
Abstract
Recent research highlights a seemingly flexible and automatic form of cognitive control that is triggered by potent contextual cues, as exemplified by the location-specific proportion congruence effect–reduced compatibility effects in locations associated with a high as compared to low likelihood of conflict. We investigated just how flexible location-specific control is by examining whether novel locations effectively cue control for congruency-unbiased stimuli. In two experiments, biased (mostly compatible or mostly incompatible) training stimuli appeared in distinct locations. During a final block, unbiased (50% compatible) stimuli appeared in novel untrained locations spatially linked to biased locations. The flanker compatibly effect was reduced for unbiased stimuli in novel locations linked to a mostly incompatible compared to a mostly compatible location, indicating transfer. Transfer was observed when stimuli appeared along a linear function (Experiment 1) or in rings of a bullseye (Experiment 2). The novel transfer effects imply that location-specific control is more flexible than previously reported and further counter the complex stimulus–response learning account of location-specific proportion congruence effects. We propose that the representation and retrieval of control settings in untrained locations may depend on environmental support and the presentation of stimuli in novel locations that fall within the same categories of space as trained locations.
Keywords
Context-specific control refers to a seemingly flexible and automatic form of cognitive control that is triggered reactively by contextual cues that predict differing likelihoods of conflict, as evidenced by “location-specific proportion congruence” (LSPC) effects (Crump, Gong, & Milliken, 2006). The LSPC effect refers to the pattern whereby compatibility (i.e., congruency) effects in conflict paradigms are reduced in mostly incompatible (MI) locations in which conflict (incompatible trials) has occurred frequently in the past relative to mostly compatible (MC) locations in which conflict has occurred infrequently.
In the first demonstration of an LSPC effect, Corballis and Gratton (2003) presented flanker stimuli in three locations—centrally and in left and right locations. When the stimuli appeared centrally, each trial was equally likely to be compatible (e.g., HHHHH) as incompatible (e.g., SSHSS). However, one of the outer locations was biased to have more compatible trials, and the other was biased to have more incompatible trials. For example, if the left location was a MC location comprising 75% compatible stimuli and 25% incompatible stimuli, then the right location was a MI location (25% compatible and 75% incompatible). The magnitude of the flanker compatibility effect (i.e., degree of slowing on incompatible relative to compatible trials) was reduced in the MI location compared to the MC location. Such LSPC effects have now been evidenced across a range of cognitive control paradigms and in response to various contextual cues (e.g., flanker task variants: Bugg, 2014; King, Korb, & Egner, 2012; Vietze & Wendt, 2009; Wendt, Kluwe, & Vietze, 2008; Stroop: Crump et al., 2006; Crump, Vaquero, & Milliken, 2008; dual-task performance: Fischer, Gottschalk, & Dreisbach, 2014; priming: Reuss, Desender, Kiesel, & Kunde, 2014).
The central question in the current study regarded the specificity versus generality of LSPC effects—that is, whether the mechanism underlying LSPC effects operates under conditions of transfer. To our knowledge, only one study to date has examined this question (but see Reuss et al., 2014, for transfer of context-specific control to unbiased items based on nonlocation cues). Crump and Milliken (2009) presented a colour word (e.g., “blue”) centrally in white followed by a square that appeared in an upper or lower location on screen; the participants’ task was to indicate the colour of the square. The authors biased the upper and lower locations with MC (e.g., many trials displayed the word “blue” followed by a blue square) or MI stimuli (e.g., many trials displayed the word “blue” followed by a yellow square). Importantly, the authors also presented a novel set of unbiased items (i.e., 50% compatible items equated in stimulus selection history, cf. Awh, Belopolsky, & Theeuwes, 2012) comprising different colour words (“green” or “red”) and coloured squares (in either green or red) in the upper and lower locations. Revealing transfer, the authors found a smaller Stroop effect for unbiased items appearing in the MI location than for unbiased items appearing in the MC location.
Crump and Milliken's (2009) transfer effect demonstrates that location-specific control is flexible in the sense that it appears to be “stimulus blind”. That is, it generalizes to novel, unbiased stimuli when such stimuli appear in the same locations as training stimuli. Importantly, the evidence for transfer to novel stimuli provides strong support for the view that the LSPC effect stems from an abstract cognitive control mechanism, and not a mechanism that is bound to specific features of the biased stimuli (e.g., stimulus–response learning; flanker response priming). Indeed Crump and Milliken interpreted the transfer pattern as supporting an episodic retrieval account of LSPC effects. By this account, an abstract control setting (e.g., devote less attention to flankers) becomes bound to the contextual cue (e.g., lower MI location) such that upon onset of a stimulus in a particular context, the associated setting is retrieved and is applied to the present stimulus.
The primary aim of the present set of experiments was to examine whether LSPC effects generalize to unbiased stimuli when such stimuli appear in novel (untrained) locations. We defined a novel location as one that differs from the locations in which the stimuli were presented during training. If a transfer effect were observed in untrained locations, it would suggest an even more flexible control system than that suggested by Crump and Milliken's (2009) findings, namely one that continues to operate in the face of contextual variation (i.e., one that is relatively “context blind”) and thereby calls into question the label “location-specific control”. This label presupposes that the context (e.g., upper location) in which a stimulus is presented needs to exactly match the context that was trained for location-specific control settings associated with that context to be triggered, an assumption that has not been empirically examined.
