
Abstract
The relative importance of routes of transmission of human T cell lymphotropic virus type 1 (HTLV-1) in Guinea-Bissau is largely unknown; vertical transmission is thought to be important, but there are very few existing data. We aimed to examine factors associated with transmission in mothers and children in Guinea-Bissau, where HTLV-1 is endemic (prevalence of 5% in the adult population). A cross-sectional survey was performed among mothers and their children (aged <15 years) in a rural community in Guinea-Bissau. A questionnaire to identify risk factors for infection and a blood sample were obtained. HTLV-1 proviral load in peripheral blood was determined and PCR was performed to compare long terminal repeat (LTR) sequences in mother–child pairs. Fourteen out of 55 children (25%) of 31 HTLV-1-infected mothers were infected versus none of 70 children of 30 uninfected mothers. The only factor significantly associated with HTLV-1 infection in the child was the proviral load of the mother; the risk of infection increased significantly with the log10 proviral load in the mother's peripheral blood (OR 5.5, 95% CI 2.1–14.6, per quartile), adjusted for weaning age and maternal income. HTLV-1 sequences of the LTR region obtained from mother–child pairs were identical within pairs but differed between the pairs. Vertical transmission plays an important role in HTLV-1 transmission in this community in Guinea-Bissau. The risk of transmission increases with the mother's proviral load in the peripheral blood. Identical sequences in mother–child pairs give additional support to the maternal source of the children's infection.
Introduction
H
HTLV-1 is endemic in Guinea-Bissau. In rural areas, where the current study was conducted, the adult prevalence was 4.9% in 2007. 10 The relative importance of different routes of transmission has not been established in this population but it is thought that sexual transmission is most common. 11 Two studies among adults in the capital Bissau found no relationship between HTLV-1 infection in mothers and their children and it was suggested that vertical transmission does not importantly contribute to the endemic HTLV-1. 11,12 However, cohort studies in Japan and Jamaica have described MTCT risks of approximately 20% 2,3,13 and the main risk factor, prolonged breastfeeding, is very common in Guinea-Bissau. 14 An earlier study from rural Guinea-Bissau found a significantly higher prevalence of HTLV-1 infection among adult children of HTLV-1-positive mothers. 10
To study the role of MTCT in the HTLV-1 epidemic in Guinea-Bissau, factors associated with HTLV-1 infection in children from infected and uninfected mothers were examined.
Materials and Methods
Study samples were collected in 2004 in Caió, a rural village in northwestern Guinea-Bissau consisting of 10 settlements dispersed amid cashew forests and rice fields. The total population is approximately 10,000 and most people are subsistence farmers. In 1997 an HIV/HTLV-1 population survey was performed and from the study participants all HTLV-1-infected women (aged 20–70 years) and age-matched HTLV-1-uninfected women were selected for this study. A fieldworker visited the women and children (<15 years) at home and after obtaining informed consent administered a questionnaire and took a 2-ml venous blood sample from the mother and her child(ren). The upper age limit of children was chosen assuming that sexual activity has mostly not started by this age in this population. 10 The area of living was defined as central or peripheral as previous studies showed different prevalences of HIV and HTLV-1 in the central and peripheral areas of Caió. 10 Medical care was provided by the project's physician when needed. At the time of the study no anti-retroviral therapy was available in Guinea-Bissau.
Diagnostics
Plasma samples of the mothers and the children were tested for the presence of antibodies to HTLV-1 and −2 by the Murex HTLV-1+2 (Abbott Murex Diagnostics, Dartford, UK). Reactive samples were retested by the same ELISA. HTLV-1 infection status was further determined by PCR using sequences targeted to the tax/rex gene. 15 No Western blot assay was performed. Each reaction was spiked with 10 copies of phage lambda DNA. This was coamplified in the same reaction to control for nonspecific inhibition. Because these tax/rex primers amplified both HTLV-1 and HTLV-2 sequences, amplicons were digested with the restriction enzyme Sau 3A (which cuts only HTLV-1) to distinguish the two viruses. In this study all PCR-positive samples were confirmed to be HTLV-1 by Sau 3A digestion.
