
Abstract
When multiple cues are presented in compound and trained to predict an outcome, the cues may compete for association with an outcome. However, if both cues are necessary for solution of the discrimination, then competition might be expected to interfere with the solution of the discrimination. We consider how unequal stimulus salience influences learning in configural discriminations, where no individual stimulus predicts the outcome. We compared two hypotheses: (1) salience modulation minimises the initial imbalance in salience and (2) unequal stimulus salience will impair acquisition of configural discriminations. We assessed the effect of varying stimulus salience in a biconditional discrimination (AX+, AY−, BX−, BY+). Across two experiments, we found stronger discrimination when stimuli had matched, rather than mismatched, salience, supporting our second hypothesis. We discuss the implications of this finding for Mackintosh’s model of selective attention, modified elemental models and configural models of learning.
Keywords
Introduction
Many theories of learning assume that the rate at which an association between stimuli is acquired depends in part on the associability of the stimuli (e.g., α). Stimulus associability is influenced by absolute properties, such as the physical intensity of stimuli, as well as by relative properties. Mackintosh (1975) proposed that the associability of a stimulus (α) will increase when it is a better predictor of an outcome than other presented stimuli. Thus, stimulus associability is dynamic, varying over training as a function of its relative predictiveness.
If two stimuli are equally predictive and presented in a compound, overshadowing may occur. The associative strength acquired by either stimulus will be less than that acquired through training in isolation (Pavlov, 1927). When stimulus associability is unequal, the stronger stimulus generally overshadows the weaker stimulus, reducing acquisition of associative strength for the weaker stimulus (Mackintosh, 1976). While overshadowing may facilitate learning by encouraging processing of informative stimuli, many discrimination tasks require learning about the co-occurrence of stimuli. For instance, in a biconditional discrimination (AX+, BX−, AY−, BY+), each stimulus is paired with the presence and absence of an outcome, such that the solution of the discrimination requires learning about both stimuli (Saavedra, 1975). Animals, including humans, can learn such discriminations (e.g., Byrom & Murphy, 2016; Gonzalez, Welch, & Colwill, 2013; Harris & Livesey, 2008; Lober & Lachnit, 2002).
Studies of configural learning, interested in how stimuli are configured to support discriminations such as the biconditional, tend to consider learning under conditions in which each stimulus is equally salient. However, beyond the controlled conditions of the laboratory, stimuli rarely have equal salience. Theories offer different predictions for the effect of unequal salience. Theories that propose learning results in salience modulation, such as that by Mackintosh (1975), suggest that unequal stimulus salience may have a minimal effect on a biconditional discrimination. Alternatively, configural and modified elemental models might be expected to predict unequal salience to decrease discrimination substantively. We summarise the theoretical background supporting these alternate hypotheses before outlining the experiments conducted.
Elemental models, such as Mackintosh’s (1975) attentional theory, did not consider configural discriminations. As no individual stimulus in a biconditional discrimination predicts the outcome, the individual stimuli cannot be used in isolation to solve the discrimination. To solve configural discriminations, elemental models require that elements have different properties in the presence of different stimuli (AX+, AY−). One assumption is that peripheral units are generated by the co-occurrence of stimuli, and that these added or inhibited units contribute to learning (McLaren & Mackintosh, 2002; Rescorla, 1972; Wagner & Brandon, 2001; Wagner & Rescorla, 1972). During the biconditional discrimination, when two stimuli, for example, A and X, are presented together, new elements are assumed to be activated by their co-occurrence which were latent in the component stimuli. Furthermore, some models include inhibited elements that are present in the component stimuli, but not activated by the compound.
An alternate approach from Pearce (1987) proposed that the co-occurrences form unique representations, different from the sum of the elements. Stimuli are assumed to excite corresponding input units and recruit configural units that are active for the configuration of stimuli (Pearce, 1994). As learning is supported by configural units, rather than the individual stimuli, this model readily solves the biconditional discrimination.
Both Pearce’s (1987) configural model and McLaren and Mackintosh’s (2002) modified elemental model predict that unequal salience will interfere with biconditional discrimination. Increasing the salience of stimuli A and B and decreasing the salience of stimuli X and Y will increase generalisation between AX and AY and decrease generalisation between AX and BX. Thus, unequal salience may be expected to advantage some facets of the discrimination and disadvantage others. However, as X and Y are crucial to the solution of the biconditional discrimination, a shift in learning towards A and B will reduce the acquisition of the biconditional discrimination.
In contrast, to static salience differences, Mackintosh (1975) suggested that salience is labile but that initial differences in stimulus salience may have only a minimal impact on a configural discrimination. Mackintosh’s (1975) theory of selective attention suggests that changes in associability will be influenced by trial order and will fluctuate over training. For instance, on the first AX+ trial, A and X become minimally predictive of the outcome. However, if the next trial is AY−, stimulus A is a relatively weak predictor of the absence of outcome, and hence, the associability of A should decrease, while the associability of Y should increase. Similarly, on the subsequent BX− trial, the associability of X should decrease. Because each individual stimulus is paired with the presence and absence of an outcome, associability should fluctuate across training. While some stimuli may start with a high salience, all individual stimuli are poor predictors of outcome and as such, the initial high salience may be expected to fall. Introducing salience modulation based on predictiveness to either configural models (George & Pearce, 2012) or McLaren and Mackintosh’s (2000, 2002) model would be predicted to limit the effect of initial salience difference on a configural discrimination.