The theoretical importance of examining whether location-specific control transfers to untrained locations relates to two distinct issues. First, when thinking about the broader utility of reactive control mechanisms, for instance “in the wild”, one would not expect to consistently encounter imperative stimuli in precisely the same location as in the past. For example, suppose a military radar operator frequently encounters a threatening signal in a particular location near the upper right area of the radar screen. If the location-specific control mechanism subsequently triggered a more focused control setting only when the operator encountered a signal precisely in that same location (versus when a signal appeared in the upper-right area more generally), location-specific control would appear to be quite limited. By contrast, if the mechanism responded to the signal occurring in the upper-right area more generally, this would not only provide support for a relatively flexible mechanism but additionally offer insights into the underlying representations (episodes in which information about space and the likelihood of interference is linked) that support the triggering of varying control settings.
The question is also relevant to a second theoretical issue, which relates to the mechanism supporting LSPC effects. LSPC effects have often been interpreted as evidence for the flexible allocation of cognitive control under conditions in which one cannot prepare for a particular likelihood of conflict prior to stimulus onset (because one cannot predict the location in which a stimulus will appear in advance of its onset, and blocks are composed of 50% compatible trials, thereby contrasting with proactive or list-wide control). More specifically, retrieval of a control setting that minimizes attention to irrelevant features is thought to be triggered by presentation of a stimulus in the MI location whereas a setting incorporating a broader scope of attention is retrieved upon presentation of a stimulus in the MC location (for discussion of the stimulus–attention associations presumed to support location-specific control, see Bugg & Crump, 2012; Crump & Milliken, 2009). Such triggering of attentional settings is often described as relatively automatic, as opposed to intentional, because the allocation of context-specific control does not appear to depend on awareness (e.g., of the context manipulation, Crump et al., 2006, 2008; or the conflict; Reuss et al., 2014).
However, as Crump et al. (2006) noted, an alternative account of LSPC effects is that they reflect complex stimulus–response learning. On this view, the LSPC pattern does not reflect retrieval of an attentional setting in response to the conflict history associated with a given contextual cue (i.e., a particular location); rather, it reflects differential event frequencies across locations (e.g., more presentations of incompatible stimuli in the MI location; Logan, 1988). For instance, participants may be retrieving the response that was most frequently associated with the irrelevant dimension (i.e., the flanking arrows in a flanker task) for a given stimulus in a particular location (cf. Schmidt & Besner, 2008). The examination of transfer has important implications for these contrasting theoretical accounts. Were transfer (i.e., modulation of compatibility effects for congruency unbiased items in untrained locations) to be observed in the present experiment, it would converge with the findings of Crump and Milliken (2009) in providing strong evidence against the complex stimulus–response learning account.
Current approach
In the two experiments reported herein, participants performed a four-choice flanker task by responding to a central target arrow while ignoring flanking arrows. Each experiment comprised three training blocks followed by a single transfer block. During training blocks, stimuli appeared in compatibility-biased locations (e.g., MC, MI). Importantly, these locations were not random locations on screen. Rather, to facilitate learning 1
We do not intend to imply that learning is explicit (see Crump et al., 2006, for evidence to the contrary).
Experiment 1
We investigated for the first time whether LSPC effects transfer to untrained locations that are uniquely linked to trained MC or MI locations (see Wendt et al., 2008, for use of untrained locations that were equally linked to MC and MI locations to examine hemispheric vs. location-specific processing selectivity). During training, stimuli were presented in three locations along an imaginary linear function, with the two endpoints of the function being congruency biased (see Figure 1). During transfer trials, unbiased (50% compatible) stimuli were presented between the compatibility biased endpoints and the centre. If location-specific control transfers to novel locations (i.e., novel x/y coordinates), the flanker compatibility effect should be reduced for unbiased items presented near the interference-heavy (i.e., MI) portion of the function rather than the interference-light (i.e., MC) portion. If location-specific control is, however, location-specific (i.e., bound to particular x/y coordinates), then interference should be equivalent across the two transfer locations.
