
Abstract
Niemann–Pick disease type C (NPC) is an inherited lipid storage disorder characterized by a defect in intracellular trafficking of exogenous cholesterol and glycosphingolipids. A goal for therapeutic treatment of NPC is to decrease/normalize cholesterol accumulation. We developed a functional genomics-based assay, combining high-throughput RNA interference (HT-RNAi) screening with high-content fluorescence imaging to identify specific genes in NPC cells that will result in more normal cholesterol levels in the diseased cells. Conditions for siRNA tranfections were optimized for 2 NPC fibroblast cell lines (GM03123, GM18453) and a normal fibroblast cell line (GM05659). RNAi screening was done using a focused-set siRNA library targeting 40 cholesterol trafficking-associated genes, knowledge mined from the existing literature on NPC disease, and/or their association with NPC1/NPC2 genes. We utilized filipin staining as a measure of cholesterol accumulation in fixed NPC cells. Data analysis of these screens confirmed several genes including LDLR and RAB9A that reduced cholesterol content in NPC cells. Nine genes were validated using filipin staining to detect unesterified cholesterol as well as cholesteryl BODIPY® esters to study lipid trafficking. Gene silencing was also confirmed using qRT-PCR. Our results show that this technology can be applied to larger screens to identify genes responsible for lipid accumulation and/or trafficking in NPC disease, which could be instrumental in developing innovative therapies for individuals afflicted with NPC disease.
Introduction
Identification of gene products that act as modulators of cholesterol accumulation/trafficking will facilitate the design and development of better therapeutic strategies for the treatment of Niemann–Pick disease. Recently, several groups have performed compound screening to identify biologically active compounds that correct the NPC phenotype. 10,12 Pipalia et al. developed a high-throughput technology based on filipin staining to identify unesterified cholesterol accumulation in cultured Chinese hamster ovary cells with an NPC phenotype. 10
In this study, our experiments were driven by the hypothesis that the targets identified by high-throughput RNA interference (HT-RNAi) screening of the NPC cells should serve as important targets for drug development in NPC disease. To accomplish this task, we combined a HT-RNAi screen with high-content cell-based analysis to identify modulators of cholesterol accumulation specifically in NPC fibroblast cells. To demonstrate and validate our approach, we chose 40 genes that are known to play a role in cholesterol accumulation or trafficking from the previously published articles. Using the HT-RNAi and high-content analysis-based approach, together with robust statistical analysis methods using intensity and area changes in filipin staining, we functionally validated the role of several target genes. To our knowledge, this is the first report showing the modulation of cholesterol accumulation specifically in NPC cells using a high-throughput platform. In addition, we based our image analysis on cellular data by reporting cell-by-cell distribution of filipin staining intensity and area, instead of considering the average values. This robust platform offers a method for large functional genomics screening for the identification of functional drug targets for NPC as well as for other lipid storage disorders.
Materials and Methods
Cell Culture
The human NPC fibroblast cells GM03123 and GM18453 and the normal fibroblast cell line GM05659 were obtained from the Coriell Institute for Medical Research (Camden, NJ). Cells were grown in minimal essential medium (MEM) supplemented with 10% fetal bovine serum (FBS), 2 mM
Reagents
Short interfering RNA targeting the genes of interest and nonsilencing control were obtained from Qiagen (Valencia, CA). Primer sets for quantitative real-time polymerase chain reaction (PCR; qRT-PCR) were also obtained from Qiagen. SYBR green for qRT-PCR analysis was from Applied Biosystems (Carlsbad, CA). Filipin III was obtained from Sigma (St. Louis, MO). DRAQ5™ was from Axxora (San Diego, CA) and cholesteryl 4,4-difluoro-5-(2-pyrrolyl)-4-bora-3a,4a-diaza-s-indacene-3-undecanoate (cholesteryl BODIPY® 576/589 C11) was obtained from Invitrogen.
RNAi Screening
High-throughput RNA interference (HT-RNAi) was performed using a custom focused-set siRNA library targeting genes associated with cholesterol transport and accumulation and obtained from Qiagen. The library contained 4 siRNAs targeting each gene for screening as single siRNA per well. Stock siRNA was diluted in siRNA buffer (Qiagen) and 9.3 ng of each individual siRNA was printed onto black clear bottom Corning 96-well plates (Fisher Scientific, Pittsburgh, PA). HT-RNAi was done by reverse transfection of cells. 13 The transfection conditions were rigorously optimized for maximal transfection efficiency with minimum toxicity to the cells. In brief, diluted siLentFect reagent (Bio-Rad, Hercules, CA) in OptiMEM (Invitrogen) was added to the wells and allowed to complex with siRNA for 30 min at room temperature. Cells were resuspended in growth media without antibiotics and plated at a final concentration of 3,000 cells/well. Plates were incubated at 37°C with 5% CO2 for 96 h. The final siRNA concentration in the wells was 13 nM.
Cholesterol Measurement
We quantified cellular unesterified cholesterol using filipin. Cells were washed with phosphate-buffered saline (PBS) and fixed with 4% paraformaldehyde. Cells were then blocked for an hour with tris-buffered saline (TBS) containing 5% normal goat serum, 0.02% sodium azide, 0.2% bovine serum albumin (BSA), and 0.1% Igepal CA-630. Fixed cells were stained with 25 μg/mL filipin and 5 μM DRAQ5™ for 1 h at room temperature. Stained cells were washed twice with PBS.
Cholesteryl BODIPY Ester Transport
To evaluate cholesterol trafficking, a fluorescent lipid probe, cholesteryl BODIPY® 576/589 C11 was added at a concentration of 1 mM in complete Dulbecco's modified Eagle's medium (DMEM) media to each well and further incubated for 24 h. The plates were then fixed with 4% paraformaldehyde, blocked, and stained with 25 μg/mL filipin and 5 μM DRAQ5™ at room temperature for 1 h. Stained cells were washed twice with PBS.