However, there are other ways that the individual value of cues might impact on discrimination learning. The biconditional discrimination can also be thought of as a set of four cues with equivalent reinforcement histories, since each cue is paired with each outcome 50% of the time. Similarly, in a transitive inference problem A+B−, B+C−, C+D− and D+E−, each of the middle cues in the sequence (B, C and D) is associated with an equivalent reinforcement history. Animals and humans can solve these discriminations but will also show a transitivity effect by choosing B > D (e.g., Couvillon & Bitterman, 1992; Von Fersen, Wynne, Delius, & Staddon, 1991). Circular versions in which the end points (A and E) also have equivalent histories are also more complex to learn (A+B−, B+C−, C+D−, D+E− and E+A−), and both animals (e.g., Davis, 1992; Wynne, 1997) and humans find these discriminations more difficult (Henley, Horsfall, & De Soto, 1969). However, what this task demonstrates is that local contrasts, as well as global reinforcement history, contribute to solution of discrimination. The relative value of the cues might be expected to support learning of the biconditional discrimination if each pair of cues involved one cue that was more salient than the other, as with the transitive inference discrimination. So, for instance, if the validity of the four cues was such that X+ > A− > Y+ > B− > X+, a form of circular transitive problem, then there is a solution to each pair of cues.
Given the role that differing salience might play in solving a complex discrimination, in the two experiments described here, we sought to assess whether unequal stimulus salience affects the acquisition of a biconditional discrimination. Experiment 1 assessed the effect of a difference in the physical salience of stimuli, while Experiment 2 extended this study training predictive validity prior to a biconditional discrimination.
Experiment 1
This experiment varied the physical salience of stimuli to assess the effect of stimulus salience on acquisition of a biconditional discrimination. In the experimental condition, participants completed a biconditional discrimination, in which the relevant stimuli were either of matched or of mismatched salience. All participants were trained with the same stimulus configurations, as shown in Table 1. The configurations contained two discrimination-relevant dimensions and one irrelevant dimension. The three dimensions were used so that both groups learnt about the same stimuli. Two stimulus dimensions had a high physical salience: a line drawing (A/B) and a background colour (X/Y). One stimulus dimension had a low physical salience: the angle of lines in the background colour (S/R).
Discrimination designs for Experiments 1 and 2.
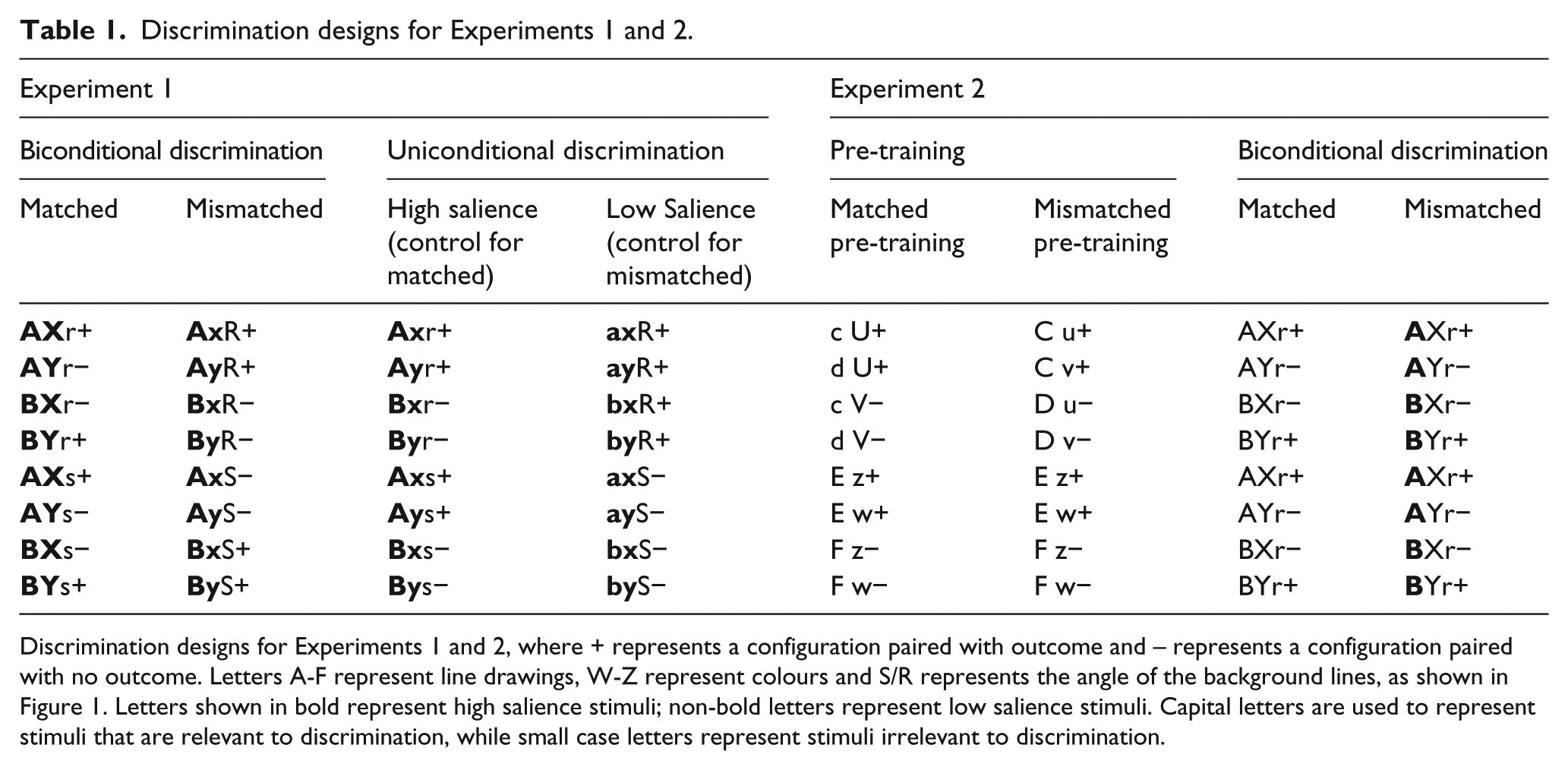
Discrimination designs for Experiments 1 and 2, where + represents a configuration paired with outcome and – represents a configuration paired with no outcome. Letters A-F represent line drawings, W-Z represent colours and S/R represents the angle of the background lines, as shown in Figure 1. Letters shown in bold represent high salience stimuli; non-bold letters represent low salience stimuli. Capital letters are used to represent stimuli that are relevant to discrimination, while small case letters represent stimuli irrelevant to discrimination.
