
Abstract
Objectives:
The goal of this research is to design a novel model using protein folding to study Therapeutic Touch, a noncontact form of energy manipulation healing. Presented is a feasibility study suggesting that the denaturation path of ribonuclease A may be a useful model to study the energy exchange underlying therapeutic touch.
Design:
The folding of ribonuclease A serves as a controlled energy-requiring system in which energy manipulation can be measured by the degree of folding achieved. A kinetic assay and fluorescence spectroscopy are used to assess the enzyme-folding state.
Results:
The data suggest that the kinetic assay is a useful means of assessing the degree of refolding, and specifically, the enzyme function. However, fluorescence spectroscopy was not shown to be an effective measurement of enzyme structure for the purposes of this work.
Conclusions:
More research is needed to assess the underlying mechanism of therapeutic touch to complement the existing studies. An enzyme-folding model may provide a useful means of studying the energy exchange in therapeutic touch.
Introduction
During a TT session, the healer assesses, modulates, redistributes and, when necessary, supplements the patient's energy in order to restore his or her energy balance. The practitioner assesses the patient's energy field by moving his or her hands a few inches directly above the surface of the patient's body in sweeping motions for several seconds in order to determine the patient's energy distribution. In the case of the human system, this energy can be either balanced, as in healthy people, or imbalanced, as in the unhealthy person. The practitioner then interacts with the imbalanced areas, and pushes and redirects energy flow where it is needed through physical hand movements. 1
TT was initially introduced as an intervention for nurses and is now widely used in a variety of settings. The efficacy of TT has been studied in a variety of clinical settings, particularly in palliative care. 3 –5 One clinical study assessed the usefulness of treating cardiovascular patients with TT versus contact therapy. To test the placebo effect associated with this method, sham TT (where TT movements were mimicked but no efforts to center and focus a healing intent were made) was given to a control group. Anxiety levels of patients were surveyed before and after treatments. The results suggested a significant decrease in post-stress state anxiety scores in subjects tested with TT relative to those tested with sham TT. On average, TT was found to reduce stress levels by 17% in the hospital setting compared to sham TT therapy. 6 A similar clinical study, in which surveys and psychologic tests were used for assessment, TT was found to be significantly more effective in relieving the anxiety of institutionalized elderly patients than massage therapy. However, TT was not found to be significantly more effective than sham TT performed by a TT practitioner who blocked the “intent to heal.” 7
A review of literature between 1997 and 2007 on the effects of TT on pain found a statistically significant effect attributed to TT in four out of five of the studies reviewed. TT was found to be useful in alleviating musculoskeletal, arthritic, and burn pain. However, not all of these studies controlled for the placebo effect. Those that did control for the placebo effect did not necessarily show a significant difference between TT and sham TT. 8 An earlier review found evidence to support TT's ability to reduce anxiety but noted that there was much variation and incongruity in the methods of analysis, administration, and conceptual basis for experiments seeking to study TT. 2
In an effort to better research the theoretical basis of TT, Krieger designed a series of studies to demonstrate TT's effect on circulating hemoglobin as a measure of system oxygenation. Using both in vivo and in vitro models, she found that TT was able to significantly increase hemoglobin levels in both systems 9,10 ; however, Krieger's work has been questioned with regard to randomization, double-blind precautions, and sufficient controls. Her work was improved upon by more recent research that assessed TT's effect on hemoglobin and hematocrit levels in young healthy volunteers presenting with hemoglobin levels of <12 g/dL. The results of this work found that both TT and sham TT were associated with a significant increase in hemoglobin levels above controls. 11 These studies are important because the focus is to directly test the target group of this method, namely, sick patients. Although these studies demonstrate an effect of TT, it is unclear whether they are due to a placebo effect or the proposed mechanism of TT.
Two (2) recent in vitro studies suggest that TT is capable of significantly increasing cell proliferation in culture of a variety of human cell types and increasing DNA synthesis, differentiation, and mineralization in human osteoblasts, while decreasing differentiation and mineralization in osteosarcoma cells. 12,13 This work adds significantly to the body of research characterizing TT's efficacy by negating placebo effects, though it does not fundamentally address the proposed mechanism underlying TT. Similarly, several studies on related energy manipulation techniques such as Reiki and qigong have been done that suggest an effect but give little clarity regarding the mechanisms underlying these energy manipulation modalities. 14 –17
TT is widely utilized by health care providers in palliative care, hospice, and inpatient settings to alleviate a variety of symptoms. Currently, it is being researched in the clinical setting; however, many of the studies supporting its use fail to adhere to rigorous research standards and to adequately control for placebo effects. Although less prevalent, promising in vitro research that intrinsically negates placebo effects suggests the potential value that TT holds. However, the current published work, both clinical and in vitro, has focused on characterizing TT's effects rather than defining and testing its underlying mechanism. In order to fundamentally understand TT and validate its use in the clinical setting, its mechanism of action must be better characterized. This work presents a novel model to investigate the mechanism of action of TT directly by borrowing from biochemistry the concepts of enzyme folding.
