
Abstract
During their differentiation to antibody-secreting plasma cells, B lymphocytes undergo dramatic changes in metabolism, structure, and function. Here we show that this transition entails extensive intra- and extracellular redox changes. Lipopolysaccharide (LPS)-driven activation and differentiation of naïve murine B splenocytes is paralleled by increased production of reactive oxygen species (ROS) from different sources, followed by a strong antioxidant response. This response includes upregulation of thioredoxin and of the cystine transporter xCT, and increased production and extracellular release of nonprotein thiols, mainly glutathione (GSH) and cysteine. Although ROS levels are higher in late-differentiating B cells, an early oxidative step is likely required to start the differentiation program, because inhibition of NADPH oxidase–dependent early ROS production impairs B-cell activation and differentiation. Addition of reducing agents such as 2-ME results in increased IgM secretion per cell, suggesting that the antioxidant response not only is aimed at restoring the redox homeostasis but also plays a functional role. A highly reduced environment coincident with the presence of large ROS-producing cells is observed in histologic sections of spleens from immunized mice, indicating that the redox modifications observed in LPS-induced B-cell differentiation in vitro occur also in vivo during physiologic immune responses. Antioxid. Redox Signal. 13, 1145–1155.
Introduction
On activation, B cells undergo profound modifications in gene and protein expression necessary to assist the massive immunoglobulin (Ig) production that will take place in plasma cells (9). In a B-cell lymphoma model, the first proteins upregulated during differentiation in vitro are mitochondrial and cytosolic chaperones, followed by metabolic enzymes, required to sustain energy and protein production. Endoplasmic reticulum resident proteins and proteins involved in the redox balance, such as peroxiredoxins and glutathione S-transferase, linearly expand during differentiation (42).
In vivo, B-cell differentiation to plasma cells takes place in the germinal centers of lymphoid organs (21) and is paralleled by strong upregulation of antioxidant systems (7). Remarkably, a hallmark of immunized lymph nodes is the massive production of nonprotein thiols (NPSH) that permeate the lymphoid tissues (7). A number of hemopoietic cells, including dendritic cells (DCs) (2), activated B cells (7), and monocytes (38) have been reported to generate low-molecular-weight thiols in vitro. The mechanism underlying thiol release has been recently described in lymphoma cells and called the “cystine/cysteine redox cycle” (3). This cycle involves (a) expression of xCT, the functional subunit of the Xc-transporter (36); (b) uptake of cystine, the oxidized form of the amino acid cysteine that prevails extracellularly (24); (c) intracellular conversion to reduced cysteine (2, 38); and (d) secretion of cysteine with consequent reduction of the extracellular medium (3, 8, 38).
Typically, upregulation of antioxidant systems represents a defense mechanism against oxidative stress (6). A definite analysis of the oxidative hits occurring along the differentiation process of B cells is so far missing. In principle, ROS production in activated B cells may occur through various sources. First, ROS can be produced by NADPH oxidase, activated by BCR (31) or TLR (29) triggering. ROS might also be generated by the process of oxidative folding (see 25 and references therein). Because Ig molecules are rich in disulfide bonds, this process could result in the production of large amounts of ROS (16). Furthermore, the higher metabolic requirements in plasma cells may cause a corresponding increase in mitochondrial ROS generation. Other enzymatic sources of ROS may include P450 isoenzymes (44) or lipoxygenase (17).
ROS were originally considered only a waste product of cell metabolism that can have dangerous effects. However, compelling evidence showed that ROS are crucial intracellular signaling molecules (14). Similarly, enzymatic or nonenzymatic antioxidant systems, upregulated to counteract ROS, may have a role in signaling (32). For example, the oxidoreductase thioredoxin Trx (22) regulates a number of transcription factors in the B-cell lineage (30). The cystine/cysteine redox cycle, in addition to representing an antioxidant response to an oxidative challenge, serves a signaling function, because it concurs to promote interleukin-1β processing and secretion by LPS-stimulated monocytes (38).
