
Abstract
Δ9-Tetrahydrocannabinol (Δ9-THC), the primary psychoactive component in marijuana, is FDA approved to ameliorate AIDS-associated wasting. Because cannabinoid receptors are expressed on cells of the immune system, chronic Δ9-THC use may impact HIV disease progression. We examined the impact of chronic Δ9-THC administration (0.32 mg/kg im, 2 × daily), starting 28 days prior to inoculation with simian immunodeficiency virus (SIVmac251; 100 TCID50/ml, iv), on immune and metabolic indicators of disease during the initial 6 month asymptomatic phase of infection in rhesus macaques. SIVmac251 inoculation resulted in measurable viral load, decreased lymphocyte CD4+/CD8+ ratio, and increased CD8+ proliferation. Δ9-THC treatment of SIV-infected animals produced minor to no effects in these parameters. However, chronic Δ9-THC administration decreased early mortality from SIV infection (p = 0.039), and this was associated with attenuation of plasma and CSF viral load and retention of body mass (p = NS). In vitro, Δ9-THC (10 μm) decreased SIV (10 TCID50) viral replication in MT4-R5 cells. These results indicate that chronic Δ9-THC does not increase viral load or aggravate morbidity and may actually ameliorate SIV disease progression. We speculate that reduced levels of SIV, retention of body mass, and attenuation of inflammation are likely mechanisms for Δ9-THC-mediated modulation of disease progression that warrant further study.
T
Using a well-established nonhuman primate model of HIV disease, we examined the impact of chronic intramuscular Δ9-THC (provided by the National Institute on Drug Abuse, Research Technical Branch, Rockville, MD) administration on the early phase of simian immunodeficiency virus (SIV) infection in age-matched (4–6 years old) and body weight-matched healthy male Indian-derived rhesus macaques. Chronic administration of Δ9-THC [or 0.05 ml/kg vehicle (VEH)] was initiated prior to SIV with 0.18 mg/kg, a dose that eliminated responding in a complex operant behavioral task in almost all of the subjects. The dose was subsequently increased for each subject to 0.32 mg/kg, over a period of approximately 2 weeks, and maintained for the duration of the study. Almost all of the animals demonstrated tolerance to the behavioral effects of this dose prior to SIV inoculation (reported elsewhere). 6 Approximately 1 month after initiating chronic Δ9-THC administration, eight animals were inoculated intravenously with 100 times the TCID50 (50% tissue culture infective dose) of SIVMAC251. The pathogenicity of this SIV isolate is similar to HIV. 7 The progression of SIV disease was monitored through clinical and biochemical parameters, and viral load in plasma and cerebrospinal fluid (CSF).
Contrary to what we expected, chronic cannabinoid administration did not significantly increase viral load or exacerbate immune dysfunction. Mean viral load in VEH/SIV+ animals was 6.56 ± 0.16 log copies at 7 days and 6.04 ± 0.6 log copies/ml at 2 months postinfection (Fig. 1A). Viral load remained elevated (6.35 ± 0.76 and 5.8 ± 0.06 log copies/ml at 3 and 6 months, respectively) throughout the duration of the study. Viral load in THC/SIV+ was lower (4.83 ± 0.56 log copies/ml) as compared to VEH/SIV+ beginning at 2 months post-SIV infection (p = NS). Viral mRNA in CSF peaked at day 14 postinfection and remained elevated up to 2 months postinfection in the VEH/SIV+ and THC/SIV+ animals (Fig. 1B). Thereafter, CSF viral load fluctuated between levels below the limit of detection and low counts in both SIV-infected groups. Across all time points, 60% of VEH/SIV+ CSF samples were above the limit of detection for the assay. In contrast, only 40% of CSF samples from THC/SIV+ animals had viral loads above the limit of detection.