Conceptual Model
In order to work within the TT construct and design a model that is satisfactory to both TT practitioners and the scientific community, the assumptions of TT must be considered.
1
People must be considered dynamic, open systems, and energy can be exchanged between the individual and the environment. Thus, the energy associated with a person does not stop at the skin but rather extends beyond that person. The universe is composed of energy at its most basic level, and humans are also embodiments of that energy. The energy field exerted by humans is not only perceivable but is organized and patterned in a constant and symmetric way in a healthy individual. TT cannot improve upon a healthy system but rather can only put into balance an otherwise imbalanced system. Finally, TT offers a form of communication, or intervention, into this characteristic energy in order to restore balance.
This research was designed to work within assumptions 1–4, and to study assumption 5. The model system must exhibit some quantifiable energy change or state to directly test TT's mechanism. The proposed enzyme-folding model is sensitive to changes in energy state and thus is useful in studying the underlying mechanism of TT's effects. It should be stated that science views energy as existing in organized forms (i.e., heat, light, electricity, all of which obey physical laws). Although TT literature evokes quantum theory to characterize the energy transfer, some schools of thought suggest that the energy associated with TT does not obey the same physical laws as conventional forms of energy. 1 By utilizing an enzyme model that responds to changes in physically defined energy states, this work assumes that TT affects conventional energy that obeys physical laws of science.
An enzyme, a protein that performs a function, is a string of amino acids—each having unique chemical and energetic properties. The enzyme's ability to perform its function is dependent upon structure, or the orientation of these amino acids with regard to each other in space. This orientation depends on various properties (charges, hydrophobicity, or hydrophilicity, acidity or basicity), which in turn depend on the energetics of the environment, or the solution in which they exist. When an enzyme folds, some of its amino acids become more closely associated with one another. The process of folding, or reorienting the amino acids in space, represents an energetic path on which the enzyme's overall energy progressively decreases as lower-energy conformations (folding patterns) are achieved. A tenet of biochemical theory explicated by Anfinsen et al. states that the lowest energy, and therefore most stable, conformation of an enzyme is its active state, the state in which it can perform its function, otherwise known as its native state. 18 The folding, or renaturing, of an enzyme, and the energy of the system in which it exists are interconnected. Therefore, the enzyme's ability to perform its function and the overall energy of the enzyme itself are interconnected.
