responses, such as key-presses, and observers were mostly asked to maintain fixation during an experimental trial. Hence, results from a highly constrained laboratory situation served as reference for cognitive models of response selection (e.g., De Jong, Liang, & Lauber, 1994; Kornblum, Hasbroucq, & Osman, 1990). In the present study, we wondered whether these cognitive models are able to accommodate behaviour in more naturalistic settings that involve different types of orienting behaviour and different types of manual responses. Imagine an everyday situation that involves saccadic eye movements and manual reaching movements: When starting your car, you notice that a red light is flashing in the lower left part of your dashboard. Even though you know that this red light warns you to release the parking brake, you will make a saccade to foveate the light to make sure that it is indeed the presumed signal. Then, you will release the parking brake with your right hand, at least when seated in a car in continental Europe. The question is whether the incongruence between the position of the warning light (on the left) and the goal of your hand movement (on the right) will interfere with response selection as predicted by cognitive models. In a typical laboratory task, this complex sequence would be reduced to a right key-press when a left stimulus is presented while the participant looks at the central fixation mark. However, saccades play a critical role in the example and in many situations of our daily life. In addition, most human actions are goal-directed and aim at objects in the environment. In general, eye movements toward the target of the action accompany or precede hand movements, notably because target fixation enhances reaching accuracy (Bekkering, Adam, Van den Aarssen, Kingma, & Whiting, 1995; Prablanc, Echallier, Komilis, & Jeannerod, 1979). These aspects of human performance are rarely considered in laboratory tasks.
The aim of the present study was to study effects of different hand movements (symbolic and reaching responses) under different eye movement instructions (spontaneous eye movements, saccades, and fixation). We chose a type of S–R compatibility effect in which the location of the stimulus is irrelevant for the task, the Simon effect (cf. Simon & Rudell, 1967; for an overview see Lu & Proctor, 1995; Simon, 1990). For instance, if participants are instructed to press a left key in response to a green square and a right key in response to a red square, left responses will be shorter when the green square appears on the left (corresponding trial) than when it appears on the right side (noncorresponding trial). Cognitive models state that the Simon effect occurs at the stage of response selection and that it results from a conflict between automatic and intentional response selection processes (cf. dual-route models, e.g., De Jong et al., 1994; Kornblum et al., 1990). While the irrelevant stimulus location automatically activates the spatially corresponding response, the intentional route translates the relevant stimulus dimension into the correct response according to the instructions. A conflict arises when automatic and intentional response codes differ. In this case, extra time is needed to solve the conflict, and consequently reaction times (RTs) are longer. The automatic activation of corresponding responses was confirmed in studies recording lateralized readiness potentials (e.g., Wascher & Wauschkuhn, 1996), which showed a rise in the activation over the motor cortex contralateral to the stimulus location. This stimulus-related activation will then decay over time (Hommel, 1994).
The attention-shift account of the Simon effect
In the framework of dual-route models, it is important to determine how the spatial stimulus code is formed. It was shown that the relative location between (moving) objects, the body midline, the visual context, and the directional information resulting from the planning of saccadic eye movements can serve as reference frame for spatial code formation (e.g., Abrahamse & Van der Lubbe, 2008; Bosbach, Prinz, & Kerzel, 2005; Hommel & Lippa, 1995; Lamberts, Tavernier, & d'Ydewalle, 1992; Umiltà & Nicoletti, 1992). Attentional accounts claim that the formation of the spatial code involves attention shifts (e.g., Proctor, Lu, & Van Zandt, 1992; Simon, 1969; Stoffer, 1991; Umiltà & Nicoletti, 1992; Verfaellie, Bowers, & Heilman, 1990). For instance, Umiltà and colleagues (Rubichi, Nicoletti, Iani, & Umiltà, 1997; Umiltà & Nicoletti, 1992) suggested that the spatial code of the stimulus is created with respect to the direction of the last shift of attention, which in turn is related to the generation of a saccade (Rizzolatti, Riggio, Dascola, & Umiltà, 1987). However, Ivanoff and Peters (2000) argued that a shift of attention in itself was not sufficient to generate the spatial code (see also Ivanoff, Klein, & Lupiañez, 2002); rather the Simon effect arose only when the shift of attention was performed from an intentionally defined reference. Importantly, these models focused on spatial code formation after covert or overt shifts of attention but never considered the time needed to shift attention or to make a saccade.
Purpose of the study
To our knowledge, the effect of saccades on the Simon effect has never been investigated as participants were required to maintain fixation in most studies. In the present article, we do not go into the debate on the formation of spatial codes (see above) or on the relationship between saccades and attention shifts (e.g., Rizzolatti et al., 1987; van der Lubbe, Neggers, Verleger, & Kenemans, 2006). We only assume that saccadic eye movements are coupled to attention shifts because spatial attention allows selecting positions in space that will be relevant for saccadic behaviour. On the assumption that it takes time to plan and execute a saccade, it follows that the number of saccades performed in a task may affect the time needed to complete the task. The simple issue of time consumption is not taken into account by current models of the Simon effect, but may explain the Simon effect in situations allowing for eye movements: Initially, the presentation of the target stimulus attracts the eyes because it represents an abrupt onset and is task relevant (Folk, Remington, & Johnston, 1992; Yantis & Jonides, 1990). In noncorresponding trials, the eyes may subsequently move toward the correct but empty response location. This second saccade may help observers to find the correct response, or may “embody” their decision. In any case, the delay incurred by the second saccade would explain the slower RTs in noncorresponding trials.