Proviral load
To determine the proviral load of HTLV-1, 600 ng of extracted DNA from whole blood was amplified using the same primer set described above. The first round PCR was amplified for only 20 cycles and the second round PCR was amplified in a real-time thermal cycler using SYBR-green detection (Qiagen). Viral loads were determined with a standard curve generated from DNA extracted from the 8166 T cell line, for which a value of 3 integrated proviruses was used. 16 Results were expressed as proviral copies/105 cells. The limit HTLV-1 detection was 4 copies/105 cells, thus undetectable viral loads were assigned a value of 2 copies/105 cells.
The HIV status of all maternal subjects was obtained from previous studies; the testing and diagnostic algorithms have been described. 17
PCR and sequencing
Amplification of the proviral DNA was performed with a heminested PCR using standard conditions. The LTR region was amplified as two overlapping fragments with modified primers derived from Salemi et al. 18 : an LTR-gag PCR of 438 bp with MO195–MO196 as outer and MO197–MO198 as inner primers and a tax-LTR PCR of 475 bp with MO199–MO200 as outer and MO201–MO202 as inner primers. The primers that were used were as follows.
LTR to gag fragment
MO195 [outer forward (OF)] AACTAGCAGGAGTCTATAAAAGCG (based on AV117);
MO196 [outer reverse(OR)] AAAGATTTGGCCCATTGCCTAG (based on AV118);
MO197 [inner forward (IF)] ACAGTTCAGGAGGGGGCTC (based on AV119);
MO198 [inner reverse (IR)] TAGGGAATAAAGGGGCGCTC (based on AV120).
tax to LTR fragment
MO199 (OF) ACTCACACGGCCTCATACAG (based on AV121);
MO200 (OR) ACGCAGTTCAGGAGGCAC (based on AV122);
MO201 (IF) CTGTTTGAAGAATACACCAACATCC (based on AV123);
MO202 (IR) CTCAACCGGCGTGGATGG (based on AV124).
Purified PCR products were obtained after 1% agarose gel electrophoresis using Qiaquick Gel Extraction (Qiagen) and were sequenced using the inner PCR primers from both directions by Macrogen (
Statistics
Data were analyzed using STATA 11 (Stata Corporation, USA). Comparisons between normally distributed variables were made by the t-test and nonnormally distributed data were compared by the Wilcoxon rank sum test. Discrete data were compared by the chi-square or Fisher's exact test. Associations between continuous data were assessed by Spearman's correlation coefficient. Variables for mother's age, children's age, and log proviral load were divided into quartiles, as was done previously, 3,13 and weaning age and birth order divided in < and ≥ the median. To account for clustering of multiple children per mother, a general estimation equation logistic model was used to examine the factors associated with HTLV-1 infection in children. Statistical significance was determined by the Wald statistic.
The study was approved by the joint Ethics Committee of the Medical Research Council/Gambia Government and by the National Ethics Committee of Guinea-Bissau.
Results
Study participants
From previous studies in the village, 90 HTLV-1-positive women and 90 age-matched HTLV-1-negative women were selected. Of the 180 mothers, 5 had died and 42 had moved away from the study area or had temporarily traveled. A total of 133 (74%) mothers were successfully located by the field team and 130 (98%) agreed to participate.