Current theories of enzyme folding suggest that the process of protein folding resembles a funnel and follows a path of decreasing energy as each progressive state represents a slightly more stable form (Fig. 1). However, during the folding process, intermediate structures are formed, which create “bumps” in the funnel. These bumps represent not only a partially folded conformation of the enzyme, but also local energy minima in which the enzyme can be temporarily stuck. These conformations have a lower energy state than the fully unfolded (denatured) enzyme, but do not represent the lowest energy state (i.e., the native state), and are in this way less energetically stable. Therefore, relative to the fully denatured enzyme, the intermediate species must overcome a lower energy barrier before assuming the native state. 19 –21 It has been well documented that forms of energy such as heat, pressure, or changes in pH can be used to overcome this energy barrier. However, to the best of our knowledge, TT's ability to overcome this energy barrier has not been tested.
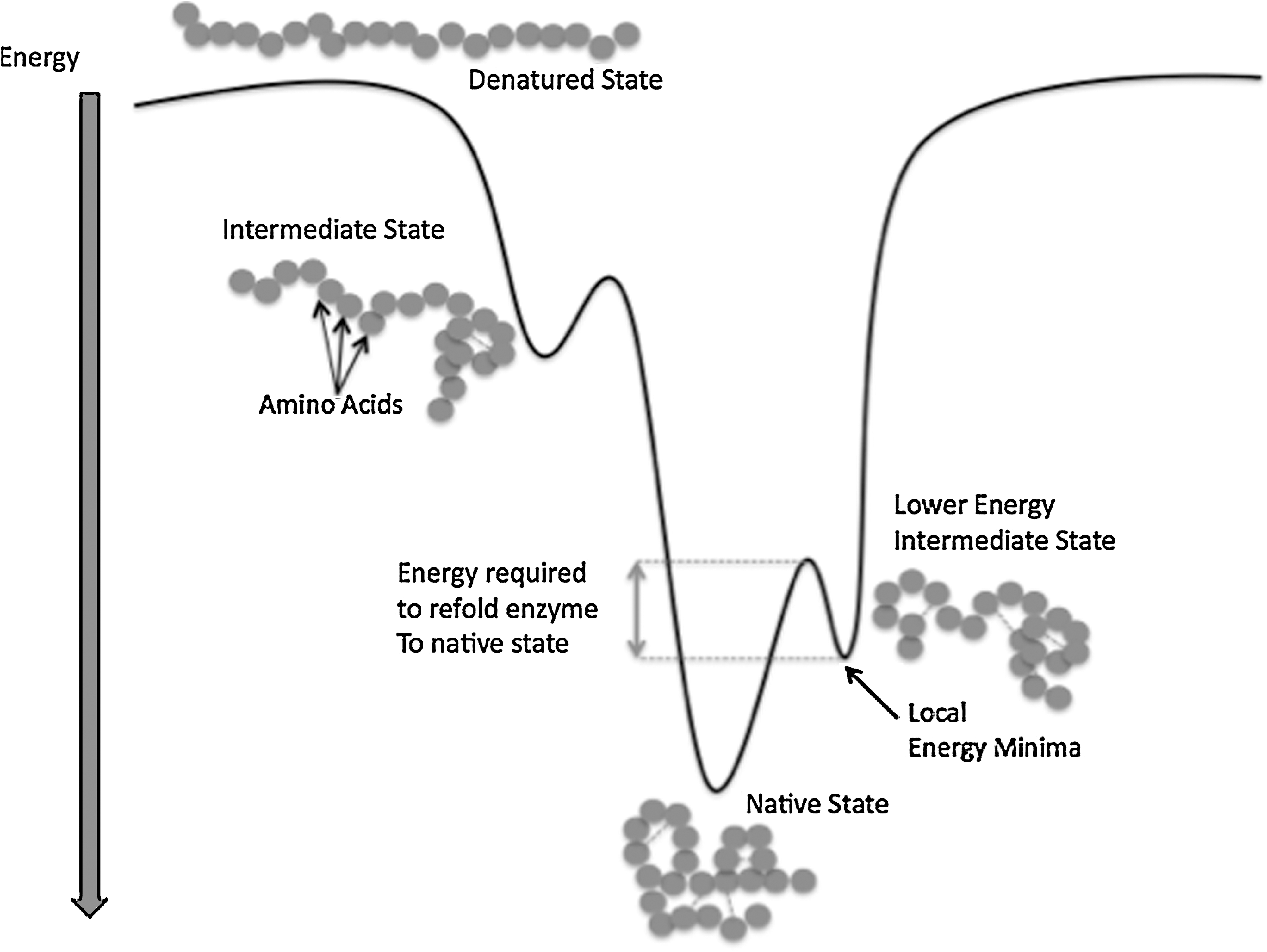
Enzyme folding state. This diagram shows the energetic path of enzyme folding. The local energy minima are shown as “bumps” in the path in which the intermediate species form. The dashed lines represent the energy barrier that must be overcome in order for an intermediate species to be folded into the native species.
Historically, the pre-eminent research on folding states was performed on ribonuclease A, a small, single-chain enzyme. Its primary function is to degrade RNA. It is predominately fixed in its native folding pattern by four disulfide bonds and autorenatures to a measurable extent. 18 In particular, the folding of ribonuclease A is sensitive to environmental energetic changes, such as heating and cooling, and is known to have a multidimensional folding/unfolding energetic path, which is suggestive that its folding path includes multiple intermediate species. 22
We propose that the intermediate state represents a system of energetic instability, or imbalance relative to the native state lowest energy conformation. The presence of intermediate states in ribonuclease A, therefore, makes it an appropriate model system in which to study TT's ability to effect energetic change in a system of relative energetic instability.
In order to determine whether this model is capable of testing the mechanism underlying TT, studies were performed following the denaturation and renaturation methods described here.
Materials and Methods
Denaturation
Denaturation is the process by which proteins and enzymes unfold, or lose their three-dimensional structure. Ribonuclease A is denatured using β-mercaptoethanol (BME) and concentrated urea solutions and renatured through progressive dialysis steps, which remove first BME and then urea. The removal of BME in the presence of urea is sufficient to cause the enzyme to remain in the partially or incorrectly folded intermediate state. These steps are described below and illustrated in Figure 2.