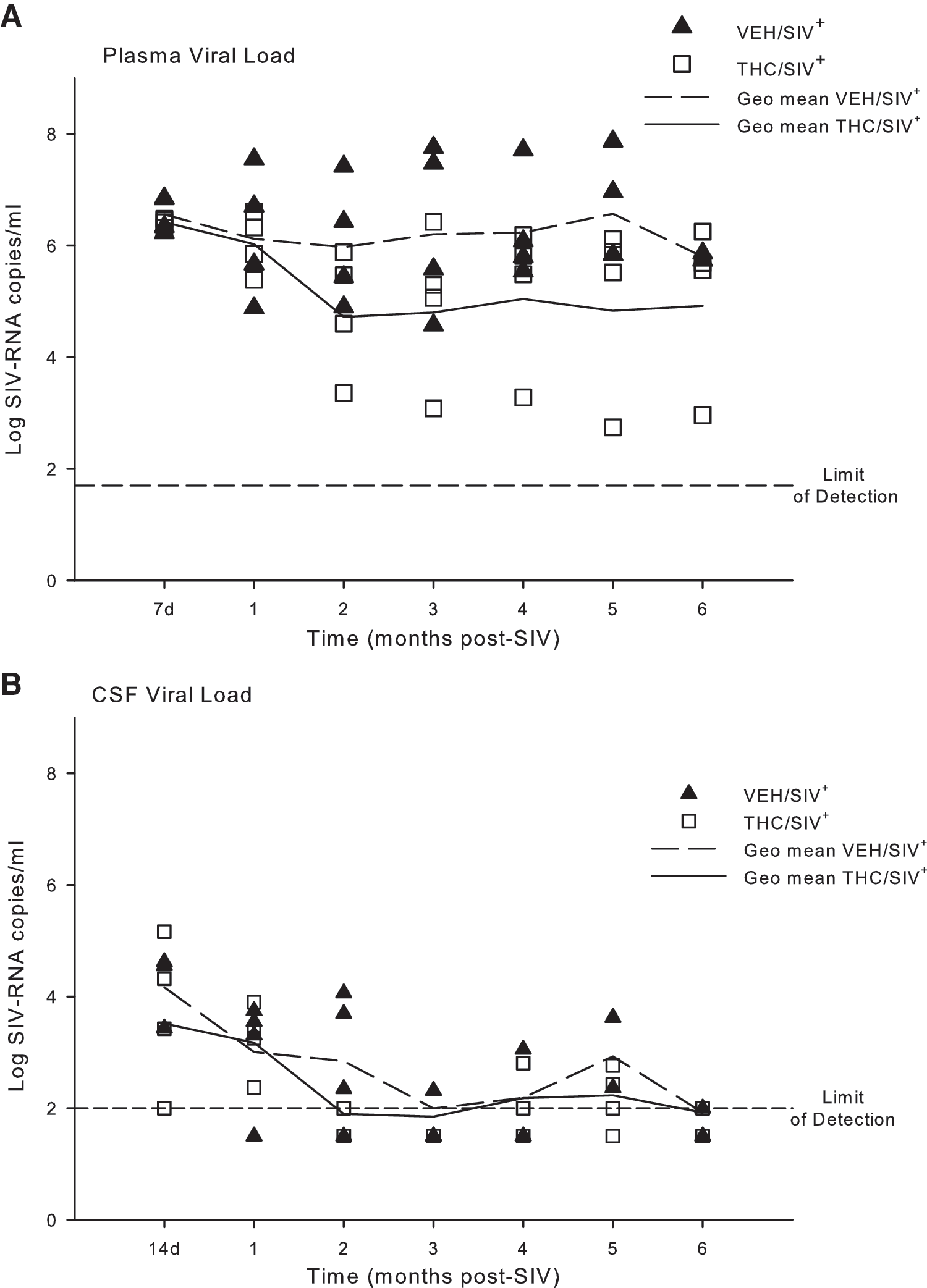
Viral load detected in
Complete and differential blood counts were performed using a Beckman Coulter LH755 for total leukocyte counts and Wright–Giemsa staining of blood smears for leukocyte differentials. Blood lymphocyte subsets were determined by flow cytometry as previously described. 8 The CD4+ to CD8+ ratio decreased significantly from preinfection values (p = 0.0001) in both VEH- and THC-treated animals (Fig. 2B). The marked decrease in CD4+ lymphocyte counts reached statistical significance (p = 0.023) at 0.5 and 1 months post-SIV infection in VEH/SIV+ animals, and beginning at 2 months in THC/SIV+ (p = 0.037). In contrast, CD8+ counts showed a short-lived increase at 2 weeks postinfection, which reached statistical significance only in THC/SIV+ animals (p = 0.029) (Fig. 2D). CD4+ and CD8+ lymphocyte expressions of Ki67 and caspase 3 (Cas3) were measured as indices of proliferation and apoptosis. No significant changes were noted