
Abstract
Selective attention can be improved under conditions in which a high perceptual load is assumed to exhaust cognitive resources, leaving scarce resources for distractor processing. The present study examined whether perceptual load and acute stress share common attentional resources by manipulating perceptual and stress loads. Participants identified a target within an array of nontargets that were flanked by compatible or incompatible distractors. Attentional selectivity was measured by longer reaction times in response to the incompatible than to the compatible distractors. Participants in the stress group participated in a speech test that increased anxiety and threatened self-esteem. The effect of perceptual load interacted with the stress manipulation in that participants in the control group demonstrated an interference effect under the low perceptual load condition, whereas such interference disappeared under the high perceptual load condition. Importantly, the stress group showed virtually no interference under the low perceptual load condition, whereas substantial interference occurred under the high perceptual load condition. These results suggest that perceptual and stress related demands consume the same attentional resources.
We sometimes feel that cognitive operations are effortful and beyond our capability, especially when these operations are difficult. The perspective that attention can be likened to a limited resource or capacity (Norman & Bobrow, 1975) predicts successful completion of a task as long as the total amount of the resource required is less than the maximum that is available. According to this principle, participants can perform multiple tasks within the limits of available resources. When the task demands exceed such limits, however, impaired performance ensues.
The finding that the distractor interference observed when a target appears alone under the low perceptual load condition is reduced when the target is embedded among nontarget items under the high perceptual load condition supports this perspective (Lavie, 1995). This result implies that no spare resources were left for distractor processing (i.e., no distractor interference) under the high perceptual load condition. In other words, attentional selectivity as indexed by distractor interference depends on the amount of resources available. As perceptual load is not a single determinant that modulates the availability of resources, other internal factors such as stress, fatigue, and reduced motivation should also modulate attentional selectivity.
We focused specifically on the effect of acute stress on attentional selectivity because stress research has assumed that acute stress consumes resources used in efforts to selectively direct attention (Chajut & Algom, 2003; Keinan, Friedland, Kahneman, & Roth, 1999). Thus, the present question is whether perceptual load and acute stress drain a common attentional resource. If this were the case, these two factors would interact, and stress may enhance attentional selectivity under optimal conditions.
Studies using the negative priming effect as an index of selective attention have generally supported this prediction. For example, Skosnik, Chatterton, Swisher, and Park (2000) found reduced negative priming (i.e., less selectivity) under stressful circumstances relative to instances in which no stress manipulations were introduced. Similarly, Braunstein-Bercovitz (2003) demonstrated that the effect of negative priming was reduced by high stress under a low perceptual load condition, implying impaired selectivity. Interestingly, high stress improved selectivity under the high perceptual load condition in that study. Although the original account of negative priming posited that this effect reflected the inhibition of attention directed at irrelevant objects (e.g., Neill, 1977; Tipper, 1985), some researchers have argued that the effect reflects memory retrieval rather than inhibitory mechanisms involved in selective attention (Mayr & Buchner, 2006; Neill, Valdes, & Terry, 1995). Thus, it is premature to interpret the results of the negative priming experiments as implying an interaction between stress and perceptual load in selective attention. To determine whether these two factors interact, the aforementioned results must be understood in the context of evidence gathered from a different perspective.
Thus, the present study examined whether stress and perceptual load interact by using a flanker-interference effect (Eriksen & Eriksen, 1974), which has been established as a hallmark of impaired spatial selective attention. In this task, reaction times for identifying a target are delayed when the flanker letters (i.e., incompatible distractors) are associated with an inconsistent response to the target relative to when the flankers (i.e., compatible distractors) are associated with a consistent response to the target. We defined the differences between the reaction times for identifying a target in the incompatible trials and those for doing so in the compatible trials as the interference effect. Lower levels of attentional selectivity reflect greater interference from the distractors and greater differences in reaction times between the incompatible and compatible conditions. The relevant literature shows that interference is decreased when the perceptual load is high (Lavie, 1995; Lavie & de Fockert, 2003). Following the procedures used in these studies, we varied the number of nontarget items to manipulate the perceptual load in the present study.