Further, we investigate whether the propensity to make a saccade to the correct location in noncorresponding trials changes as a function of response mode. Importantly, symbolic and goal-directed responses vary with respect to the coupling between hand and eye. When a target has to be reached with the hand, a saccade toward the target precedes the start of the hand movement (e.g., Prablanc et al., 1979). Because key-presses are not goal directed, we think that the coupling between ocular and manual systems will be less strong. In a Simon task, this weaker eye–hand coupling may be visible in a smaller rate of saccades from the imperative stimulus location toward the correct response location in noncorresponding trials (i.e., only one saccade toward the imperative stimulus is expected). Therefore, we expect the Simon effect to decrease. Conversely, the Simon effect is expected to increase when the number of saccades toward the correct response location in noncorresponding trials is high. This may occur when the programming of a reaching movement necessitates an eye movement toward the correct (but empty) response location in noncorresponding trials.
Finally, our study provides the first data detailing where and when the eyes move in a Simon task. Because of the importance of saccades in everyday activities, this evaluation is interesting in itself. To get an idea about the spontaneous eye movement behaviour, we started the experimental session by recording eye movements without any particular instruction (unconstrained condition). Next, in separate blocks of trials, participants were required either to maintain central fixation throughout the trial (fixation condition) or to make a saccade toward the imperative stimulus (saccade condition). Following the first saccade, participants were free to move their eyes (see Figure 1). The role of response mode was assessed by requiring symbolic and goal-directed responses. In the finger-lift condition participants lifted their left or right index finger. In the reaching condition, participants reached one of the two response boxes with the index finger of the right hand (see Figure 1). In both conditions, responses were executed on a flat-panel screen that was also used to display the stimuli.
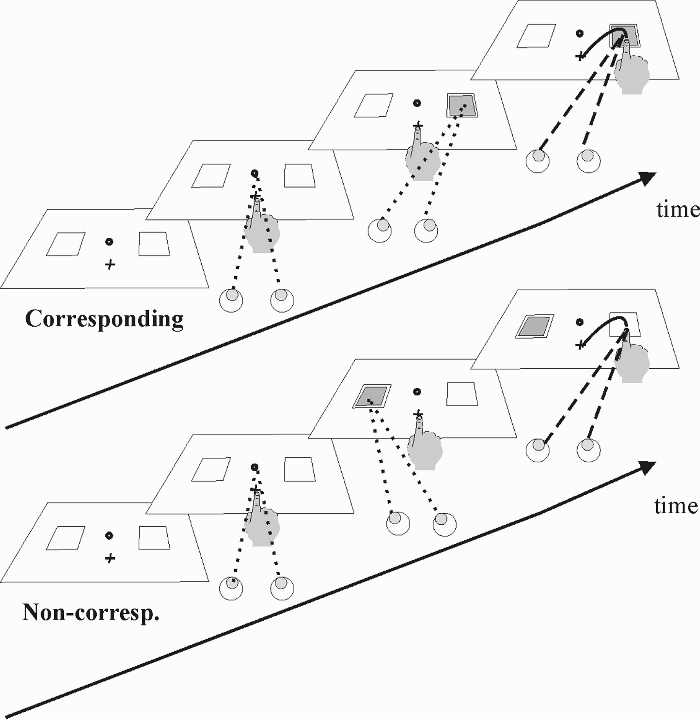
Illustration of a corresponding and noncorresponding trial (upper and lower panels, respectively) in the saccade condition with reaching responses. A trial began when gaze (dotted lines) was on the central circle, and the right index finger was on the cross. At stimulus onset, participants had to perform a saccade toward the coloured stimulus and carry out the reaching response as quickly as possible. The continuous line illustrates the trajectory of the reaching response. After the first saccade, participants were allowed to move the eyes at will. In corresponding trials, we expect the eyes to stay on the stimulus, while in noncorresponding trials, we expect a saccade toward the correct response location (dashed line).
EXPERIMENT 1
Method
Participants
A total of 16 university students (12 females, mean age of 21 years, right-handed) participated in the experiment. All were naïve with respect to the purpose of the experiment and had normal or corrected-to-normal vision. The study was approved by the ethics committee of the Faculté de Psychologie et des Sciences de l'Education of the University of Geneva.
Apparatus and stimuli
An Eyelink II (Osgood, Ontario, Canada) eye tracker was used to record eye movements. The eye tracker was set to pupil-only mode at 500-Hz sampling rate. Before each block, a pseudorandom 9-point calibration procedure was performed. The X, Y, and Z coordinates of manual movements were recorded by an ultrasonic system (CMS20S, zebris Medical GmbH, Isny im Allgäu, Germany) at a sample frequency of 150 Hz by means of a marker positioned on the nail(s) of the finger(s) used to perform the task.
Hand movements were executed on a flat panel screen. The screen was placed in a frame and was attached to the edge of a table, which allowed changing the inclination of the screen relative to the table. We chose an angle of about 38° between the screen and the table. Two empty boxes (2 × 2 cm) were displayed on a black background, and the distance from the fixation circle in the screen centre to the centre of the boxes was about 7.8 cm. A cross was displayed 7.8 cm below the fixation circle and served as a starting position for reaching movements. The imperative stimulus was a green or red square of about 1.7 × 1.7 cm that was presented in one of the two boxes (see Figure 1).
Procedure
The experiment took place in a dimly lit room. Participants’ eyes were at a distance of approximately 46 cm from the flat panel screen. A distance of 1 cm on the screen corresponded to approximately 1.2° of visual angle. We instructed participants to respond as fast and as accurately as possible to the colour of the square, irrespective of its location. In the finger-lift condition, trials were initiated by placing both index fingers on the screen. The left and right index fingers were placed about 3 cm below the left and right boxes, respectively. Participants responded by lifting either the left or the right index finger. 1 In the reaching condition, the screen was touched with the right index finger only. A trial began when the index finger was on the cross. Participants were instructed to lift the finger from the screen surface, move the hand toward the correct location, and touch the box with their index finger. In both response modes, the coloured square appeared randomly between 0.3 and 1.3 s after the trial was initiated. After 1.5 s, the coloured square vanished, and a new trial could be initiated.