High-Content Image Acquisition
In the primary screen, the IN Cell Analyzer 3000 (GE Healthcare, Piscataway, NJ) equipped with an argon laser (360 nm, 488 nm) and krypton laser (647 nm), a 40× Nikon ELWD objective 0.6 numerical aperture, and three 12-bit CCD cameras (1,340× 100 pixels; Princeton Scientific, Princeton, NJ) were used to acquire images. One image was acquired at the center of each well using no binning. Filipin-stained images were acquired using a 364-nm excitation and 450/25-nm emission filters. DRAQ5™ images were acquired using a 647-nm excitation and 695/55-nm emission filters.
In the confirmation screens, the IN Cell Analyzer 1000 (GE Healthcare) equipped with an inverted Nikon Eclipse TE2000-U microscope, a 100 W Xenon lamp (BD Biosciences, San Jose, CA), a 10× Plan Apochromat objective 0.45 numerical aperture, and a 12-bit Photometrics CoolSNAP camera (1,392 × 1,040 pixels; Roper Scientific, Tucson, AZ) was used to acquire images. The IN Cell Analyzer 1000 was preferably used instead of the IN Cell Analyzer 3000 in the confirmation screen because of its optimal image acquisition parameters. Two images per well were acquired using no binning. Filipin-stained images were acquired using a 360/40-nm excitation and 460/40-nm emission filters. Cholesteryl BODIPY® C11 stained images were acquired using a 535/50-nm excitation and 620/60-nm emission filters with a 650-ms exposure time. DRAQ5™-stained images were acquired using a 620/60-nm excitation and 700/75-nm emission filters with a 500-ms exposure time. FF410/504/582/669-DIO1-25 × 36 dichroic filter (Semrock, Rochester, NY) was used to acquire all images.
High-Content Image Analysis
Image analyses for initial screening using filipin and DRAQ5 were performed using the IN Cell Investigator software (GE Healthcare). Images of filipin-stained cells were segmented and analyzed using the morphology analysis module. The average filipin intensity, total filipin staining area, and total filipin intensity were calculated and used to perform a cell-by-cell data analysis. Images of DRAQ5™-stained nuclei were segmented and used to calculate nuclear count. For confirmation screens, filipin, DRAQ5, and cholesteryl BODIPY-stained cells were imaged. Images of cholesteryl BODIPY-stained cells were segmented and analyzed using a granularity analysis module of the IN Cell Investigator software package. The total granule intensity per cell was calculated and used to perform data analysis.
Data Analysis
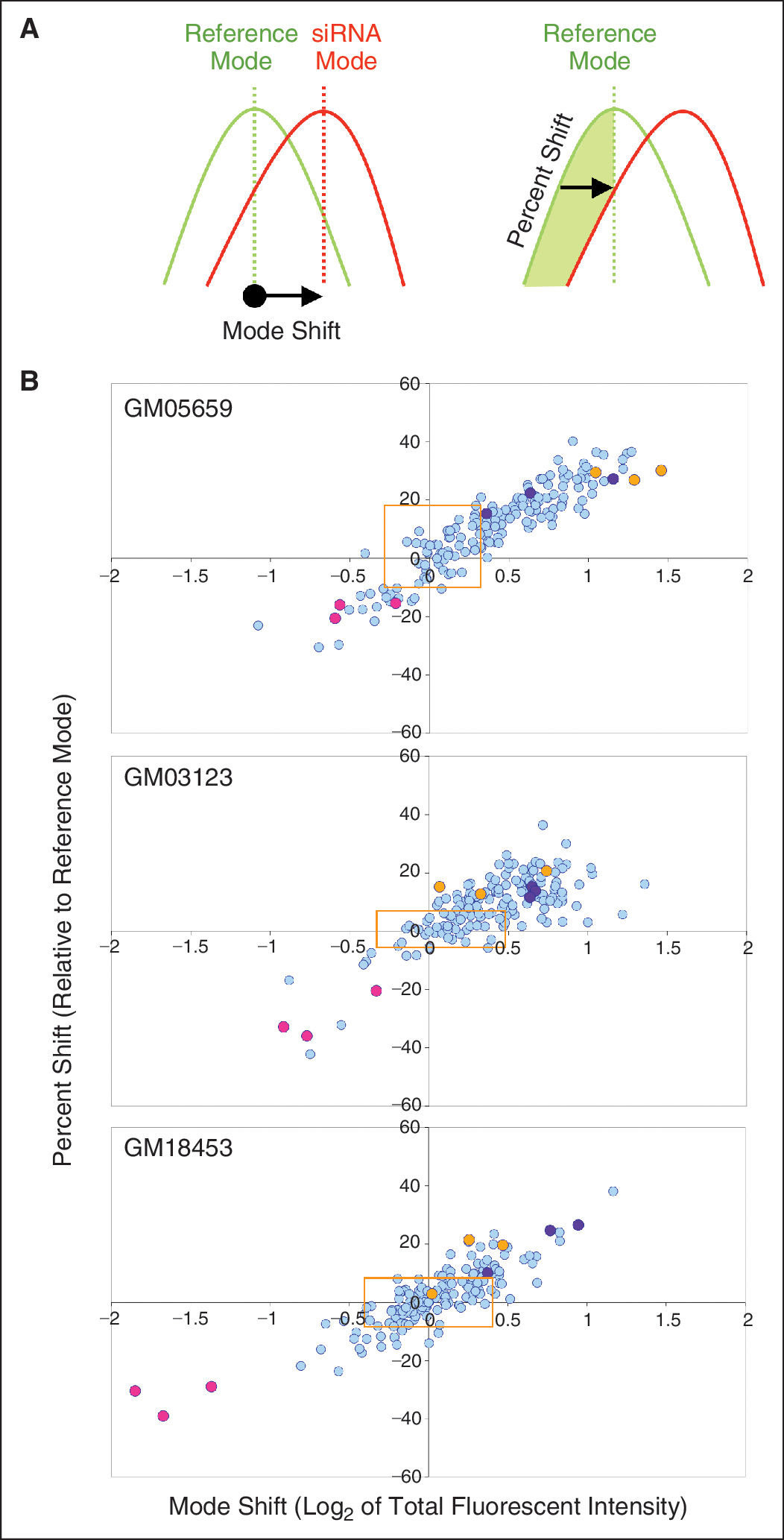
Cell-by-cell measurements (average filipin intensity, total filipin staining area, and total filipin intensity) were log2 transformed before the distribution was calculated (MATLAB; Mathworks, Inc., Natick, MA). Wells without enough cells were removed from further analysis. The cells from all the untreated wells were pooled and their distribution was used as reference (reference distribution) and their mode was used as reference mode. Distribution of measurements for each siRNA was compared with reference distribution and 2 statistics were computed: (1) mode shift, which is the extent (with direction) of the mode