We used the Trier Social Stress Test (TSST; Kirschbaum, Pirke, & Hellhammer, 1993), in which participants delivered a brief speech and received a mental calculation task while a video recorder and an examiner recorded participants' nonverbal behaviours to manipulate acute stress. It has been shown that oral presentations are stressful for students, and the impact of such stress is reflected in salivary cortisol concentrations (Preuss, Schoofs, Scholtz, & Wolf, 2010). We predicted that the interference effect would be decreased under the high perceptual load relative to the low perceptual load condition in the low stress group, as demonstrated by previous studies (e.g., Lavie, 2005). Critically, when stress is high, interference would be decreased the under the low perceptual load condition because the stress would consume the residual resources.
Method
Participants
Thirty-nine Japanese male participants, recruited from the National Institute of Advanced Industrial Science and Technology (AIST, Tsukuba, Japan) subject pool participated in exchange for monetary remuneration. Participants were randomly assigned to the stress (N = 19, mean age = 21.0 years) or the control (N = 20, mean age = 21.1 years) group. To avoid the effects of a menstrual hormonal cycle, only male subjects were selected. The experiment was conducted in the afternoon (between 14:00 and 17:00). All participants provided written informed consent and were tested individually.
General procedure
The experiment consisted of four stages: pre-stress measurement, stress manipulation (if any), the flanker-interference task, and poststress measurement. The State–Trait Anxiety Inventory (STAI) Form JYZ (Hidano, Fukuhara, Iwawaki, Soga, & Spielberger, 2000) was administered to both groups at the beginning of the experiment. Saliva samples were collected from the stress group. The stress group participated in the stress manipulation for 15 min immediately after the first saliva sampling, whereas the control group engaged in a filler task for the same period of time. Both groups then participated in the flanker-interference task, which was followed by the second STAI questionnaire (and saliva sampling for the stress group only). Participants were debriefed before they left.
Pre- and post-stress measurements
The STAI consists of 40 items, each of which was rated on a 4-point Likert scale, which measure state and trait anxiety. We used only the state-anxiety items (20 items). Half the 20 items measured the presence of anxiety, and the other half measured the absence of anxiety. Saliva samples were collected from the stress group with salivettes after the completion of the STAI questionnaire. The samples were centrifuged at 4,000 g for 5 min, and the filtrates were stored at –40°C. We were unable to collect and measure salivary cortisol from the control group because of budgetary limitations.
Stress manipulation
Each participant was instructed to prepare a five-minute speech regarding his strengths and weaknesses and deliver it in front of an examiner. Five minutes later, the participant was brought to a podium that was located in a different room in which an examiner was waiting. At the beginning of the TSST procedure, the examiner initiated recoding of the speech with a video camera mounted on a tripod in front of the participant. During the speech, the examiner monitored participants' nonverbal behaviour using a tally counter and a checklist that included eye-blink frequency and number of times the participants looked away or down. After the speech, participants completed a five-minute mental calculation task in which they serially subtracted 13 from 1,022 as fast and as accurately as possible. Following each failure, the examiner instructed participants to restart the subtraction from 1,022.
Flanker-interference task
Stimuli for the flanker-interference task (Eriksen & Eriksen, 1974) were displayed as white letters on the black background of a computer monitor. A trial began with a fixation display in which a small circle (radius = 0.5° visual angle), was presented at the centre of the screen for 1,500 ms. The viewing distance was approximately 57 cm. The fixation display was followed by a task display consisting of a target letter (N or Z) embedded in the central array (three columns of three potential locations) of nontargets (Xs). Perceptual load, defined as the number of nontarget stimuli, was set at low or high (two or five nontarget items, respectively; Figure 1). The locations of the targets and nontargets were determined randomly for every trial. Each letter subtended 1.0° (H) × 0.5° (W), and the centre-to-centre distance was 1.2°. The central array was flanked by two distractor letters, one on each side. Under the compatible condition, the distractor letters were identical to the target (e.g., when the target was Z, the distractors were also Zs.) Under the incompatible condition, the distractor letters differed from the target (e.g., when the target was Z, the distractors were Ns). Participants identified the target as quickly and accurately as possible, by pressing one of the designated keys. Reaction times and the identities of the pressed keys were recorded. The task display remained visible until participants responded.