The 130 participating women had 799 children who were alive; 67 mothers (32 HTLV-1 infected and 35 HTLV-1 uninfected) had children aged less than 15 years (140 children total). A blood sample was obtained from 125 of these 140 children, thus including 61 mothers (31 HTLV-1 infected and 30 HTLV-1 uninfected) with 125 children (55 born to HTLV-1-infected and 70 born to HTLV-1-uninfected mothers) in the main analysis (Fig. 1). All children were singletons. All study participants had at least one positive ELISA and a positive PCR, except for one mother whose sample was ELISA positive but PCR negative (Table 1
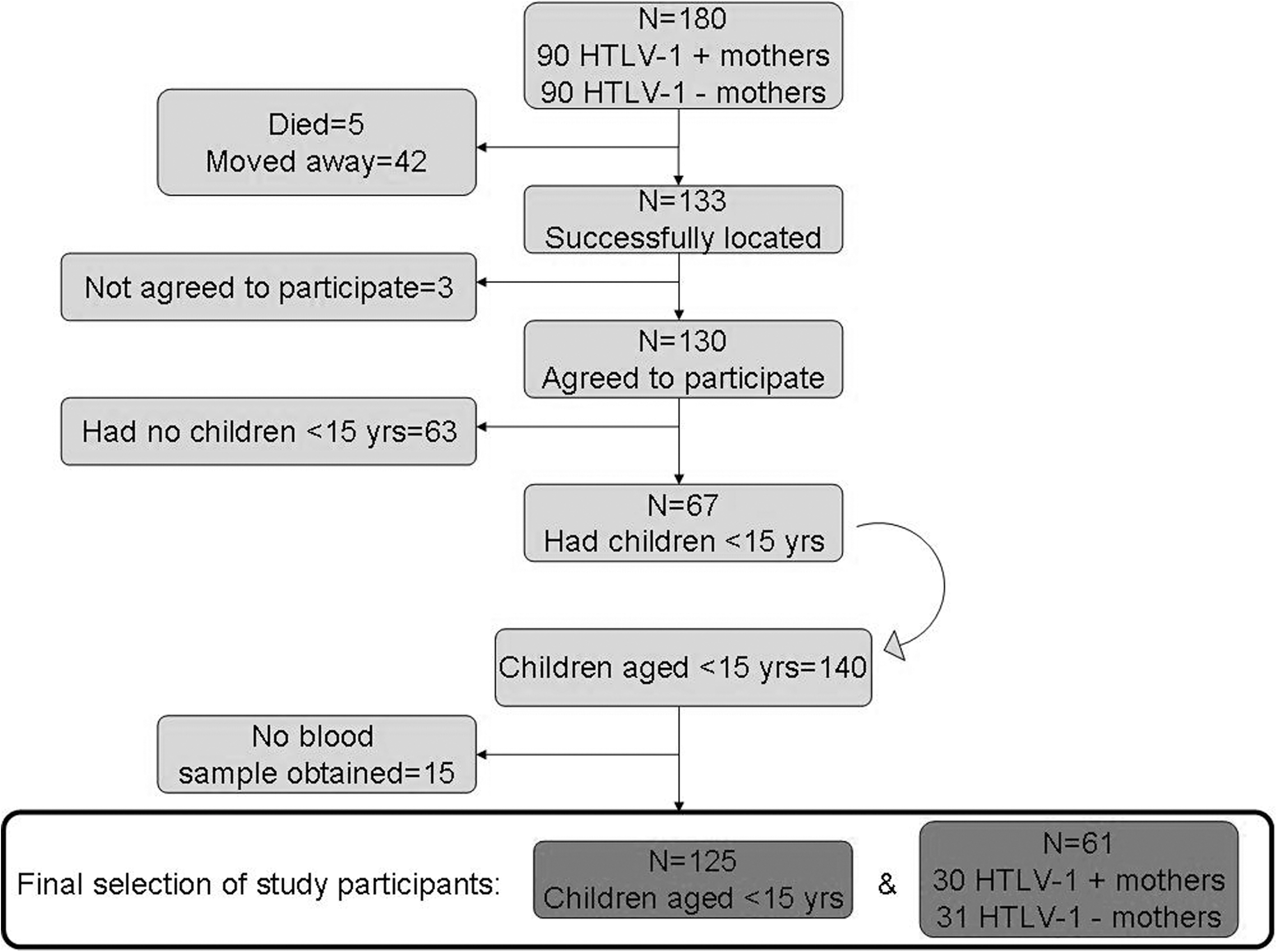
Flow diagram of study participants: The selection of mothers and their children, Caió, 2004.
The median age of HTLV-1-infected mothers at delivery was similar to that of uninfected mothers (both 34 years; rank sum test, p=0.9). The HTLV-1-infected mothers had had 179 live born children in total, of whom 52 (29%) had died. This was similar to the HTLV-1-negative mothers, who had had 188 live born children of whom 56 (30%) had died (chi square, p=0.9). Twenty-five (81%) of the 31 infected mothers and 18 (60%) of the 30 noninfected mothers lived in the central area of the village (chi square, p=0.08).
Proviral load and transmission
None of the children of HTLV-1-negative mothers were HTLV-1 positive. Of the 55 children born to HTLV-1-positive mothers, 14 (25%) were infected (p<0.001). Eleven HTLV-1-positive mothers had infected children: one mother had three infected children, one mother had two infected children, and the other nine mothers had one infected child each.