In this study, we discovered the redox changes that occur along the transformation of a resting B cell to an Ab-secreting plasma cell, and investigated the relevant role of oxidative stress and of the antioxidant response in the development and outcome of B-lymphocyte differentiation. Our data indicate that ROS levels increase with differentiation and are counterbalanced by several antioxidant mechanisms. Both ROS production and the subsequent redundant antioxidant response are essential at different steps for an efficient transition of B cells to plasma cells.
Materials and Methods
Chemicals
Diphenylene iodonium (DPI), 5,5'-dithiobis-(2-nitrobenzoic acid) (DTNB), DTT, LPS, 2-ME, N-ethylmaleimide (NEM), cycloheximide (CHX), rotenone, metyrapone, and nor-dihydroguaiaretic acid, were obtained from Sigma-Aldrich. 2',7'-Dichlorofluorescein diacetate (H2DCF-DA), monochlorobimane (MCB), and calcein-AM were from Molecular Probes.
PeroxyGreen1 (PG1) was obtained by Dr. C.J. Chang (27).
Cell preparation and culture
BALB/c mice from Charles River (Lecco, Italy) were maintained in pathogen-free facilities. All protocols were approved by the Institutional Review Board of the Istituto Nazionale per la Ricerca sul Cancro (Genova, Italy). Spleens were removed from dead mice, and B cells were purified by positive selection on MACs columns by using anti–CD19-conjugated magnetic beads (Miltenyi Biotec), according to the manufacturer's instruction. Selected B cells were plated (1 × 106/ml) in complete RPMI supplemented with 50 μM 2-ME and activated with 20 μg/ml LPS (28, 42).
Intracellular ROS and reduced GSH detection
At each time point, cells were washed and resuspended in PBS containing 10 μM H2DCF-DA to assess intracellular ROS (10) or 100 μM MCB, to assess intracellular GSH (11). After incubation of 20 min in the dark, cells were washed, resuspended in PBS, and FITC fluorescence (H2DCF-DA) or Violet-1 fluorescence (MCB) was immediately measured with a flow cytometer (CyAn, Beckman Coulter) and analyzed by using Summit V4.3 software. In double-staining experiments, cells were incubated in PBS containing both fluorescent dyes. H2DCF-DA was trapped inside cells by deesterification (40) and was also used to distinguish viable cells. To correct for any increase in fluorescence due to increased cell volume, cells were loaded in parallel with 10 μM calcein-AM and analyzed with flow cytometry (4, 18).
ELISA
IgM content in supernatants was determined with ELISA by using the Mouse IgM Quantitation Kit (Bethyl Laboratories) according to the manufacturer's instructions.
Determination of NPSH in culture media
Supernatants (0.1 ml) of murine B cells plated at 1 × 106/ml in 24-well plates in complete RPMI medium for different times were reacted with 10 mM DTNB, and the absorption was measured at 412 nm (38).
Real-time PCR
Total RNA was isolated from murine B-cells by using TriPure Isolation Reagent (Roche) and reverse-transcribed by Superscript III Reverse Transcriptase (Invitrogen), according to the manufacturer's instructions. Real-time PCR determination of cDNA was performed by using SYBR greenER qPCR Super Mix for iCycler reagent (Invitrogen). The specific primers used were forward 5′-aaacccaagtggttcagacg-3′ and reverse 5′-atctcaatcctgggcagatg-3′ (for xCT), forward 5′-atggccttccgtgttcctac-3′ and reverse 5′-gcttcaccaccttcttgatgtc-3′ (for GAPDH). Relative expression was determined by using the ΔCt method (23).
Western Blot analysis
Triton X-100 cell lysates were resolved on 12% SDS-PAGE and electrotransferred (38). Filters were probed with rabbit anti Trx (S3.1; kind gift of Dr. F. Clarke, Brisbane, Australia) and rabbit anti heme-oxygenase-1 (HO-1; Stressgen) polyclonal antibodies, or anti β-actin monoclonal Ab (Sigma-Aldrich), followed by the relevant secondary Ab (GE Healthcare), and developed with ECL-plus (GE Healthcare).
Immunization
Twelve-week-old animals (three per group) were immunized with four weekly i.p. injections of 2 × 108 sheep red blood cells (SRBC; 7). One week after the last injection, mice were killed, and the spleens removed and analyzed. All protocols were approved by the Institutional Review Board of the Istituto Nazionale per la Ricerca sul Cancro (Genova, Italy).