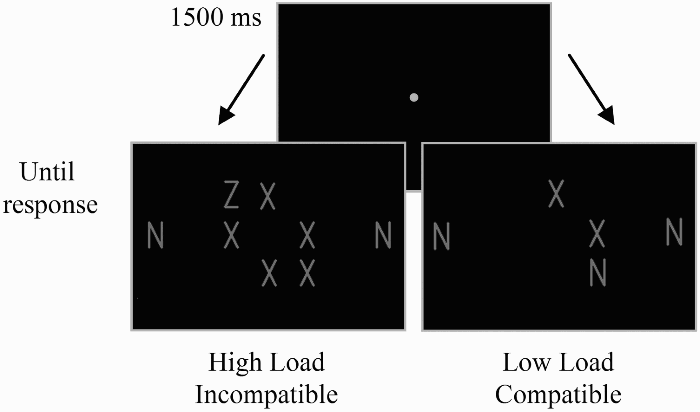
Schematic representation of the stimulus displays.
We used a 2 × 2 × 2 factorial design with one between-subjects factor, stress (present or absent), and two within-subject factors, perceptual load (low or high) and compatibility (compatible or incompatible). Participants engaged in 10 practice trials followed by 144 experimental trials. The flanker-interference task lasted approximately 10 min so that the salivary cortisol response induced by the TSST procedure (Kudielka, Schommer, Hellhammer, & Kirschbaum, 2004) could develop and reach measureable levels by the end of the task.
Results
Stress manipulation
Cronbach's alpha coefficient for the STAI, used as a measure of internal consistency reliability (Cronbach, 1951), was .91. This result indicates adequate internal consistency. The sum of the STAI scores for anxiety presence and anxiety absence (scores reversed) was calculated to evaluate state anxiety and is plotted in Figure 2a as a function of pre- versus post-stress manipulation for each group. A 2 (group: stress vs. control) × 2 (time: pre vs. post) mixed analysis of variance (ANOVA) indicated a significant main effect of time, F(1, 37) = 11.88, p < .01. The interaction between group and time was also significant, F(1, 37) = 6.82, p < .05. An F-test for simple main effects indicated a significant effect for time in the stress group, F(1, 37) = 18.36, p < .01, suggesting that participants in the stress group experienced stronger anxiety during the second measurement than during the first measurement.
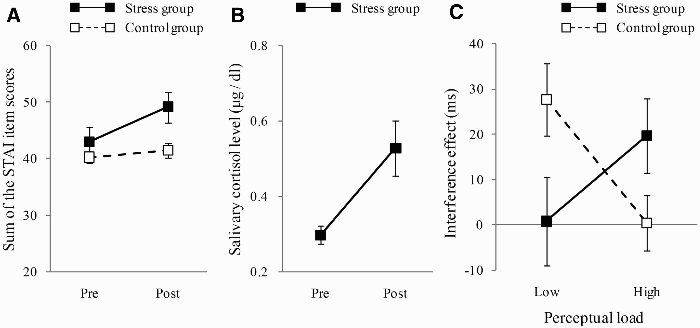
(a) Sum of the STAI (State–Trait Anxiety Inventory) scores and (b) mean salivary cortisol levels for each group on the pre- and post-stress manipulation. (c) Mean flanker interference effect (ms) for high and low perceptual load conditions in the stress and control groups.
The salivary cortisol levels in 200-ml samples of saliva were measured using an RIA (radioimmunoassay) assay (Gamma Coat; Dia Sorin, Stillwater, MN, USA). Assay results showed a range of 0.05–3.00 µg/dl and intra-assay coefficients of variation of <5% and <10% for the pre-stress and post-stress assays, respectively. Figure 2b presents cortisol concentrations in the stress group as a function of time. A within-participant t test indicated a significant difference between the pre- (M = 0.30, SD = 0.02) and the post-manipulation (M = 0.53, SD = 0.07) results, t(14) = 3.14, p < .01, Cohen's d = 0.97.
Flanker interference
Mean reaction times and error rates under each perceptual load and compatibility condition are shown in Table 1 for the stress and control groups. To confirm the effect of compatibility on reaction times, we conducted a within-participant t test in the control group. The result indicated that reaction times under the incompatible condition (M = 648.32 ms, SD = 94.37) were longer than those under the compatible condition (M = 634.44 ms, SD = 99.07), t(19) = 2.67, p < .05, Cohen's d = 0.15.