The median proviral load of the infected mothers was 31 copies/105 cells (range undetectable–2669). The median proviral load of infected children was 8 copies/105 cells (range undetectable–11,233). There was no correlation between the proviral load in mothers with that of their children (Spearman's r=−0.04, p=0.9).
Odds ratios (OR) were estimated for possible risk factors of HTLV-1 infection among the 55 children born to HTLV-1-infected mothers (Table 2). None of the HTLV-1-positive children had ever received a blood transfusion. A significantly increased OR of 3.5 [95% confidence interval (CI) 1.6–7.7] was found for each quartile increase in mother's log10 proviral load. A nonsignificantly increased OR for higher weaning age was observed (2.7, 95% CI 0.5–14.5). In the multivariable model, mother's log10 proviral load, weaning age, and any means of making money were kept in the model, because these are known risk factors for HTLV-1 infection. Any means of making money (OR 4.1, 95% CI 0.8–19.5) and weaning age above 19 months (OR 3.1, 95% CI 0.6–15.1) both confounded the OR of mother's log proviral load by >10%, but were themselves not statistically significant predictors of HTLV-1 infection. The adjusted OR for mother's log10 proviral load was 5.5 (2.1–14.6) per quartile (p=0.001).
One mother did not have a proviral load measurement.
Missing data for two mothers.
Four children were still breastfed at the time of the study (ages 2.5, 6, 8.5, and 12 months) and were excluded from this analysis.
OR, odds ratio; CI, confidence interval.
For some children, the HTLV-1 diagnosis of their mother was made after their birth. To exclude the possibility of misclassification of these children, an additional analysis was performed only including children whose mother's HTLV-1 status was known at birth. Thirty-one children born to HTLV-1-positive mothers were then included, of whom 10 (32%) (p<0.001) were HTLV-1 infected. The unadjusted OR for mother's log10 proviral load was 4.1(1.5–11.5) and the adjusted (for weaning age and any means of making money) OR was 3.6 (1.2–10.3). Another analysis was performed to ascertain that sexual activity had not yet started; girls aged 13 and 14 years old (n=8) were excluded and the adjusted OR for mother's log10 proviral load [4.8 (1.6–14.1)] remained significantly increased.