change relative to reference mode and (2) percent shift, which indicates the change in the percentage of cells that are to the left or right of reference mode relative to that in reference distribution. These data are represented as 2D scatter plots of mode shift vs. percent shift for each siRNA treatment of the 3 cell lines (see Fig. 1). Z′ factors for the assay using each cell line were calculated according to Zhang et al. 14 using total intensity data from a 96-well plate containing specific siRNA controls as shown in Appendix Table 1
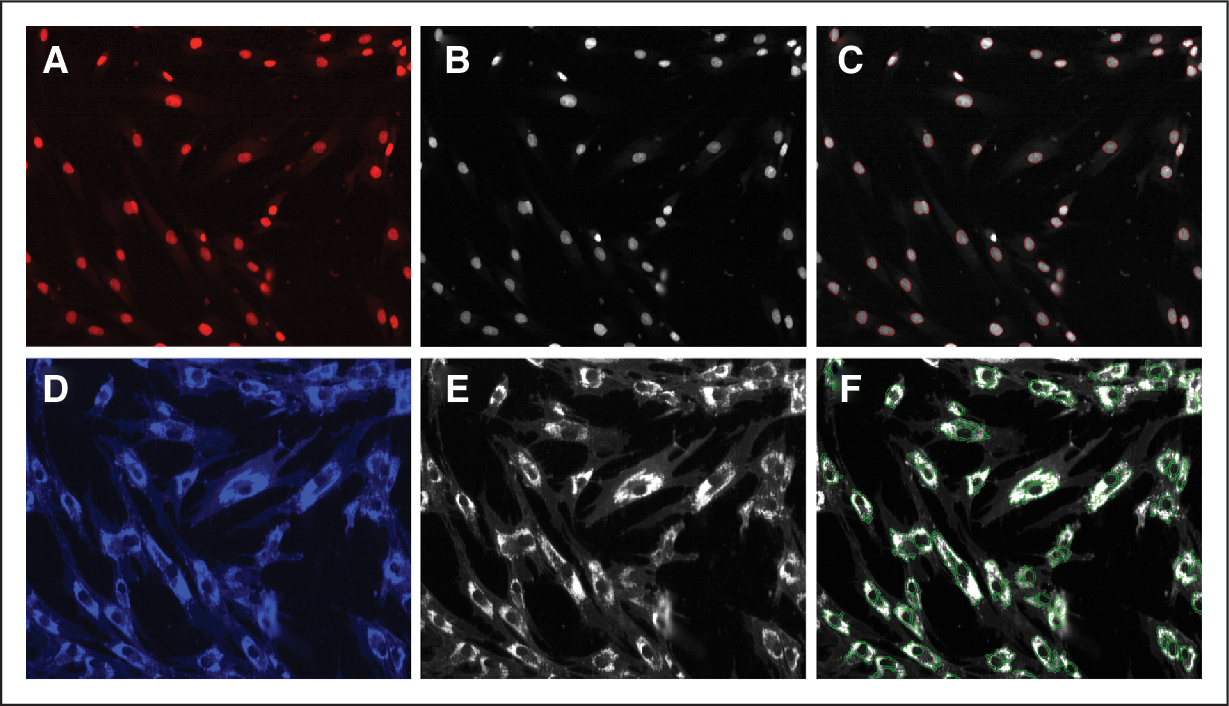
Quantitation of cholesterol accumulation by measuring filipin fluorescence. The NPC fibroblast cell line GM03123 was grown to sub-confluency, fixed with paraformaldehyde and stained with DRAQ5 and filipin. Fluorescent images were acquired using the IN Cell Analyzer 3000 with a 40 × objective for DRAQ5 staining (
Results
Knowledge Mining and Selection of the Focused-Set Genes
A customized set of genes was selected from the literature using the text-mining software Pathway Studio 6.0 (Ariadne Genomics, Rockville, MD). These genes were selected to serve as a putative positive control set for the detection of NPC1/NPC2-regulated activity. Pathway Studio 6.0 is pathway/network database created through a custom natural language processing (NLP) method. NPC1 and NPC2 were seeded into a blank pathway and a direct interaction network from each node was used to build all interactions in the database form each node. The resulting list of interactions was manually inspected to remove any false positive/negative interactions. Forty genes were selected for assembly of a focused-set siRNA for development of the cholesterol modulation assay (Table 1). A custom siRNA set was obtained targeting the selected 40 genes using 4 siRNAs per target (Appendix Table 2).
Genes Chosen for the Focused-Set siRNA Library
Establishment of a High-Content Assay for Measuring Cholesterol Accumulation in NPC Fibroblasts
In order to identify genes that modulate cholesterol accumulation in disease-relevant cells, we utilized the NPC fibroblast cell lines GM03123 and GM18453 and one normal fibroblast cell line GM05659 for assay development. A high-content imaging assay was developed using a fluorescent marker for cholesterol accumulation. Measurement of cholesterol accumulation was done by staining the fixed NPC/normal fibroblast cells with filipin, which is a specific marker for unesterified cholesterol and DRAQ5™, which stains the nuclei of cells. 10,15 Using the IN Cell Analyzer 3000, images were acquired for both DRAQ5 and filipin for all the 3 cell lines. Figure 2A and 2D show DRAQ5 and filipin fluorescent staining of GM03123 NPC cells, respectively. Images were converted to grayscale (Fig. 2B and 2E) and fluorescent staining was segmented using IN Cell Investigator software (Fig. 2C and 2F). Total filipin fluorescent intensity was obtained by multiplying mean fluorescent intensity and total fluorescent area. Cholesterol accumulation in the NPC fibroblast cells as seen by filipin staining was measurably higher than in the normal fibroblast cells (Fig. 3A–3D).