A further experiment was run to consider potential differences between the current symbolic finger-lift responses from more typical key-press responses. The apparatus and the stimuli were the same as those in the unconstrained condition of the present study with the exception that eye movements were not measured. Finger-lift and key-press responses were performed on the screen, about 6 cm below the lateralized boxes. The results indicated that reaction times and the mean Simon effect were similar in finger-lift and key-press responses: mean reaction times, 424 and 416 ms, F(1, 9) = 0.28, MSE = 2,140.79, p = .61; mean Simon effect, 19 and 15 ms, F(1, 9) = 0.37, MSE = 132.84, p = .56.
Finger-lift and reaching responses were performed in three eye movement conditions. In the unconstrained condition, no instruction concerning eye movements was given. The aim of this condition was to record the spontaneous eye movements of participants. In the fixation and saccade conditions, eye movements were constrained. In the fixation condition, gaze should stay on the fixation circle from the beginning of the trial until the end of the manual response. In the saccade condition, participants were required to look at the fixation circle until stimulus presentation. When the coloured square appeared, they were required to make a saccade to the stimulus location (see Figure 1). Participants were told that they could move their eyes as they wanted after the first obligatory saccade. Error feedback (see Analyses section for error types) was displayed on the screen, and the respective trial was repeated in the remainder of the experiment.
Design
There were three separate sessions on separate days during two weeks. In the first session, all participants performed the Simon task without eye movement constraints. In the second session, half of the participants performed the fixation and the other half the saccade condition. In the third session, the remaining eye movement instruction was administered. The colour–response mapping and the order of response modes (“finger-lift” vs. “reaching”) in each block was balanced across participants. Each block was preceded by about 10 practice trials. In each block, the four possible combinations of two square locations and two square colours (or response locations) were repeated 50 times for a total of 200 trials per response mode.
Analyses
For both manual responses, we defined movement onset as the first sample exceeding a velocity of 50 mm/s with the constraint that the velocity stayed above this value for at least 100 ms. Movement offset was defined differently for finger-lift and reaching responses. For reaching responses, a position criterion of 4 mm to the screen was used. This position criterion could not be adopted for finger-lift responses, given that participants were only instructed to lift the finger, but not to immediately lay it back on the screen. Most participants kept the finger in the upper position for some time before lowering it again. Therefore, we defined the offset of the movement as the sample at which peak velocity was reached. This sample occurred while raising the finger, close to the highest position. Finally, we used the Eyelink II parser to detect saccades, but only considered saccades with amplitudes larger than 2.5° (2.1 cm). Remember that the targets were presented at 9.4°. Saccades smaller than 2.5° were therefore likely to represent corrections of fixation, rather than changes in the object that was looked at.
Several ocular and manual errors were determined. For all eye movement conditions, blinking during stimulus presentation and saccades before stimulus presentation were counted as errors. In the fixation condition, saccades larger than 2° were considered fixation errors. In the saccade condition, saccadic latencies longer than 800 ms were considered missing, and first saccades not directed toward the coloured stimulus were considered saccadic choice errors. Manual latencies shorter than 100 ms and longer than 800 ms were anticipations and missed trials, respectively. In the finger-lift condition, manual choice errors were trials in which the wrong index finger was lifted. In the reaching condition, manual choice errors were trials in which the wrong box was touched; hand movements should not glide along the screen (i.e., the hand/finger had to be lifted). Finally, loss of the ultrasonic signal was detected.
Mean RTs (see Table 1 and Figure 2) and the percentage of choice errors for manual responses were calculated for corresponding and noncorresponding trials. The unconstrained condition was analysed separately from the other two conditions, because block order was confounded (i.e., the unconstrained condition was always run first). We ran analyses of variance (ANOVAs) on RTs and on the percentage of choice errors. The remaining error types were excluded from analysis. We corrected the degrees of freedom using Huynh–Feldt's correction when the assumption of sphericity was violated. In follow-up analysis, paired-samples t tests were conducted.
Mean reaction time and between-subject standard error for corresponding and noncorresponding trials
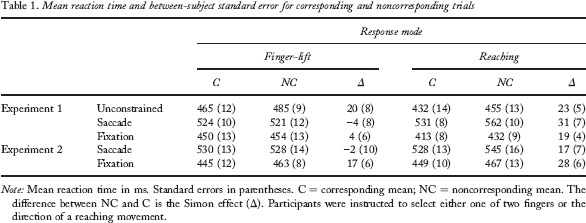
Note: Mean reaction time in ms. Standard errors in parentheses. C = corresponding mean; NC = noncorresponding mean. The difference between NC and C is the Simon effect (δ). Participants were instructed to select either one of two fingers or the direction of a reaching movement.
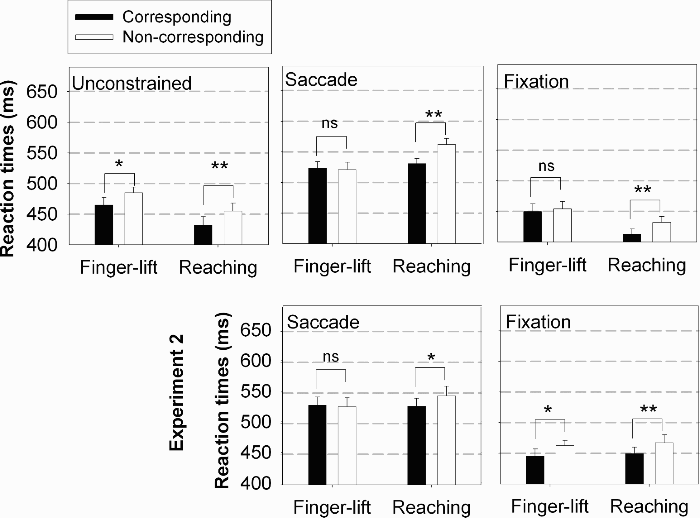
Mean reaction times in the unconstrained, saccade, and fixation conditions of Experiments 1 and 2 as a function of correspondence (corresponding and noncorresponding) and response type (finger-lift and reaching). The p values below the .05 and .01 levels are indicated by one (*) and two (**) asterisks, respectively. Nonsignificant p values are indicated by ns.