in the percent of either Ki67+ or Cas3+ on CD4+ lymphocytes in either of the experimental groups (data not shown). In contrast, both markers were increased significantly in CD8+ lymphocytes at 2 weeks postinfection, suggesting increased activation of these cells during the acute response to infection (Fig. 3A). The CD8+ Ki67 expression was increased 10-fold at this time point, and was significantly higher in THC/SIV+ animals when compared to the VEH/SIV+-infected animals (p = 0.049). This rise in CD8+ lymphocyte proliferation was transient in both groups with only values for THC/SIV+ animals remaining higher (p = 0.05) than controls at 1 month post-SIV infection. Values for the VEH/SIV+ and THC/SIV+ groups were not different from control beyond 1 month postinfection. The changes in the number of Ki67+ CD8+ lymphocytes were paralleled by those in number of Cas3+ CD8+ lymphocytes. Cas3+ CD8+ lymphocytes showed a transient and significant (p = 0.001) 7- to 8-fold increase that peaked at 2 weeks post-SIV infection in both VEH/SIV+ and THC/SIV+ animals (Fig. 3B). Although a similar pattern of higher levels of Cas3+ CD8+ lymphocytes was observed in the THC/SIV+ animals, this difference failed to reach statistical significance in comparison to the VEH/SIV+ animals (p = 0.06). This rise in CD8+ lymphocyte apoptosis was short-lived, and values after 1 month post-SIV infection were not different from uninfected control values.
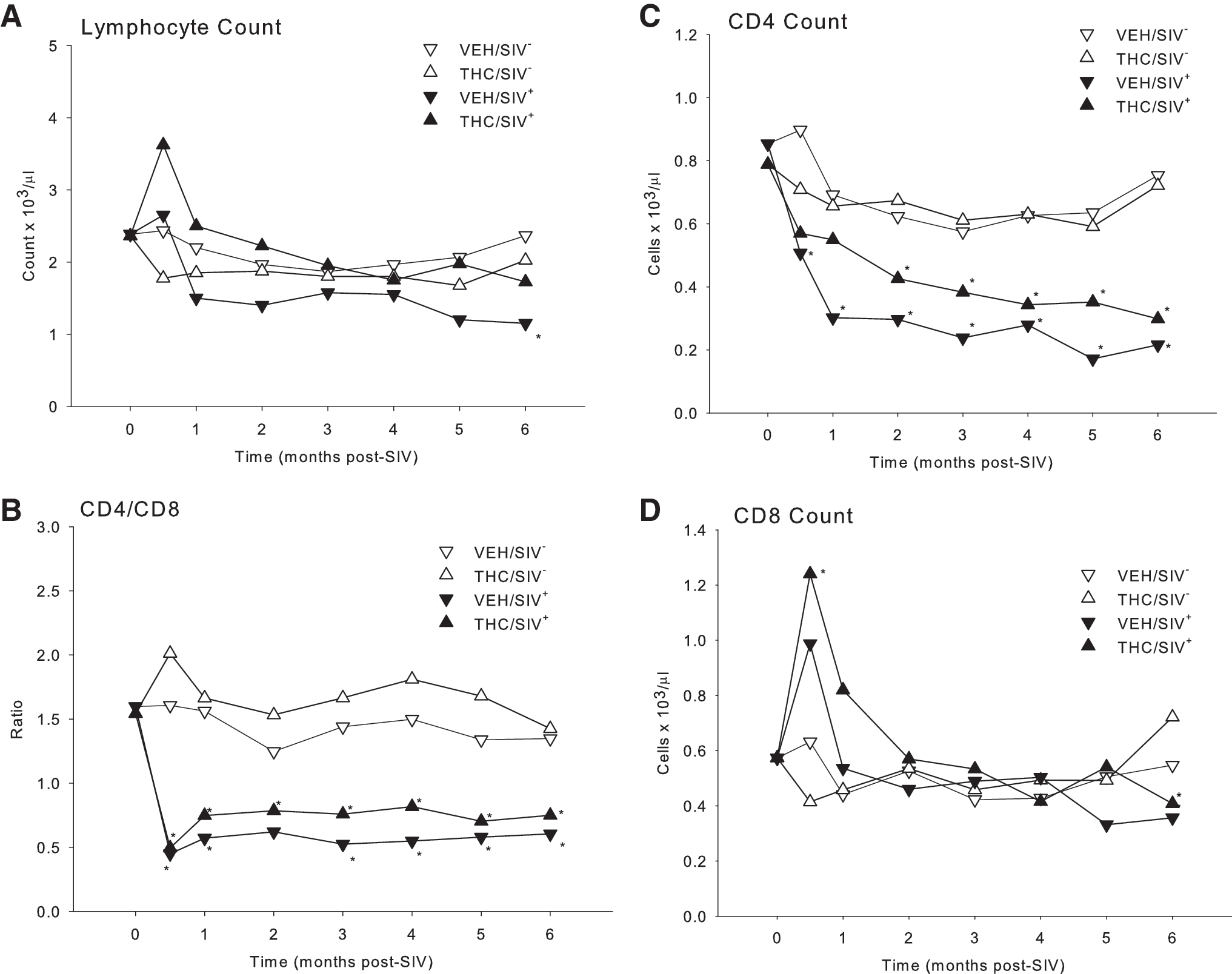
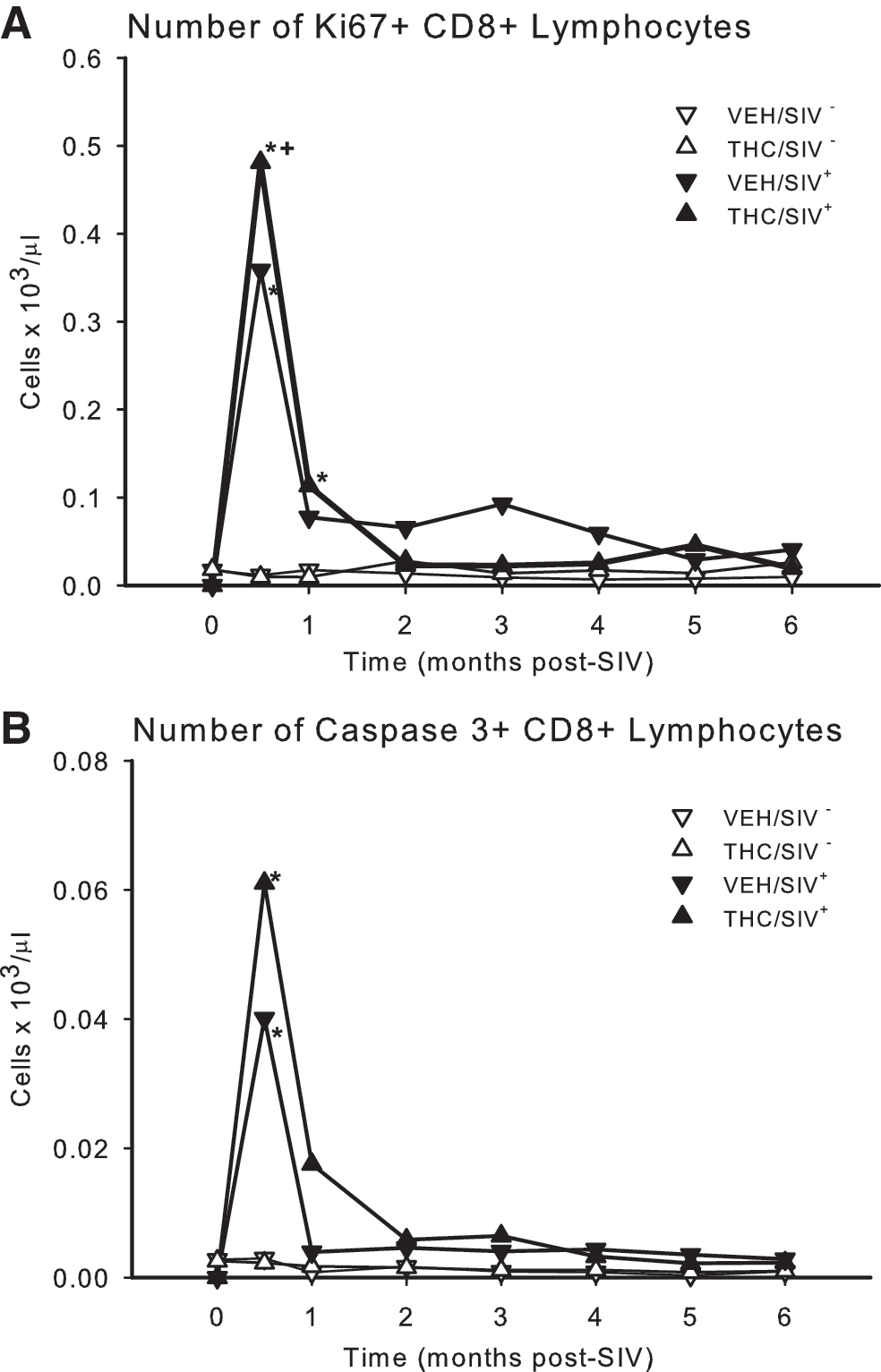
Numbers of
No deaths were noted during the first 5 months post-SIV infection in VEH/SIV+ or THC/SIV+ animals (Fig. 4). Two of the VEH/SIV+ animals succumbed to SIV infection shortly after 5 months, and a third reached end stage at 7 months. Among the THC/SIV+ animals, the first animal did not reach end stage until 11 months post-SIV inoculation. A summary of the clinical indications for euthanasia and the most salient necropsy findings is presented in Table 1.