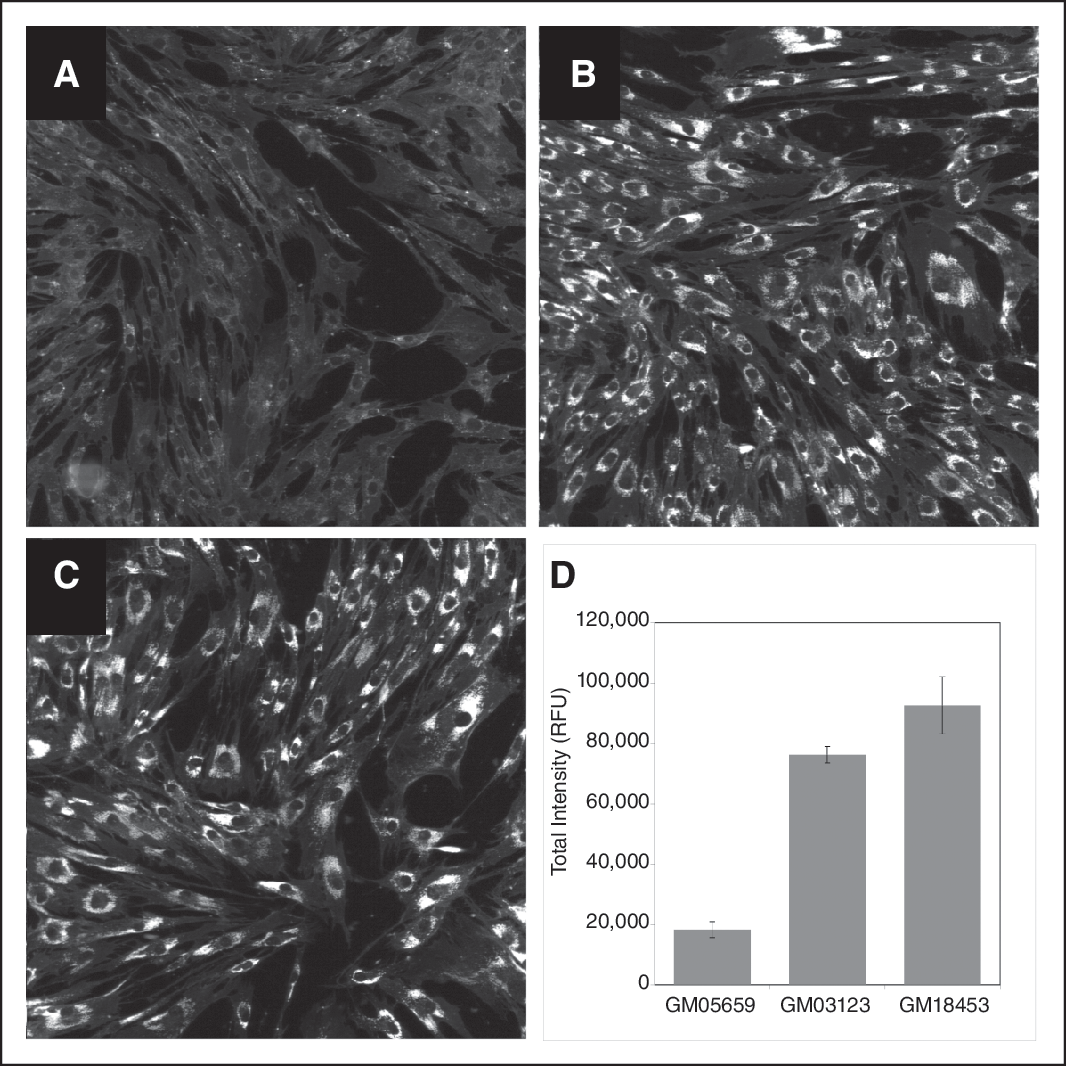
Cholesterol accumulation in NPC and normal human fibroblasts. Normal human fibroblast cell line (
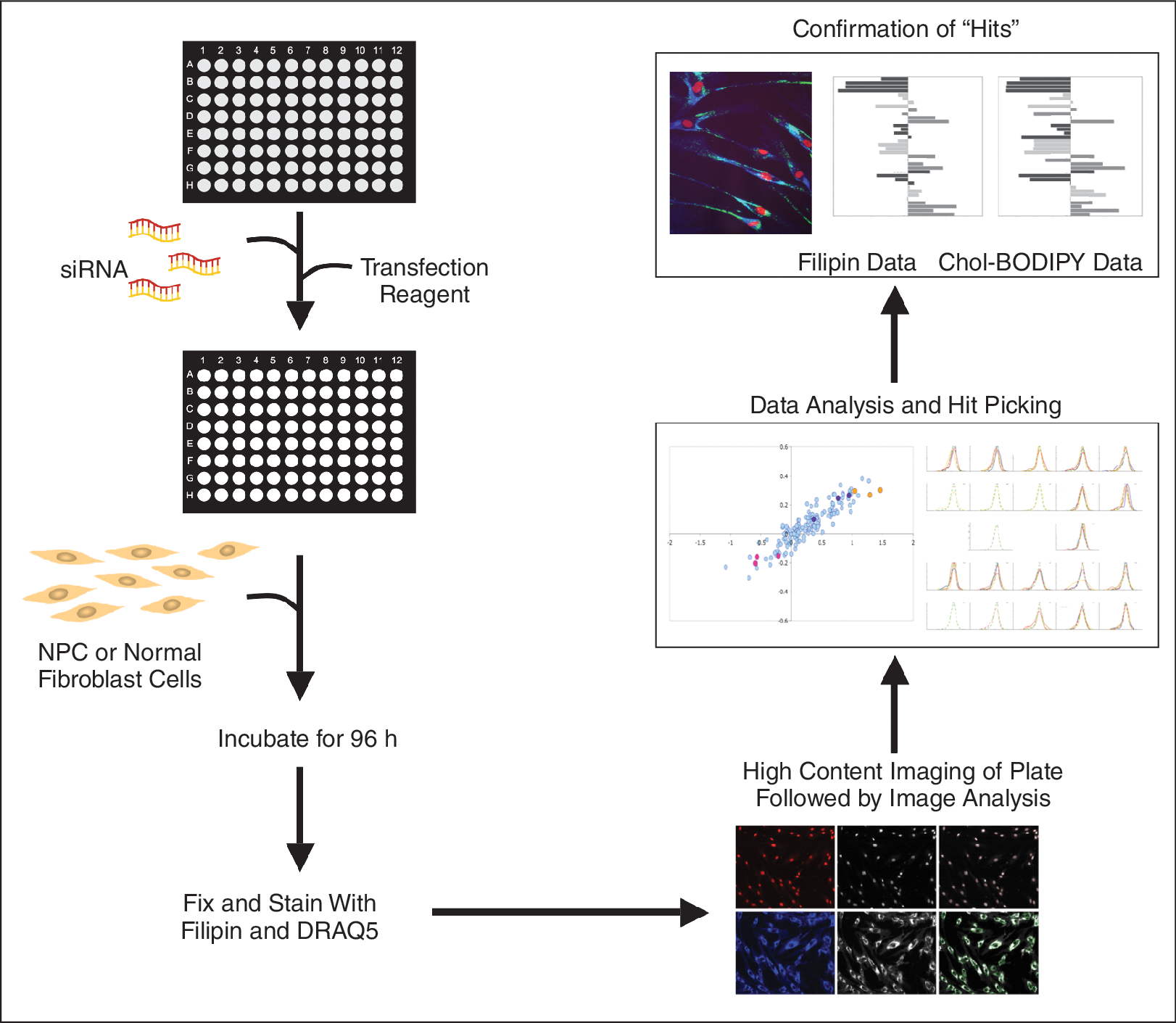
Schematic of high throughput, high content siRNA screening in NPC cells. This flow diagram shows the various steps in the HT-RNAi screening to identify modulators of cholesterol accumulation and trafficking in NPC cells.
RNAi Screening for Modulators of Cholesterol Accumulation in NPC Cells
To identify genes that modulate cholesterol accumulation in NPC cells, we first established conditions for effective gene silencing using siRNA for each individual fibroblast cell line. Transfection optimization was performed using a panel of commercially available transfection reagents and siLentfect at a ratio of 6:1 with OptiMEM showed the best transfection efficiency (Appendix Fig. 1). Other parameters including cell number and incubation times were also optimized. The result was a HT-RNAi assay that allowed for high-efficiency siRNA transfection of NPC fibroblast cells by cationic lipids in 96-well plates (Fig. 4).
High content siRNA screening for modulators of cholesterol accumulation. (
The RNAi screen involved transfecting either the normal fibroblast cell line GM05659 or the NPC cell lines GM03123 or GM18453 with a custom siRNA library targeting 40 cholesterol regulating genes with 4 siRNAs per gene in order to accumulate more robust data (Appendix Table 2). After 96 h, plates were fixed and stained with filipin and DRAQ5, wherein filipin stained the cholesterol containing vacuoles and DRAQ5 was used to stain the nuclei to get cell count in every field of image acquisition (Fig. 4). Image analysis was done using the morphology module on IN Cell Investigator software (Fig. 2C and 2F). The data collected included nuclear count using DRAQ5-stained nuclei, filipin fluorescent intensity (I), filipin fluorescent area (A), and total intensity (I × A) values for each cell in the acquired field of every well. These data were analyzed based on cell-by-cell measurement according to the protocol outlined in subsection “Data Analysis” under “Materials and Methods.” Log2 values of the total intensity were