The inference that these stimulus dimensions differ in salience was assessed through a categorisation task. Participants (n = 14, including nine women, with an average age of 33 years) were presented with these stimuli and asked to divide the stimuli into two groups. While nine participants categorised the stimuli based on colour and five participants categorised the stimuli based on shape, none of the participants categorised the stimuli based on the angle of the lines in the background colour.
If differences in the physical salience of stimuli presented in compound influences configural learning, acquisition of the biconditional discrimination would be weaker in the mismatched salience condition than in the matched salience condition. However, poor biconditional discrimination in the mismatched condition may also reflect a general inability to learn about the low salience stimulus dimension (S/R). To rule out this possibility, a control group completed a uniconditional discrimination with exactly the same stimulus configurations as used in the biconditional discrimination. In this control discrimination, only one stimulus dimensions reliably predicted the paired outcome, and participants did not have to learn about stimulus configurations. Within the control group, half of the participants learnt a discrimination in which a high salience stimulus dimension (A/B) predicted the outcome, and half learnt a discrimination in which the low salience stimulus dimension (S/R) predicted the outcome. If overshadowing influences configural learning, beyond simply impairing participants’ ability to learn about low salience stimuli, the difference between the matched and mismatched groups should be greater in the biconditional than uniconditional discrimination.
Method
Participants
Participants were university students (n = 48), who were reimbursed £6 for their time. The average age was 22.27 years (standard deviation [SD] = 2.59). Seventeen participants were male. Average digit span (a measure of working memory capacity; Lezak, 1995) was 7.54 (SD = 0.59) digits. Demographics across the participant groups are shown in Table 2.
Demographics for Experiment 1.
Design
A factorial design was employed with discrimination task (uniconditional and biconditional), stimulus salience (matched and mismatched) and stimulus counterbalancing (learning with stimulus sets A, B or C) as between-subjects factors. The outcome type (+, −) and the trial block (1-12) were incorporated as within-subjects factors.
Materials
All stimuli were programmed and presented using Visual Basic software. The stimulus configurations included three dimensions: a line drawing (A/B), a background colour (X/Y) and the angle of lines in the background (S/R). To ensure that effects were not driven by a specific stimulus, partial counterbalancing was accomplished by using three different sets of stimuli: A, B and C. The pairs of line drawings, and example of stimulus configurations, are shown in Figure 1. Colour pairs were yellow versus red, pink versus peach and blue versus green. The angle of the lines were 26° versus 103°, 51° versus 129° and 77° versus 154°.
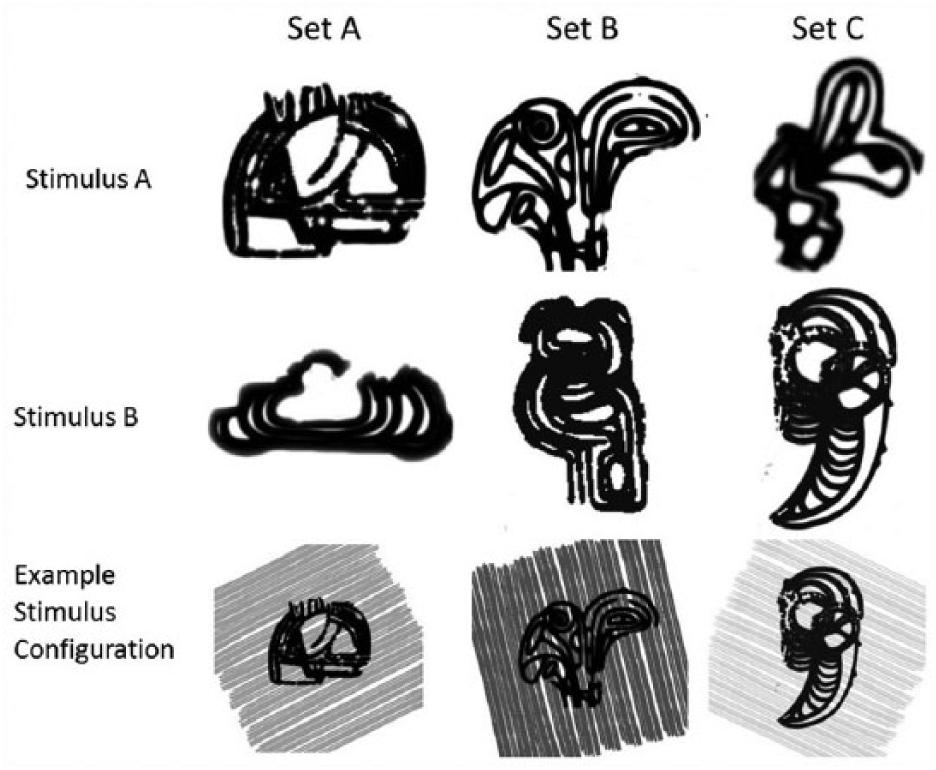
The six different black line drawings used for one dimension of the stimuli. Within each stimulus set, two different black shapes were used, allowing this dimension to vary between two points.
