
Abstract
Vaginal SHIVSF162P3 acquisition in pigtail macaques (Macaca nemestrina) is dependent on time point during the menstrual cycle. Susceptibility is higher around menstruation and lower at ovulation in mid cycle. This complicates the design of repeat low-dose (RLD) SHIV exposure studies because virus challenges given during low susceptibility periods have lower chances to infect. To account for fluctuating susceptibility, we analyzed menstrual cycles rather than exposures until infection following virus challenges. We first reanalyzed infection data of 41 macaques receiving placebo or no treatment during once (n=18) or twice (n=23) weekly virus exposures. The same number of cycles was required for infection with either challenge frequency, while it took a median four or six challenges for once or twice weekly exposures, respectively. More virus challenges to infection likely reflect frequent unsuccessful exposures in frequently exposed animals. When reanalyzing two previously reported biomedical HIV intervention studies, we found 1% tenofovir gel was 74% or 86% efficacious based on cycles or exposures (p=0.019 or p=0.003, respectively, Fisher's exact test), while 1% raltegravir gel was 84% or 89 % efficacious, respectively (p=0.047 or p=0.031). Evaluating the number of menstrual cycles rather than exposures until infection can account for varying susceptibility during the menstrual cycle. Our observations have implications for future study designs such as planning the frequency of virus exposures. Menstrual cycle analysis may also avoid potential overestimation of efficacy against vaginal challenges during low susceptibility periods in the cycle that are unlikely to cause infection.
T
For experiments with vaginal RLD SHIV exposures, 2,7 our group uses pigtail macaques (Macaca nemestrina) because they can easily be vaginally infected, have stable year-round median 32-day menstrual cycles, 8,9 and are of favorable size for gynecological examinations. 10 However, their susceptibility to vaginal SHIV infection fluctuates during the menstrual cycle, peaking 1 week prior to and 1 week after the onset of menstruation. 9,11 This complicates the analysis of the number of exposures required for infection, as repeated exposures during the cycle thus have unequal chances to infect and depend on exposure initiation time within the cycle. As an alternative, we have refined the RLD model for evaluating biomedical intervention studies by reporting the number of menstrual cycles required for infection. 12 –15 To accomplish this, we have since so far used calendar months, 28-day time segments, or actual menstrual cycle analysis with differing cycle lengths for each animal, 12 –15 and have attempted to initiate virus exposures at select menstrual cycle phases. 14 We here report our experiences, and provide an in-depth description of our analysis methods.