Method for Experiment 1. Black stimuli indicate training locations (which could be mostly compatible, MC; mostly incompatible, MI; or 50% compatible), and bolded blue stimuli indicate transfer locations (all 50% compatible). The green box identifies a MC (75% compatible) location; the red box identifies a MI (25% compatible) location, and the grey box identifies a 50% compatible location during training. All items actually appeared unbolded in black and one at a time regardless of their location during the experiment. Figure not drawn to scale. To view this figure in colour, please visit the online version of this Journal.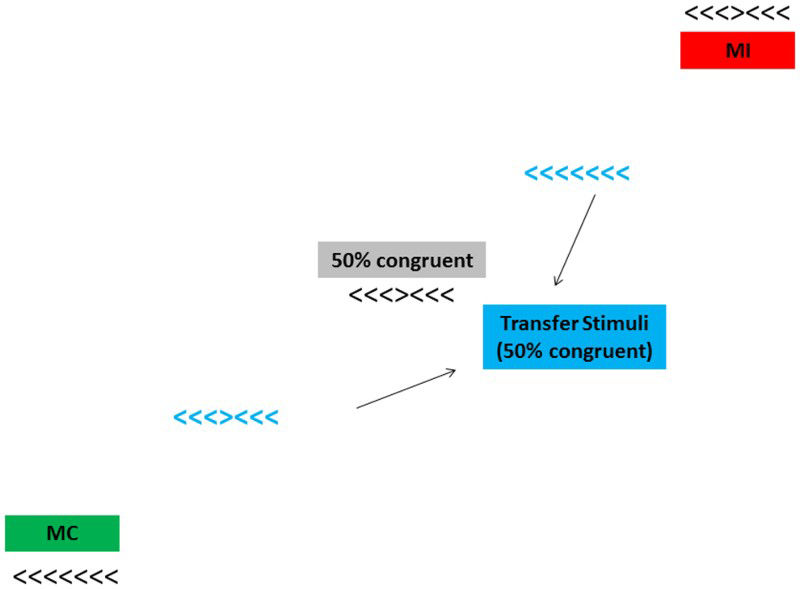
Method
Participants
Twenty-nine undergraduates participated for payment or partial course credit. 2
In Experiment 1 we aimed to collect data from 28 participants (approximating Crump & Milliken's, 2009, Experiment 2, sample size). We then roughly doubled the sample size for Experiment 2 because we expected smaller effects with the increased number of locations to learn during training and/or transfer. Participant sign-ups by the end of the data collection period dictated the final sample size for both experiments.
Stimuli
Participants sat in a well-lit room and viewed the display binocularly from approximately 70 cm. On each trial a single black flanker stimulus measuring approximately 7 cm wide and 1 cm high appeared on a white background. A four-choice task was used such that there were 16 possible flanker stimuli—four compatible (all arrows pointing left, right, up, or down; e.g., <<<<<<<) and 12 incompatible (central arrow pointing in direction opposing flanker arrows; e.g., >>><>>>). Participants responded to the direction of the central arrow with their right index finger on the number pad (“2” for down, “4” for left, “6” for right, “8” for up).
Design and procedure
On each trial, a black fixation cross appeared centrally for 1000 ms prior to the onset of each flanker stimulus, which remained visible until the participant made a valid response (e.g., pressed the 2, 4, 6, or 8 key). Participants completed a practice block of 12 trials, followed by four blocks of test trials—three identical training blocks followed by a single transfer block. During training blocks, stimuli randomly appeared one at a time in one of three locations that created a linear function—one location was at the centre of the screen, and two were equally distant in opposite corners of the screen (see Figure 1). For all participants the central location was used to anchor the function and was not compatibility biased (i.e., 50% of trials were compatible, and 50% were incompatible; we refer to this as the 50% compatible location). However, the two corner positions were compatibility biased—one location for each participant was the MC location (75% compatible trials), and the other was the MI location (25% compatible trials). The direction of the function (positive or negative) and assignment of location (top or bottom) to the MC or MI condition was counterbalanced across participants. For ease of exposition, assume that the participants were exposed to a positively sloping function with MC at the bottom and MI at the top (as in Figure 1). Each training block consisted of 144 trials, with an equal number of trials (48) appearing in each location. In the MC location, each of the four compatible stimuli appeared nine times (36 total), and each of the 12 incompatible stimuli appeared once (12 total). Those frequencies were reversed in the MI location (i.e., each unique stimulus appeared three times for a total of 12 compatible and 36 incompatible trials). In the central, 50% compatible, location, each compatible stimulus appeared six times (24 total), and each incompatible stimulus appeared twice (24 total). Each trial in every block was chosen randomly without replacement from the group of possible trials in that block.
The transfer block consisted of 240 trials, 144 of which were compatibility biased and identical to those presented in the preceding training blocks. In addition, 48 unbiased (50% compatible) stimuli randomly appeared in each of two new locations that were equidistant between the centre location and one of the compatibility-biased end points (locations) along the function (see the bolded blue stimuli in Figure 1). Importantly, the set of stimuli that appeared in these two locations was identical—they only differed in whether they appeared near the MC or MI location along the function (referred to hereafter as near MC and near MI, respectively).
Results
Mean reaction times and error rates for training locations and 50% compatible transfer locations in Experiment 1
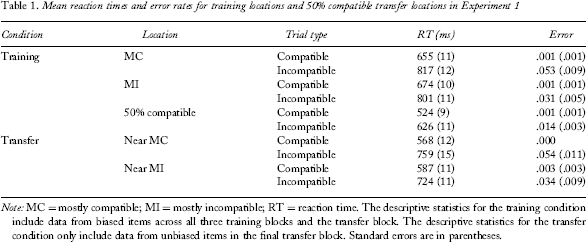
Note: MC = mostly compatible; MI = mostly incompatible; RT = reaction time. The descriptive statistics for the training condition include data from biased items across all three training blocks and the transfer block. The descriptive statistics for the transfer condition only include data from unbiased items in the final transfer block. Standard errors are in parentheses.