Procedure
The task involved a cover story developed and described elsewhere (Byrom & Murphy, 2016). Stimuli were presented as works of art in a fictitious art gallery. The participants’ task was to predict the outcome (stimulus popularity) associated with each stimulus configuration. The design of the discrimination tasks is shown in Table 1. Stimulus configurations were displayed 12 times, giving a total of 96 trials. On each trial a stimulus configuration was presented, and participants were asked to predict the outcome using a Likert scale on the keyboard, ranging from 1 (unpopular), through 5 (unsure), to 9 (highly popular). Following the prediction, feedback was presented showing the paired outcome (the stimulus’ popularity). This remained on the screen until the participant pressed a button for the next trial. Feedback was represented visually. Where a stimulus was popular (+), participants were shown a room more than 60% full of people. Where a stimulus was unpopular (−), participants were shown a room that was less than 40% full. Feedback was probabilistic, so that on popular trials (+) the room was shown as 70%, 80% or 90% full, and on unpopular trials (−) the room was shown as 10%, 20% or 30% full. Probabilistic feedback was used to maintain the participant’s engagement with the task. For each of the 12 trial blocks, the participant’s average outcome rating was calculated for stimuli paired with a positive outcome and separately for stimuli paired with a negative outcome.
Results
The relative salience of the stimuli had an impact on acquisition of the biconditional discrimination but not the uniconditional discrimination, as shown in Figure 2. Successful discrimination in the uniconditional task indicates that participants can use the low salience stimuli, when presented alongside higher salience stimuli, to resolve a discrimination. However, when the same stimuli needed to be configured with more salient stimuli acquisition of the discrimination was impaired. These observations were supported by analysis of variance (ANOVA) with the factors of outcome (+ vs −), trial block (1-12), stimulus counterbalancing sets (A, B, C), salience condition (matched and mismatched) and discrimination (uniconditional and biconditional).
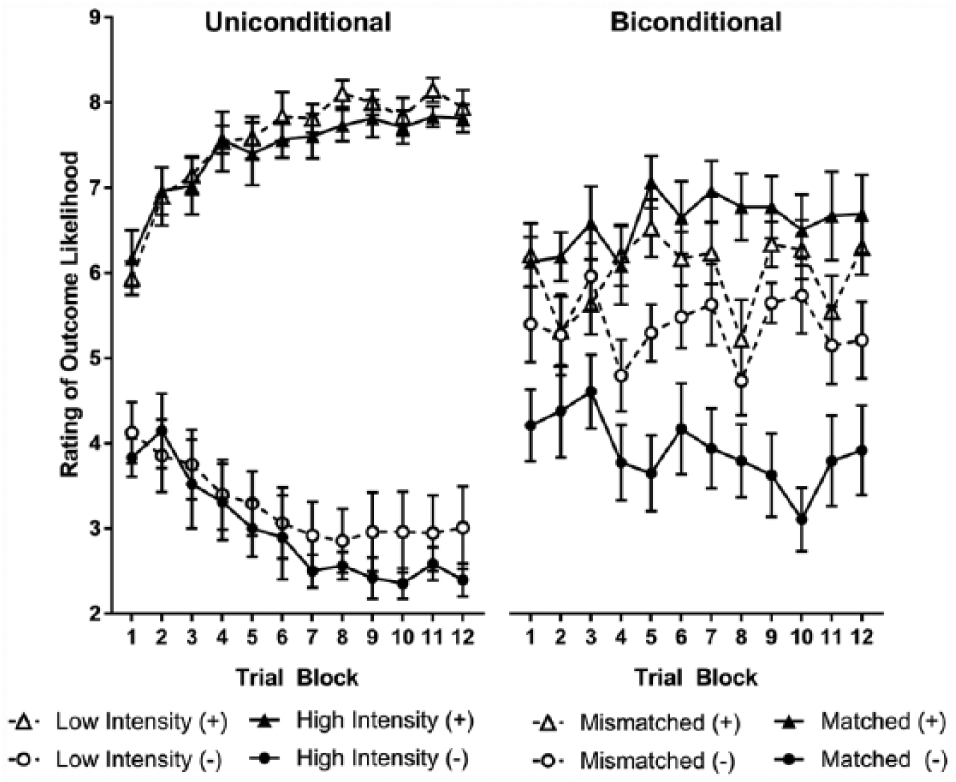
Discrimination learning in Experiment 1, showing change in outcome likelihood ratings over training in the uniconditional discrimination (left-hand panel) and biconditional discrimination (right-hand panel). Error bars show standard error of the mean.
Indicating overall learning, there was a significant main effect of outcome, with participants giving higher ratings to stimulus configurations paired with a positive outcome than a negative outcome, F(1, 44) = 174.97, p < 0.001,
For participants completing the biconditional discrimination, there was a main effect of outcome, F(1, 22) = 16.47, p < 0.001, qualified by a significant interaction with salience condition, F(1, 22) = 6.34, p = 0.020. Considering all training trials, participants in the matched salience condition discriminated between stimuli based on the paired outcome, F(1, 11) = 15.69, p = 0.002,
For participants completing the uniconditional discrimination, there was a main effect of outcome, F(1, 22) = 429.05, p < 0.001,
Discussion
The relative salience of the stimuli within a configuration influenced the acquisition of a biconditional discrimination. Where salience was mismatched, such that one stimulus dimension in the configuration had a high salience while another had a low salience, participants failed to acquire the biconditional discrimination. This suggests that despite the irrelevance of individual stimuli to the discrimination, the salience of these stimuli influenced discrimination. The pattern of results supports configural and modified elemental models and does not support the suggestion of salience modulation within a configural discrimination. We suggest that failure to solve the biconditional discrimination in the matched condition occurs because the low salience stimuli (R/S) are overshadowed by the high salience stimuli.