To illustrate the effects of two different analysis methods, we provide a side-by-side comparison of the number of exposures until infection occurred and of cycles to infection. In Fig. 1, we plotted Kaplan–Meier infection survival curves of 41 SHIVSF162P3-infected female control animals. Adult female pigtail macaques were infected at the CDC according to humane care guidelines 16 and with IACUC (Institutional Animal Care and Use Committee) approval. They were control- or placebo-treated animals from various prevention trials 2,7,12,13,17 with different study designs; their menstrual cycle determination, infection, and susceptibility during the cycle have been reported 9,11 and tabulated. 11
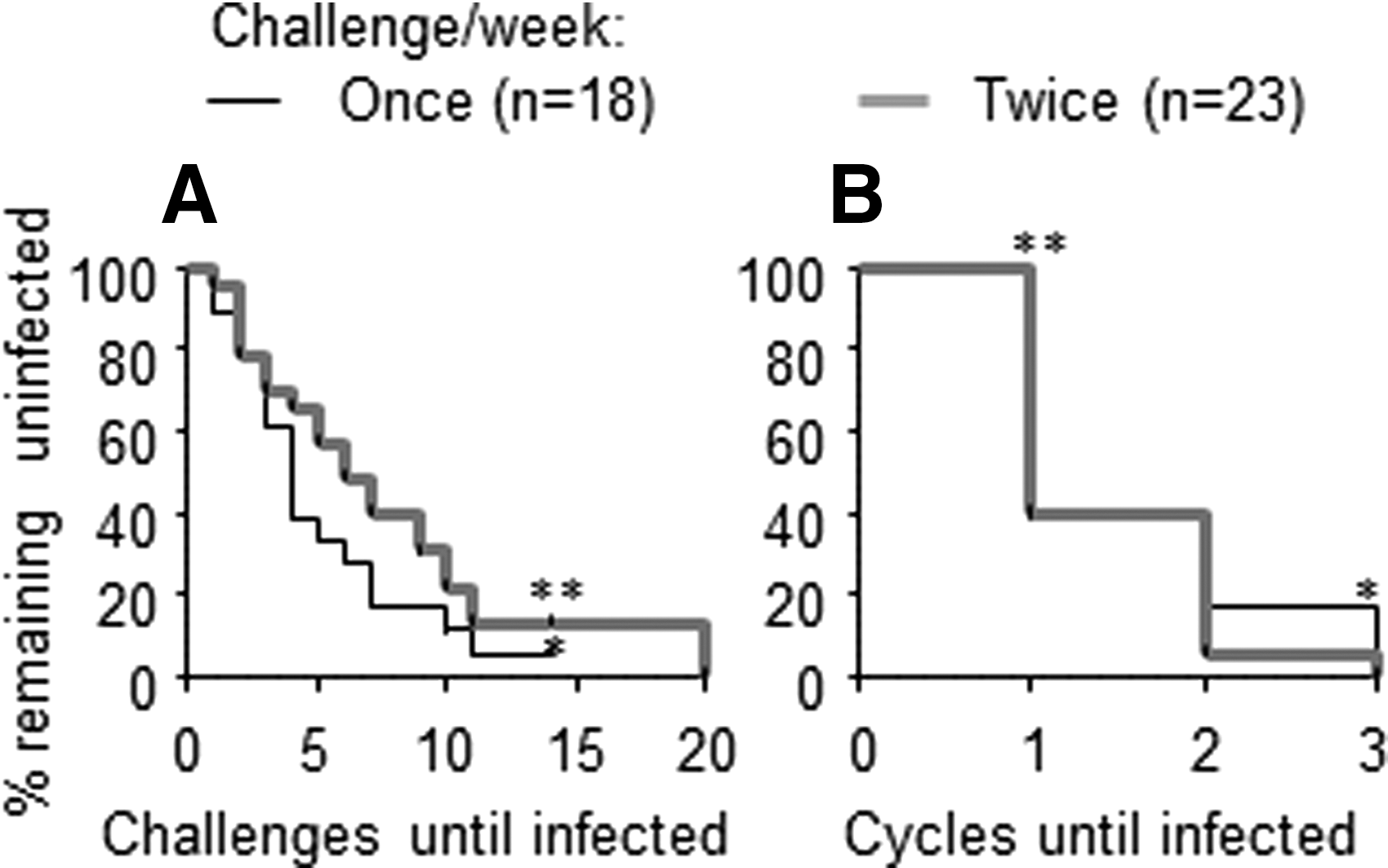
Comparison of analysis methods for data from 41 untreated or placebo-treated control macaques. Forty-one pigtail macaques were repeatedly exposed to intravaginal SHIVSF162P3 with random start time relative to their menstrual cycle (previously summarized
11
).
Macaques received one (n=18) or two (n=23) low-dose SHIVSF162P3 exposures 18 (10–50 TCID50) per week for up to 14–20 exposures, in accordance with their original study designs. 2,7,12,13,17 Exposures started randomly within the menstrual cycle, continued throughout menstruation, and were stopped when animals became SHIV positive. The infecting virus exposure was identified as the challenge given 7 days prior to SHIV detection of at least 50 copies/ml. To determine the number of menstrual cycles required for infection, we applied 28-day periods from the start of challenges, regardless of actual cycle data. Logistical hurdles prevented the implementation of 32-day periods because resulting 8-day intervals between virus challenges meant animal procedures would fall on different weekdays each week, conflicting with operational procedures in our animal facility. Alternatively, if challenges occurred on the same weekday each week, some 32-day periods would have four and others would have five virus exposures, complicating comparisons of infections during menstrual cycles. For one exposure per week and using 28-day periods, SHIV infection in the first, second, third, and fourth cycle meant macaques received 1–4, 5–8, 9–12, or 13–16 exposures before their first positive viral load, respectively.