LSPC effects
A 2 (proportion compatibility, PC: MC or MI) × 2 (compatibility: compatible or incompatible) within-subjects analysis of variance (ANOVA) was conducted on mean RT for the compatibility-biased items in the trained locations collapsed across the four test blocks (i.e., this analysis included biased items from both training and transfer blocks; see Table 1 for means). 3
Due to a programming error, for a subset of participants (n = 6) one stimulus during each of the three training blocks was presented in the wrong location. That trial is excluded from the analyses presented. Note that exclusion of those participants from the analyses did not change the patterns or statistical reliability of the LSPC and transfer effects.
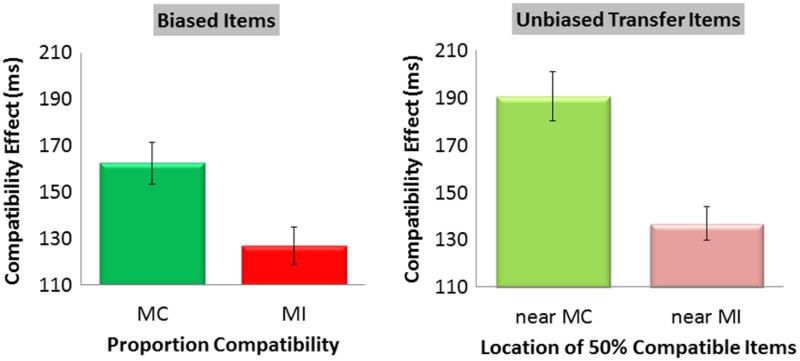
Flanker compatibility effects for biased items (mostly compatible, MC; and mostly incompatible, MI) and unbiased transfer items (near MC and near MI) presented in novel locations from Experiment 1. Error bars represent standard errors of the mean. To view this figure in colour, please visit the online version of this Journal.
Mean error rates are presented in Table 1. For error rate, the same analysis revealed a main effect of congruency with more errors on incompatible than on compatible trials, F(1, 26) = 48.00, MSE = .001, p < .001,
Transfer effects
The transfer analysis focused on the unbiased (50% compatible) trials presented in untrained locations that were near the MC or MI locations (as a reminder these trials occurred only in the final block; see Table 1 for means). The main question was whether the magnitude of the flanker compatibility effect in these two locations differed based on their location along the function. A 2 (location: near MC or near MI) × 2 (compatibility: compatible or incompatible) ANOVA revealed that compatible trials were faster than incompatible trials, F(1, 26) = 495.71, MSE = 1459.78, p < .001,
Mean error rates are presented in Table 1. The error rate data for transfer trials revealed a similar pattern. Participants made fewer errors on compatible than on incompatible trials, F(1, 26) = 30.09, MSE = .002, p < .001,
Discussion
The current experiment demonstrated for the first time that LSPC effects transfer to unbiased items presented in novel, untrained locations. When two sets of identical 50% compatible stimuli appeared in novel locations along the function, there was a larger flanker compatibility effect in both RT and error rate for stimuli that were near the MC location than for stimuli that were near the MI location. As expected, the typical LSPC effect was also observed—participants showed a larger flanker compatibility effect in both RT and error rate in the MC than in the MI location.
The transfer effect is of theoretical importance for two reasons. First, the transfer effect points to a highly flexible control system that can accommodate variation in the contextual cue (location) itself, as might often be encountered outside of laboratory settings. In other words, it appears that location-specific control is not only stimulus blind as prior findings demonstrated (Crump & Milliken, 2009), but may also be somewhat “context blind” (i.e., not location-specific), at least for nearby locations where biased items were never presented. Note that “context blind” here implies that a control setting is not fixed to the exact physical location in which biased stimuli were presented, but can extend to unbiased stimuli in nearby locations that are associated with the biased location (see discussion below regarding how such locations are being coded). Importantly, it does not imply that context is being ignored; location still serves as the cue differentiating the high-conflict from low-conflict area. Second, the transfer pattern challenges the complex stimulus–response learning account of LSPC effects (see also Crump & Milliken, 2009). If such a mechanism were at play, we should not have found a difference in compatibility effects across transfer locations. This is because the same set of unbiased stimuli were presented in each transfer location such that, for example, any learning about the likelihood of a particular response for a given location/irrelevant dimension conjunction should have been equivalent.
One way to account for the primary finding of transfer of location-specific control to novel locations is via a categorical coding system that relates control settings to categories of space (e.g., MI setting is associated with “upper-right” spaces and MC setting with “lower-left” spaces; cf. Kosslyn, 1987) as opposed to distinct x/y coordinates, as in a coordinate coding system. The implication is that novel locations that are categorized in the same way as trained locations may trigger control settings similarly to trained locations. One intriguing possibility is that participants learned there was an increasing likelihood of interference progressing from the lower-left to the upper-right area of the visual field. Then, when unbiased items appeared in novel locations during transfer, participants retrieved the control setting that became associated with the interference-light (lower-left) or interference-heavy (upper–right) portion of the function (space). 4
One potential objection to such an account is that the 50% compatible location presented at fixation did not reveal a compatibility effect that was intermediate to that of the outer MC and MI locations (see Table 1). However, caution is needed when comparing compatibility effects from the central, 50% compatible location to the MC and MI locations. Stimuli in the central, 50% compatible location were presented at fixation, and therefore eye movements were not necessary (whereas the eyes were required to travel identical distances to the MC and MI locations). Indeed, Corballis and Gratton (2003) found a similar pattern of reduced compatibility effects for the 50% compatible location at fixation (compared to the congruency-biased locations to the left or right) and excluded the 50% compatible trials from some of their analyses. Following on their work, we elected to use the central 50% location as an anchor point to facilitate learning of the function and did not expect it to inform our interpretation of the coding system that supported transfer.
