
Abstract
While sodium-glucose transporter 2 inhibitors (SGLT2i) demonstrate urate-lowering effects, their causal role in Gout prevention remains controversial. This study employs advanced Mendelian randomization (MR) techniques to dissect immune-mediated mechanisms underlying this relationship. Using bidirectional two-sample MR and mediation analysis, we analyzed genetic instrument variables for SGLT2i (10 single-nucleotide polymorphisms, F-statistic >20), Gout risk (6,810 cases/477,788 controls), and 731 immune cell phenotypes. Pleiotropy and heterogeneity were also assessed to ensure robustness. The study confirmed a significant indirect effect of SGLT2i, which exhibited a 2.6% reduced Gout risk (Odds Ratio [OR]: 0.9738, 95% confidence interval [CI] = 0.9623, 0.9854, P = 1.12e-05). Thirty-five immune cell phenotypes were identified as significantly affecting Gout development, with key phenotypes such as CD86 on myeloid Dendritic cell (DC) (OR: 0.9966; 95% CI = 0.9930, 0.9995), contributing to 12.8% of the overall mediation effect. No evidence of heterogeneity or pleiotropy was detected and reverse-direction MR corroborated these findings. Our study first established SGLT2i as Gout-protective agents through DC-mediated immunomodulation, offering mechanistic insights for targeted prevention strategies in clinical practice.
Keywords
BACKGROUND
Sodium-glucose transporter 2 inhibitors (SGLT2i) are a novel class of antidiabetic drugs that inhibit SGLT2 in the renal tubules, thereby reducing the reabsorption of free glucose and enhancing glucose excretion in the urine. 1 Recent studies have found that in addition to lowering blood glucose levels, SGLT2i have the potential to reduce serum uric acid levels and decrease the risk of Gout attacks.2,3 However, the precise mechanisms by which SGLT2i provide anti-Gout benefits and improve clinical outcomes remain unclear, and are subject to ongoing research and debate.4,5 Gout is an inflammatory disease caused by the deposition of urate crystals in joints and other tissues. It is closely associated with immune cell function. 6 Neutrophils and monocytes are considered the primary cell types involved in Gout’s uric acid crystal-induced inflammation. 7 According to estimates, there were approximately 55.8 million individuals with Gout globally in 2020, with the prevalence in males being 3.26 times higher than in females. The number of cases is projected to increase to 95.8 million by 2050. 8 Gout not only affects the quality of life of patients, but also imposes a significant economic burden on health.
Mendelian randomization (MR) is a statistical method that employs genetic instrumental variables (IVs), typically single-nucleotide polymorphisms (SNPs), to infer potential causal relationships from nonexperimental data in a manner analogous to treatment allocation in randomized controlled trials. 9 This strategy enhances the credibility of the causal inferences in observational studies. MR mediation analysis uses genetic variants as IVs to assess the impact of exposure factors on disease risk via specific biological pathways. The value of mediation analysis lies in its ability to elucidate the mechanisms through which independent variables influence dependent variables via intermediary factors, thus enhancing our understanding of causal pathways in research. 10
To investigate the association between SGLT2i and the risk of Gout, we employed a bidirectional, two-sample MR approach to evaluate this relationship and used a two-step MR analysis with 731 types of immune cells as mediators to explore potential pathways through which SGLT2i affect Gout risk. By representing the therapeutic effect of SGLT2i with genetic variations and integrating large-scale genome-wide association study (GWAS) data, we assessed the association between these variants and Gout, revealing the mechanisms through which SGLT2i may regulate immune pathways in the prevention and treatment of Gout. 11 This study aimed to elucidate the potential mechanisms of SGLT2i in Gout prevention and treatment, with a genetic focus on the pathways by which they affect Gout risk by modulating inflammatory responses. This study not only provides new genetic evidence for the use of SGLT2i in Gout treatment but also contributes to an in-depth understanding of their mechanism of action and provides theoretical support for clinical practice. Despite observational evidence linking SGLT2i to reduced Gout risk, conflicting results persist due to residual confounding (e.g., concurrent diuretic use) and short-term follow-up in clinical trials. MR, by leveraging lifelong genetic exposure to SGLT2i, offers a unique opportunity to clarify these relationships while identifying mediating pathways.
MATERIALS AND METHODS
Study Methodology
In this study, we first employed a two-sample MR approach using SGLT2i as the exposure factor and Gout as the outcome derived from the latest research. 12 In the second step, we employed a two-step mediation MR method to assess summary data from GWAS of 731 immune cells, which included counts of leukocytes, eosinophils, basophils, neutrophils, lymphocytes, and monocytes as mediators in the analysis (Table 1). To ensure the reliability of the MR analysis, three assumptions had to be satisfied: (1) There is a robust association between genetic variants and the exposure factor; (2) genetic variants must be independent of confounding factors; and (3) the impact of genetic variants on the outcome is mediated solely through the exposure factor, with no alternative pathways. This analytical framework aids in a more accurate understanding of the genetic mechanisms by which SGLT2i influence Gout. This study adhered to the STROBE-MR guidelines. 13 Figure 1 outlines the workflow of this study. In our MR analysis, we utilized only publicly available datasets, thereby negating the need for additional ethical approval. For detailed information on the ethical approval and participant consent for each GWAS, please refer to the original publications.