Uninfected animals were evaluated at completed cycles, even if further exposures happened in additional, but incomplete cycles. For example, a macaque remaining uninfected after 14 weekly challenges completed three cycles but not four, and received two superfluous challenges that were not evaluated. For two exposures per week, infection in the first, second, or third cycle meant macaques received 1–8, 9–16, or 17–24 exposures before their first positive viral load, respectively. Kaplan–Meier survival graphs were constructed and analyzed with GraphPad Prism5.03 (San Diego, CA), also used for Fisher's exact, two-tailed outcome comparisons.
When we separately analyzed macaques receiving one or two challenges per week, we found four and six median numbers of exposures were required for infection, respectively (Fig. 1A). This difference was not statistically different (log-rank test, p=0.19). Infection rates (infections/number of exposures) were also not significantly affected by challenge frequency (p=0.271, Fisher's exact test). Three macaques remained uninfected after 14 exposures. Although the difference in challenges required for infection was not statistically significant for the two challenge frequencies, the observation is consistent with expectations due to varying susceptibility during the cycle. If a portion of animals started challenges in a low susceptibility period, more unsuccessful exposures will happen in twice-weekly exposed animals compared to once-weekly exposed animals until a high susceptibility period is reached.
The survival curves for menstrual cycle analysis for macaques receiving one or two challenges per week were strikingly similar for the two frequencies (Fig. 1B; log-rank test, p=0.51). One median cycle was required for infection, regardless of exposure frequency. Thirty-nine percent of the macaques remained uninfected after the first menstrual cycle of challenges for each frequency. These findings indicate that exposing animals twice per week is not advantageous over once per week. This new observation has implications for future study designs with reduced virus challenge frequency, potentially improving animal health due to reduced anesthesia frequency and lowering virus needs and animal technician time.