Of course, participants may have failed to notice the low salience stimuli all together, and thus, have been unable to learn about these stimuli. This would provide a simpler explanation for our results. However, this explanation can be ruled out by the control condition, where participants learnt about identical stimulus configurations to those presented in the biconditional discrimination. In the control condition, only one stimulus dimension predicted the outcome, making these uniconditional discriminations. Participants acquired the uniconditional discrimination readily whether the outcome was predicted by a high or low salience stimulus, demonstrating that participants can recognise and learn about the low salience stimuli (R/S) when presented in configurations.
In combination, these results suggest that mismatched stimulus salience creates a unique problem for learning the biconditional discrimination. In the mismatched salience condition, the outcome in the biconditional discrimination was predicted by the combination of a stimulus dimension that participants used to categorise stimuli and a dimension that participants did not use to categorise stimuli. While participants did not use the low salience stimulus dimension to categorise stimuli, they had no problem learning that this dimension was a reliable predictor of a paired outcome, acquiring the uniconditional discrimination readily. Participants were significantly impaired when they had to learn about the combination of high and low salience stimulus dimensions.
An additional component of the design may have contributed to the poor discrimination in the mismatched condition. In the matched high salience condition, the irrelevant stimulus dimension had a low salience. In the mismatched condition, the irrelevant stimulus dimension had a high salience. Therefore, both the consistency of the salience matching and the type of irrelevant dimension were possible sources of the difference between conditions. In Experiment 2, we conducted a systematic replication of the main effect of interest, while removing any effect due to the irrelevant dimension by reverting to a standard biconditional with only two dimensions.
Experiment 2
This experiment extended the findings of Experiment 1, explicitly manipulating the associability of stimuli through relative validity training, prior to a biconditional discrimination (Baker, Murphy, & Mehta, 2003; Le Pelley & McLaren, 2003). Prior to learning a biconditional discrimination (AX+, BX−, AY−, BY+), participants completed either matched training, where both stimulus dimensions were trained to be predictors of an outcome, or mismatched training, where only one stimulus dimension was trained to be a reliable predictor of an outcome, as shown in Table 1. We refer to stimulus dimensions because different stimuli, from the same dimension, were used in pre-training and in the biconditional discrimination to ensure that the effects of training on associability were not confounded by acquired associative strength. For instance, the colours red and yellow may have been used in associability training, while pink and peach were used in the biconditional discrimination.
This procedure follows the pattern of intra-dimensional and extra-dimensional shifts (e.g., George & Pearce, 1999; Mackintosh & Little, 1969). In the matched condition, a stimulus dimension predictive of the outcome in training became relevant to the biconditional discrimination. In the mismatched condition, a stimulus dimension that was not predictive of the outcome in training became relevant to the biconditional discrimination. To establish two stimulus dimensions as equally relevant to the discrimination, without providing prior training in a configural discrimination, the matched training included, for both dimensions, stimuli that were and were not relevant to the outcome, as shown in Table 1. In this way, while the matched condition in Experiment 1 contained two high salience stimulus dimensions, the matched condition, here, contained two relatively neutral stimulus dimensions.
Participants
Participants were university students (n = 18), who were reimbursed £6 for their time. The average age was 22.29 years (SD = 2.56). The average digit span was 7.53 (SD = .58) digits. Key demographics across the participant groups are shown in Table 3.
Demographics for Experiment 2.
Design
A factorial design was employed with stimulus pre-training (matched vs. mismatched) and stimulus counterbalancing (sets A, B or C) as a between-subjects factor. The paired outcome in the discrimination task (+ vs −) and the trial block (1-12) were incorporated as within-subjects factors.
Materials
Stimuli were as described in Experiment 1; however, the third stimulus dimension, the angle of lines in the background (the low salience stimulus used in Experiment 1), was held constant across all stimulus configurations.
Procedure
The procedure was as described in Experiment 1, with the inclusion of a pre-training phase. There was a pause between pre-training and test phases, in which a new computer programme was loaded and participants were presented again with task instructions. The design for pre-training, consisting of 12 trial blocks and a total of 96 trials, is shown in Table 1. Pre-training used the same cover -story and, hence, comparable procedure as that described in Experiment 1. As described in Experiment 1, partial counterbalancing was used with three groups of participants learning about three different sets of stimuli, as shown in Table 4.
Stimuli groups used in Experiment 2.

Stimulus sets are shown in Figure 1.