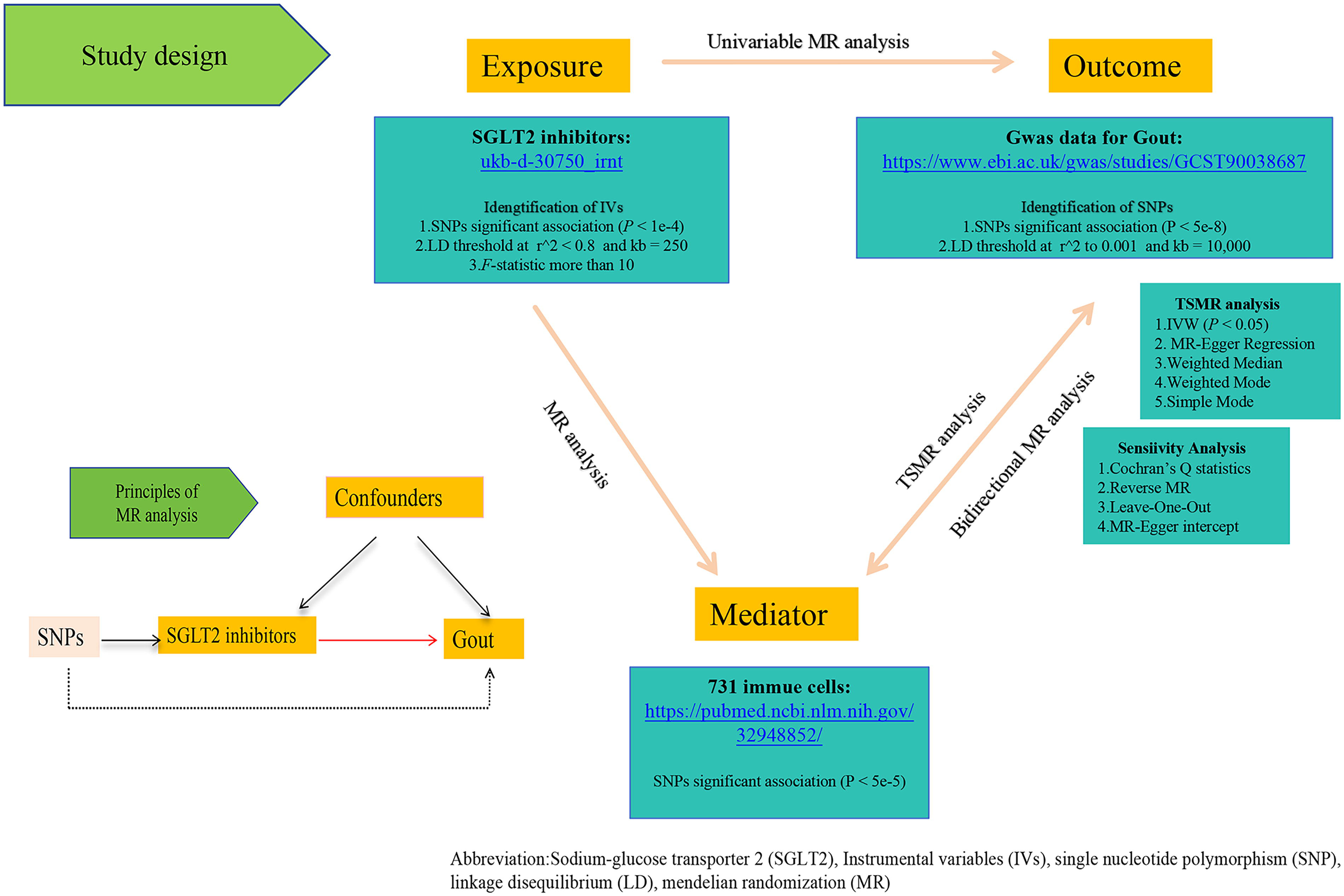
The framework of this study.
Dataset Utilized in Our Study
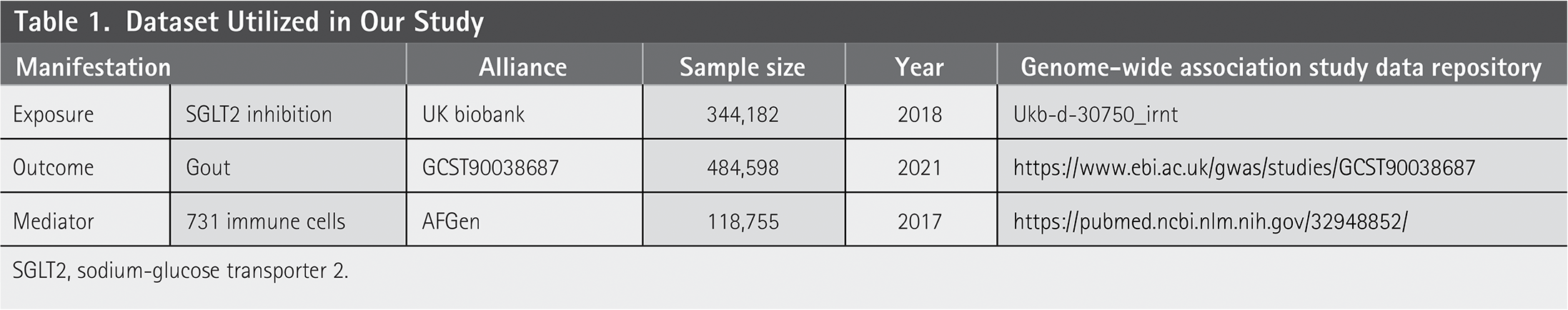
SGLT2, sodium-glucose transporter 2.