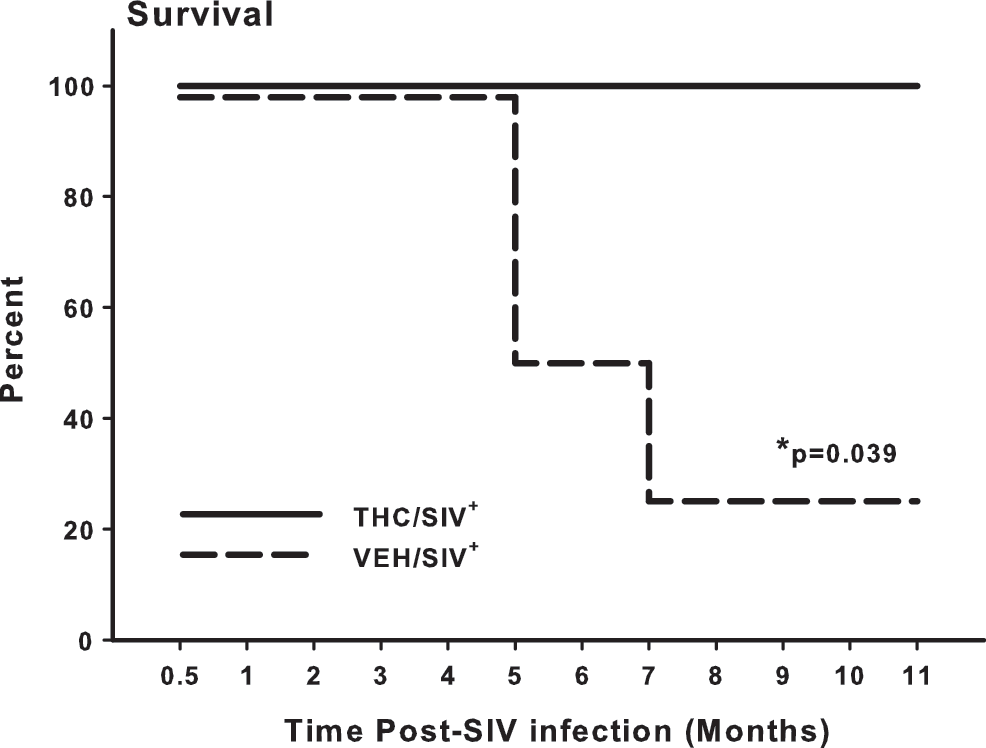
Percent survival of vehicle-treated infected (VEH/SIV+) and THC-treated infected (THC/SIV+) rhesus macaques over the initial 11 months post-SIV infection, the timepoint when the first THC/SIV+ animal reached end-stage criteria. Log rank statistic for the survival curve (Kaplan–Meier survival analysis) is greater than would be expected by chance (p = 0.037).