calculated for each cell in an image field and plotted to obtain the mode. Plots of siRNA-treated cells were compared with a reference of untreated (no siRNA) controls to obtain mode shift and percent shift (Appendix Fig. 2). Mode shift and percent shift values were calculated for each cell line with each siRNA treatment (Appendix Table 3). Screening data for each cell line was plotted as mode shift vs. percent shift of total intensity for each siRNA treatment (Fig. 1). Z′ factors were also calculated according to Zhang et at. 14 using control siRNA NPC2_6 and LDLR_5 and are shown in Appendix Table 1.
This statistical analysis used mode over mean or median to allow for potential mass shifts of subpopulations. The data show predominant unimodal distributions, as indicated by the good correlation of mode shift to percent shift. As we plan to use these statistics for data analysis in the future and the distributions are not always symmetrical, we found that the mode statistics are better indicators of where the majority of the mass is located as compared to mean or median. Comparison of mode shift or percent shift with mean total intensity also shows good correlation highlighting the unimodal distribution of the data (Appendix Fig. 3). A gene was called a “hit” if at least 2 siRNAs were outside the 1.65 standard deviation of the nonsilencing siRNA values for mode shift and percent shift (Appendix Fig. 4). Analysis of the siRNA screening data showed that knockdown of low-density lipoprotein receptor (LDLR), APOL1, and RAB9A significantly reduced cholesterol accumulation in NPC cells, while the knockdown of NPC1, NPC2, NPC1L1, STARD3, STARD4, and sphingomyelin phosphodiesterase 1 (SMPD1) showed increased accumulation of cholesterol in the NPC cells. Of the 40 genes, these 9 gene targets were selected for further validation based on their ability to modulate cholesterol accumulation (Table 2). To demonstrate the silencing efficiency of the siRNA targeting the 9 gene hits, we performed qRT-PCR using specific primers for the target genes. The qRT-PCR results showed that all the siRNAs silenced their respective genes and showed at least 70% knockdown (Appendix Fig. 5).