Genetic Source of SGLT2i
We identified genetic variants linked to SLC5A2 (SGLT2) expression from the Genotype-Tisssue Expression (GTEx v8 kidney cortex) and eQTLGen consortium. SLC5A2-mRNA expression data and expression quantitative trait loci (eQTL) data were derived from the GTEx and eQTLGen databases. GTEx provides gene expression data across various tissues and associated genetic variants, 14 whereas eQTLGen offers extensive eQTL analyses. 15 These variants, which are associated with Hemoglobin A1c levels, a marker for diabetes management, were selected based on a significant association (P < 1e-4) from the GWAS data of 344,182 nondiabetic Europeans. The data preprocessing method was sourced from the literature. 16 We set the linkage disequilibrium (LD) threshold at (r2 < 0.8) and limited the genetic distance within 250 kb. We estimated the strength of IVs using the F-statistic, where F < 10 indicates weak instrument bias. The F-statistic was calculated as F = [(n − k − 1)/k] * [R2/(1−R2)], where R2 = 2 × EAF × (1−EAF) × 2. Here, EAF represents the effect allele frequency, n is the sample size, k denotes the number of IVs, and β2 quantifies the effect 17 (see Supplementary Table S1 for details on the IVs for causality).
Outcome Data for Gout
In this study, the GWAS of Gout was derived from the GCST90038687 dataset, which utilizes genome-wide genotyping array technology. The study included 6,810 Gout cases and 477,788 controls of European descent, representing the largest scale of GWAS meta-analysis to date for this condition. 18 We selected outcome SNPs related to Gout using a significance threshold (P < 5e-8). We set the LD parameter (r2) to 0.001 and the genetic distance to 10,000 kb to ensure independence among SNPs, thereby eliminating interference due to LD. 16 The findings were reported as odds ratios (ORs) accompanied by their respective 95% confidence intervals (CIs).
GWAS Data Sources for Mediation Analysis
Data of mediation MR analysis was obtained from the GCST0001391 to GCST0002121 dataset in the GWAS repository encompassed 731 immunophenotypes analyzed by flow cytometry, including absolute cell counts (118), median fluorescence intensity (389), morphological parameters (32), and relative cell counts (192) with P < 5e-5 (https://gwas.mrcieu.ac.uk/datasets/). A cluster analysis was conducted. The Sardinian GWAS dataset included 3,757 cases and 3,027 controls of European descent, aged 18–102, with 43% male and 57% female participants. 19
Data Quality Control and Exclusion Criteria
To minimize potential biases and ensure data quality, we applied the following exclusion criteria across all datasets: palindromic and ambiguous SNPs were excluded. SNPs with a minor allele frequency >0.42 were removed to avoid strand ambiguity. Phenotypes showing horizontal pleiotropy were excluded based on MR-Egger intercept P-values <0.05. Immune cell datasets with >10% missing flow cytometry parameters were also excluded. 20
Univariable and Two-Step MR Analysis
Univariable MR analysis
Initially, we conducted a univariate MR analysis to assess the direct impact of SGLT2i on Gout. This analysis used genetic variants strongly associated with the use of SGLT2i as IVs, which helped to ensure a robust understanding of the direct link between SGLT2i and Gout.
Bidirectional and Two-Sample MR Analysis
Subsequently, we implemented a bidirectional two-sample MR approach to estimate the potential effects of Gout on 731 immune cell phenotypes. For immune cell phenotypes that demonstrated statistically significant associations with SGLT2i, we conducted two-sample MR analysis to assess their potential effects on Gout.
Mediation Analysis
To quantify the mediating role of immune cells in the causal pathway between SGLT2i and Gout, we employed a two-step MR mediation framework. 21 The analysis proceeded as follows:
① Total effect estimation
The total effect (β_total) of SGLT2i on Gout was first estimated using univariable MR analysis, as described in the “Univariable MR Analysis” section.
② Mediation effect decomposition
Step 1: We assessed the causal effect of SGLT2i on candidate immune cell phenotypes (mediators) using MR analysis (β_exposure→mediator).
Step 2: For mediators showing significant associations with SGLT2i (P < 0.05), we further evaluated their causal effects on Gout (β_mediator→outcome).
③ Indirect effect calculation
The indirect effect (β_indirect) mediated through immune cells was calculated as the product of the two estimates:
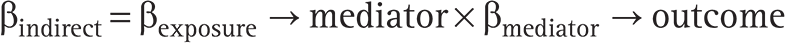
The proportion of the total effect mediated by immune cells was derived using the Delta method:
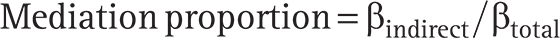
The Delta method was also applied to estimate the standard error of the mediation proportion, enabling hypothesis testing and confidence interval construction.
