
Abstract
To prevent HIV-1 infections in females, vaccines that elicit antibodies in vaginal secretions are much desired. To induce these antibodies, intravaginal vaccinations are sometimes recommended. However, the benefit of intravaginal vaccination remains a topic of debate, and parallel studies of intravaginal and intramuscular vaccination routes are rarely performed. Here we describe tests of mucosal and systemic antibodies after mouse vaccinations by several routes with HIV-1 envelope protein formulated in adjuvant. Response magnitudes were as follows: intraperitoneal > intramuscular = intravaginal tissue ≥ subcutaneous. We found that the well-accepted and logistically feasible intramuscular immunization was similar to intravaginal tissue immunization for the induction of antibodies in blood and vaginal secretions. Results suggest that a routine comparison of intravaginal and intramuscular immunizations may serve as a beneficial gatekeeper for the development of new intravaginal HIV-1 vaccines.
H
In a preliminary experiment, we compared adjuvants. We mixed 5 μg HIV-1 envelope protein with CFA/IFA [Sigma, Freund's adjuvant complete or incomplete; envelope/phosphate-buffered saline was emulsified in CFA (for the first injection) or IFA (for booster injections) using equal volumes of antigen and adjuvant], alum (Imject®Alum, Pierce Cat #77161; equal volumes of antigen and adjuvant were mixed per the manufacturer's recommendations), nonoxynol (Options Conceptrol Vaginal Contraceptive Gel with 4% nonoxynol-9, Ortho, 10 μl/dose 5 ), or mannan (Sigma M7504, 0.6 mg/dose, prepared as described previously 6 ). Vaccines were injected superficially into vaginal tissue (ivag-t, 50 microliters per mouse injection) to ensure retention of product at the site. We found that CFA/IFA generated the best response among the tested adjuvants (data not shown). The next best response was with alum or nonoxynol, and because alum is a licensed product, it was selected for continued study.
Parallel tests of different injection routes were then performed with the envelope protein/alum vaccine. In this case, animals received priming and booster immunizations separated by a 3- to 4-week interval. As before, 5 μg protein in alum were used per injection in a 50 μl final volume. Animals were not pretreated to synchronize estrus cycles before vaccinations.
Figure 1A and B shows the results from an experiment in which i.m. (in the hind leg) and ivag-t immunizations were compared. Animals either received their priming and booster dose by the same route (homologous, i.m. followed by i.m., or ivag-t followed by ivag-t) or by different routes (heterologous, i.m. followed by ivag-t, or ivag-t followed by i.m.), 7 and anti-envelope antibodies were tested 2 weeks after the booster by ELISA.
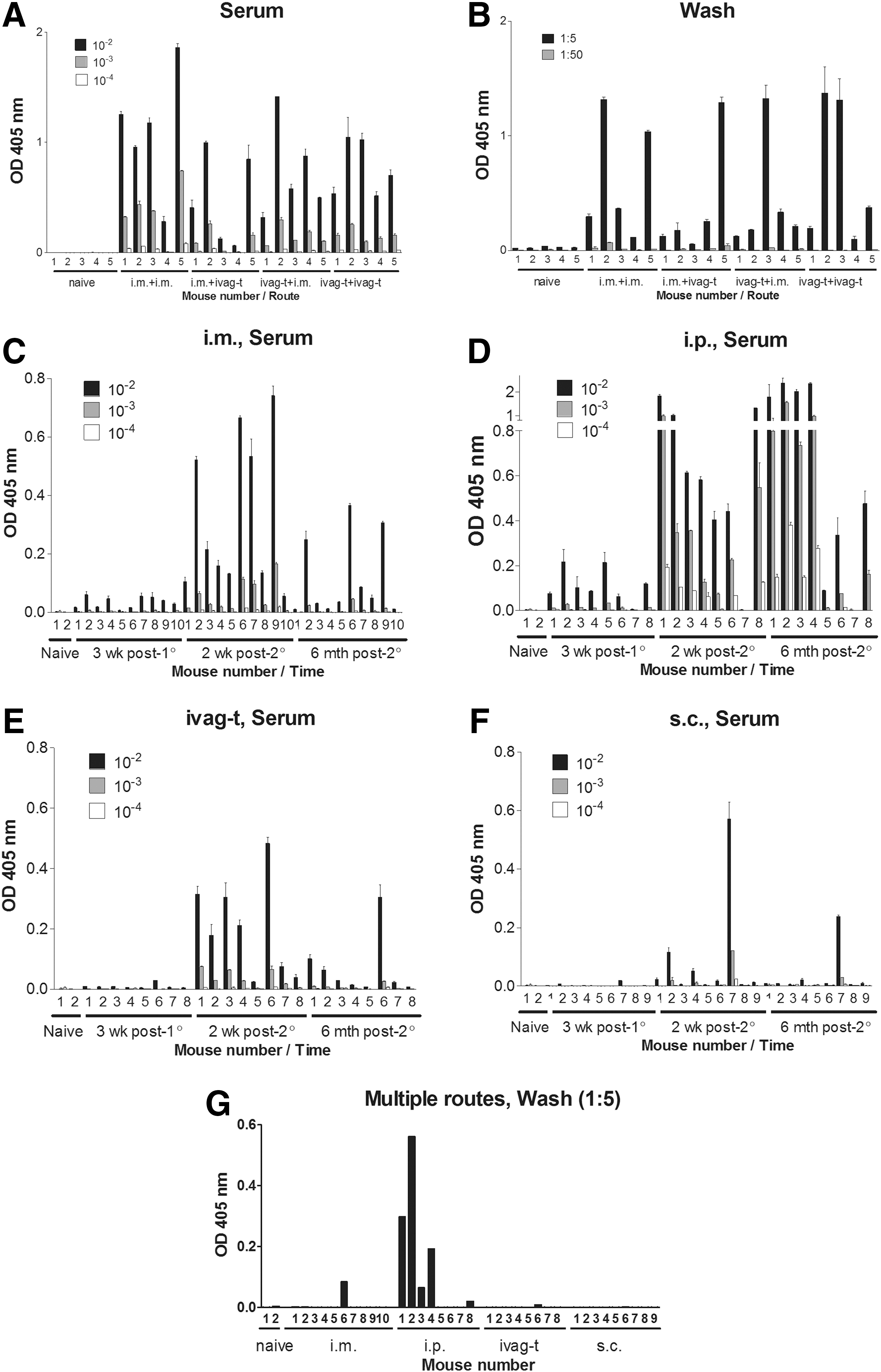
Comparing antibody responses toward HIV-1 envelope protein/alum after immunizations by different routes. HIV-1 envelope-specific antibody responses are shown in sera
As demonstrated in Figure 1A and B, the homologous i.m. immunizations were similar to homologous ivag-t immunizations or heterologous immunizations (i.m. followed by ivag-t, or ivag-t followed by i.m.) for the induction of serum (Fig. 1A) and vaginal wash (Fig. 1B) antibody responses. One-way analysis of variance (ANOVA) with the Tukey Multiple Comparisons Test was performed and showed no significant differences between i.m. and ivag-t immunizations, but showed that antibodies in vaccinated animals were significantly improved compared with controls (p < .05).