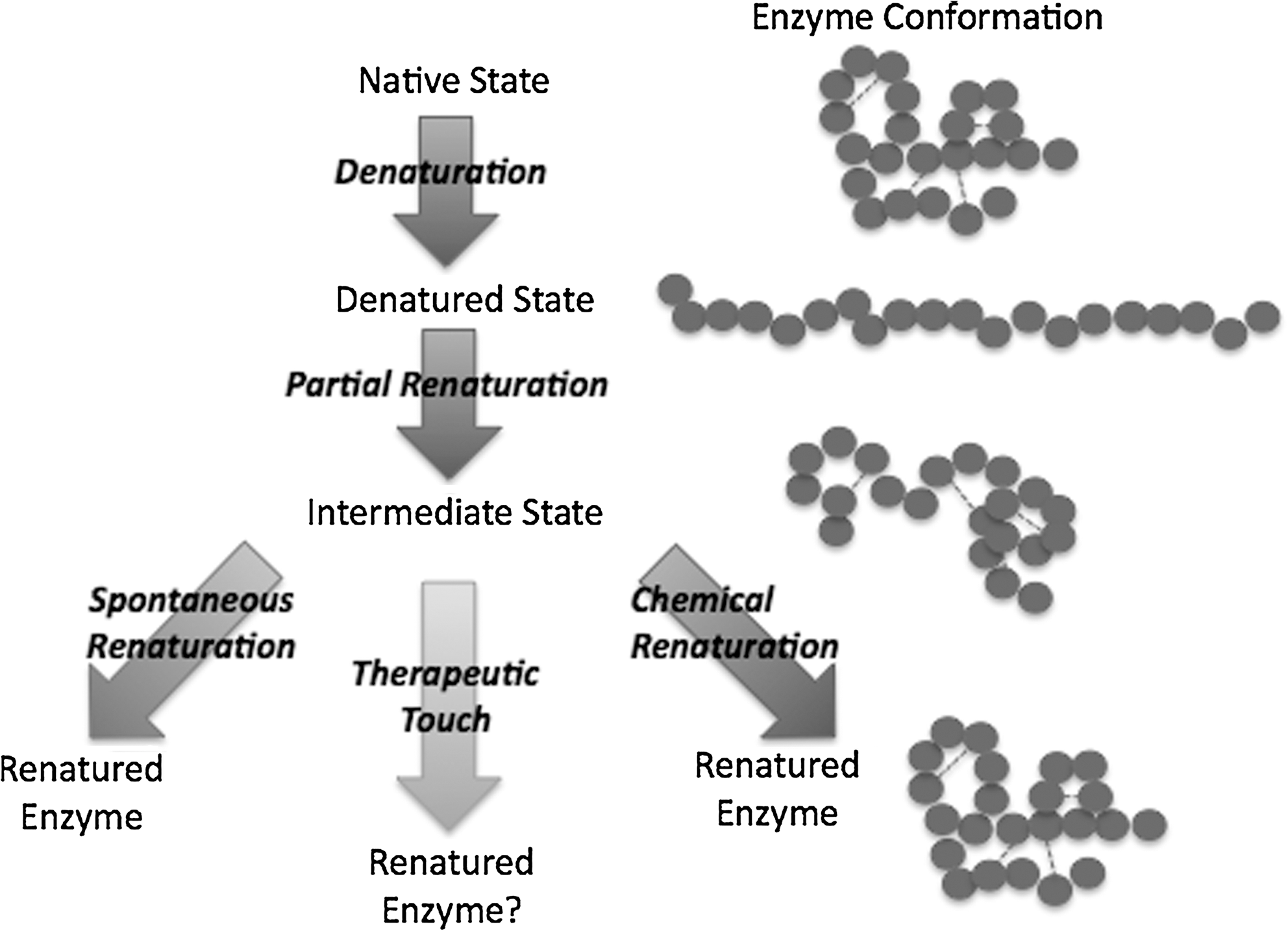
Reaction schematic. This reaction schematic shows the process by which the enzyme samples were folded and tested.
Step 1: Unfolding
Bovine pancreatic ribonuclease (3.6 × 10−6 mol/L) was incubated in buffered urea (8.0 mol/L) solution with BME (1.4 × 10−3 mol/L) for 60 minutes. 23 –26 All solutions were prepared in 0.01 mol/L (3-(N-morpholino) propanesulfonic acid) (MOPS) and 2 mmol/L ethylenediaminetetraacetic acid (EDTA) buffer at pH of 7.5.
Step 2: Controlled refolding to intermediate state
Renaturation is the process by which a protein or enzyme is refolded and assumes its three-dimensional structure. Partial renaturation was performed in order to encourage the formation of misfolded protein in an intermediate energy state. The enzyme solution from the denaturation step was transferred to dialysis bags (pore size 3000 D), which were then dialyzed for 1.5 hours in 8.0 mol/L urea at 30°C in approximately 1:100 ratio of enzyme: dialyzing solution to remove BME and encourage fast, improper renaturation. 18,24,25
Step 3: Refolding chemically, spontaneously, and by TT
Complete renaturation was performed in order to affirm that the enzyme could be chemically renatured to its native state. The dialysis bags from the partial renaturation step were then dialyzed in 300-mL solutions of 0.01 mol/L MOPS buffer with 2 mmol/L EDTA at a pH of 7.5. In this step, trace BME (1.4 × 10−4 mol/L) was added to encourage the breaking and proper reformation of the disulfide bonds. This solution was exchanged three times, after 30, 90, and 150 minutes.