Experiment 2
In Experiment 2, we aimed to conceptually replicate the finding that location-specific control transfers to novel, untrained locations and also to examine whether such transfer would be evident when a spatial configuration was adopted that precluded use of discrete categorical codes of a left–right (Corballis & Gratton, 2003) or up–down (e.g., Crump et al., 2006) nature. Stimuli were presented in locations within a three-ring bullseye pattern. During training, multiple locations within each ring were associated with a unique proportion compatibility such that, for example, the probability of interference increased in locations farther from the bullseye. Then, on transfer trials, unbiased stimuli appeared in novel locations within each ring. Because the net left–right and up–down dispersion for stimuli appearing in each ring was equal (e.g., was 0; see Figure 3), a transfer effect would (a) provide converging evidence that novel locations can trigger the allocation of location-specific control, and (b) extend Experiment 1 findings by showing that transfer may also be supported by a different set of categorical codes, namely the coding of “inner” versus “outer” space, and their relationship with the probability of interference. Notably, a transfer effect would additionally provide further evidence against complex stimulus–response learning accounts of the LSPC effect (see Experiment 1; Crump & Milliken, 2009).
Method for Experiment 2. In this experiment the region of space represented by a ring was associated with a compatibility bias (mostly compatible, MC; or mostly incompatible, MI) during training. Black and bolded blue stimuli represent training and transfer items, respectively. All items actually appeared in unbolded black and one at a time regardless of their location during the experiment. Figure not drawn to scale. To view this figure in colour, please visit the online version of this Journal.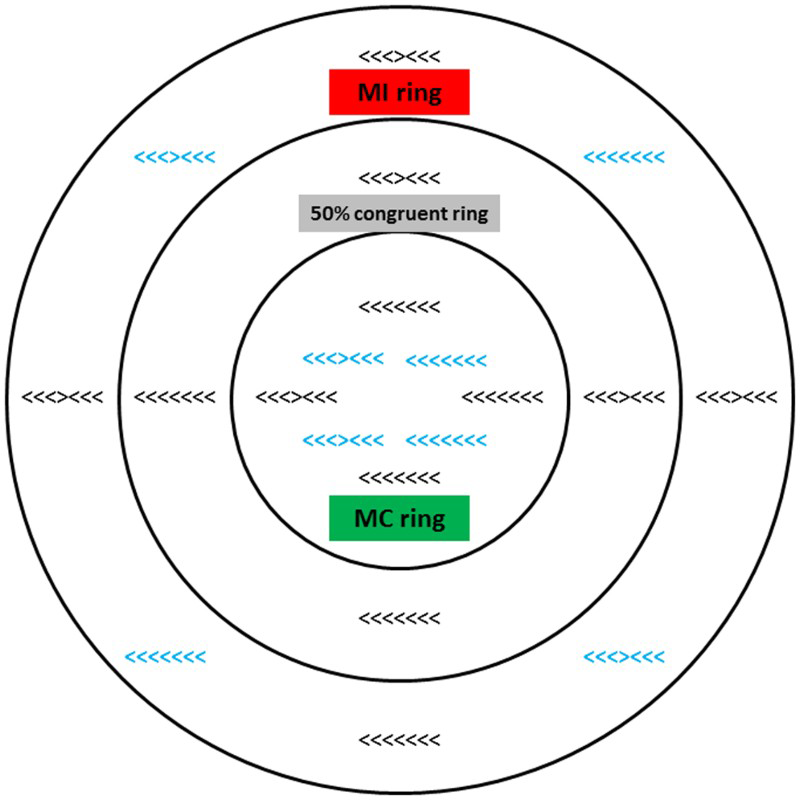
Method
Participants
Fifty undergraduates meeting the same criteria as those in Experiment 1 participated (see footnote 2). Data were excluded for two participants—one for not adhering to task instructions (she or he did not perform a single incompatible trial correctly during practice) and one for falling asleep during testing.
Stimuli, design, and procedure
The method was conceptually similar to that of Experiment 1 with the following exceptions. The flanker stimuli were approximately 3.5 cm wide and 0.5 cm high. Instead of the stimuli appearing against a blank white background, a circular bullseye was present throughout fixation and stimulus presentation during practice and the four test blocks (see Figure 3). During the three training blocks, stimuli could appear in one of 12 locations (four in each ring) presented along the (invisible) vertical and horizontal axes. Each of the three stimulus locations to the left of centre on the horizontal was equally offset from the centre as its counterpart location to the right of centre on the horizontal. Similarly, the three locations above centre on the vertical were equally offset from centre as the three locations below centre on the vertical. Thus, each ring has equivalent left–right and up–down dispersion (the net dispersion across the stimuli in each ring is 0).