④ Sensitivity adjustment
To account for multiple testing across 731 immune phenotypes, we applied the Benjamini-Hochberg procedure to control the false discovery rate (FDR), with statistical significance defined as PFDR < 0.05. 22
MR Analysis
To predict the effect of SGLT2i on Gout, we used five MR methods: IVW, MR-Egger, Weighted Median, Weighted mode, and Simple mode. Our primary method, IVW, is effective if all SNPs are valid IVs. MR-Egger remains valid even with some invalid IVs but is imprecise. 9 The weighted Median is robust against pleiotropic effects and superior to the MR-Egger method in terms of error rate and power. Causality was confirmed if it was consistent across methods and significant at P < 0.05. Consistent with prior research, a PFDR value below 0.05, indicating a significant causal association, beta coefficients, and standard errors were obtained from the GWAS data on Gout, and any ambiguous SNPs were either adjusted or removed. We conducted two-step, two-sample MR analyses using R (version 4.2.2, Vienna, Austria, 2024-05-11) and “TwoSampleMR” packages.
Reverse MR Analysis
Before further study, we validated the directional judgment of the causal effects. We conducted reverse MR analysis of the 731 immune cells to detect Gout.
Sensitivity Analysis
To maintain the integrity of the MR analysis, we conducted a sensitivity analysis using several methods to detect heterogeneity and pleiotropy. These included the Cochran’s Q test and the MR-Egger intercept. A significant Cochran Q test (P < 0.05) indicated heterogeneity and the MR-Egger intercept evaluated the instrumental variable validity for horizontal pleiotropy impacts. To address outliers and influential data points, we implemented three distinct methods to assess the robustness of our MR study results. Initially, leave-one-out analysis: Iteratively removing each SNP to assess its impact on effect estimates; MR-PRESSO: Detecting outliers via global heterogeneity tests (P < 0.05); Concurrently, scatter plot analysis facilitates a visual assessment of the relationship between the strength of the instrumental variables and the magnitude of their effects; Furthermore, funnel plot analysis was employed to detect potential biases such as publication bias or genetic heterogeneity. All statistical results retain original computational precision.
RESULT
Effects of SGLT2i on Gout Outcomes
We selected 10 independent SNPs as IVs for SGLT2i, and each SNP exhibited an F-statistic exceeding 10 (Supplementary Table S2). Our analysis revealed a notable link between SGLT2i and lowered risk of Gout (OR: 0.9738; 95% CI = 0.9622–0.9854, P = 1.12e-05, β = −0.027). Using the Steiger test, we further validated that these SNPs predominantly affect SGLT2i rather than the risk of Gout, thereby ensuring correct directional causality. The extremely low p-value of 4.705969e-23 strongly supports the validity of this conclusion. Sensitivity analyses confirmed the absence of horizontal pleiotropy (PEgger intercept = 0.3513) and heterogeneity (PQ = 0.9111), see Figure 2 and Supplementary Tables S3 and S4 for details.
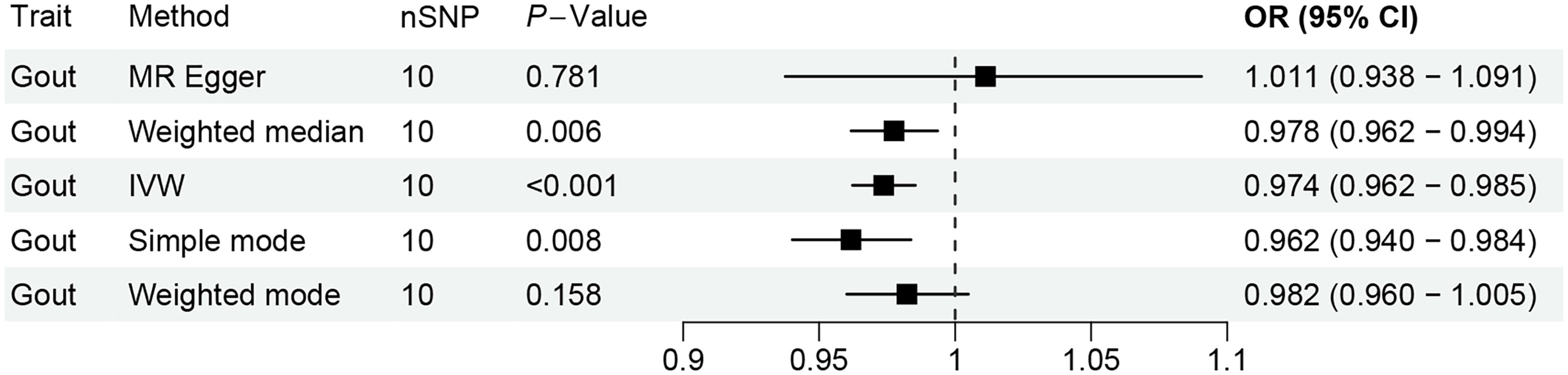
MR analysis of SGLT2i on Gout. MR, Mendelian randomization; SGLT2i, sodium-glucose transporter 2 inhibitors.
Genetic Influence of Immune Cells on Gout
We conducted an extensive statistical analysis to explore how immune cell phenotypes might influence the development of Gout (Figure 3). To ensure the accuracy of our findings, we applied FDR corrections to the IVW results and identified factors with PVAL (FDR) greater than 0.05. A thorough review of the data revealed that 35 immune cell phenotypes significantly impacted the risk of developing Gout, with PFDR less than 0.05, as demonstrated using the IVW method. These phenotypic traits included six Absolute Counts (AC), 13 MFI, two MP, and 14 Relative Counts (RC). Among these immune cells, 14 were associated with an increased risk of Gout, while 11 served as protective factors against Gout (Supplementary Table S5). Additionally, the reverse MR analysis did not reveal any positive findings.