Functional Description of the Role of 9 Genes in Cholesterol Accumulation or Trafficking
Confirmation of Modulators of Cholesterol Accumulation in NPC Cells
In order to confirm our screening results for modulators of cholesterol accumulation, we performed siRNA and high-content screening in combination with treatment of NPC and normal fibroblasts with cholesteryl BODIPY® 576/589 C11. Cells were fixed and stained with filipin and DRAQ5 as described earlier. Image analysis included an additional image capture at 535/620 to capture fluorescent staining of the cholesteryl BODIPY 576/589 C11 uptake by the cells. Knockdown of NPC1, NPC2, NPC1L1, STARD3, STARD4, and SMPD1 increased filipin staining in the NPC cells confirming that there is a marked increase in cholesterol accumulation in these cells. In addition, we also saw a surge in the cholesteryl BODIPY in these cells showing that there is a marked increase in cholesterol trafficking. Most importantly, we see that in NPC cells with knockdown of NPC1, NPC2, and SMPD1, there is a significant increase in cholesterol accumulation as well as in trafficking (Fig. 5). Moreover, knockdown of LDLR, RAB9A, and APOL1 reduced the filipin staining in the NPC cells suggesting that there is a reduction in the accumulated cholesterol in these cells. In addition, we also see a significant reduction in the cholesteryl BODIPY in these cells showing that there is a marked reduction in cholesterol trafficking. Of significant interest is the finding that upon LDLR knockdown there is negligible amount of cholesteryl BODIPY in the NPC cells suggesting a near complete shutdown of the trafficking mechanism (Fig. 6).
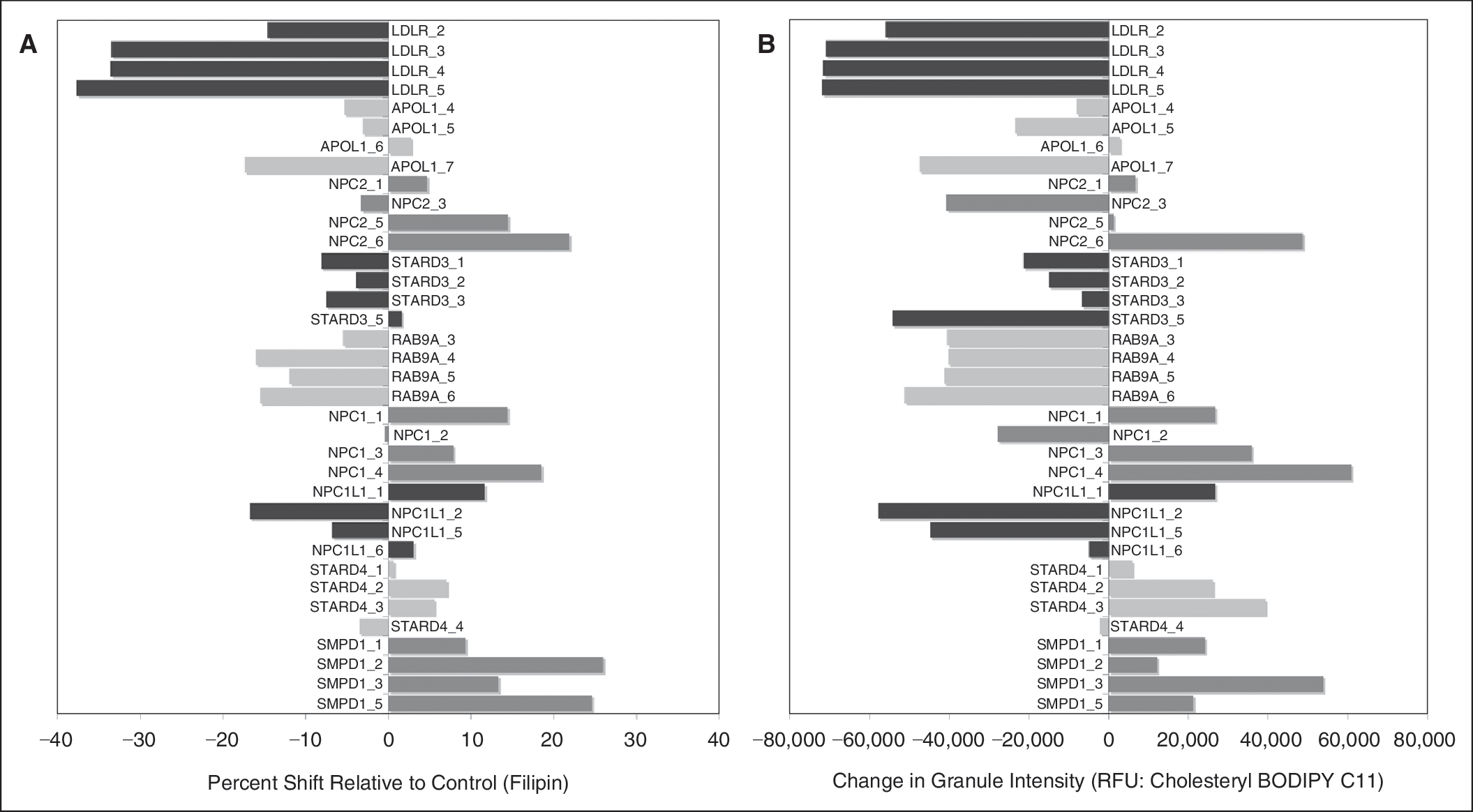