In this experiment, instead of a single location being associated with a particular compatibility bias (e.g., MC) during training (as in Experiment 1, and prior research), four distinct locations within each ring were associated with a particular compatibility bias, and the bias differed across the three rings of the bullseye. Stimuli presented in the central ring were 50% compatible for all participants, and the inner and outer rings were either MC or MI (location of MC or MI rings counterbalanced across participants). Each of the training blocks comprised 144 trials, with 12 stimuli appearing at each of the 12 locations. Due to the increased number of possible locations for stimuli to appear (and thus reduced number of stimuli in each location), on each “compatible” or “incompatible” trial throughout the experiment the programme randomly selected amongst each of the four or 12 possible stimuli, respectively. For example, in a given MI location there were nine randomly selected incompatible stimuli and three randomly selected compatible stimuli presented over the course of each block.
During the single 192-trial transfer block, no stimuli were presented in the central ring (i.e., the only 50% compatible items presented were the transfer items). Half of the transfer block trials replicated the compatibility-biased items (MC and MI items) presented in the trained locations in the inner and outer rings during the training blocks. In addition, 96 of the 50% compatible items appeared at eight new locations—four within the inner ring and four within the outer ring (see bolded blue items in Figure 3). Importantly, the compatibility of these items was identical regardless of their location, however half of them were presented in the ring associated with an MC bias during training trials, and half were presented in the ring associated with an MI bias.
Results
Mean reaction times and error rates for training locations and 50% compatible transfer locations in Experiment 2
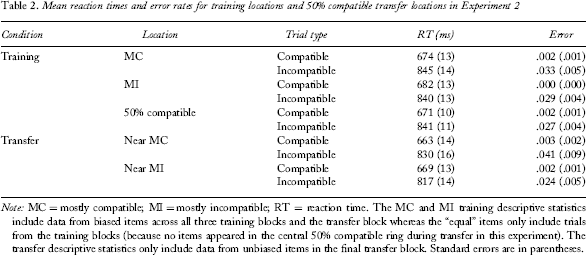
Note: MC = mostly compatible; MI = mostly incompatible; RT = reaction time. The MC and MI training descriptive statistics include data from biased items across all three training blocks and the transfer block whereas the “equal” items only include trials from the training blocks (because no items appeared in the central 50% compatible ring during transfer in this experiment). The transfer descriptive statistics only include data from unbiased items in the final transfer block. Standard errors are in parentheses.
LSPC effects
A 2 (PC: MC or MI) × 2 (compatibility: compatible or incompatible) ANOVA was conducted on RT data from biased trials in the three training blocks and the transfer block (see Table 2 for means). There was a main effect of compatibility (Mcompatible = 678 ms, SE = 10, Mincompatible = 842 ms, SE = 11), F(1, 47) = 1233.84, MSE = 1050.87, p < .001, Flanker compatibility effects for biased items (mostly compatible, MC; and mostly incompatible, MI) and unbiased transfer items (near MC and near MI) presented in novel locations from Experiment 2. Error bars represent standard errors of the mean. To view this figure in colour, please visit the online version of this Journal.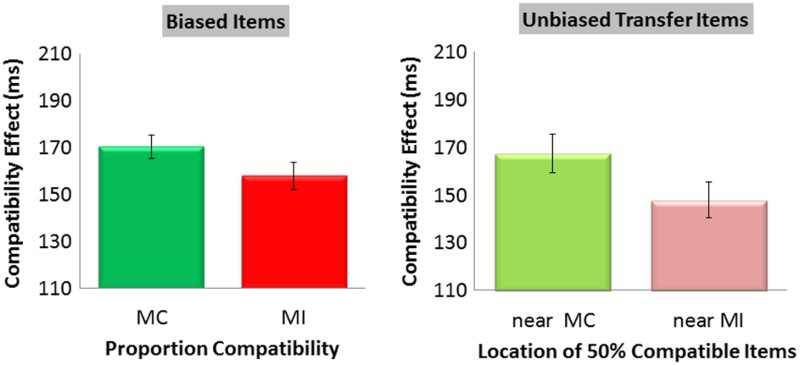
Transfer effects
For consistency with Experiment 1, we continue to refer to 50% compatible transfer items appearing within the MC ring as near MC items and 50% compatible transfer items appearing within the MI ring as near MI items. A 2 (location: near MC or near MI) × 2 (compatibility: compatible or incompatible) ANOVA was conducted on RT for the 50% compatible trials presented in the MC or MI rings during the final transfer block (see Table 2 for means). There was a main effect of compatibility (Mcompatible = 666, SE = 10, Mincompatible = 824, SE = 12), F(1, 47) = 617.83, MSE = 1927.89, p < .001,
The comparable 2 × 2 analysis on error rates revealed a main effect of compatibility, F(1, 47) 30.26, MSE = .001, p < .001,
Discussion
Providing a conceptual replication and theoretical extension of Experiment 1, we observed an LSPC effect for biased items in trained locations and, most importantly, unbiased items (i.e., transfer) in novel locations within compatibility-biased rings of a bullseye pattern. As in Experiment 1, there are two theoretical implications of the transfer pattern. First, recall that due to the spatial configuration of the locations used in the present experiment, the transfer effect cannot be accounted for by a left–right (Corballis & Gratton, 2003) or upper–lower (e.g., Crump et al., 2006) categorical code. Instead, participants may have learned that “inner” space was associated with less interference than “outer” space (or vice versa). This finding, in conjunction with Experiment 1, supports the view that location-specific control settings may not be exclusively location-specific (coordinate based); rather, they may also be coded categorically (e.g., lower left/upper right or inner/outer), and such codes may support transfer of control to novel locations that fall within the relevant categories of space. As such, these codes offer a mechanism via which location-specific control might operate in the wild, in the face of contextual variation. Returning to the example of the radar operator, this may suggest that if the upper right area of the radar screen corresponded to a particular country, then the occurrence of a signal in a novel location within (but not outside of) the borders of that country would trigger location-specific control. Second, the transfer effect further challenges the complex stimulus–response learning account of LSPC effects.