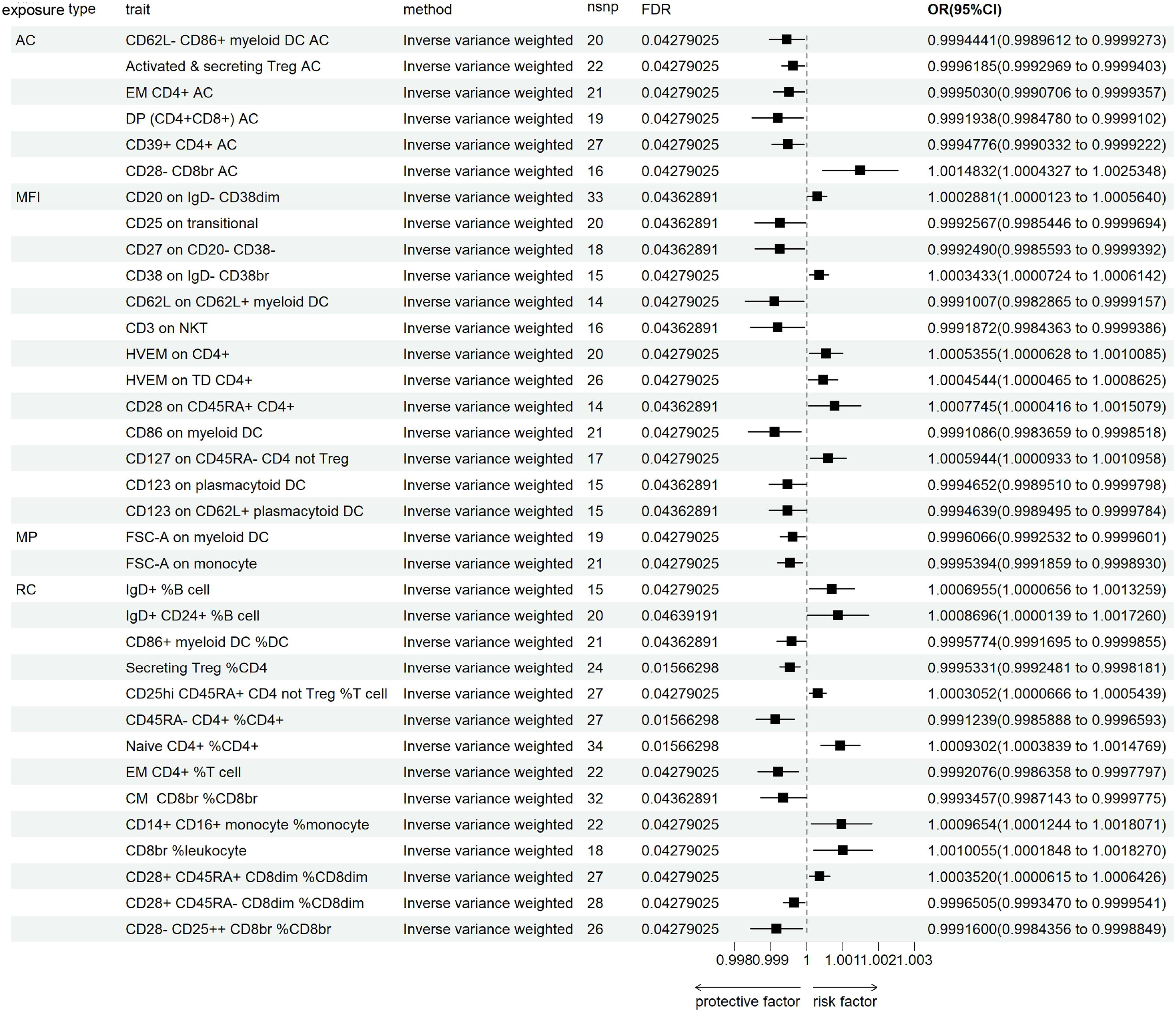
MR analysis of SGLT2i on Gout.
In B cells, the MFI phenotypes of CD25 on transitional and CD27 on CD20− CD38− were associated with a reduced risk of Gout, whereas all CD38 phenotypes were negatively correlated with the progression of Gout. Among the 10 immune phenotypes in Treg cells, two were related to MFI, five to RC, and three to AC, with CD28 being the most common phenotype (50%). In addition to CD28+ CD45RA− CD8dim %CD8dim and CD28− CD25++ CD8br %CD8br, which lowered the risk of Gout, other CD28-related phenotypes increased the risk of developing Gout. In the maturation stages of T cells, the phenotypes CD45RA− CD4+ %CD4+, Effector Memory (EM) CD4+ AC, EM CD4+ T cells, and Central Memory (CM) CD8br %CD8br were associated with a reduced risk of Gout, while the remaining four phenotypes increased the risk. In conventional Dendritic Cells (cDC), of the eight immune phenotypes, four were related to MFI, one to RC, one to AC, and two to MP, with the most common marker being myeloid DC (mDC) (50%). Including the MFI phenotype of CD86 in mDC, all cDC cell phenotypes exhibited a protective effect against Gout. The causal relationship between CD86 on mDC and Gout was illustrated in Supplementary Figure S1, including funnel plot, forest plot, scatter plot, and leave-one-out analysis plot. These visualizations provided an intuitive representation of the potential causal associations between CD86 on mDC and Gout, offering data support for further mechanistic investigations. The sensitivity analyses did not demonstrate any significant pleiotropy. Heterogeneity was not observed among the 32 immune cell phenotypes, see detail in Supplementary Tables S6 and S7).