The median proviral load of mothers who transmitted was 348 copies/105 cells (range 13–1134) and of mothers who did not transmit 6 copies/105 cells (range undetectable–2669) (rank sum test, adjusted for clustering; p<0.001) (Fig. 2).
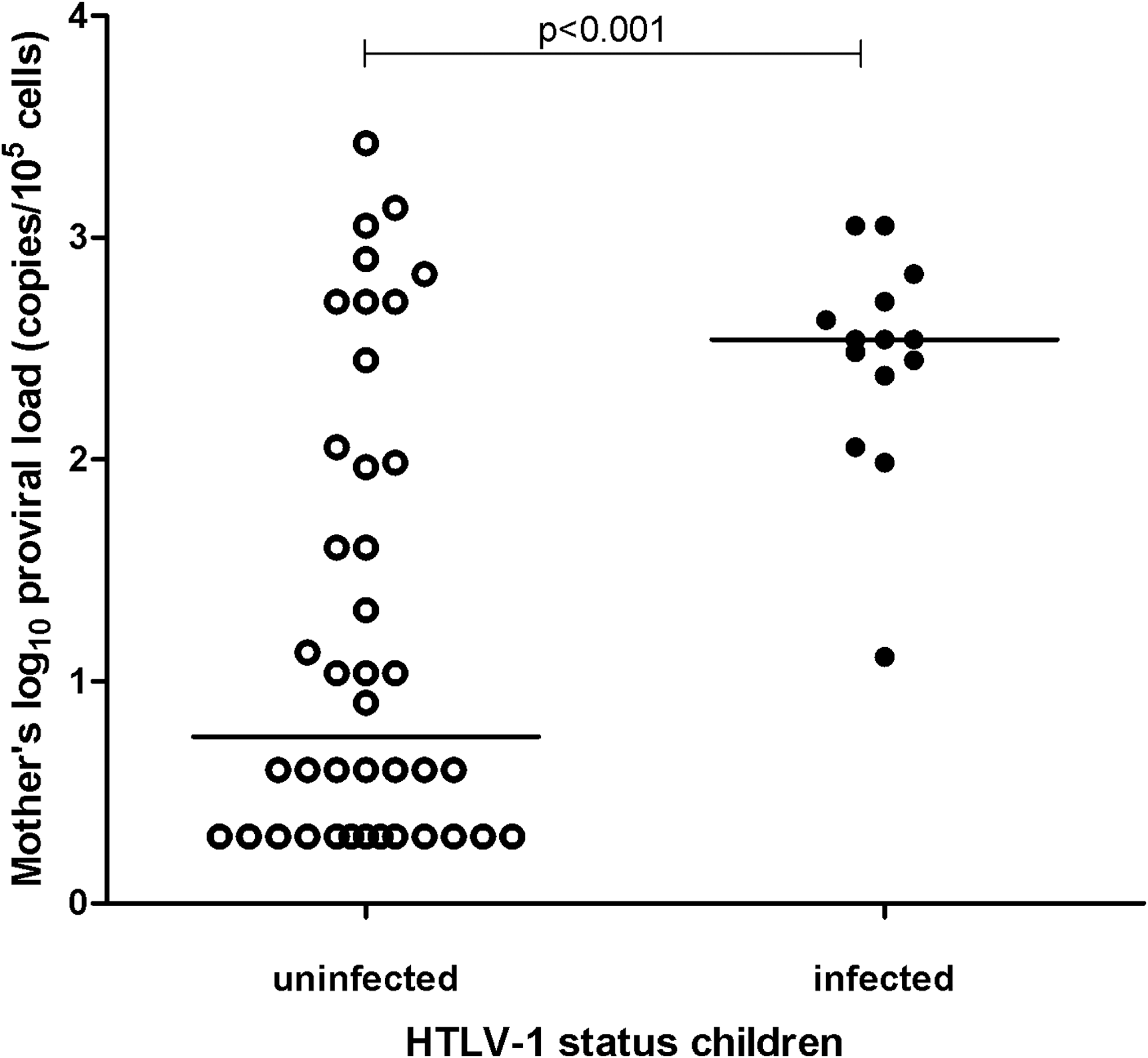
Mothers' human T cell lymphotropic virus type 1 (HTLV-1) log10 proviral load by HTLV-1 infection status of their children in Caió, Guinea-Bissau. Horizontal lines indicate median log10 proviral load.
The youngest child that tested HTLV-1 positive was 2.5 months. The proviral load of this child was 264 copies/105 cells and he was born to a mother whose proviral load was 348 copies/105 cells and whose two other sons that participated were also HTLV-1 positive.
Among the 31 HTLV-1-infected mothers, 16 (52%) mothers had one child participating in the study and 15 (48%) mothers had two or more children that participated. Six of the 15 (40%) mothers had children with discordant HTLV-1 status. Of one mother (3%) all enrolled children were HTLV-I infected and of eight mothers (26%) all enrolled children were uninfected.
Sequence analysis
In an effort to confirm the maternal source of HTLV-1 infection, HTLV-1 sequence data were obtained from mother–child pairs. As HTLV-1 evolution is very slow, 21 we hypothesized that identical sequences would be observed for mother–child pairs if vertical transmission had occurred whereas infection from another source would appear as nonidentical mother–child sequence pairs. Twenty-three samples of mother–child pairs were still available for sequencing (other samples were depleted or missing) and from 16 of these LTR sequences could be obtained. Samples in which sequencing failed had a lower proviral load compared to samples on which sequencing was successful. Complete sequences were obtained from five mother–child pairs and only the tax-part of the LTR sequence was obtained from two mother–child pairs (age range children: 3–14 years). These sequences all belonged to the cosmopolitan subtype 1a, subgroup D, which has also been described in pregnant women in the capital of Guinea-Bissau, Bissau. 22 The complete pair sequences were compared to another Caió HTLV-1 sequence (Caió4142, unpublished observations); differences from this Caió master sequence are marked in Fig. 3. There is 100% identity between sequences within pairs, but each pair has unique differences from the master sequence.