General Discussion
The current study presented novel evidence that LSPC effects transfer to unbiased (50% compatible) items presented in novel, untrained locations. In Experiment 1, the magnitude of compatibility effects differed for unbiased items in novel locations based on learned interference for nearby locations along a linear function. In Experiment 2, the magnitude of compatibility effects differed for unbiased items that were presented in novel locations in rings of a bullseye, based on the learned interference associated with a given ring.
The current demonstrations of transfer suggest that location-specific cognitive control is notably more flexible than previously documented. The findings support the view that location-specific control may not (always) be location-specific and raise the possibility that a categorical (e.g., “inner” versus “outer”) coding system may support transfer of location-specific control. However, before elaborating on our preferred interpretation of the transfer patterns as demonstrating a flexible, reactive control system that can accommodate cue variation (i.e., different locations), we discuss potential alternative explanations for the observed results.
Alternative accounts of transfer
The complex stimulus–response learning account attributes the LSPC effect to the learning of responses that are associated with location cue + distractor compounds (Crump et al., 2006; cf. Schmidt & Besner, 2008). While such an account might explain the LSPC effect on training trials, it cannot explain the transfer effect. The stimuli presented in each of the novel transfer locations were 50% compatible, yet they yielded different flanker compatibility effects based on their location. If participants were simply learning to predict responses in particular locations based on distractor information, then equivalent compatibility effects should have been observed across the two locations. Crump and Milliken's (2009) findings similarly ruled out the complex stimulus–response learning account.
A similar account that merits consideration is a practice-based location–flanker–response priming account, which posits that irrelevant flankers prime responses differentially across MC and MI locations. 5
We thank an anonymous reviewer for suggesting this possibility.
Finally, the control account of the present effects may also be contrasted with another account that has been proposed to explain proportion compatibility effects. The temporal learning account attributes such effects to participants learning the rhythm of responding in a mostly easy (i.e., MC) compared to a mostly difficult (i.e., MI) condition (Schmidt, 2013a, 2013b). While the account was initially developed to explain list-wide proportion compatibility effects, Schmidt, Lemercier, and De Houwer (2014) recently showed that temporal learning expectancies can be learned for specific contexts in a nonconflict task (i.e., in the absence of a congruency manipulation), allowing individuals to adjust the rhythm of responding on a location-by-location basis. The authors suggested that temporal learning might therefore explain LSPC effects in conflict tasks, including patterns of transfer such as those observed by Crump and Milliken (2009). One way to evaluate whether this account explains the present findings is to examine the data for evidence that participants learned different rhythms of responding in the MC and MI locations. In Experiment 1 and Experiment 2, there was no main effect of PC on the training trials. While this appears inconsistent with the temporal learning account, as noted by Schmidt et al. (2014), the main effect of PC reflects a contrast of marginal means and does not take into account the relative frequency of compatible and incompatible trials in each location (for each PC level). A comparison of average RTs in each location regardless of trial type supports that participants were overall faster in the MC location (MExp1 = 693 ms, MExp2 = 715 ms) than in the MI location (MExp1 = 768, MExp2 = 800 ms), suggesting that temporal learning may have contributed to the LSPC training effects. Turning to the novel finding of transfer effects, because all transfer locations comprised 50% compatible (i.e., equally difficult) stimuli, one can rely on the main effects of location to inform the account. In neither experiment was there a main effect of location, suggesting similar temporal expectancies across the transfer locations. In other words, compatibility effects should have been equivalent across transfer locations were temporal learning solely responsible for the effect. One interesting possibility is that location-based temporal learning transferred from the training trials to the nearby locations representing the transfer trials; however, the absence of a main effect of location for the transfer locations is difficult to reconcile with this view.
The current account
In contrast to the accounts discussed in the preceding section, we suggest that the observed LSPC effect in unbiased novel locations is the result of a reactive (cf. e.g., Braver, Gray, & Burgess, 2007) allocation of the appropriate abstract control setting based on the cue of stimulus location. 6
A similar category of effects, known as item-specific proportion congruence (ISPC) effects, have been suggested to reflect “automatic control” (Jacoby, Lindsay, & Hessels, 2003). Location-specific control may similarly be automatic in the sense that retrieval of the appropriate settings for a given location does not appear intentional (strategic) given the rapidity with which it occurs and participants’ lack awareness of the PC of particular locations (e.g., Crump et al., 2006).