Validation of genes that modulate cholesterol accumulation. The Niemann–Pick disease type C (NPC) fibroblast cell line GM18453 was transfected with siRNA targeting low-density lipoprotein receptor (LDLR), APOL1, NPC1, NPC1L1, NPC2, STARD3, STARD4, RAB9A, and sphingomyelin phosphodiesterase 1 (SMPD1) by reverse transfection. After 54 h, cells were treated with 1 mM of cholesteryl BODIPY C11 for an additional 18 h. Cells were washed, fixed with paraformaldehyde, and stained with filipin. Fluorescent image analysis was done using the IN Cell Analyzer 1000. (
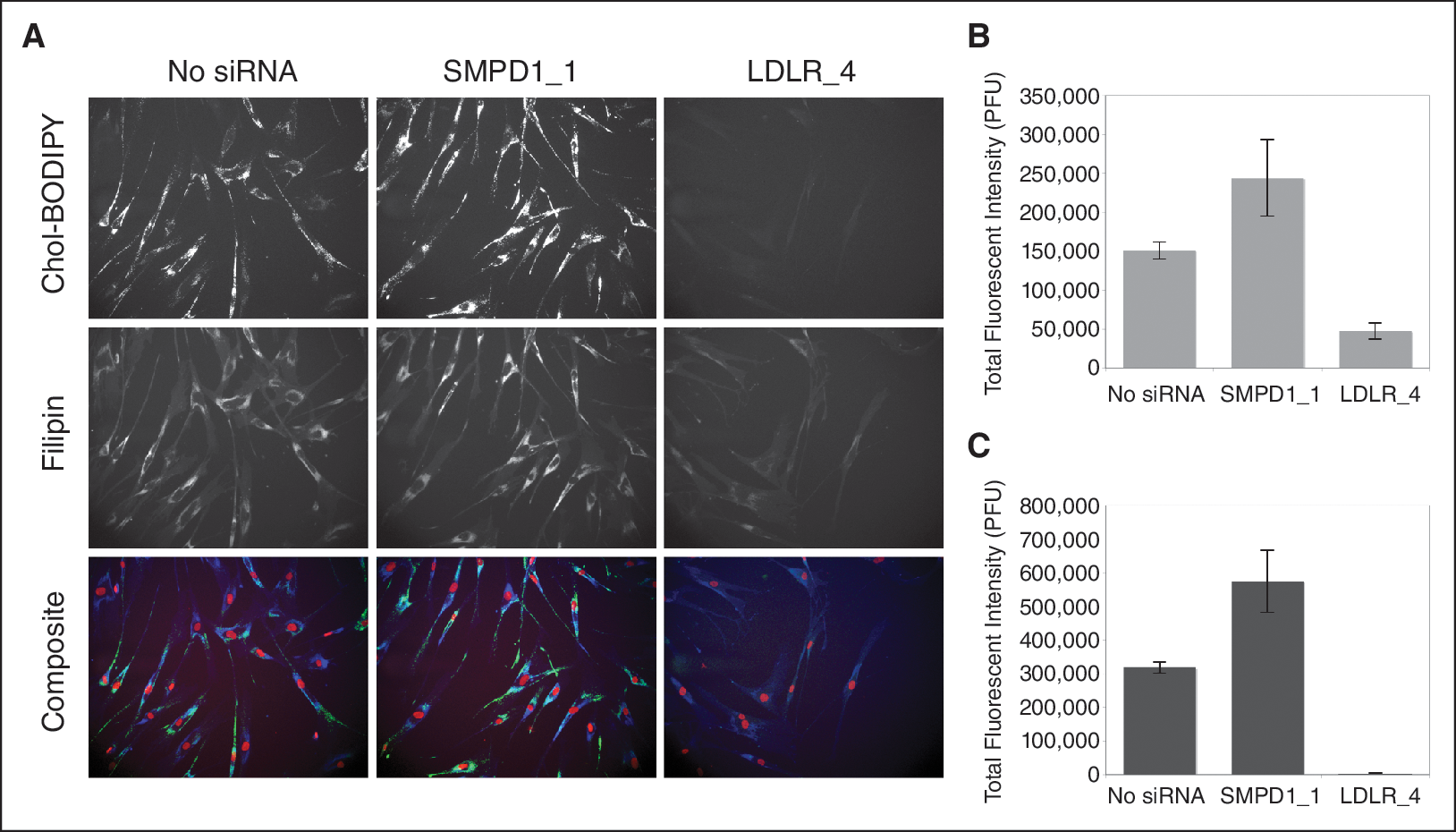
Modulation of cholesterol uptake and accumulation by silencing of sphingomyelin phosphodiesterase 1 (SMPD1) and low-density lipoprotein receptor (LDLR). The Niemann–Pick disease type C (NPC) fibroblast cell line GM03123 was transfected with siRNA targeting LDLR and SMPD1 by reverse transfection. After 54 h, cells were treated with 1 mM of cholesteryl BODIPY C11 for an additional 18 h. Cells were washed, fixed with paraformaldehyde, and stained with filipin. (
Discussion
Niemann–Pick disease type C is caused by a defect in either NPC1 or NPC2 and is characterized by accumulation of cholesterol and glycosphingolipids in acidic compartments (late endosomes and/or lysosomes) of the cell. 10 This disease is ultimately fatal and currently untreatable. Therefore, it is imperative to identify targets for therapeutic intervention in the NPC cells. To work in that direction, there is an urgent need for developing robust high-throughput assays to study multiple genes functionally and follow the cellular phenotypic changes. A recent study by Bartz et al. used RNAi screening on HeLa cells and identified ∼20 genes that could play an important role in cholesterol accumulation and trafficking in these cancer cells. 16 RNAi screening for modulators of cholesterol accumulation in NPC cells would allow for identification of NPC disease-relevant genes.