Results
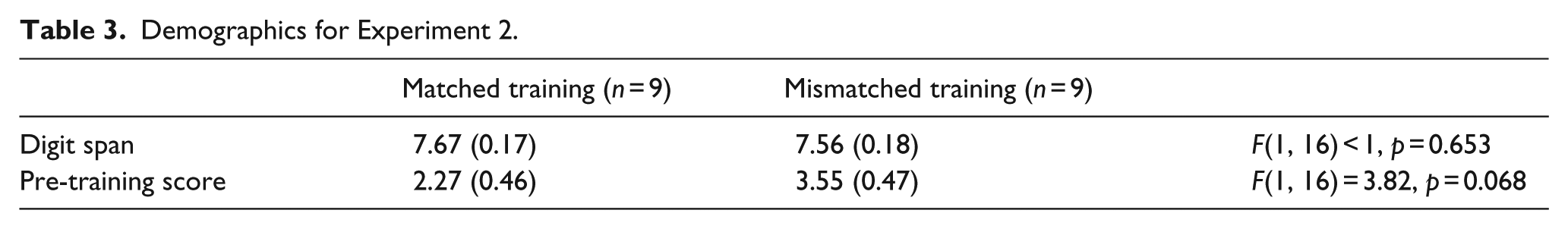
We outline the results of the pre-training phase and then address the acquisition of the biconditional discrimination. Pre-training used a uniconditional discrimination in both the matched and mismatched conditions, as shown in Table 1. As shown in the left-hand panel of Figure 3, participants successfully learnt the pre-training discrimination. Discrimination scores (rating for stimuli paired with + less rating for stimuli paired with −) increased over training trials, from 0.10 (standard error of the mean [SEM] = 0.30) in the first block of training to 4.33 (SEM = 0.48) in the final block of training, F(11, 176) = 22.80, p < 0.001,
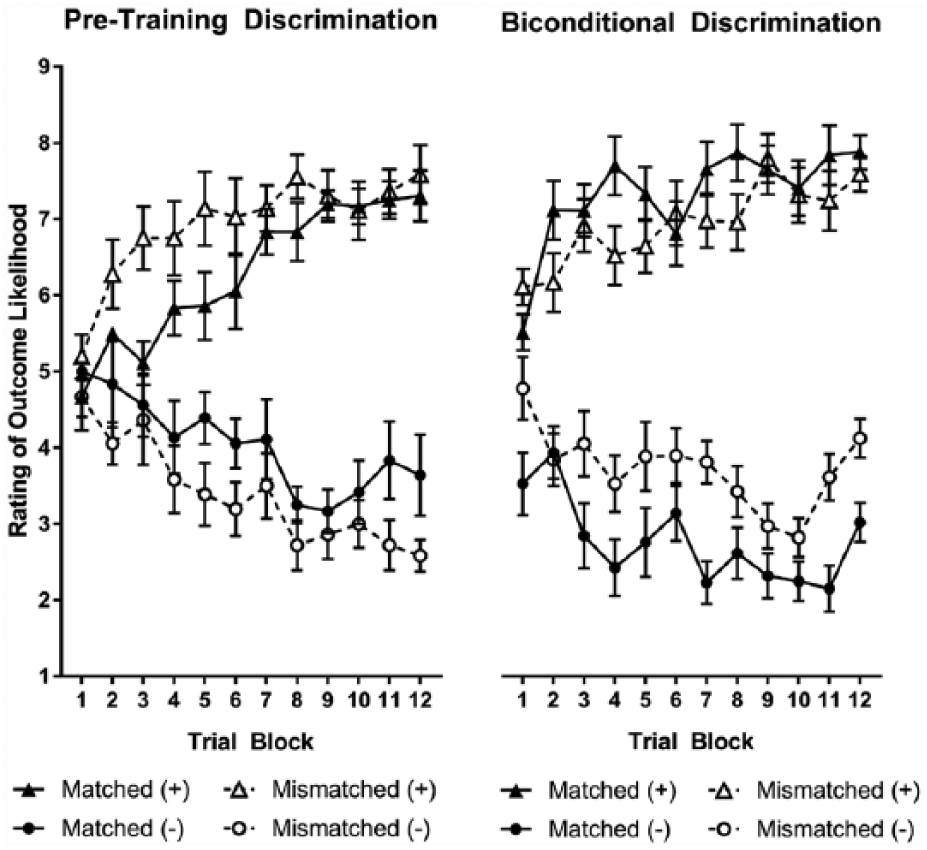
Discrimination learning in Experiment 2, showing change in outcome likelihood ratings over training. Error bars show standard error of the mean.
As shown in the right-hand panel of Figure 3, participants in the mismatched associability condition showed slightly weaker biconditional discrimination than those in the matched associability condition. However, this observation was not supported by the analysis of variance, with the factors of outcome (+, −), trial block (1-12), stimulus-counterbalancing set (A, B, C) and trained salience condition (matched and mismatched); the interaction between outcome, trial block and trained salience condition was not significant, F(11, 176) < 1.
However, acquisition of the pre-training discrimination might be expected to influence the extent to which stimulus associability is altered by training. We repeated the ANOVA, with pre-training performance (average discrimination score) held as a covariate throughout analysis. Assumptions for analysis of covariance (ANCOVA) were met. There was independence of the covariate and treatment effect; the training discrimination (i.e., covariate) did not differ significantly between the two treatment conditions: F(1, 16) = 3.82, p = 0.068. Regression slopes were homogeneous; performance on the training discrimination and biconditional discrimination were correlated, r = 0.688, p < 0.001, and this correlation did not vary across treatment groups; performance in the training discrimination and biconditional discrimination were correlated for both the matched, r = 0.738, p = 0.023, and mismatched, r = 0.846, p = 0.004 conditions.
In the ANCOVA, discrimination varied between associability conditions, with a significant interaction between associability condition and outcome, F(1, 11) = 5.42, p = 0.040. As reported in Experiment 1, there was no interaction with counterbalancing stimuli groups, F(2, 11) < 1, p = 0.62. Analysed separately, with the covariate of pre-training performance, discrimination approached significance in the matched associability condition, F(1, 7) = 4.76, p = 0.065,
The relative associability of stimuli within a configuration, established through prior relevance training, had a small influence on the acquisition of a biconditional discrimination. This interaction was dependent on the strength of prior training. Where the associability of paired stimuli was mismatched, with one stimulus dimension previously trained to be relevant, while the other was trained to be irrelevant, acquisition of the biconditional discrimination was weaker than it was in a condition in which stimuli in the configuration were treated equally in pre-training.
To achieve the salience imbalance in this experiment, we trained only one dimension to be relevant to the discrimination in the mismatched training condition. While we think this unlikely, this may be considered to have effectively trained participants to use an elemental strategy in discrimination learning, encouraging a reliance on only one dimension to solve the discrimination. However, this does not provide a full account of the difference between the two conditions. While participants were trained to use both dimensions to solve discriminations in the matched training condition, this remained a uniconditional discrimination, in which the outcome on any trial was predicted by only one dimension of the configuration. This would provide the same training as the mismatched condition, again, encouraging a reliance on only one dimension to solve the discrimination. In training with pigeons, George and Pearce (1999) adopted an alternative approach, using a biconditional discrimination with a redundant dimension in pre-training, prior to switching to a new biconditional discrimination involving stimuli from either (1) both the previously relevant dimensions or (2) one previously relevant and one previously irrelevant dimensions.