Mediation Analysis of the Causal Relationship between SGLT2i and Gout
This study confirmed an independent causal relationship between SGLT2i and Gout. Previous research has linked SGLT2i and immune cells to Gout. Following the principles of the two-step MR mediation analysis, we investigated whether specific immune cell phenotypes mediate the effect of SGLT2i on Gout. First, we established the total effect (β). To analyze the effect of SGLT2i on immune cell phenotypes, we primarily used the IVW method to adjust for genetic effects (β2), as detailed in Supplementary Table S8. Our study indicates that SGLT2i have a significant causal role in modulating immune cell levels. Specifically, SGLT2i significantly downregulated monocyte levels and upregulated DC and Treg cells. Specifically, SGLT2i significantly downregulated monocyte levels (CD14+ CD16+ monocyte: β = −2.1123, P = 0.0025) and upregulated DC activation (CD86 on mDCs: β = 3.8190, P = 7.8768e-06) and regulatory T cell subsets (activated & secreting CD4+ Treg: β = 2.6753, P = 0.0002). These findings provide a crucial molecular basis for the role of SGLT2i in immune response regulation and their potential anti-inflammatory effects. Additionally, we calculated the indirect mediation effect of SGLT2i on Gout outcomes, derived by assessing the mediator’s influence on the outcome and the impact of exposure on the mediator, using the delta method for effect ratio computation. We identified the mediation effect of CD86 on mDC (OR: 0.9966; 95% CI = 0.9930, 0.9995) accounted for 12.8% of the total effect, indicating a statistically significant association with Gout risk, even after excluding the direct effects of SGLT2i (Figure 4 and Supplementary Table S9). This suggests that SGLT2i may reduce the risk of Gout by modulating these immune cell phenotypes. Furthermore, to explore the consistency and robustness of these findings, we assessed horizontal pleiotropy and heterogeneity in the instrumental variables used. These results are presented in Supplementary Tables S10 and S11, which provide a detailed analysis of the stability and reliability of the associations observed. This supports the potential mediating role of SGLT2i in the onset of Gout, offering new insights into the mechanisms by which SGLT2i affect immune function and Gout risk.
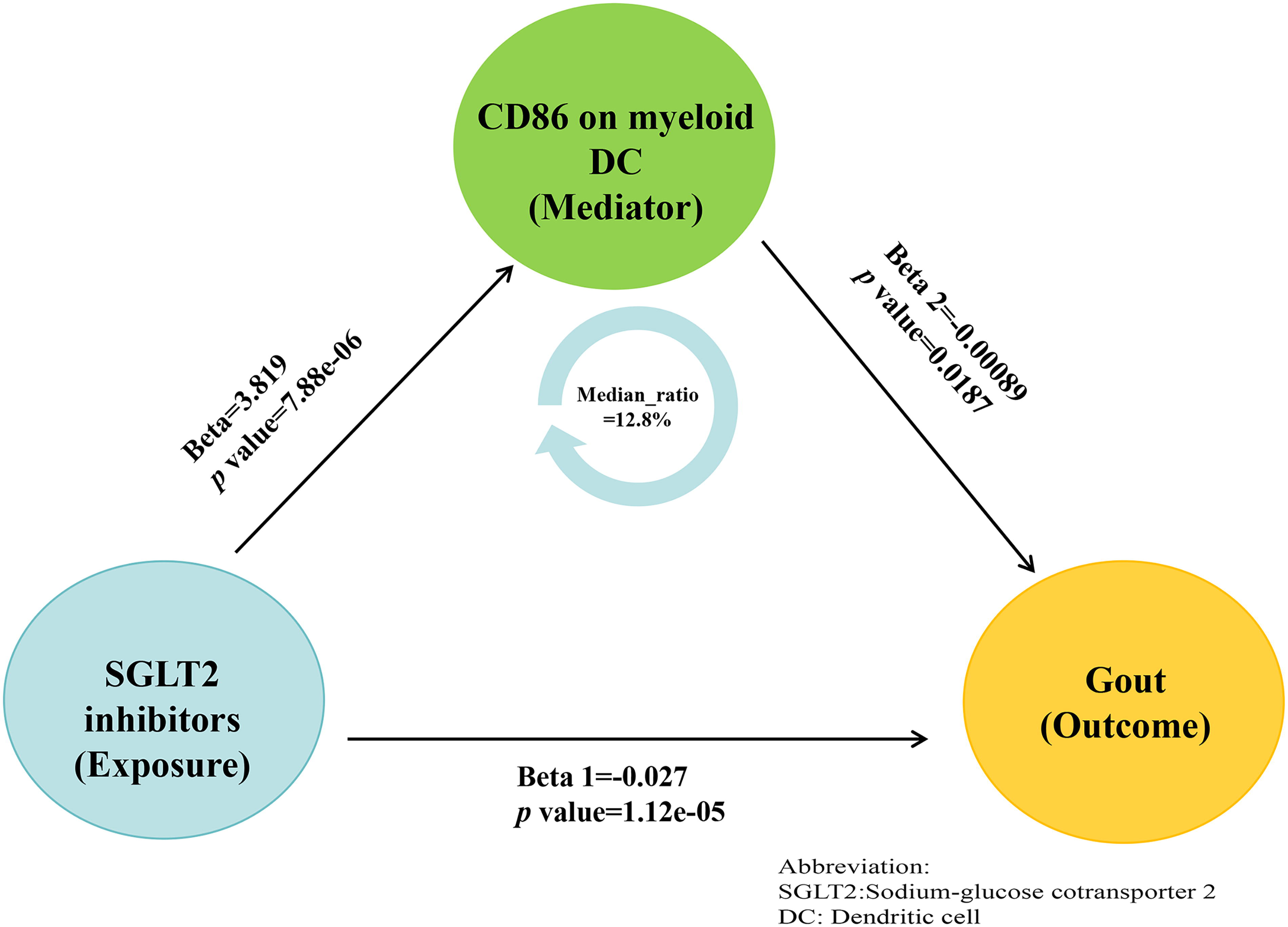
Mediation analysis with causal link between SGLT2i and Gout.