Animals were studied during the pretreatment period (prior to initiating the Δ9-THC administration) and following SIV infection during the “asymptomatic phase of infection” prior to any of them reaching criteria for euthanasia. This period was defined as the period of time following SIV infection during which none of the animals showed clinical signs of disease including weight loss greater than 5% of basal, secondary infections that required antibiotic treatment, neurological manifestations of disease, severe diarrhea, and dehydration. Animals were sacrificed and necropsy performed when they met predefined clinical indications including the presence of three or more of the following: weight loss greater than 15%, hypoalbuminemia (<3 mg/dl) in the presence of edema, anemia and thrombocytopenia, 3 days of complete anorexia, major organ failure or medical conditions unresponsive to treatment (including respiratory distress, intractable diarrhea, or persistent vomiting, secondary infections that required antibiotic treatment), neurological manifestations of disease, severe diarrhea, and dehydration. Survival was tracked throughout the first year postinfection, at which time all animals still on study were euthanized and necropsy was performed.
Notably different patterns in weight gain were observed between the initial 3 month post-SIV infection period and the subsequent 3- to 6-month post-SIV period (Fig. 5). This was particularly true for the VEH/SIV+ animals, which showed a tendency for greater (p = NS) weight loss than the THC/SIV+ animals. Echocardiography (Toshiba Aplio at 8.5 MHz) performed prior to (baseline) and 3 and/or 6 months after SIV infection in the two SIV-infected groups (VEH and THC) failed to show significant alterations in cardiac systolic or diastolic function (data not shown).
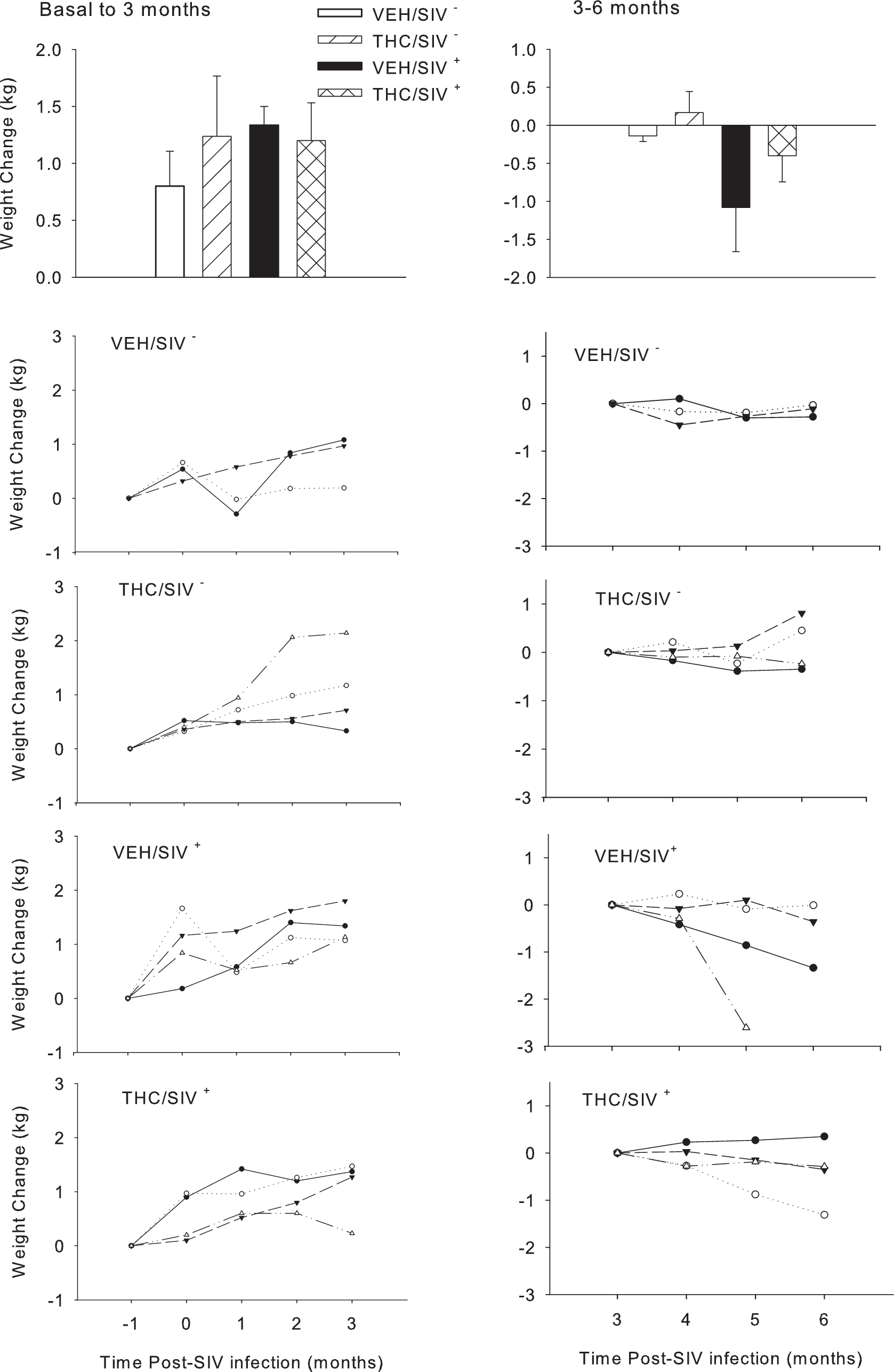
Body weight change (kg) during the initial 3 months post-SIV infection (left panels) and during the subsequent 3- to 6-month period of infection. Top panels depict change in body weight in vehicle-treated uninfected (VEH/SIV−), THC-treated uninfected (THC/SIV−), vehicle-treated infected (VEH/SIV+), and THC-treated infected (THC/SIV+) rhesus macaques expressed as means ± SEM over time post-SIV infection. Because of the individual variability, data are shown for each animal during the initial 3 months post-SIV infection and during the later 3–6 months post-SIV infection in the lower panels, showing the trends in body weight changes. The symbols in the line graphs represent individual animals in the specified treatment group and carry over across time.