General discussion
Across two experiments, overshadowing effects influenced discrimination when neither stimulus was predictive of the outcome and both stimuli were needed to solve the discrimination. Where stimuli had an unequal salience, biconditional discrimination was either retarded or completely abolished. In Experiment 1, participants failed to learn a biconditional discrimination when the stimuli were mismatched in physical intensity. In Experiment 2, with prior training held as a covariate, discrimination was weaker when stimuli had a mismatched relative validity history. We suggest that when stimuli of mismatched salience were presented in compound, the high salience stimulus is over-represented, impairing acquisition.
Although there has been considerable exploration of the factors that influence configural learning, including stimulus properties and training (for review, see Melchers, Shanks, & Lachnit, 2008) and individual differences (Byrom & Murphy, 2014, 2016; Haddon et al., 2014), the influence of relative stimulus salience has received less attention. This is curious, as beyond the laboratory, the stimulus configurations routinely encountered do not consist of equally associable components. For instance, contexts are often used in configuration with discrete stimuli to predict outcomes. The findings reported here suggest that the ability to acquire configural discriminations may be substantially influenced by the relative stimulus salience.
George and Pearce (1999) published a comparable experiment with pigeons, and our findings provide a good indication that their results can be extended to human participants. Pigeons were trained to solve a biconditional discrimination with a redundant stimulus, in much the same design as described here in Experiment 1. Following initial training, one of two new biconditional discriminations was introduced. For one group (matched salience), the new discrimination contained stimuli from the two previously relevant stimulus dimensions. For the other group (mismatched salience), the new discrimination contained stimuli from one of the previously relevant dimensions and from one of the previously redundant dimensions. Pigeons in the matched salience condition acquired the discrimination faster than the mismatched salience condition, indicating both that the amount of attention paid to a stimulus dimension is determined by its relevance to the discrimination and that this allocation of attention influences configural discrimination.
We set out to compare two possible hypotheses for the effect that unequal stimulus salience would have upon biconditional discrimination. First, following configural and modified elemental models, we might expect that unequal salience would retard learning, as the salience of one component of the compound might reduce activation of unique configural or peripheral units and increase generalisation between the compounds to be discriminated. Alternatively, following Mackintosh (1975), as no individual stimulus was a good predictor of the outcome, salience modulation may be expected to reduce the salience of individual stimuli, minimising the effect of any imbalance in salience. Salience modulation has been incorporated into Pearce’s configural model (George & Pearce, 2012) and could play a role in modified elemental models (Livesey & McLaren, 2011).
The findings reported here do not provide strong support for the second hypothesis. The Mackintosh (1975) model predicts that associability is updated based on the predictive validity of individual stimuli. Contrary to this, differences in stimulus salience continued to influence acquisition across biconditional discrimination. However, Mackintosh’s (1975) model does allow for the pre-training, conducted in Experiment 2, to influence the rate of learning observed in the subsequent configural discrimination. The effect of mismatched salience was substantially weaker in Experiment 2 and dependent on the strength of prior training. This may reflect a weakness in our predictive validity training or less than perfect generalisation of trained associability from the training stimuli. The difference in the size of effect between Experiments 1 and 2 might also reflect the contribution of the redundant stimulus in Experiment 1. Furthermore, the manipulations in Experiments 1 and 2 focused on different facets of stimulus salience. In Experiment 1, physical salience was manipulated. In Experiment 2, prior training was used to influence stimulus associability. These facets of salience, physical salience and learnt associability may be different. It is conceivable that changes in learnt associability, as described in models of salience modulation, occur more readily than changes in the influence of physical salience.
The findings reported here are accommodated by our first hypothesis that unequal stimulus salience will retard configural discrimination. Both configural and modified elemental models allow stimulus salience to influence similarity between configurations or proportions of overlapping elements (e.g., McLaren & Mackintosh, 2002; Pearce, 1987). For instance, Pearce (1987) proposed that generalisation between configurations occurs on the basis of similarity, where the proportion of elements representing a stimulus may be determined by the intensity (or salience) of the stimulus. Specifically, similarity is calculated as set out in Equation (1):

where Pcom is the perceived intensity of the stimuli that are common between presented configurations, and
This approach to generalisation allows us to predict the results of the biconditional discriminations reported in Experiments 1 and 2, as shown in Figure 4. For instance, if we assume that the intensity of A is substantially greater than X or Y (say a ratio of 9-1) the similarity between AX and AY will be high (i.e., 0.81), while the similarity between AX and BX will be low (i.e., 0.01). As shown in the left-hand panel of Figure 3, the biconditional discrimination is acquired more slowly for the mismatched salience condition, reflecting the results of Experiment 2. Experiment 1 included a redundant stimulus, expected to influence the similarity between compounds, and thus influence generalisation, much like a context cue (Vallee-Tourangeau, Murphy, & Baker, 1998). In particular, the redundant stimulus introduces generalisation between compounds that ostensibly share no cues (i.e., AX and BY), as these configurations share a redundant stimulus (i.e., RAX and RBY). Where the redundant stimulus has high salience, this generalisation is more substantive than a condition where the salience of the redundant stimulus is low. The influence of this on a biconditional discrimination is shown in the right-hand panel of Figure 3; inclusion of a redundant stimulus reduces biconditional discrimination in both matched and mismatched salience conditions; however, the effect is more pronounced in the mismatched salience condition, effectively eliminating discrimination. This simulation is similar to the findings of Experiment 1.