DISCUSSION
In our study, utilizing a two-step approach and bidirectional two-sample MR technique, we first observed a significant negative causal effect of SGLT2i on Gout, revealing their potential protective role in reducing Gout risk. Further bidirectional MR and mediation effect analyses indicated that SGLT2i may reduce the risk of Gout by modulating specific immune cell phenotypes. Specifically, CD86 on mDCs, accounting for approximately 12.8% of the association between SGLT2i and Gout risk, suggests that SGLT2i may exert significant indirect effects on Gout risk through the activation and secretion of mediating mDCs. Moreover, the application of reverse MR and sensitivity analyses further strengthened the robustness and reliability of our findings. This mediation MR study provides strong scientific evidence for the role of SGLT2i in the prevention of Gout.
Several studies have shown that SGLT2i users have a reduced risk of Gout by 11%–61%.23–25 For instance, studies in Taiwan 5 and by the U.S. Veterans Affairs 26 support these findings, demonstrating that SGLT2i significantly reduce the incidence of Gout. SGLT2i significantly reduce the risk of Gout in patients with type 2 diabetes and/or heart failure. 27 These results support the potential of SGLT2i in reducing Gout incidence. Further analysis indicates that SGLT2i reduce the occurrence of Gout by significantly lowering blood uric acid levels, a mechanism likely related to their inhibition of the renal tubule urate transporter 1, promotion of uric acid excretion, and anti-inflammatory effects. 28 However, despite some mechanisms being elucidated, the specific immunological mechanisms remain incompletely understood, requiring further research for validation.
However, existing studies are not without contradictions. Some observational studies show no significant effect of SGLT2i on Gout risk. 4 Additionally, there remains controversy over whether SGLT2i independently lower uric acid regardless of renal function.29,30 These discrepancies may arise from differences in study design, patient populations, and confounding factors such as comorbidities and concomitant medications. The impact of SGLT2i compared with other second-line diabetes medications (such as glucagon-like peptide 1 receptor agonists31,32 and dipeptidyl peptidase 4 inhibitors5,33,34) on Gout risk also varies. These differences suggest that SGLT2i may have unique mechanisms of action, offering advantages in Gout prevention. Our analysis using large-scale GWAS data confirms a significant negative causal relationship between SGLT2i and Gout, aligning with these clinical observations. However, these studies have not fully addressed the complexity of immune cell involvement in Gout or how SGLT2i reduce Gout risk and improve prognosis, which requires further clarification.
SGLT2i may primarily affect Gout-related inflammatory pathways through the following mechanisms. First, they promote the polarization of M2 macrophages, thereby reducing the activity and number of pro-inflammatory M1 macrophages, which is crucial for anti-inflammatory effects and tissue protection. 35 Second, SGLT2i can inhibit the activation of the NOD-like receptor family pyrin domain containing 3 (NLRP3) inflammasome, thereby reducing the release of pro-inflammatory cytokines (such as interleukin-1 beta [IL-1β]), which play a vital role in the inflammatory pathways of Gout.5,36,37 Additionally, SGLT2i activate AMP-activated protein kinase (AMPK), enhancing the function of M2 macrophages, further alleviating inflammation and promoting cellular energy balance and anti-inflammatory pathways. 38 In the context of Gout, the number of regulatory T cells (Tregs) decreases, and enhancing Treg function helps suppress excessive inflammatory responses. 39 During acute Gout attacks, genes associated with inflammation and cytokines in classical monocytes are generally upregulated systemically. 40 Our study indicates that SGLT2i play a significant role in controlling the immune response in Gout, particularly in regulating the maturation and function of immune cells, especially DCs and Tregs. SGLT2i may modulate the immune response in Gout by affecting the maturation, activity, and surface markers of these immune cells. Specifically, DCs and Tregs. SGLT2i significantly reverse changes in the maturation of memory CD4+ T cells, the function of Tregs, alterations in B cell surface markers, and the numbers of monocytes and effector T cells. These immunological changes suggest that SGLT2i counteract Gout by modulating the characteristics of immune cells, further supporting their potential as immunomodulatory treatments. Our findings not only complement existing literature but also further highlight the therapeutic significance of SGLT2i in immune modulation. Specifically, our study suggests that these inhibitors exert protective effects against Gout by downregulating CD86 on mDCs and Tregs, while upregulating monocyte levels.