With respect to eye movements, we observed that participants performed two saccades at maximum. The percentage of first and second saccades for Experiments 1 and 2 was calculated for each eye movement and response mode, as a function of correspondence (Figure 3, black dots). More precisely, the percentages of first and second saccades were obtained by dividing the number of trials with a saccade by the overall number of trials in the respective condition. Then, we also evaluated the percentage of saccades opposite to the imperative stimulus location, for corresponding and noncorresponding trials (Figure 3, white dots). To obtain the total proportion of opposite saccades (see Figure 3, rightward part of each graph), we summed up percentages of opposite saccades obtained in the first and second saccades when the first saccade could be directed away from the stimulus (i.e., in the unconstrained condition of Experiment 1). Otherwise, the first saccade was always directed toward the stimulus (by instruction), such that the percentage of “total opposite” is equal to the percentage of second saccades away from the stimulus.
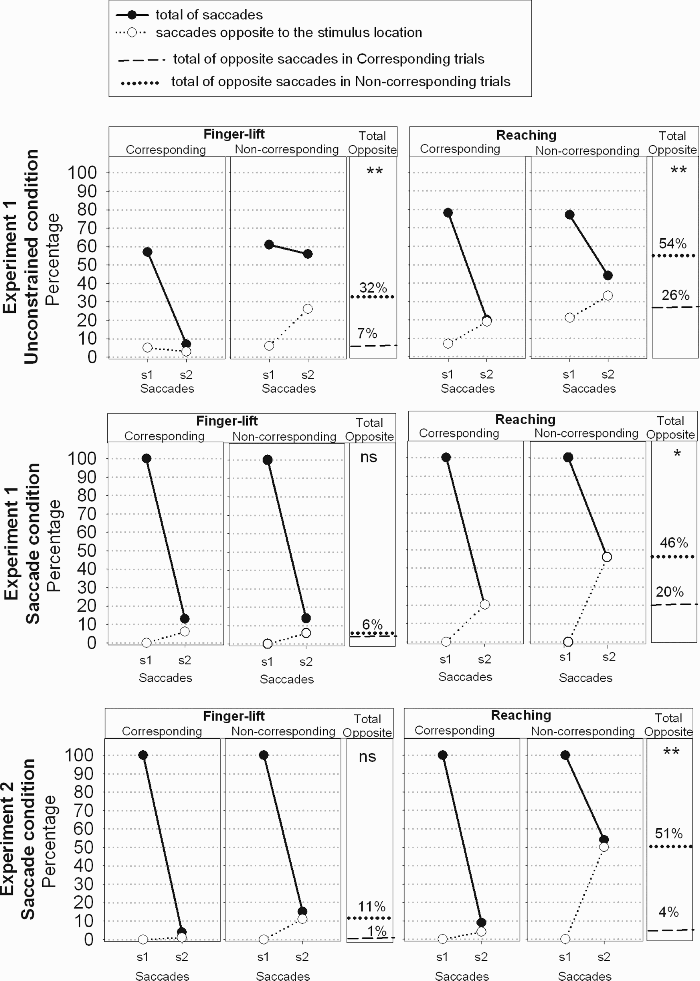
Percentages of saccades in the finger-lift and reaching conditions of Experiments 1 and 2. The filled circles show the percentage of first and second saccades (s1 and s2, respectively). Unfilled circles show the percentage of saccades directed away from the imperative stimulus. All percentages were calculated with respect to the overall number of trials in a given condition. The graph on the right of each subgraph (“total opposite”) shows the total percentage of saccades that were directed opposite to the imperative stimulus in corresponding and noncorresponding trials. We compared the percentages of “total opposite” saccades by t test. Probabilities below the .01 and .001 level are indicated by one (*) and two (**) asterisks, respectively. Nonsignificant p values are indicated by ns.
The percentage of opposite saccades is fundamental to our hypothesis. We started from the assumption that in any case, participants had to shift their eyes or attention toward the imperative stimulus to gather the relevant information. However, each saccade opposite to the stimulus location is considered as an additional process that lengthens the time to response selection. We chose to sum up percentages of first and second saccades in the unconstrained condition of Experiment 1 because it quantifies “saccade time” in corresponding and noncorresponding trials. Finally, an ANOVA was run on the total percentages of saccades opposite to the stimulus location. We also verified whether a Simon effect was present in saccadic latencies of the first saccade, but no significant effects were observed (ANOVA results not shown). Saccadic latencies of the first and second saccades are shown in Figure 4.
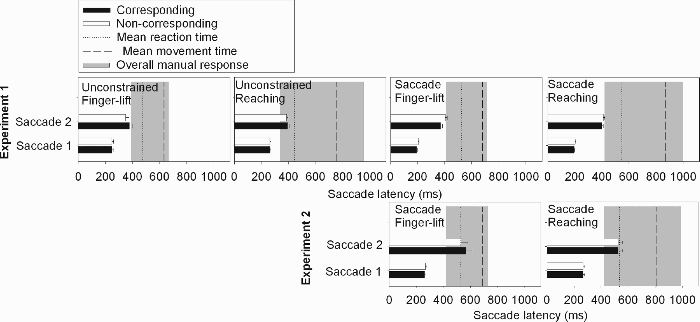
Saccadic latency (ms) for the first and second saccades as a function of correspondence. The dotted line represents mean reaction time (i.e., movement initiation), and the dashed line the mean movement time (i.e., end of movement execution). The grey surface indicates the interval during which the hand was in motion, starting at 2.5% of the fastest reaction times and ending at 97.5% of the slowest movement times.