Cannabinoids, including Δ9-THC, have been shown to have immunomodulatory effects 9,10 on cytokine production and lymphocyte phenotype, function, and survival 11,12,13 as well as cell-mediated immunity. 14 Similar immunosuppressant effects on lymphocyte 15 and alveolar macrophage 16 function have been reported in nonhuman primates. Furthermore, the potential of cannabinoids to regulate the activation and balance of human Th1/Th2 cells by a CB2 receptor-dependent pathway has been supported by findings from several studies. 17,18 More recent studies have provided evidence that the synthetic cannabinoid WIN 55,212-2 can potently inhibit HIV-1 expression in CD4+ lymphocytes and microglial cell cultures in a time- and concentration-dependent manner. 19 Moreover, Rock et al. 20 demonstrated the involvement of CB2 receptors in cannabinoid antiviral activity in microglial cells. To examine the possibility that the improved survival of SIV-infected rhesus could have been due to direct suppression of viral replication, we examined the in vitro effects of 1 and 10 μM Δ9-THC on viral replication in MT4-R5 cells (human T cell line) 21 infected with SIVMAC251 (10 TCID50). Pretreatment with 10 μM Δ9-THC (3.2 μg/ml) resulted in a significant decrease in cell-associated viral load in cells collected at 3 and 6 days post-SIV inoculation (Fig. 6). Several possible mechanisms could be involved in this suppression of viral replication, including effects on cell cycle, interaction of cannabinoids with viral coreceptor, and viral integration into the host genome. Moreover, the contribution of the specific cannabinoid receptor subtypes to the overall effects also remains to be examined.
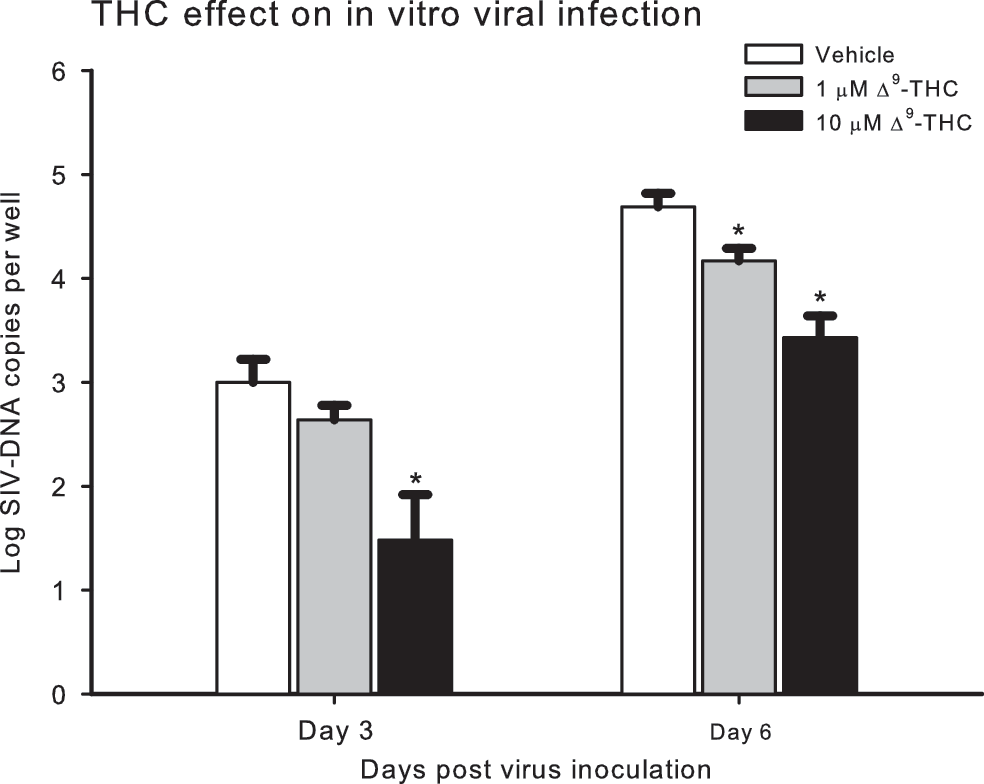
MT4-R5 cells (human T cell line) were seeded at a cell density of 5 × 105/ml in RPMI 1640 containing 10% heat-inactivated fetal calf serum, 2 mM glutamine, 100 U/ml penicillin, 100 μg/ml streptomycin, and 1 μg/ml puromycin in T25 flasks and maintained at 37°C with 5% CO2 in the presence of Δ9-THC (1 or 10 μM) or vehicle for 11 days. Culture media was replaced at 3–4 day intervals. After 11 days, 1.5 × 106 MT4-R5 cells from each treatment group were inoculated with 10 TCID50 SIVmac in fresh media containing Δ9-THC or vehicle, plated in 96-well dishes at a density of 1 × 105 cells/well, and maintained at 37°C with 5% CO2. Uninfected cultures of Δ9-THC- and vehicle-treated cells were maintained identically for use as controls. One-half of the replicate cultures were harvested 