To serve as a control for the TT treatment, partially renatured solutions were allowed to slowly, spontaneously renature after removal of the urea through dialysis in buffer. No BME was added to these solutions so that chemically induced renaturation would not occur. The dialysis solution was also exchanged 3 times, after 30, 90, and 150 minutes.
TT was performed on dialysis bags in 100 mL of buffer for 7 minutes and the dialysis bags were then transferred to 300 mL of buffer and dialyzed as for spontaneous renaturation. The practitioner did not touch either the beaker holding the dialysis bags or the bags themselves at any point. TT was performed with hands 2–4 inches away from the vessel, as shown in Figure 3, by a Nurse Healers–Professional Associates International (NH-PAI)–certified TT practitioner. A single practitioner was utilized due to limited resources and time to complete the study.
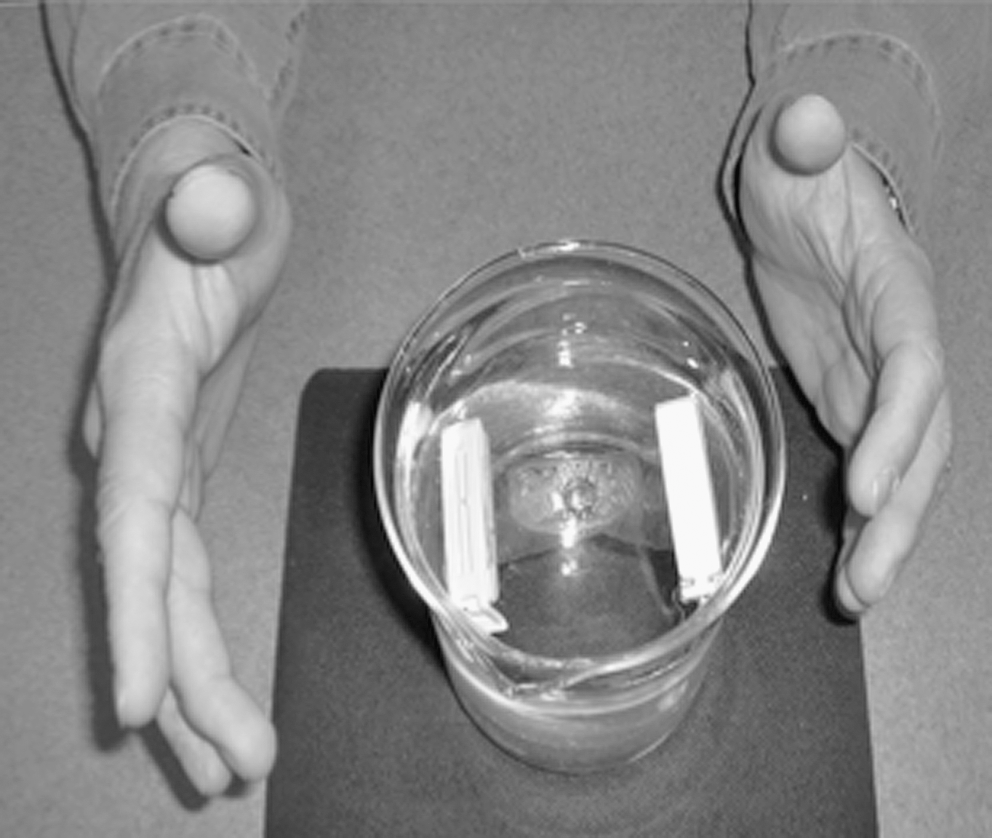
Therapeutic Touch performed during dialysis.
All solutions were maintained at 30°C throughout the experiment and were kept in the same room to ensure equal light exposure.
Measurement
Measures of structure and function, a kinetic assay and fluorescence spectra, respectively, were performed to indirectly test the energy state at each step.
Ribonuclease activity was assayed by monitoring the change in absorbance at 688 nm of 500 μL of 4.97 × 10−4 mol/L methylene blue intercalated into 50 μL of 10 mg/mL RNA for 10 minutes, upon addition of 500 μL ribonuclease (3.6 × 10−6 mol/L) as described by Greiner-Stoefflele et al. 21 Average rates were obtained through a kinetic analysis over 1 minute.
Fluorescence emission spectra of the enzyme solutions (3.6 × 10−6 mol/L) were recorded from 200 to 700 nm following excitation at 278 nm. In theory, the degree of denaturation correlates with a decrease in peak intensity at 305 nm due to the increased solvent exposure of tyrosine residues. 27
Samples were assayed with both methods after denaturation, partial renaturation, and complete renaturation steps. In total, three trials with replicates were performed to test TT. All samples were analyzed blindly.
Results
A kinetic assay was used to quantify the enzyme's activity in order to better understand the enzyme's energy state as it relates to the native state, which is defined by the ability to function. The activity assessed is ribonuclease A's ability to degrade RNA by providing substrate RNA intercalated with methylene blue. As RNA is degraded, methylene blue is released, which corresponds to a decrease in the solution's absorbance at 688 nm over time. Therefore, a more active sample of ribonuclease A will show a decrease in absorbance more quickly than a less active sample.