Results
Unconstrained eye movements
RTs
A two-way ANOVA (Response Mode × Correspondence) on manual RTs showed longer RTs with finger-lift than with reaching responses (475 vs. 443 ms, respectively), F(1, 15) = 13.41, MSE = 1,189.43, p < .01, and in noncorresponding than in corresponding trials (470 vs. 448 ms, respectively), F(1, 15) = 11.44, MSE = 626.76, p < .01. The interaction between response mode and correspondence did not reach significance, F(1, 15) = 0.29, MSE = 140.79, p = .60. The Simon effect was significant with finger-lift (20 ms), t(15) = –2.36, p < .05, and with reaching responses (23 ms), t(15) = –4.36, p < .001. Figure 2 plots the Simon effect for the three eye movement conditions as a function of response mode.
Errors
A two-way ANOVA (Response Mode × Correspondence) on the percentage of choice errors indicated a larger error rate in noncorresponding than in corresponding trials (4% vs. 3%, respectively), F(1, 15) = 5.91, MSE = 0.001, p < .05. The main effect of response mode and the interaction between response mode and correspondence were not significant (Fs < 0.67, MSE = 0.001).
Eye movement analyses
We analysed the percentage of trials in which a saccade opposite to the stimulus location (henceforth referred to as “opposite saccades”) was performed in corresponding and noncorresponding trials (see Figure 3, upper row). The two-way ANOVA (Response Mode × Correspondence) showed a larger rate of opposite saccades in reaching than in finger lift responses (40% vs. 19%), F(1, 15) = 21.85, MSE = 324.90, p < .001, and in noncorresponding than in corresponding trials (43% vs. 17%), F(1, 15) = 107.63, MSE = 102.43, p < .001. The difference between corresponding and noncorresponding trials was similar with finger-lift and reaching responses (24% vs. 28%), F(1, 15) = 0.51, MSE = 52.56, p = .49, and significant for both response modes, t(15) = –7.97, p < .001, and t(15) = –6.95, p < .001, respectively. Thus, in noncorresponding trials, saccades away from the irrelevant stimulus toward the correct response location occurred regardless of response mode.
Fixation and saccade conditions
RTs
We performed a three-way ANOVA (Response Mode × Eye Movement Condition × Correspondence) on manual RTs, yielding a large number of significant main effects and two-way interactions. Importantly, all significant effects were qualified by a three-way interaction. To avoid repetitions, we forgo reporting the full ANOVA in the text and refer to Table 2. Instead, we present the results of follow-up ANOVAs (Eye Movement Condition × Correspondence) for each response mode separately.
Statistics of the three-way ANOVA on reaction times and choice errors for Experiments 1 and 2
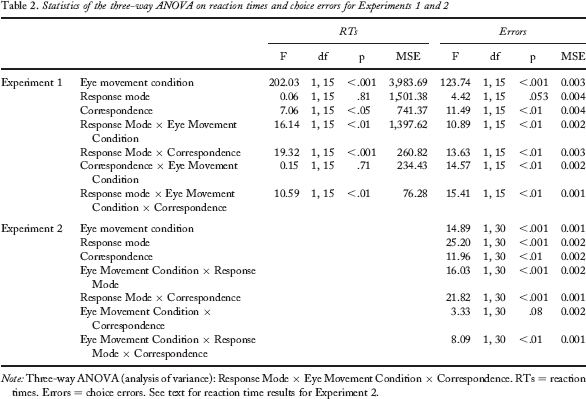
Note: Three-way ANOVA (analysis of variance): Response Mode × Eye Movement Condition × Correspondence. RTs = reaction times. Errors = choice errors. See text for reaction time results for Experiment 2.
For reaching responses, RTs were shorter with fixation than with saccades (422 vs. 546 ms, respectively), F(1, 15) = 275.35, MSE = 892.09, p < .001, presumably because of the dual-task situation when a saccade had to be executed. RTs were shorter in corresponding than in noncorresponding trials (472 vs. 497 ms, respectively), F(1, 15) = 25.70, MSE = 399.68, p < .001. The interaction of eye movement condition and correspondence was nearly significant (a Simon effect of 19 ms and 31 ms was found for the saccade and fixation conditions, respectively), F(1, 15) = 4.10, MSE = 142.88, p = .06 (see Figure 2).
For finger-lift responses, RTs were also shorter in the fixation than in the saccade condition (452 vs. 523 ms, respectively), F(1, 15) = 39.98, MSE = 2,006.90, p < .001. Surprisingly, the Simon effect was absent (487 ms for both corresponding and noncorresponding trials), F(1, 15) = 0.01, MSE = 602.51, p = .97. Finally, the effect of eye movement condition was not qualified by correspondence, F(1, 15) = 1.52, MSE = 167.83, p = .24. The mean Simon effect was about –4 ms in the saccade condition and 4 ms in the fixation condition (see Figure 2).
Errors
The three-way ANOVA (Response Mode × Eye Movement Condition × Correspondence) again showed many significant effects, including the three-way interaction. To avoid redundancies, statistics of the three-way ANOVA are shown in Table 2.
For reaching responses, the two-way ANOVA (Eye Movement Condition × Correspondence) showed that the error rate was larger in the saccade than in the fixation condition (17% vs. 3%, respectively), F(1, 15) = 73.02, MSE = 0.004, p < .001, and in noncorresponding than in corresponding trials (14% vs. 6%), F(1, 15) = 23.21, MSE = 0.004, p < .001. Correspondence was also qualified by eye movement condition, showing that the difference between corresponding and noncorresponding trials was larger in the saccade than in the fixation condition (13% vs. 2%, respectively), F(1, 15) = 19.43, MSE = 0.003, p < .001. This means that participants had a strong tendency to reach to where they looked. In noncorresponding trials, reaching toward the endpoint of the first saccade (the stimulus) resulted in an error.
For finger-lift responses, choice errors were more frequent in the saccade than in the fixation condition (12% vs. 3%, respectively), F(1, 15) = 120.03, MSE = 0.001, p < .001. Neither the main effect of correspondence, F(1, 15) = 0.08, MSE = 0.003, p = .78, nor the interaction between correspondence and eye movement condition reached significance, F(1, 15) = 0.52, MSE = 0.001, p = .48.