We next conducted a reanalysis of two previously published HIV intervention studies 15,19 and provide a side-by-side evaluation of the number of exposures until infection occurred (Fig. 2, left panels), compared to the published evaluation using cycles to infection (Fig. 2, right panels). Only partially efficacious interventions were reanalyzed, because the completely protective interventions we have reported 7,12,13 are fully efficacious, regardless of the analysis method. The first intervention included a 1% tenofovir (TFV) gel modality in which six macaques received vaginal gel and were challenged with virus 30 min and 72 h later, 19 and compared to 10 placebo-treated animals. A second intervention had 1% raltegravir (RAL) gel given 3 h after SHIV exposure as postexposure prophylaxis 15 (six experimental and four placebo-treated macaques). Efficacy of preventing infection was calculated as Efficacy (%)=1 – (P i/P c) with P (infection rate; i=intervention, c=control)=number of infections/number of challenges or cycles.
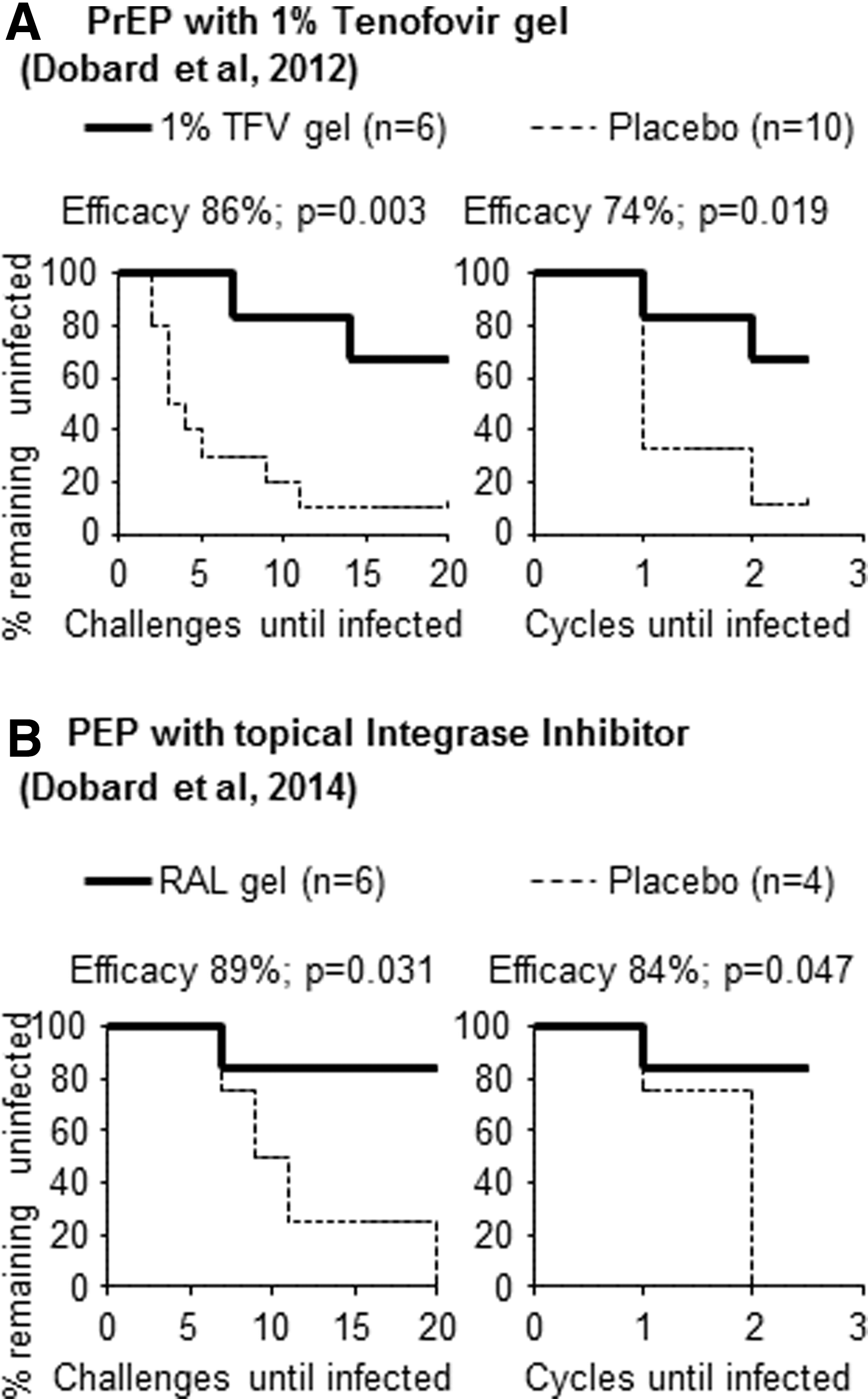
Comparison of analysis methods for intervention data. The graphs show Kaplan–Meier survival graphs to plot the percentage of pigtail macaques remaining uninfected after the number of vaginal SHIVSF162P3 challenges received (left panels) or after the number of menstrual cycles with virus challenges (right panels).
For the first intervention study evaluating the efficacy of 1% TFV gel to prevent vaginal SHIVSF162P3 infection, 19 analysis of exposures to infection showed 86% efficacy (Fig. 1A, left panel, p=0.003, Fisher's exact test) compared to 74% efficacy (Fig. 1A, right panel, p=0.019) when efficacy measurements were analyzed by number of exposures or cycles to infection, respectively. Likewise, for 1% RAL gel, 15 efficacy was calculated at 89% or 84% (Fig. 1B, p=0.031 or p=0.047, respectively, by Fisher's exact test). Thus, the two interventions found partially efficacious with cycle analysis were also partially efficacious using exposure number as the analysis method. In both examples, menstrual cycle analysis yielded a slightly lower efficacy value. However, this observation could not be supported by statistical comparisons, as only two example trial results were available. Larger sample sizes and additional statistical analyses might be able to ascertain this observation with statistical significance if and when more trial results become available.
It is possible, but not proven by our observations, that analysis by exposure number may overestimate efficacy, potentially because unsuccessful virus challenges during low susceptibility periods contribute to efficacy calculations. This is particularly true for protected animals who survive many challenges uninfected, while most control animals become infected as soon as a high susceptibility period is reached. Therefore, low susceptibility periods may affect efficacy calculations for intervention animals more than control animals when analyses are based on exposure numbers.