Figure 4 shows the activity data collected for a single trial having three replicates. There was a significant difference in activity between the intermediate state samples and the chemically renatured, spontaneously renatured, and TT-treated samples. There was also a significant difference between the chemically renatured and the spontaneously renatured and TT-treated samples. However, the data do not suggest a significant difference between the TT-treated samples and the spontaneously renatured samples.
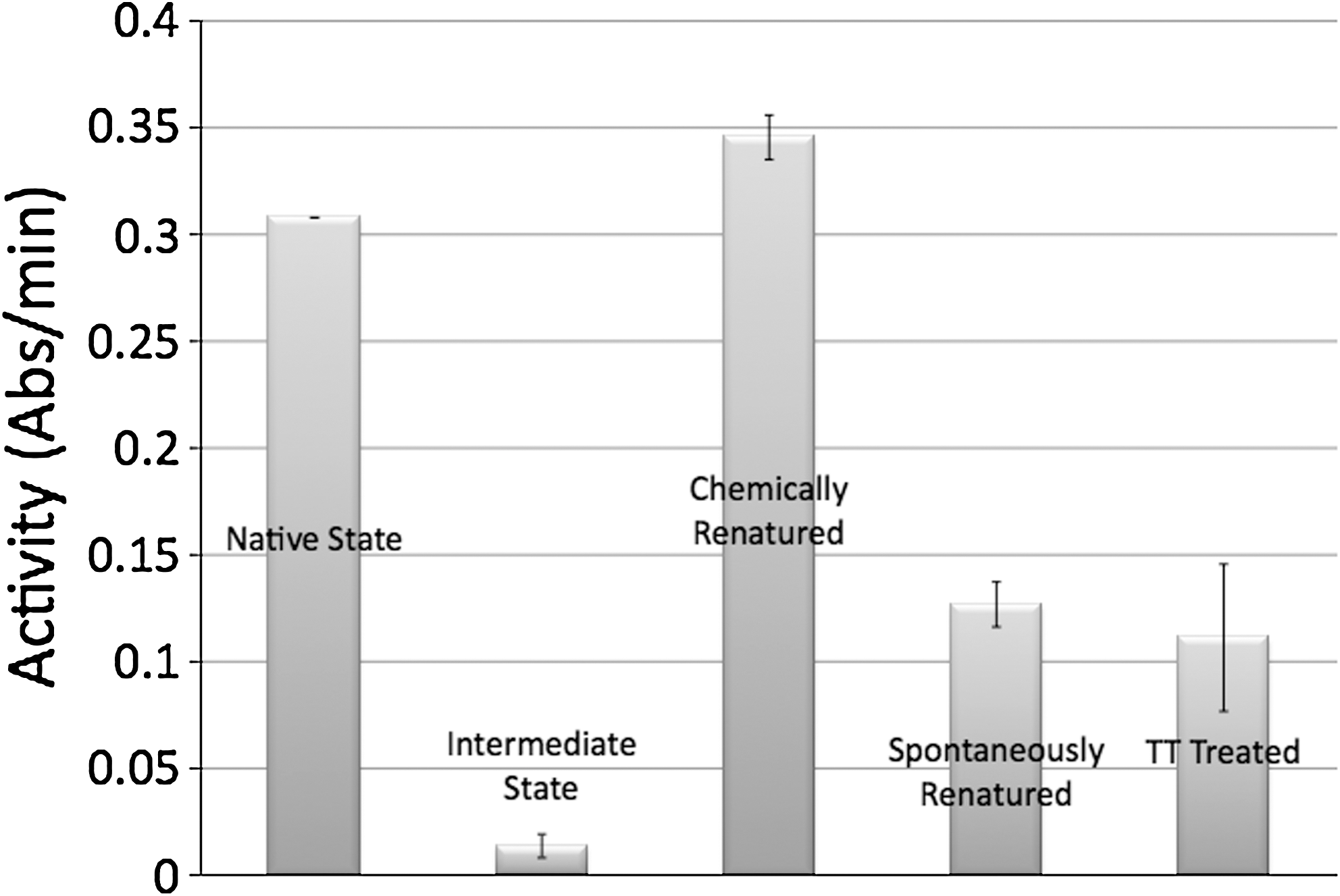
Activity data for optimized trial. Error bars = ± standard deviation. It should be noted that the native state sample was prepared in bulk (n = 1), and then the denatured sample was divided into three samples for partial renaturation to the intermediate-state samples (n = 3). Each of these samples was then divided into three replicates for each treatment, as follows: chemically renatured, spontaneously renatured, and Therapeutic Touch (TT)–treated samples (n = 3).