Eye movement analyses (saccade condition only)
The ANOVA (Response Mode × Correspondence) on the percentage of saccades opposite to the stimulus location showed more opposite saccades in reaching than finger-lift responses (33% vs. 6%), F(1, 15) = 47.63, MSE = 251.19, p < .001, and in noncorresponding than in corresponding trials (26% vs. 13%), F(1, 15) = 16.58, p < .001 (see Figure 3, middle row). Finally, the difference between corresponding and noncorresponding trials was larger in reaching than in finger-lift responses (26% vs. 0%), F(1, 15) = 13.23, p < .01, and was only significant with reaching responses, t(15) = –3.9, p < .001.
Further, we wanted to know whether the percentages of opposite saccades observed when eye movements were constrained (saccade condition) differed from the ones found in the unconstrained condition. For corresponding trials, t tests indicated a similar percentage of redirections in the unconstrained and saccade conditions, both for finger-lift and for reaching responses: mean differences of 1%, t(15) = 0.30, p = .77, and 6%, t(15) = 1.69, p = .11, respectively. For noncorresponding trials, the percentage of opposite saccades was similar in the unconstrained and saccade conditions when reaching responses were executed (difference of 8%), t(15) = 1.01, p = .33, but differed with finger-lift responses (difference of 25%), t(15) = 4.99, p < .001. In sum, observers behaved as during unconstrained viewing when asked to saccade and reach, but behaved differently when asked to saccade and lift the finger. Interestingly, the size of the Simon effect in the unconstrained and saccade conditions was unchanged for reaching responses (23 and 31 ms), t(15) = –1.5, p = .15, but differed for finger-lift responses (20 and –4 ms), t(15) = 3.21, p < .01. Hence, occurrence of the Simon effect was coupled to the occurrence of saccades toward the correct response location in noncorresponding trials.
Discussion
The aim of the present study was to investigate whether the execution of saccades contributed to the Simon effect. We expected that more saccades opposite to the stimulus location in noncorresponding trials increase the Simon effect because each saccade takes time. Conversely, similar rates of opposite saccades in corresponding and noncorresponding trials should diminish the Simon effect.
We replicated the Simon effect in manual RTs and error rates when the task was performed without any oculomotor constraints: RTs were longer, and responses were less accurate in noncorresponding than in corresponding trials. Surprisingly, the Simon effect in RTs and errors completely disappeared with finger-lift responses when eye movements were controlled (saccade and fixation condition). In contrast, it persisted with reaching movements. Because the automatic activation of response codes by the irrelevant stimulus location is known to decay over time (Hommel, 1994), one may argue that the relatively long RTs eliminated the Simon effect in the saccade condition with finger-lift responses. However, RTs were just as long as those with reaching responses, and a Simon effect was confirmed in this condition. Thus, the absolute RT values cannot explain the absence of a Simon effect with finger-lift responses in the saccade condition. Analysis of eye movements suggests an alternative explanation. Notably, all conditions showing a significant Simon effect presented a larger rate of saccades opposite to the stimulus location in noncorresponding than in corresponding trials (difference of more than 24%). This agrees with our hypothesis that a larger number of saccades slows down response selection in noncorresponding trials. Of course, the relationship between saccades and Simon effect could only be verified in conditions in which eye movements were allowed. However, we believe that the same covert redirection of attention occurred when fixation was maintained, but we have no evidence to support this other than that the RT data with fixation mirrored the RT data with an initial saccade to the stimulus.
Further, with respect to previous findings indicating a strong coupling between manual and ocular systems (e.g., Prablanc et al., 1979), our results confirmed that observers performed more saccades with reaching than with finger-lift responses. In the unconstrained eye movement condition, the eyes were directed toward the correct response location on a substantial proportion of noncorresponding trials. Interestingly, this was true for both reaching and finger-lift responses. These redirections toward the correct response location in noncorresponding trials may have facilitated the selection of the target on the screen in reaching responses or the selection of the effector in finger-lift responses (remember that the index fingers were placed on the screen just below the lateralized boxes). Unexpectedly, we also observed saccades opposite to the imperative stimulus location in corresponding trials, especially with reaching responses. The purpose of these saccades away from the imperative stimulus in corresponding trials remains unclear.
Furthermore, the eye movement behaviour changed drastically with finger-lift responses and the constraint to saccade to the imperative stimulus. Instead of redirecting gaze at the correct location in noncorresponding trials, participants looked at the imperative stimulus and stayed there. In contrast, the pattern of eye movements with reaching movements was the same as that in the unconstrained condition: In noncorresponding trials, the eye was redirected to the correct location after the initial saccade to the imperative stimulus. The difference between response modes may be explained by the necessity to select the to-be-reached target location as the goal of the reaching response and the absence of a goal-directed movement with finger-lift responses.
In sum, the Simon effect in Experiment 1 vanished with saccades to the stimulus and symbolic responses. The reduced number of saccades in this condition may account for the absence of the Simon effect. However, given the novelty of the present findings, we wished to replicate the main results of Experiment 1. In Experiment 2, we ran the saccade and fixation conditions with finger lift and reaching responses but decided to test separate groups of participants in each condition (i.e., a between-subjects design).
EXPERIMENT 2
Method
Participants
A total of 16 students (10 females, mean age of 22 years, right-handed) from the same pool as that in Experiment 1 participated in the fixation condition, and another 16 students in the saccade condition.
Apparatus and stimuli
The same apparatus as that in Experiment 1 was used.
Procedure, design, and analyses
Two different groups of participants performed the saccade and fixation conditions. Each group performed the two types of manual response (finger-lift and reaching conditions). The design and analyses were the same as those in Experiment 1 with the exception that separate groups of participants performed the saccade and fixation conditions.