The choice of analysis method by number of cycles or exposures likely matters most when experimental groups are composed of animals that are not distributed equally in all possible parts of their cycles at study start, i.e., have some degree of synchronicity. Then it is possible that animals of one group but not the other start exposures in low susceptibility periods. It is difficult to distribute animals equally into study groups according to cycle status. Progesterone data are easiest to interpret in retrospect when longitudinal trajectories are available, i.e., not at study start. We have also encountered individual animals with consistent cycling patterns until experimentation started, only to find that the stresses of increased animal handling and fasting for anesthesia unpredictably affected cycles. We therefore suggest it is prudent to choose analysis by assessing menstrual cycles post hoc rather than anticipating menstrual cycles before study start.
Menstrual cycle analysis solves the problem of lacking cycle synchronicity at study start, but it does not eliminate the problem that animals may be cycling irregularly, and are unequally distributed in study groups. For example, if in a study comparing interventions, animals in one group are regularly cycling but in the other group animals are not cycling or have irregular cycles, their proportions of time in periods of high susceptibility will vary and affect the interpretation of intervention efficacy. However, this biological issue of differing susceptibility confounds both analysis types, and is not solved by switching analysis method.
When we implemented these methods a few issues arose regarding experimental design. We have since chosen to use 28-day increments rather than actual, measured cycle lengths as previously explored. 14 This had resulted in varying number of virus challenges per cycle due to short or long cycles. Not relying on actual cycle length determination also avoids lengthy cycle monitoring before study start to determine day 1 of each cycle, and exclusion of animals with incomplete cycle information, saving time and allowing all challenge data to be used. We prefer to use 28-day analysis units over the median 32 days of pigtail cycles 8,9 to allow convenient once-weekly exposures on the same week day in each unit, as is desirable for workflow in our animal facility.
We acknowledge the effect of the 4-day difference from median 32-day cycle length has not yet been fully analyzed. For animals remaining uninfected or protected from infection, we have started to analyze only completed 28-day units, because partial cycles may have started during low susceptibility periods without a substantial chance of infection. Thus, we now usually plan experiments in multiples of 28-day intervals, i.e., 4, 8, 12, 16, or 20 exposures once per week. For example, the uninfected macaque after 14 weekly challenges shown in Fig. 1 received two superfluous challenges that were not evaluated because the fourth cycle was incomplete. Also, exposure interruptions are difficult to interpret with both analysis methods, as animals may miss susceptible periods. For example, one previously evaluated macaque (macaque 303 11 ) was not included in Fig. 1 because virus challenges were interrupted for 3 weeks due to a holiday. If unavoidable, a full 28-day interruption can be considered, regardless of analysis method.
A study limitation was that we did not evaluate further reduction of virus challenge frequency beyond once per week. In addition, exploring implications for statistical power of RLD studies will be helpful, as will statistical simulations of cycle synchronicity in animal groups of different sizes. This was not the focus of this report on our practical experiences with RLD studies.
Alternatives to menstrual cycle analysis are to synchronize macaques before study start, e.g., with birth control pills as we have previously done, 20 or to start virus challenges at the same cycle time point in each animal, also previously reported. 14 These methods are laborious and time intensive and are susceptible to erroneous menstrual cycle judgment and to unpredictable cycling changes during experimentation.
Footnotes
Acknowledgments
The findings and conclusions in this presentation are those of the authors and do not necessarily represent the views of the Centers for Disease Control and Prevention. SHIVSF162P3 was obtained through the NIH AIDS Reagent Program, NIAID, NIH from Drs. Janet Harouse, Cecilia Cheng-Mayer, Ranajit Pal, and the DAIDS, NIAID. We gratefully acknowledge CDC's animal care staff, the HIV Lab Animal Model staff, and other CDC colleagues who performed the studies from which previously published data were analyzed. We thank Debra Hanson for insights into animal model designs. This work was funded by the CDC and an interagency agreement (Y1-A1-0681-02) between the CDC and NIH.
Author Disclosure Statement
No competing financial interests exist.