Staining procedures and confocal analyses
Serial cryostat sections of frozen spleens from immunized or not immunized mice were double stained with the sulfhydryl-reactive dye, Mercury Orange, as previously described (7) and with the H2O2-specific dye PG1 (27), or with the ROS-specific dye H2DCF-DA (39) immediately after sectioning. Mercury Orange was incubated 5 min on ice at a final concentration of 25 μM. To confirm the specificity of Mercury Orange binding to NPSH, control sections were pretreated with 100 μM NEM for 10 min to block thiol groups. Staining with PG1 (5 μM) was performed for 10 min at 37°C. Staining with H2DCF-DA (10 μM) was performed for 30 min at 37°C. To confirm the specificity of PG1 or H2DCF-DA staining, control sections were pretreated with 1 mM DTT for 10 min. The sections were analyzed with confocal microscopy, and the images were acquired with the Fluoview FV500 software.
For double immunofluorescence, rabbit anti-mouse Trx (S3.1), rabbit anti-HO-1 (Stressgen), rat anti-CD45R B cells (R&D), and rat anti-F4/80 tissue macrophage (19, Novus Biologicals) antibodies were used. Rhodamine (TRITC)-labeled anti-rat IgG or FITC-labeled anti-rabbit IgG (Jackson Immunoresearch) were used as secondary Abs. Images were acquired with Leica DM LB-2 microscopy by using Scion Image software.
Statistical analysis
The data were statistically analyzed with a one-way ANOVA test, followed by Bonferroni posttest, or with an unpaired t test by using GraphPad software.
Results
ROS production during B cell differentiation
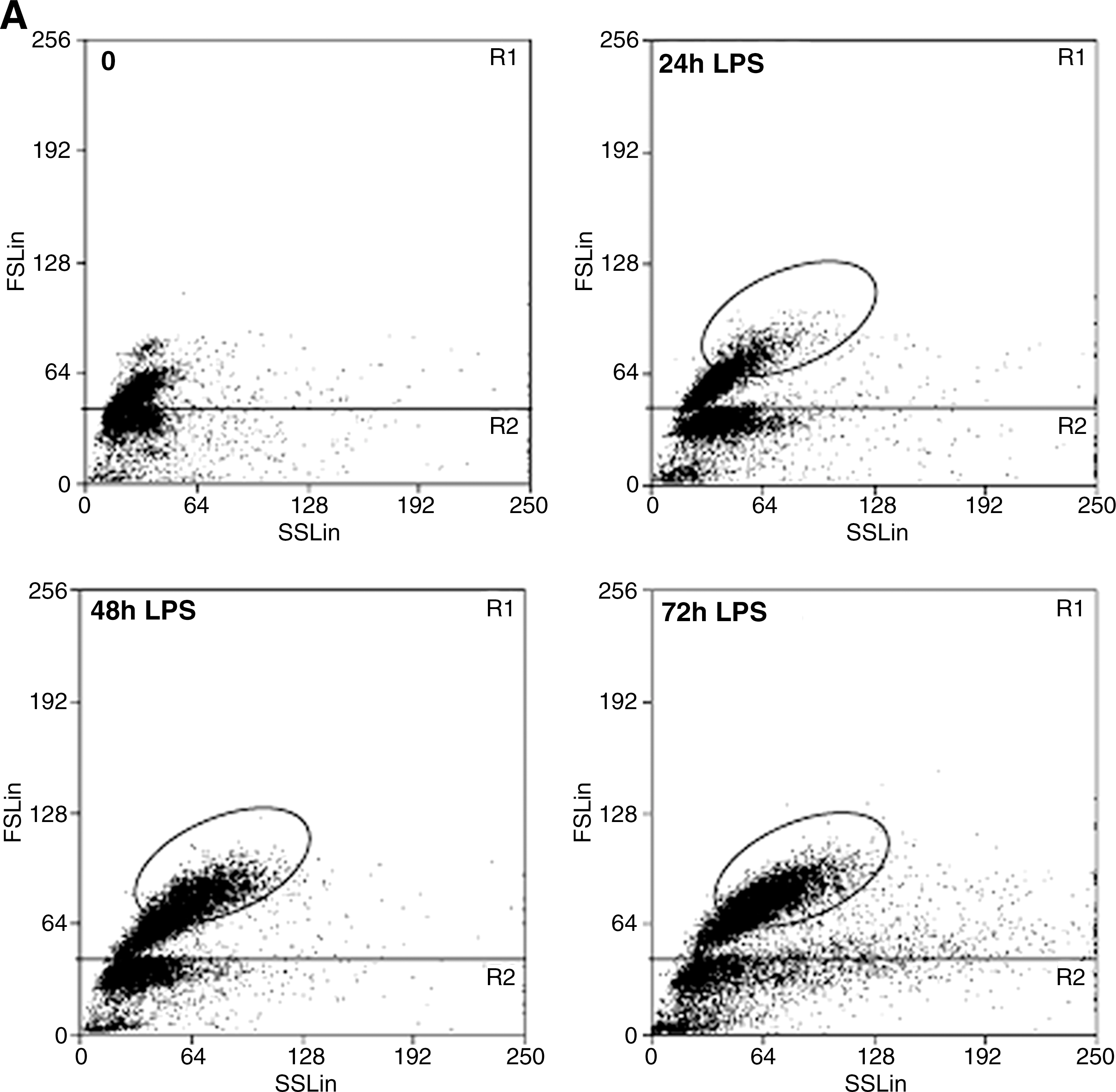
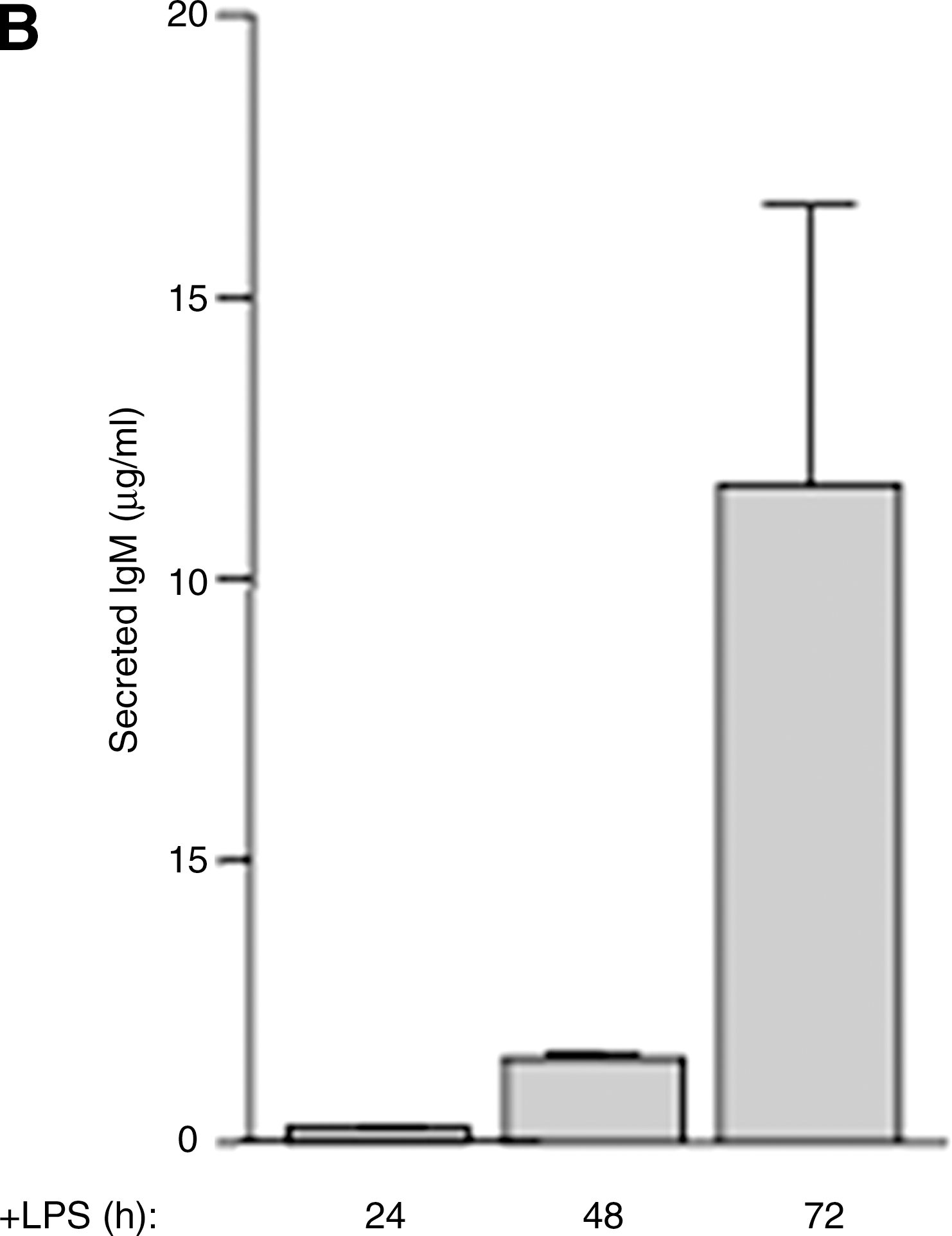
LPS stimulation in vitro of B splenocytes from naïve mice results in activation of a fraction of B cells (28). As shown in Fig. 1A, activated B lymphocytes (R1, encircled) increase in size and complexity, appearing as a rather homogeneous population of cells larger than resting B cells that emerges 24 h from LPS stimulation and increases thereafter. Nonactivated B cells remain of small dimensions and low complexity (R1, not encircled) or die (R2). Analysis of IgM secretion revealed a dramatic increase at day 3 (Fig. 1B), in agreement with the typical kinetics of secretion induced by LPS in vitro (42).