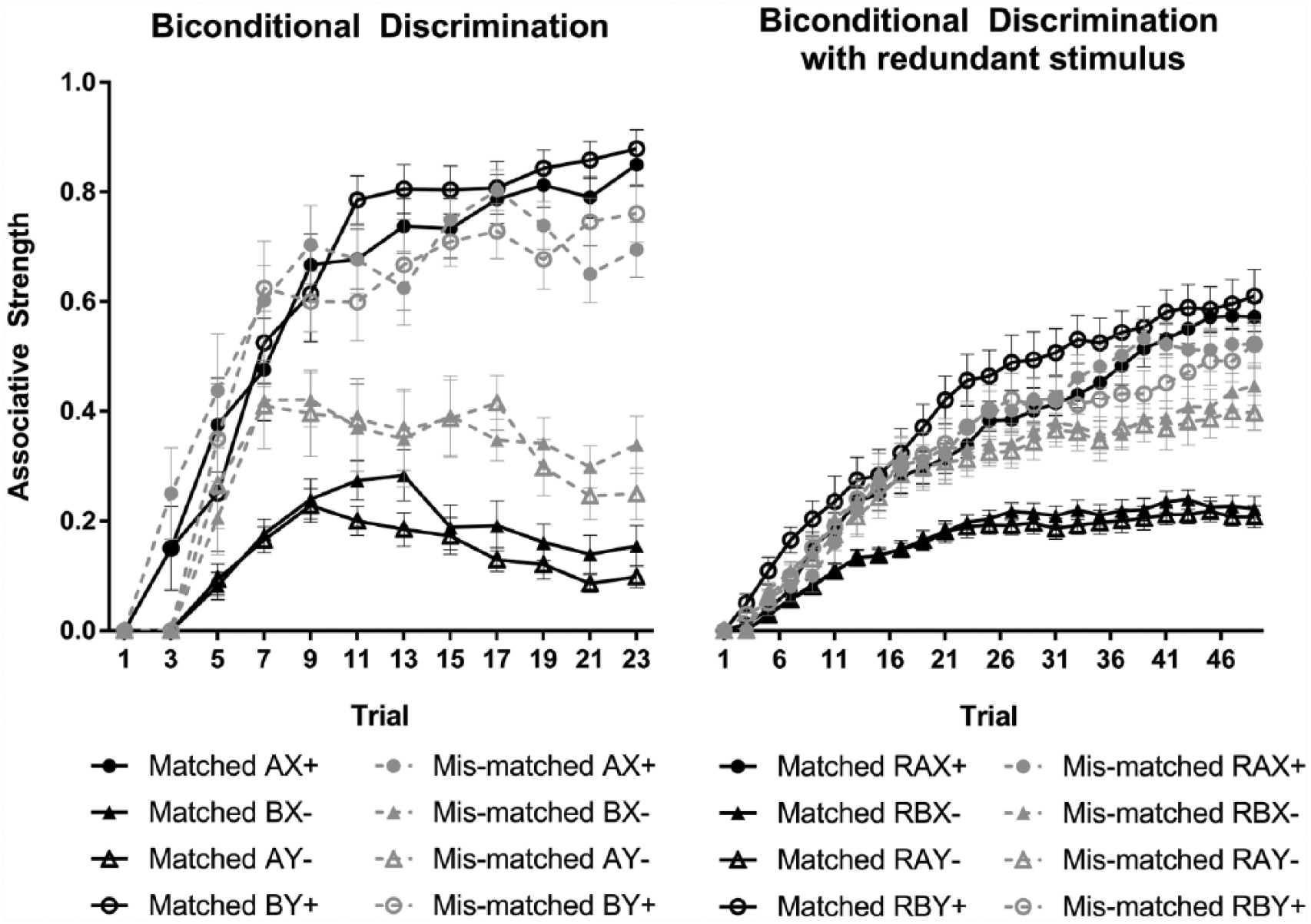
Simulated learning, using Pearce’s (1987) configural model, for biconditional discrimination with and without a redundant stimulus.
It is important to note that while these stimulations replicate the acquisition of the biconditional discriminations, Pearce’s (1987, 1994) configural model does not predict learned changes in associability (though see also George & Pearce, 2012). Furthermore, the parameters used to predict biconditional discrimination cannot replicate the uniconditional discrimination. By making the intensity of A and B nine times greater than X and Y, learning with X and Y is reduced dramatically, predicting a substantive impairment in the acquisition of the uniconditional discrimination. This suggests that the effect of unequal salience on biconditional discrimination may not be explained solely in terms of differences in the similarity of configurations and generalisation.
One suggestion is that the associative account may inadequately explain learning with the uniconditional discrimination. Instead, it is possible that participants engage in a process of hypothesis testing (Yu, Smith, Klein, & Shiffrin, 2007). This proposal suggests that perhaps hypothesis testing is the first strategy adopted by participants and that once found lacking a simpler associative mechanism takes over. This hypothesis does not predict the gradual learning that was found but is worth exploring in future experimental work.
Do the modified elemental models provide a sound account of the influence of unequal salience? Elemental models depend on replaced elements, also referred to as peripheral units, to solve configural discriminations (McLaren & Mackintosh, 2002; Wagner & Brandon, 2001). Alteration in the recruitment of core and peripheral units would influence configural discrimination; as the activation of core units relative to peripheral units increases, the model will behave more elementally and discrimination will be retarded (see also Wagner, 2003). In terms of salience modulation, recruitment of core and peripheral units might change over training; as core units, activated by elements unique to the individual stimuli are redundant in a configural discrimination, the salience of core units may decrease relative to the salience of peripheral units (see Delamater, Sosa, & Katz, 1999, for a comparable approach).
The two experiments presented here suggest that an imbalance in stimulus salience may influence configural discrimination. Confirmation of the effect of mismatched salience needs independent quantification of the differences in perceived salience of stimuli. While we have considered this from two very different perspectives (the physical intensity of salience and trained associability), there are many ways in which stimulus salience can be manipulated, and further study will be necessary to conclude that stimulus salience does, in general, influence configural discrimination.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship and/or publication of this article.