When i.p. and s.c. immunizations were added to comparisons, i.p. immunizations were found to be the best at inducing antibody responses in blood and vaginal wash, and s.c. immunizations were weak. Analyses were routinely conducted 2 weeks after the boost, but a preliminary longitudinal experiment was also conducted, in which all four routes of immunization (i.m., i.p., ivag-t, and s.c. by homologous prime and boost) were compared over an extended time course.
As shown in Figure 1C, the i.m. injections elicited responses postboost that waned considerably, but incompletely, by 6 months after the boost. This response was not as strong as that achieved following i.p. injections (Fig. 1D). Responses to ivag-t injections (Fig. 1E) were no better than i.m. injections, and s.c. injections (Fig. 1F) yielded only weak responses. The i.p. vaccination route was the only route that maintained detectable responses in several, but not all, animals in vaginal washes 6 months after the boost (Fig. 1G).
A one-way ANOVA with the Tukey Multiple Comparison Test was conducted using serum results from the 2-week postboost time point for each of the vaccination regimens in the longitudinal experiments (Fig. 1C–F) and demonstrated a significant difference between the i.p. group versus all other groups at each of the three tested serum dilutions (p < .05). The i.p. route was also significantly better than all other routes for the maintenance of antibody responses in vaginal washes 6 months after the boost (Fig. 1G).
Altogether, our results showed that the ivag-t-tissue injection was not superior to i.m. immunizations for the induction of local immunity with envelope protein/alum formulations. Intraperitoneal injections were best of all. These conclusions are made with the understanding that our experiments were conducted only in mice and that numerous untested variables (e.g., the use of a replication-competent vaccine) could have improved immunization outcomes. A recent study by Stary et al. 8 identified an attractive new adjuvant and route that may improve upon old systems for the induction of immune responses in the female reproductive tract. The new vaccination regimen was compared with s.c. vaccinations and was proven superior. A side-by-side comparison between the new strategy and i.m. immunizations is now encouraged.
As stated previously, the vaginal route of immunization has been described as noninductive by some researchers. 3 In some cases, a heterologous prime–boost strategy using a combination of local and systemic routes of immunization might improve outcome, 6,7 but in our study, we found that the homologous i.m. prime–boost was comparable to the heterologous prime–boost for the induction of systemic and local immune responses.
Other vaccination sites [e.g., intranasal (i.n.) or sublingual] were not tested as part of our analyses and may have improved outcome, particularly for the induction of both IgA and IgG (our preliminary tests revealed poor IgA responses in vaginal washes, as predicted by previous literature 3 ). In theory, the engagement of common mucosal lymphoid tissue should assist immune responses in vaginal secretions. 6 We note, however, that Barnett et al. 9 showed that when macaques were immunized by homologous i.m. immunizations, animals were protected from a vaginal challenge with a chimeric simian/human immunodeficiency virus, but homologous i.n. immunizations were not protective.
In conclusion, our results show that the i.m. vaccination route is comparable to the ivag-t route for the induction of systemic antibodies and antibodies in vaginal washes. The i.p. route was better than all others, and the s.c. route was weak. Data suggest that a routine comparison of ivag and i.m. immunizations may serve as a beneficial gatekeeper for the development of new ivag HIV-1 vaccines. The i.m. vaccination route is more attractive because it is simple and is a well-accepted route for vaccine delivery. Experience with the licensed papillomavirus vaccine (GARDASIL, a multivalent vaccine representing nine different virus types) serves as precedent, showing that an i.m. vaccination regimen protects humans from a sexually transmitted disease. 10 Possibly, i.m. injections of a multienvelope HIV-1 vaccine 7 will eventually protect humans from HIV-1 infections and AIDS.
Footnotes
Acknowledgments
This research was supported, in part, by NCI P30-CA21765 and the American Lebanese Syrian Associated Charities.
Author Disclosure Statement
JLH is named on a patent describing a multi-envelope HIV-1 vaccine. RES reports no conflicts of interest.