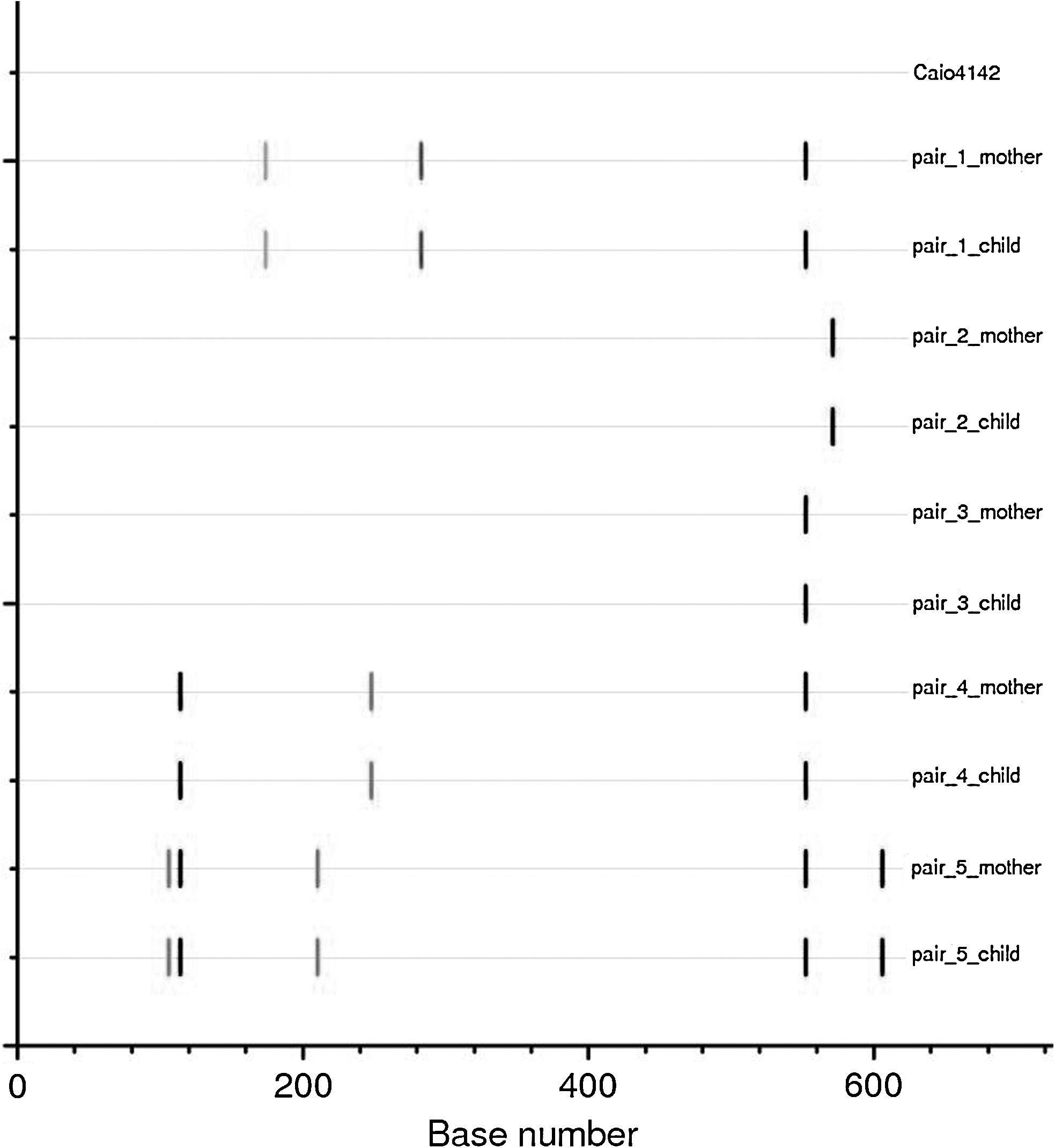
Highlighter plot of five complete LTR sequences of mother–child pairs compared to a reference strain from Cai
Discussion
This study demonstrates a very strong and highly significant association between HTLV-1 proviral load of mothers and HTLV-1 infection of their children in a rural community in Guinea-Bissau. This is in line with studies performed in Jamaica and French Guyana. 3,7 This is the first time such an association has been reported in an African study and suggests MTCT is very likely to be a major factor contributing to the HTLV-1 endemic in this area. It is not possible to rule out other transmission routes in this study due to its cross-sectional design. However, HTLV-1 loads are stable over time, 23 therefore the measured maternal load likely reflects the load around time of delivery. The only factor associated with HTLV-1 infection in children was the maternal proviral load while none of the children with an uninfected mother was HTLV-1 positive.