CD86, a key co-stimulatory molecule on DCs, regulates T helper cell 1 (Th1)/T helper cell 17 (Th17) inflammatory responses by binding to the CD28/cytotoxic T-lymphocyte-associated protein 4 receptors on T cells. 41 Studies have shown that macrophages and mDCs are the predominant cells in the synovial fluid of Gout patients, and under hyperuricemia or urate crystal stimulation, they drive the release of pro-inflammatory factors such as IL-1β and IL-6, leading to interactions between immune cells in blood and synovial cells, ultimately triggering Gout. 42 The expression level of CD86 is influenced by the metabolic environment, for example, hyperglycemia may enhance the antigen-presenting capacity of mDCs, further promoting CD86-mediated inflammation. 43 Aragón-Herrera et al. demonstrated through untargeted metabolomics that empagliflozin reduces CD86 expression in visceral fat in Zucker rats, modulating lipid metabolism and inflammatory responses. 44 SGLT2i may suppress inflammation in Gout by modulating CD86 expression, thereby exerting their therapeutic effects. 45 Furthermore, SGLT2i also demonstrates some efficacy in lowering uric acid levels, which may further help alleviate Gout symptoms. 46 Therefore, CD86 on mDC may serve as a mediator for the negative regulation of Gout by SGLT2i.
Our study highlights SGLT2i as a promising strategy for Gout prevention and management. By modulating immune cell, SGLT2i significantly reduce Gout risk. CD86 on mDC, a key mediator, accounts for 12.8% of this protective effect, suggesting SGLT2i target immune pathways to exert anti-inflammatory benefits. Clinically, SGLT2i should be prioritized in diabetic patients with hyperuricemia or early Gout, with CD86 as a potential biomarker for treatment response. Future trials should explore combining SGLT2i with mDC-targeted therapies (e.g., CD86 blockade) to enhance anti-inflammatory efficacy, potentially transforming Gout treatment through metabolic and immune modulation.
In contrast to observational studies, this research employed a bidirectional, two-step MR analysis to reduce bias from confounding factors and reverse causality, and sensitivity analysis was performed to verify the robustness of the results. Additionally, further mediation analysis was conducted to determine the proportion of mediation by immune cells. While this study provides new insights into the interactions between SGLT2i, immune cells, and Gout, it has the following limitations: (1) Representativeness of genetic variants: We used genetic variants associated with SGLT2i to model the long-term exposure effects. While this may not fully reflect the short-term treatment impact, it provides effective evidence for determining causal direction and represents the best practice under current conditions. (2) Limitations related to ethnicity and population: This study is primarily based on data from individuals of European descent, which may limit the generalizability of the results. We acknowledge this limitation and encourage future research to extend to more diverse ethnicities and populations. (3) Assumptions of genetic instrumental variables: Although MR analysis relies on stringent assumptions, we minimized confounding effects with carefully selected genetic variants. However, the presence of undetected confounders cannot be entirely ruled out. To verify and expand our findings, future studies should include data from broader ethnic and population groups, and incorporate laboratory and clinical trials, particularly those directly manipulating immune cell levels, to further explore and confirm these observations. To further validate these findings, future research could simulate the interaction between immune cells and SGLT2i through in vitro experiments, particularly using cell culture models to test immune cell function, in order to support the causal inference of this study and explore in greater depth the role of SGLT2i and immune cells in Gout. Finally, the initial study did not provide key information such as age and gender, factors closely related to Gout incidence. The lack of these data limits the in-depth analysis of different subgroups.
CONCLUSION
This study provides genetic evidence supporting the relationship between SGLT2i, immune cells, and Gout. Specifically, CD86 on mDC may play a critical role in mediating the protective effect of SGLT2i in reducing Gout.
AUTHORS’ CONTRIBUTIONS
H.Z., W.L., Y.Z., J.L., and Z.C. contributed to conceptualization. Z.C. and J.L. performed formal analysis. All authors contributed to methodology. Z.C., Y.Z., Z.C., and J.L. managed project administration. All authors provided resources. H.Z. and J.L. conducted data analysis and software. H.Z., J.L., and Z.C. conducted validation and visualization. Z.C. and Y.Z. provided supervision. Z.C., W.L., J.L., and Z.C. contributed to writing—original draft. H.Z. and Y.Z. were responsible for writing—review and editing. All authors were responsible for data curation and investigation. All authors have read and approved the final article.
Footnotes
FUNDING INFORMATION
This work was financially supported in part by research grants from
ETHICS APPROVAL AND CONSENT TO PARTICIPATE
This study, approved by local GWAS and conducted using aggregated data from an online platform, received ethical approval from local committees. Participants provided written consent. We adhered to local and institutional regulations on legality and ethics. National regulations exempted explicit written consent from participants or their next of kin. Our procedures strictly followed laws on privacy and data protection, upholding participants’ rights and ethical standards, eliminating the need for additional ethics approval.
AVAILABILITY OF DATA AND MATERIALS
DISCLOSURE STATEMENT
The authors declare that they have no competing interests.