Means and standard deviations of reaction times and error rates for each condition
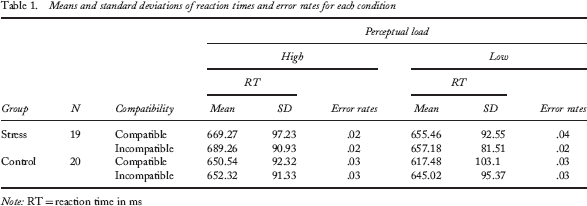
Note: RT = reaction time in ms
We defined the flanker-interference effect as the difference between the reaction times under the incompatible and the compatible conditions (a greater value indicates more interference). Figure 2c shows the interference effects in the stress and the control groups as a function of perceptual load. A 2 (group: stress vs. control) × 2 (perceptual load: low vs. high) mixed ANOVA revealed a significant interaction between the two factors, F(1, 37) = 9.65, p < .01. Paired t tests for the simple main effect indicated that the high perceptual load led to a smaller interference in the control group, t(19) = 2.66, p < .05, and led to a greater interference in the stress group, t(18) = 1.75, p < .10.
Discussion
The results indicated that the stress manipulation was effective in that a significant increase in the STAI scores was observed in the second relative to the first stress measurement in only the stress group. The salivary cortisol levels of the stress group also increased after the TSST. These results indicate that the TSST increased the stress levels of members of the stress group.
Regarding the flanker effect in the control group, we observed a significant interference from the distractor letters when the perceptual load was low, replicating a traditional flanker effect (Eriksen & Eriksen, 1974). This flanker effect disappeared when the load was high. This is consistent with the effect of the perceptual load on the flanker interference predicted by perceptual load theory (Lavie, 2005), suggesting that attentional selectivity can be improved when no spare resources are available to process flanker distractors. More importantly, the flanker interference was eliminated in the stress group when the perceptual load was low. This finding is consistent with the prediction derived from perceptual load theory, given the assumption that perceptual load and stress consume a common attentional resource.
Interestingly, flanker interference was observed in the stress group under the high perceptual load condition. Although this condition is beyond the scope of the load theory, a tentative explanation can be proposed—namely, that an excessive load (i.e., perceptual and stress) disrupts the maintenance of an optimal task set, which is supported by the dorsal frontoparietal network (Corbetta, Patel, & Shulman, 2008). As suggested by recent studies, the ventral network, which is linked with the output of the locus coeruleusnorepinephrine system, sends signals to reconfigure the attentional set maintained by the dorsal network (Aston-Jones & Cohen, 2005; Bouret & Sara, 2005). In the context of the finding that the social stress induced by delivering a speech increased norepinephrine levels (Dimsdale & Moss, 1980), we surmise that maintenance of an optimal task set (i.e., for locating and identifying a target in the central array while ignoring the flanking distractors) became fragile and sometimes failed under stress and a high perceptual load. Under such severely taxing conditions, a flanker interference effect occurred. This contrasts sharply with the results obtained when only stress or only a high perceptual load was present. Under such moderately demanding conditions, the visual system was able to maintain the task set.
Alternatively, the flanker interference observed in the stress group with high perceptual load may be interpreted in the framework of the arousal-biased competition (Mather & Sutherland, 2011), in which high arousal biases processing so that salient stimuli gain priority while less salient stimuli are further suppressed. Because the model predicts that arousal impairs keeping track of multiple items, the interference under the high perceptual load condition of the stress group could be explained in the context of this model. Clearly, the viability of this alternative needs further investigation.
In conclusion, the interaction observed in the present study suggests that perceptual load and stress drain the same attentional resources for nontarget processing, resulting in improved selective attention. We argue that attentional selectivity increases (i.e., minimum interference) when either perceptual load or stress consumes spare attentional resources. In other words, stress enhanced selective attention in that interference was eliminated in the stress group under low perceptual load condition. However, the stress group exhibited a severe interference effect when the perceptual load was high. Although the source of this impairment was not identified in the present study, we surmise that the visual system lost the ability to maintain focus on the task when both the perceptual load and the level of stress were high. Under such conditions, unstable visual selection and the occasional selection of irrelevant items produced interference by the flanker distractors.