Fluorescence spectra were used to qualitatively monitor the changes in protein structure. The denatured sample was shown to have a greater fluorescence peak intensity than a native state or renatured sample due to the increased exposure of tyrosine in the denatured state. Figure 5 shows the differences in the fluorescence peak intensity for a given sample in its intermediate, spontaneously and chemically renatured states. However, fluorescence spectra did not reliably detect a statistical difference among these states.
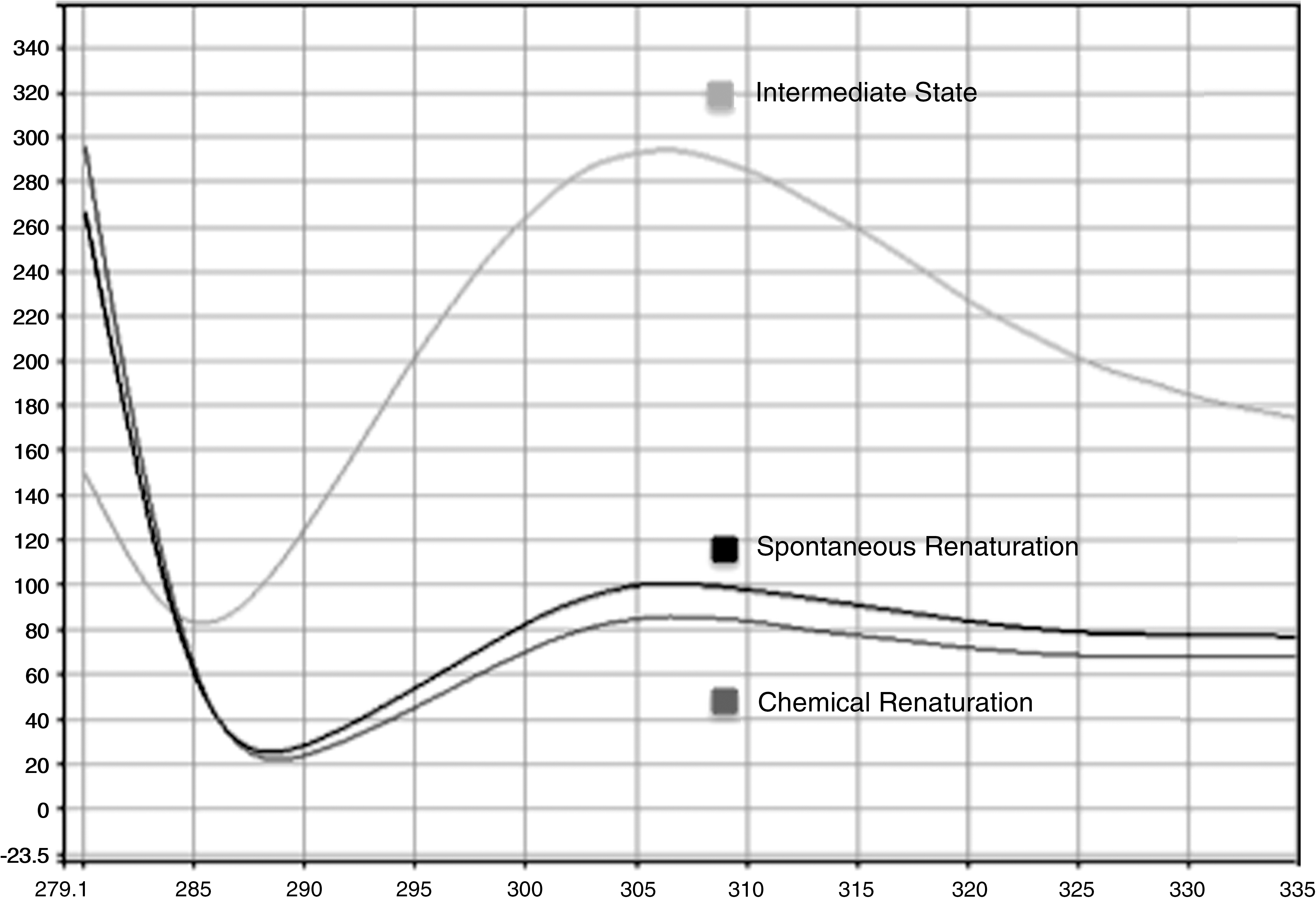
Fluorescence spectra of intermediate state, spontaneously and chemically renatured samples of a single trial. Shown here is the effect of chemical and spontaneous renaturation compared to the intermediate sample at a peak intensity at 306 nm.
Discussion
The purpose of this work was to design a model and run preliminary tests that can adequately study the energy exchange underlying the effects of TT. The study proposed was designed to satisfy the first four assumptions of TT in order to test the fifth, that TT offers a form of communication, or intervention, into this energy in order to restore balance.