In this study, we utilized a RNAi-based screening assay in NPC fibroblast coupled with high-content imaging and robust data analysis to identify and functionally validate the role of several NPC disease-related targets. To achieve our goals, we knowledge mined the existing literature on NPC as well as on NPC1 and NPC2 genes and made a comprehensive list of 40 genes that have known association with the disease. The next step involved the development of the assay, which required testing the NPC fibroblast cell lines for transfection efficiency, followed by testing various different time points for imaging plates and several other different parameters. We ran the assay on 2 different NPC fibroblast cell lines as well as on a normal fibroblast cell line. These experiments were followed by choosing the right modules for segmentation of the filipin staining so that the intensity and area values could be converted into numbers to quantitate the change in filipin staining pattern with each individual gene knockdown. For analysis of the data, we used a MATLAB application, which was based on changes in intensity and area of the filipin staining of each cell normalized to the number of cells in each field of data collection. The results for each siRNA treatment were mode shift and percent shift values that correspond to changes in filipin staining (cholesterol accumulation; Fig. 1). This statistical analysis protocol was selected to allow the cells to behave as subpopulations and thus be a better indicator than mean or median intensity values that would not be able to discriminate an overall change in all the cells, or cells in one value mode (ie, low value) being shifted into another value mode (ie, high value). Interestingly, our data showed predominantly unimodal distributions, but as the distributions are not always symmetrical, the “mode” statistics are a better indicator of where the majority of the mass is located than the mean or median. Both statistics “mode shift” and “percent shift” are designed to reflect the shift of cell distribution and should correlate well for these unimodal distributions. Furthermore, we observed that there was not much change in the cell numbers when the cells were treated with all the different individual siRNA indicating that the results were not skewed by lower cell numbers resulting from cellular cytotoxicity. Phenotypic profiling by comparison of the mode shift and percent shift values between the 3 cell lines did show higher correlation between the 2 NPC cell lines GM03123 and GM18453 (0.3–0.4) in comparison with the normal fibroblast cell line GM05659 (Appendix Fig. 6). This indicates that silencing of genes that modulate cholesterol accumulation do so more similarly in NPC cells than in normal fibroblast cells.
Analysis of the screening data of the focused-set siRNA showed that a few siRNA lowered cholesterol staining and many more siRNA increased cholesterol staining compared with control siRNA. Three genes of particular interest were LDLR, which had 3 siRNAs that lowered cholesterol staining, and NPC2 and SMPD1, which had 3 siRNAs that increased cholesterol staining in all the cell lines (Fig. 1). Silencing of LDLR significantly reduced the strong filipin staining of both NPC cell lines GM03123 and GM18453, but it also reduced the moderate filipin staining of the normal fibroblast cell line GM05659. This is expected as LDLR mediates the endocytosis of cholesterol and is in accordance with the previous findings of Zhang et al. where they showed that steroidal LDLR up-regulator GW707 induces accumulation of lysosomal-free cholesterol and inhibits LDL-stimulated cholesterol esterification, similar to that observed in U18666A-treated cells and in Niemann–Pick mutants. 17 Similarly, knockdown of SMPD1 showed increased accumulation of cholesterol in the 3 cell lines, which is in line with the findings that SMPD1 loss-of-function mutations lead to the development of Niemann–Pick disease types A and B. 18 Silencing of NPC2 also showed increase in cholesterol accumulation in not only the wild-type fibroblast cell line, which is expected as NPC2 loss-of-function leads to a disease phenotype, but also in the NPC fibroblasts, which carry NPC1 mutations. This result suggests that knockdown of NPC2, which is known to interact with NPC1 in cholesterol transport, 11 in mutant cells can lead to further disruption of this pathway and thus further cholesterol accumulation.
After extensive data analysis, we chose 9 target genes from our primary screening on 2 different NPC cell lines and a normal fibroblast cell line. These genes included NPC1, NPC2, NPC1L1, LDLR, RAB9A, APOL1, STARD3, STARD4, and SMPD1, which all have been previously associated with cholesterol modulation (Table 2 and references therein). This clearly showed that the assay performed well and was sensitive enough to pick up changes in filipin staining, which directly assesses the changes in cholesterol accumulation. Bartz et al. conducted gene expression studies to identify genes that might be involved in the regulation of cholesterol levels. 16 Their experiments included NPC2 and LDLR as controls, which were also included in our experiments and the data analysis and all the validation experiments clearly confirmed their results. In addition, they also observed that STARD4, which is a cholesterol transporter and binds free cholesterol and increases cholesteryl ester formation, showed up-regulation at all time points studied after sterol depletion, which confirms that STARD4 is involved in maintaining the cholesterol homeostasis. Our results were in concordance to their findings and we observed that when STARD4 is knocked down filipin staining increases thus showing increased cholesterol accumulation in the NPC cells.
Next, we did confirmation screening to test the roles of these genes in cholesterol trafficking and interestingly we observed that the knockdown of LDLR completely shuts down the cholesterol trafficking in our cells (Figs. 5 and 6). To the best of our knowledge, this is the first report on using HT-RNAi technology to identify targets in NPC fibroblast cells.
Our results validated the role of these 9 genes in cholesterol accumulation in NPC cells, thus demonstrating that this protocol can be effectively utilized to do much larger screens to aid in target identification and validation. In addition, we also confirmed that genes NPC1, SMPD1, LDLR, and RAB9A play an important role in cholesterol accumulation and/or trafficking in NPC cells. This work clearly shows the robustness of this technology to conduct screening for the identification of novel genes, which play important roles in cholesterol modulation in NPC and other cholesterol accumulation-associated diseases. Thus, use of this assay could eventually lead to identification of molecular targets in NPC cells that could be instrumental in the development of innovational therapies for individuals afflicted with NPC.
Footnotes
Acknowledgments
We wish to thank William Pavan at the NHGRI, Frederick Maxfield at Cornell University, and Spyro Mousses, Irma Gonzales, Kati Koktavy, Debbie Ries, and Andy Gardner at TGen for their help and support. This work was supported by a grant from the Ara Parseghian Medical Research Foundation.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations
Appendix
A high-content RNAi-screening assay to identify modulators of cholesterol accumulation in Niemann–Pick type C cells.