Results
RTs
We ran a three-way ANOVA (Eye Movement Condition × Response Mode × Correspondence) on manual RTs with eye movement condition as between-subjects factor. All main effects and interactions are discussed here because the triple interaction did not reach significance. Results showed shorter RTs in the fixation than in the saccade condition (454 vs. 533 ms, respectively), F(1, 30) = 25.52, MSE = 1,953.55, p < .001, which is probably due to the dual-task situation in the saccade condition. RTs did not differ between finger-lift and reaching (492 vs. 495 ms, respectively), F(1, 30) = 0.26, MSE = 1,606.45, p = .61. RTs in corresponding trials were shorter than those in noncorresponding trials (456 vs. 501 ms, respectively), F(1, 30) = 11.51, MSE = 621.71, p < .01; however, correspondence did not interact significantly with the eye movement condition, F(1, 30) = 2.86, MSE = 621.71, p = .10. Importantly, the difference between corresponding and noncorresponding trials was larger in reaching than in finger-lift responses (23 vs. 8 ms, respectively), F(1, 30) = 8.59, MSE = 212.58, p < .01 (see Figure 2); t tests on each response mode (across eye movement condition) confirmed that the Simon effect was significant with reaching but not with finger-lift responses, t(31) = –4.93, p < .001, and t(31) = –1.28, p = .21, respectively.
Unlike in Experiment 1, the interaction between eye movement condition and response mode did not reach significance, F(1, 30) = 0.33, MSE = 1,606.45, p = .57. Despite the absence of interaction between correspondence and eye movements, we carried out t tests on each response mode and eye movement instruction to check whether the results of Experiment 2 replicated the null Simon effect observed in Experiment 1 when finger-lift responses were performed. During eye fixation, the Simon effect was present with reaching (28 ms), t(15) = –4.44, p < .001, and with finger-lift responses (17 ms), t(15) = –2.91, p < .05. When a saccade had to be made to the stimulus, t tests showed a significant Simon effect with reaching (17 ms), t(15) = – 2.63, p < .05, but not with finger-lift responses (–2 ms), t(15) = 0.25, p = .80, replicating the results of Experiment 1.
Errors
The three-way ANOVA (Eye Movement Condition × Response Mode × Correspondence) with eye movement condition as between-subjects factor showed that all effects were significant (see Table 2). To avoid repetitions, we only discuss results of the follow-up ANOVAs for each response mode separately. The two-way ANOVA (Response Mode × Correspondence) on the fixation condition showed no significant main effect: response mode, F(1, 15) = 0.74, MSE = 0.001, p = .40; correspondence, F(1, 15) = 2.97, MSE = 0.001, p = .11. The interaction between response mode and correspondence was significant, indicating a larger Simon effect with reaching than with finger-lift responses (2% vs. 0%, respectively), F(1, 15) = 7.25, MSE = 0.001, p < .05. The two-way ANOVA (Response Mode × Correspondence) on the saccade condition showed a larger error rate with reaching than with finger-lift responses (11% vs. 4%, respectively), F(1, 15) = 31.10, MSE = 0.002, p < .001, and in noncorresponding than in corresponding trials (9% vs. 6%, respectively), F(1, 15) = 9.00, MSE = 0.003, p < .01. The response mode by correspondence interaction confirmed a larger Simon effect with reaching than with finger-lift responses (8% vs. –1%, respectively), F(1, 15) = 15.96, MSE = 0.002, p < .01. Participants had a strong tendency to reach to the endpoint of the first saccade (the stimulus) in noncorresponding trials, whereas they were far less inclined to lift the finger close to the saccadic endpoint. Again, this points to strong eye–hand coupling with reaching movements.
Eye movement analyses (saccade condition only)
The ANOVA (Response Mode × Correspondence) on the percentage of saccades opposite to the stimulus location indicated a larger rate of opposite saccades with reaching than with finger-lift responses (28% vs. 6%), F(1, 15) = 25.44, MSE = 285.66, p < .001, and in noncorresponding than in corresponding trials (31% vs. 3%), F(1, 15) = 25.57, MSE = 510.43, p < .001 (see Figure 3, lower row). The difference between corresponding and noncorresponding trials was larger with reaching than with finger-lift responses (47% vs. 10%), F(1, 15) = 35.59, MSE = 222.82, p < .001, and was only significant for reaching, t(15) = –5.78, p < .001, and t(15) = –2.03, p = .07, for reaching and finger-lift responses, respectively. Interestingly, t tests confirmed that the Simon effect in RTs in reaching responses was also larger than that in finger-lift responses (17 vs. –2), t(15) = 3.17, p < .01.
Discussion
In Experiment 2, two different groups of participants performed saccade and fixation conditions. The aim was to confirm the modulation of the Simon effect by the number of saccades performed in corresponding and noncorresponding trials. The main results of Experiment 1 were replicated in Experiment 2. The Simon effect in RTs and in manual error rates was larger with reaching than with finger-lift responses. With reaching responses, the Simon effect was significant in the fixation and saccade conditions (28 and 17 ms, respectively). With finger-lift responses, the Simon effect was present in the fixation condition (17 ms) but not in the saccade condition (–2 ms).
Analysis of saccades indicated that the difference in the number of saccades performed in corresponding and noncorresponding trials distinguished conditions in which a Simon effect occurred from conditions in which it did not. With reaching responses, we found substantial redirection of saccades in noncorresponding trials and a significant Simon effect. With finger-lift responses, fewer saccades opposite to the stimulus location occurred, and the Simon effect approached zero.
GENERAL DISCUSSION
In the present study, we provided an integrated evaluation of the perception–action relationship in the Simon effect by recording simultaneously two different types of hand movements (reaching and finger-lift responses) under three different eye movement instructions (unconstrained, fixation, saccade to stimulus). Despite the large number of goal-directed actions that we perform in everyday life, most studies investigated S–R compatibility with symbolic responses and central fixation. While symbolic responses can be executed without shifting the eyes toward the target location, goal-directed responses require the eye (e.g., Prablanc et al., 1979) and attention (Baldauf & Deubel, 2008; Baldauf, Wolf, & Deubel, 2006) to move towards the target before the response is executed. Similarly, the dominant mechanism for reaching responses is thought to be visuospatial attention, while the dominant mechanism for key-presses is effector selection (Adam & Pratt, 2004; Adam et al., 2008).