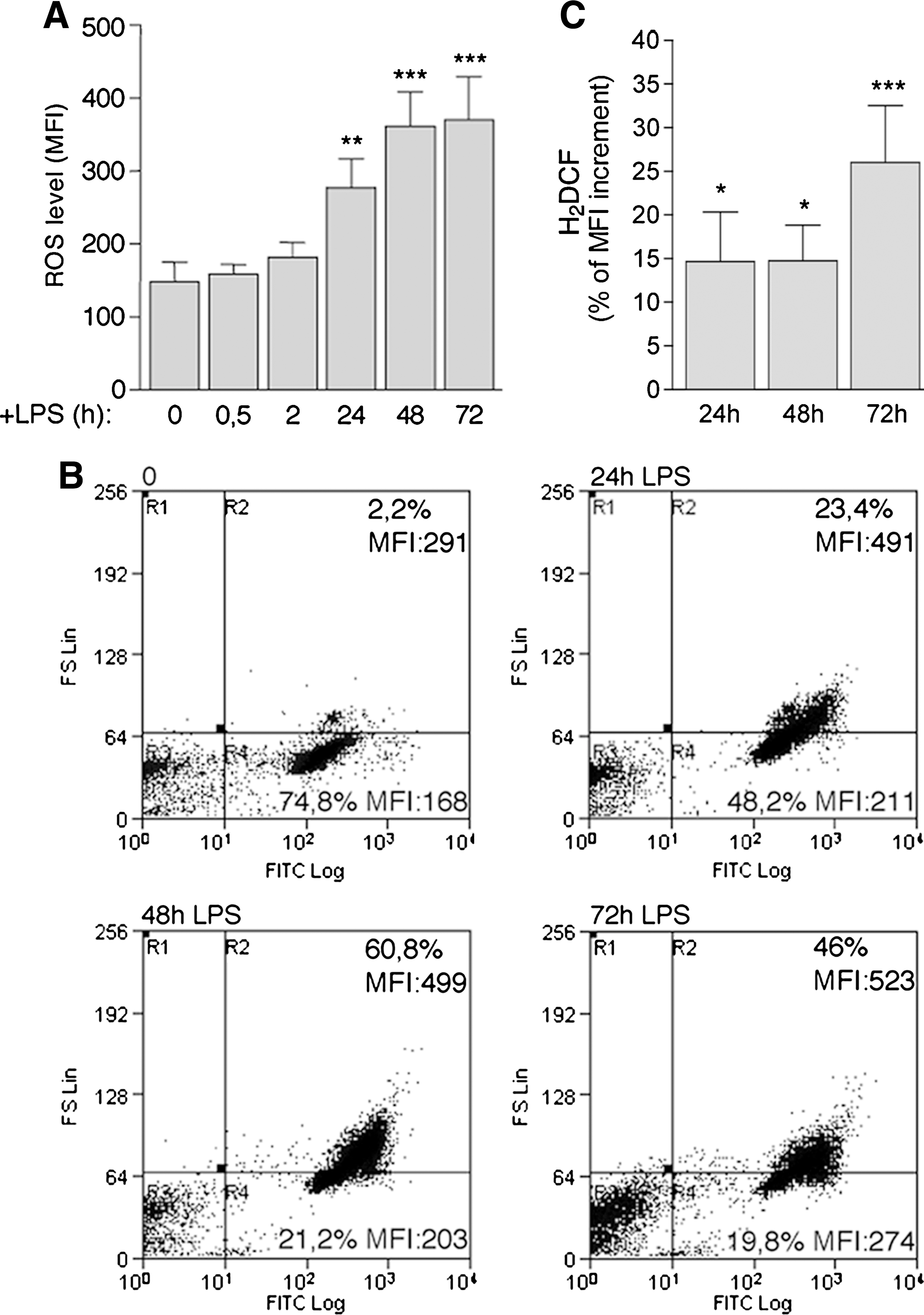
To assess ROS production, resting B splenocytes were exposed to H2DCF-DA at various times from LPS stimulation. As shown in Fig. 2A, ROS were only slightly increased in the first 2 h of exposure to the mitogen. A significant enhancement in ROS levels was detected after 24 h of culture and increased along with B-cell differentiation. Although all living cells stained with the vital dye H2DCF-DA (Fig. 2B, R2 and R4), small cells (Fig. 2B, R4) were less positive, and the highest levels of ROS were found in the large, differentiating B cells (Fig. 2B, R2). To correct for the fluorescence increase merely due to enlarged cell volume, B cells were loaded in parallel with the vital dye calcein-AM (4, 18), and the H2DCF-DA fluorescence increase at each time point was normalized versus the increase in calcein-AM (Fig. 2C). The results confirmed that the increment of H2DCF-DA fluorescence is due to increased ROS production along with B-cell differentiation (Fig. 2C and Supplemental Fig. 1; see
Different mechanisms concur to ROS production
B cells at various times from LPS stimulation were treated for 3 h with compounds that inhibit ROS generation by different sources, namely DPI, which blocks NADPH oxidase (15); rotenone, which inhibits mitochondrial ROS production (13); or with the protein-synthesis inhibitor cycloheximide (CHX). At the end of the incubation, cells were loaded with H2DCF-DA, and ROS production was assessed with FACS analysis. DPI decreased ROS generation at all times (24, 48, and 72 h from LPS stimulation), resulting in a 30–40% of inhibition with respect to control cells (Fig. 3A). Similarly, rotenone affected ROS production at all times, causing a decrease of 20–30%, whereas CHX resulted in only a slight reduction in ROS generation (Fig. 3A). Metyrapone, a potent inhibitor of cytochrome P450 (44), and nor-dihydroguaiaretic acid, which blocks the lipoxygenase activity (17), barely affected ROS production, even at high dosage (data not shown).