3 days post-SIV-infection, with the remainder harvested 6 days after infection. Supernatant and cell pellets were fractionated by centrifugation at 700 × g for 5 min at room temperature. Cell viability was monitored through cell counts using Trypan blue. DNA was prepared and the quantity of proviral SIV DNA in the cell pellets was determined by real-time PCR. DNA viral load (log SIV DNA copies/well) was quantified in cells harvested at 3 and 6 days postinoculation. Expression of CB1 and CB2 receptors was confirmed in these cells prior to using them in the study. Values are mean ± SEM of eight replicate wells. Results were analyzed with two-way ANOVA followed by pairwise multiple comparison procedures (Holm–Sidak method) *p ≤ 0.008.
In summary, this study is the first to report in vivo experimental data demonstrating that chronic Δ9-THC initiated prior to, and continued throughout the asymptomatic phase of SIV infection, does not impair the host's ability to control viral load, and does not increase morbidity and mortality from the infection. Although the small groups and natural variation in SIV disease are limitations of the study, the vehicle-treated group is representative of typical rhesus infections with SIVmac251. Δ9-THC treatment clearly did not increase disease progression, and indeed resulted in generalized attenuation of the classic markers of SIV disease (set point viral load/viral level in general). The mechanisms underlying the decreased mortality in cannabinoid-treated SIV-infected animals remain to be elucidated. However, based on our results and reports in the literature, we speculate that retention of body mass, attenuation of viral replication, and an overall immunosuppressant effect of cannabinoids may contribute to the amelioration of SIV disease progression seen in our study. The cellular mechanisms, including the potential role of the recently identified interaction between the HIV coreceptor CXCR4 and the cannabinoid system 22 as potential sites of pharmacoimmunomodulation, remain to be determined.
Footnotes
Acknowledgments
The authors would like to thank Joseph Moerschbaecher, PhD, for his helpful discussions on chronic THC administration and dosing and Dr. Catalin Filipeanu for cannabinoid receptor expression measurements. Additionally, the authors would like to acknowledge the technical expertise of Connie Porretta, Jean Carnal, Nedra Lacour, Peter Lewis, Merlin Johnson, Jessica Howard, and Jaime Hubbell. Supported by NIDA-020419-01.
Author Disclosure Statement
No competing financial interests exist.