First, the requirement that TT must be performed on an open system engaged in energy exchange is addressed by utilizing a protein system that can be structurally altered with the addition of energy. BME was used in the chemical renaturation samples to promote disulfide bond reduction and thus provided energy in the form of available electrons. Breaking disulfide bonds that have improperly formed in the intermediate structures allows the refolding of the enzyme and increases the chances that the enzyme will refold to its most stable state.
Second, as in the human model, proteins possess an inherent quality of energy, albeit much less complex. Specifically, the aggregate effect of the interactions between individual amino acids composing a protein and the environment creates a definable energy state specific to the given sequence of amino acids and the given conformation. The inability to isolate a single energy-requiring system within the human model makes it nonideal to test the ability of TT to “communicate, intervene and restore balance” to an energetically unstable system. 21
Third, a perceivable energy field must exist. Perception must be possible through both the methods of analysis and by the TT practitioner. A biochemical assessment of the energy field was possible indirectly by assessing activity, which is optimal when the enzyme is folded into its lowest energy state. The practitioner in this study was also able to perceive the energy of the samples and described it as “red, green and blue energy of overlaid, askew bubbles.”
The fourth principle is that TT cannot improve upon a healthy, balanced system but rather can only put into balance an otherwise imbalanced system. This study proposes that the intermediate state of the enzyme represents an energy deficit because it requires some energy to be refolded into its lowest energy conformation and is therefore in a state of energy imbalance.
In order to assess the changes in energy of the proposed model, this study sought to characterize the function and structure of the enzyme. The kinetic assay assessed the function of ribonuclease A by quantifying the activity of the enzyme in solution. The assay was able to distinguish between the native, denatured, chemically renatured, and spontaneously renatured samples on the basis of their activity. By including a chemical control and spontaneous control, this assay sets up a range of activity allowing for the relative assessment of TT treatment. For a given sample, the maximum activity would be the rate of the chemical control, and the baseline activity would be the rate of the spontaneous control.
The limitation of the kinetic analysis is that it cannot distinguish between the intermediate and denatured samples, since neither the intermediate nor denatured state of the enzymes is active. To address this limitation, structural analysis was performed by collecting fluorescence spectra at each assay point. The fluorescence spectra were insufficient in three ways. First, this analysis failed to adequately and consistently distinguish between the denatured and intermediate state samples, despite many efforts to optimize the procedure. It is possible that the degree of tyrosine exposure between the denatured and intermediate states is not significantly different and thus is not an appropriate parameter on which to distinguish these two states.
In addition, fluorescence spectra are very sensitive to solution effects or slight variations in pH, temperature, or concentrations, which would add variability to the results that could change between replicate sets and the progressive optimization that occurred.
For the reasons addressed above, fluorescence spectroscopy may not be the best technique to characterize the structure of ribonuclease A. We suggest that future work be done using circular dichroism spectroscopy, which would give a much more detailed picture of the enzyme structure. Although this addition could potentially complicate the project's design, we suggest that future studies include multiple practitioners. It is also possible that another enzyme model, such as rhodanese, with a clearly characterized intermediate species might produce more significant structural results. 28
This study was not sufficiently powered to make statements regarding the validity of TT due to a limited number of repetitions and the use of only one practitioner. However, the data presented do suggest that the degree of renaturation exhibited in the TT samples was not significantly different from the degree of autorenaturation, considered to be baseline, by the kinetic assay of the spontaneously renatured samples. There are many potential interpretations of this trend. It is possible that TT did provide energy to the enzymes, allowing them to refold, but was not able to refold them to their native state. The methods presented here were not sufficient to explore this possibility.
Although the results do not support or reject the ability of TT to affect enzyme folding, the model is conceptually valid to assess the physical qualities of energy exchange. This feasibility study offers a novel model and methods, which, with greater replicate sets, multiple practitioners, and a more precise means of characterizing protein structure, will elucidate and describe the mechanisms underlying TT.
Conclusions
The study of TT requires a model engaged in open change of energy that is perceivable and related to the health or function of the model itself. It is important to study the mechanisms underlying the effects of TT in order to complement the work being done to characterize TT in the clinical and in vitro settings. The presented work offers a novel means of studying TT by measuring its ability to affect enzyme refolding and energy state. The enzymatic intermediate state is, by definition, energetically trapped. It is, therefore, an excellent model for this research because in order for a conformation change to occur, energy must be added to the system. Whereas the majority of the current literature on this topic has focused on characterizing TT, the results presented here suggest that further research using this model system may provide information about the mechanism underlying TT.
Footnotes
Acknowledgments
The authors are grateful for the contributions of the Westminster College chemistry and biology departments as well as the Westminster College Drinko Center for Excellence in Teaching & Learning for its generous research and travel awards. The authors also recognize Mr. Glenn Wood for his service and expertise. The authors recognize Dr. Megan Pailler for editorial comments.
Disclosure Statement
No competing financial interests exist.