The flexibility of location-specific control
The current findings imply that location-specific control is notably more flexible than would be expected if location-specific control were exclusively supported by a coordinate system (i.e., a unique setting for each x/y stimulus coordinate; e.g., Jager & Postma, 2003). Location-specific control may be stimulus blind (Crump & Milliken, 2009) and relatively context blind (i.e., able to cope with variation of cue location—x/y coordinates), as long as the novel location is perceived to fall within the same spatial category as biased stimuli (e.g., upper-left; inner), a finding that is most easily accounted for by a categorical coding system. The diversity of categorical codes evidenced in the present experiments (e.g., a combination of vertical and horizontal, Experiment 1, and inner versus outer, Experiment 2) highlights that individuals readily pick up on a variety of statistical regularities in the environment and represent them in a form that enables the context-sensitive allocation of cognitive control, including to untrained locations. The present findings thereby converge with prior findings in highlighting the putative importance of category-level representations in facilitating transfer of reactive control mechanisms (see Bugg, Jacoby, & Chanani, 2011, for transfer of item-specific control to new exemplars from trained animal categories, and Cañadas, Rodríguez-Bailón, Milliken, & Lupiáñez, 2013, for transfer of context-specific control to novel exemplars of male and female facial categories). We note, however, two potential limitations. First, the categorization process necessary for transfer of control from one location to another in the present research was relatively simple. More specifically, control contexts could be categorized by a rule in which the contexts differed on only one feature (e.g., in Experiment 2 inner location vs. outer location). This is the easiest type of category to learn (e.g., Kruschke, 1992), so future research is necessary to determine whether location-specific control could be learned and transferred when more complicated categorization is necessary. Second, the current experiments examined only transfer of location-specific control and not other forms of context-specific control. If transfer is facilitated by categorization processes, one might expect transfer of colour-specific or shape-specific control to the extent that novel cues are members of trained (biased) categories (e.g., if training entails shape cues that are squares in the MC condition and circles in the MI condition, presentation of another four-sided shape such as a rectangle may trigger retrieval of the attentional setting associated with the MC condition whereas presentation of a triangle may not). This remains to be tested.
Furthermore, although our findings converge on the conclusion that location-specific control may operate at a categorical level, we are not arguing that location-specific control operates only at a categorical level (cf. Crump & Milliken, 2009, where transfer occurred for stimuli presented in the same location, which may be accounted for by a coordinate coding system). Categorical coding may occur selectively in paradigms such as the current that provide environmental support by presenting trained locations within invisible (Experiment 1) or visible (Experiment 2) spatial configurations that bias adoption of categorical codes (and perhaps also by presenting transfer locations within the reference frame created by such configurations; cf. Kunde, Kiesel, & Hoffman, 2003). Prior research, albeit with a very different type of task, predicts factors such as the size of the attended region of space (Michimata, Saneyoshi, Okubo, & Laeng, 2011), and whether there is overlap in receptive fields (Jacobs & Kosslyn, 1994) affect whether categorical coding is evidenced. Further research is therefore needed to identify those conditions that bias adoption towards categorical or coordinate codes in LSPC paradigms, as such factors would seem to impact heavily on whether location-specific control transfers to novel, untrained locations.
Finally, although we have argued that participants learned and transferred control based on categorical codes, we acknowledge that the transfer locations were also closer in spatial proximity to their “categorically linked” biased locations than the other biased location. Thus theoretically, spatial proximity could be fully driving the transfer effects. However, recall that in Experiment 1 the unbiased transfer locations were exactly equidistant from the 50% compatible location and one of the biased locations. Therefore, if spatial proximity was the mechanism, it seems just as likely that participants would have adopted the 50% control setting, and thus the observed transfer effects would be unlikely. Of course an alternative possibility is that the control setting adopted in the transfer locations represents the “average” of the two closest control settings (e.g., for transfer location in upper right, it reflects the average of the control setting associated with the middle location and the extreme upper right location; cf. Wendt et al., 2008). While this does not undermine the evidence for transfer, it does support the spatial proximity account. A stronger counterargument to this account stems from Experiment 2. The transfer locations in the outer (MI) ring of the bullseye were almost as close to the biased items in the inner (MC) ring as they were to the biased items in the outer (MI) ring (see Figure 3). 7
More specifically, a transfer item in the outer ring was an average 29.0 E-prime units from the training items within the outer ring and 31.2 units from the training locations in the inner ring; a difference unlikely to be noticed by participants (for comparison the inner transfer items were an average of 7.0 units from the inner ring biased items).
Conclusion
The novel findings presented herein demonstrate that LSPC effects transfer to novel, untrained locations. Novel locations that were nearby trained locations stimulated retrieval of the control settings associated with the trained locations, thereby affecting the magnitude of compatibility effects for unbiased items presented in the novel locations. These findings suggest that location-specific control settings may not be location-specific, in a coordinate sense. Rather, individuals may represent the relationship between locations and the probability of interference in the format of a categorical coding system that enables transfer of location-specific control to locations that fall within trained categories of space. The findings suggest that context-specific control seems to be not only stimulus blind (Crump & Milliken, 2009), but also (within certain parameters) context blind (i.e., can accommodate cue variation) and is therefore a highly flexible system of control.