We sought to clarify whether the type of manual response and the occurrence of eye movements modulate the Simon effect. Our basic idea was that more saccades occur in noncorresponding than in corresponding trials: Initially, a saccade is made to the task-relevant stimulus. In noncorresponding trials, a second saccade is then made to the correct (but empty) response location. The greater number of saccades in noncorresponding than in corresponding trials (two vs. one) delays response selection in noncorresponding trials and contributes to the Simon effect.
Under free viewing conditions (unconstrained condition, Experiment 1), the pattern of saccades was similar for reaching and finger-lift responses. More saccades away from the stimulus were made in noncorresponding than in corresponding trials. In addition, the Simon effect was of comparable size with reaching and finger-lift responses. However, making a saccade to the stimulus (saccade condition, Experiments 1 and 2) eliminated the Simon effect with finger-lift responses, but not with reaching responses. We believe that the number of saccades accounts for this difference. In the saccade condition, participants had to first look at the stimulus. Subsequent to the first saccade, behaviour in noncorresponding trials differed between finger-lift and reaching responses. With finger-lift responses, the eye did not move any further, even though the correct response location was opposite to the stimulus. When the hand had to reach the correct response location, a second saccade was made from the stimulus to the correct response location because of eye–hand coupling. As the Simon effect only occurred when saccades away from the stimulus occurred in a substantial number of trials, we conclude that the longer RTs in noncorresponding trials are attributable to the delay caused by the execution of saccades from the stimulus to the correct response location. Thus, the presence or absence of the Simon effect depends on the number of saccades: When a similar number of saccades is performed in corresponding and noncorresponding trials, the Simon effect is null, and when the number of saccade is larger in noncorresponding than in corresponding trials, the Simon effect is positive.
Even if we only confirmed this pattern with saccades, we speculate that it also holds for covert shifts of attention in the fixation condition. More precisely, we suppose that in the fixation condition of Experiment 1 the Simon effect did not occur because participants (covertly) attended the imperative stimulus in corresponding and noncorresponding trials (i.e., one shift of attention in both conditions). Conversely, the Simon effect was present in Experiment 2 because participants performed a supplementary shift of attention toward the correct response location (or toward the effector) in noncorresponding trials.
Comments about the generalization of the present findings and conclusions
We are aware that our task is not typical with respect to the majority of studies in the field. For instance, in the conditions in which eye movements were constrained, participants had to attend not only to their hand movements as in typical Simon tasks but also to their eye movements. Further, the response locations were also different in our tasks. Reaching and finger-lift responses occurred on the screen, and this may increase the dimensional overlap between stimulus and responses (Adam et al., 2008) and therefore the size of the Simon effect. Finally, the saccade task created a dual-task situation, which may also affect the size of the Simon effect according to models assuming a decay of activation by the irrelevant stimulus (Hommel, 1993; Kornblum et al., 1990). Even if not typical, the characteristics of our task are essential to investigate the Simon effect in more naturalistic settings. Indeed, looking at and reaching objects in our environment are frequent activities of our daily life. The surprising result of our study is that when a task-relevant object is looked at (which is the most likely response outside the lab), symbolic responses are not affected by the spatial location of the stimulus (i.e., no slowing if the object is in a noncorresponding spatial location). Although the generalization of the present findings to the Simon effect literature is limited, the implications for more ecological situations are nonetheless significant.
We are also conscious that our explanation contrasts with popular cognitive models of human performance because it does not focus on automatic and intentional response codes (Hommel, 1993; Kornblum et al., 1990). However, we believe that our account provides the simpler framework for summarizing the present results. Besides, we should keep in mind that only Simon tasks using horizontal visual stimuli and manual responses with parallel arms confirmed the assumption of automatic activation (e.g., Vallesi, Mapelli, Schiff, Amodio, & Umiltà, 2005). However, studies using auditory stimuli, vertical visual stimuli, or crossed-hand responses mostly observed stable Simon effect functions and, more importantly, no automatic activation over the motor cortex but only a delay in response selection in noncorresponding trials (e.g., Vallesi et al., 2005; Wascher, Schatz, Kuder, & Verleger, 2001; Wiegand & Wascher, 2005). Thus, our study is not the first to indicate that the Simon effect in reaction times is modulated by task demands.
Related findings
In a further analysis, we observed that the initial trajectory of reaching responses deviated toward the irrelevant spatial position (see Appendix). This effect has been amply described in two previous papers (Buetti & Kerzel, 2008, 2009). However, the present study suggests that this deviation may be more related to (covert or overt) shifts of attention that precede response execution. This idea is consistent with research on trajectory deviations of saccadic and manual movements after shifts of attention (Sheliga, Riggio, Craighero, & Rizzolatti, 1995; Tipper, Howard, & Houghton, 1998; Welsh & Elliott, 2004).
A last remark concerns the implication of current findings for response selection models. We are not the first to report that response selection depends not only on the relation between stimulus and response sets but also on the response type. For instance, Fagioli, Hommel, and Schubotz (2007) confirmed priming of action-related features: Processing of shape was favoured with grasping responses while location information was privileged with reaching responses. Modulations of cognitive processes by the response type were also shown in other paradigms, such as precueing tasks (e.g., Adam et al., 2008) and inhibition of return tasks with saccadic responses (e.g., Souto & Kerzel, 2009). Thus, to understand the human ability to select and control actions, it is necessary to consider the specific mechanisms underlying each response type.
To summarize, our study investigates the role of saccades in the Simon effect. Interestingly, the Simon effect vanished when a saccade toward the stimulus was performed, and a symbolic response was required, while it persisted with reaching movements. We claim that the redirection of attention toward the correct response location contributes to the Simon effect. The redirection did not occur with saccades and symbolic responses, but did occur with saccades and reaching responses because of eye–hand coupling.