A limitation of the study is that in some cases it was uncertain whether the mother was HTLV-1 infected at the time of delivery. We therefore performed a sensitivity analysis excluding these cases, and still showed a significantly increased OR for the mother's log10 proviral load. A significantly increased OR was also still observed after exclusion of 13- and 14-year-old girls, in order to further rule out a potential sexual route of infection. Also, seven mother–child pairs had identical HTLV-1 sequences, lending additional support to the maternal origin of the children's infection. 24 The children from whom these sequences were obtained had an age range of 3–14 years, further suggesting sexual transmission to be very unlikely in older children. Unfortunately, only seven mother–child pairs could be sequenced. This is a limitation of the study, although it seems unlikely that these sequences would not be representative of all the pairs from the study. The age at weaning, which was previously shown to be an independent risk factor, was not associated with HTLV-1 infection in our study. This lack of association could be due to the high median weaning age (20 months) and the fact that all but one child were breastfed longer than 12 months, so there was no comparison group of short-term breastfed children. 3,4 Maternal income was measured in this study by assessing whether the woman had the means to make any money. This may, in the rural setting of this study, not be the best indicator for socioeconomic status and therefore might not be significantly related to HTLV-1 infection, or there is truly no association, as opposed to what was observed in the Jamaican cohort. 3,4,25
The range of the proviral load measured in this study (0–11,233/105 cells) was in line with that in asymptomatic carriers from Japan (100–5000/105 cells) and from Brazil (0–13,800/105 cells). 26,27 The median proviral loads in the current study, however, were lower than those observed in these studies. Whether this is due to differences in methods of measuring the proviral load or selection of subjects or whether this is a true difference remains to be resolved.
The observed discordance in HTLV-1 status between siblings from HTLV-1-positive mothers suggests that factors in addition to maternal proviral load play a role in the transmission. Biggar et al. showed an independent increased risk with increase in concordance of HLA type 1 alleles of children in the Jamaican cohort 25 and Brown et al. found certain haplotypes of the IL-6 and IL-10 genes to be associated with HTLV-1 infection. In both studies, this association was independent of weaning age and the mother's proviral load. Also, a dominant gene on chromosome 6 was found to be associated with childhood infection in families of African descent in French Guyana. 28 In genetic studies among adults, specific polymorphisms in the DC-SIGN gene were associated with HTLV-1 infection in a Brazilian population 29 and CCR2-64I (a polymorphism of a chemokine receptor gene) in the Jamaican cohort. 30 A factor that has not been explored so far is the role of nonexclusive breastfeeding (versus exclusive breastfeeding), which is the rule in Guinea-Bissau and which has been found to be an important risk factor for vertical transmission of HIV-1. It is conceivable that this may influence the transmission of HTLV-1 as well. Unfortunately, this factor was not covered by our questionnaire. It would be of major interest to further explore the factors involved in the discordance of HTLV-1 infection of siblings. They represent a unique group of individuals with a very similar exposure to HTLV-1, but an apparent difference in susceptibility to the infection.
The youngest infant in this study with detectable provirus was 2.5 months old. In a Japanese cohort, 3–4% of bottle fed infants born to HTLV-1-positive mothers became HTLV-1 infected, suggesting other routes of transmission than breastfeeding. 31 In a recent review, Hino suggests placental transmission during delivery would be the most plausible route. 32 This review also highlights that bottlefeeding is preferred over short-term breastfeeding for prevention of MTCT, as there is no clear evidence that the risk of transmission with short-term breastfeeding is equal to the risk of transmission with bottlefeeding. 32 This may not apply, though, to developing countries where the risks of not breastfeeding may not outweigh the risks associated with HTLV-1 transmission.
In summary, in this study from rural Guinea-Bissau, we found 25% of children of HTLV-1-infected mothers to be HTLV-1 infected. Infection in the children was significantly associated with an increased maternal proviral load and in seven mother–child pairs, identical LTR sequences were observed.
Footnotes
Acknowledgments
We would like to thank Abdoulie Jabang for his help with the sequencing work and Roel Coutinho for his very useful suggestions and comments. A modified abstract of this article has been published online (
Author Disclosure Statement
No competing financial interests exist.