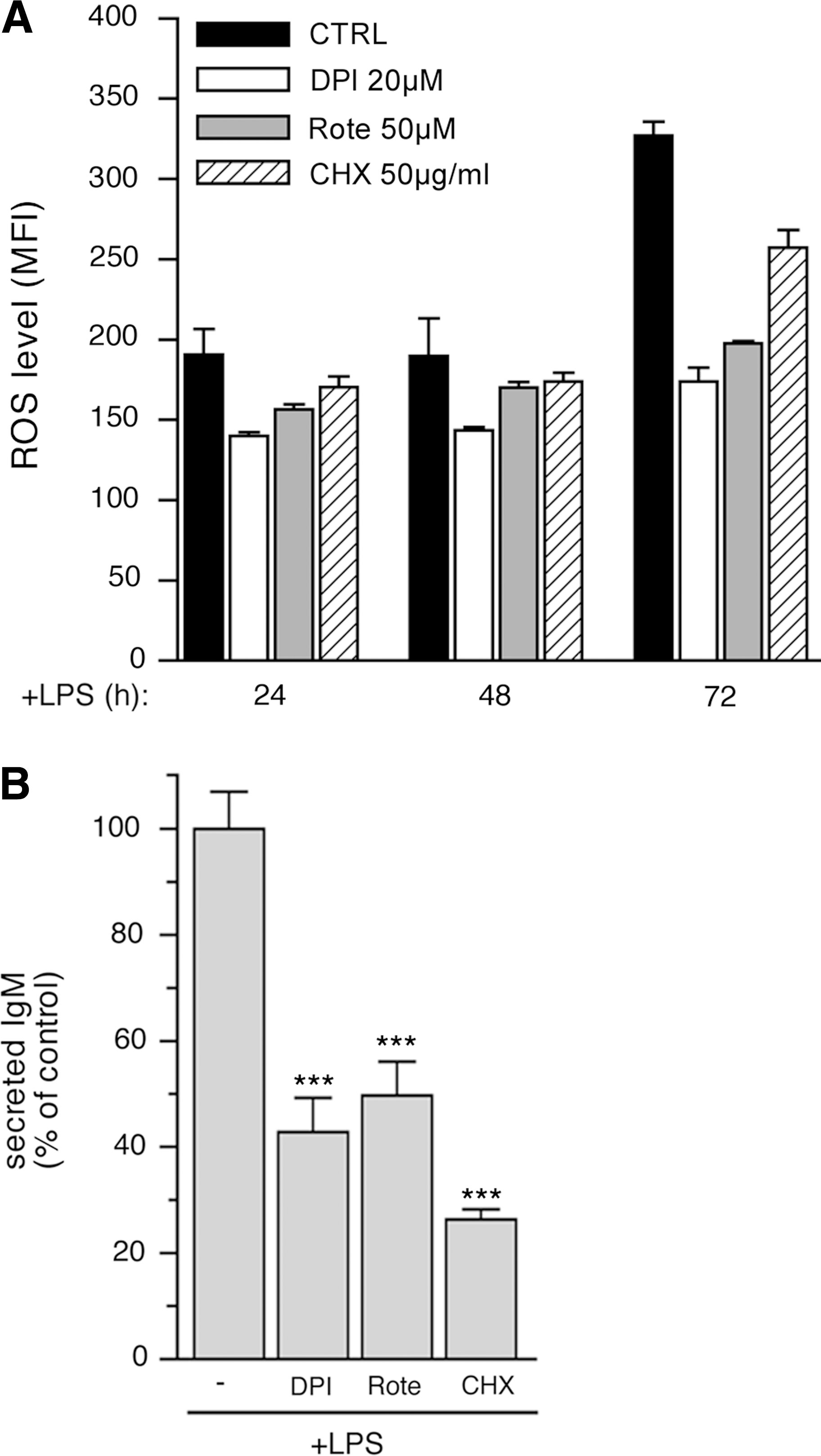
As expected, CHX treatment strongly inhibited IgM secretion (Fig. 3B). The decreased ROS production observed in 72-h LPS-stimulated B cells during 3 h of exposure to DPI and rotenone (Fig. 3A) also correlated with a decreased IgM secretion (Fig. 3B).
Activation of antioxidant responses in ROS-producing cells
Next, we examined the activation of antioxidant systems in differentiating B cells. As shown in Fig. 4, the levels of GSH, the major intracellular electron donor (26), doubled in the first 24 h of stimulation with LPS and increased thereafter (Fig. 4A and B). The highest GSH increase was observed in larger, differentiating cells (Fig. 4C, R2). Remarkably, double staining revealed that the same cells that produce more ROS also produce more GSH (Fig. 4D). Normalization versus calcein-AM staining (4, 18) at each time point confirmed that the strong enhancement in MCB fluorescence was actually due to a dramatic increase in GSH, more evident at 48 h from stimulation (Fig. 4E).
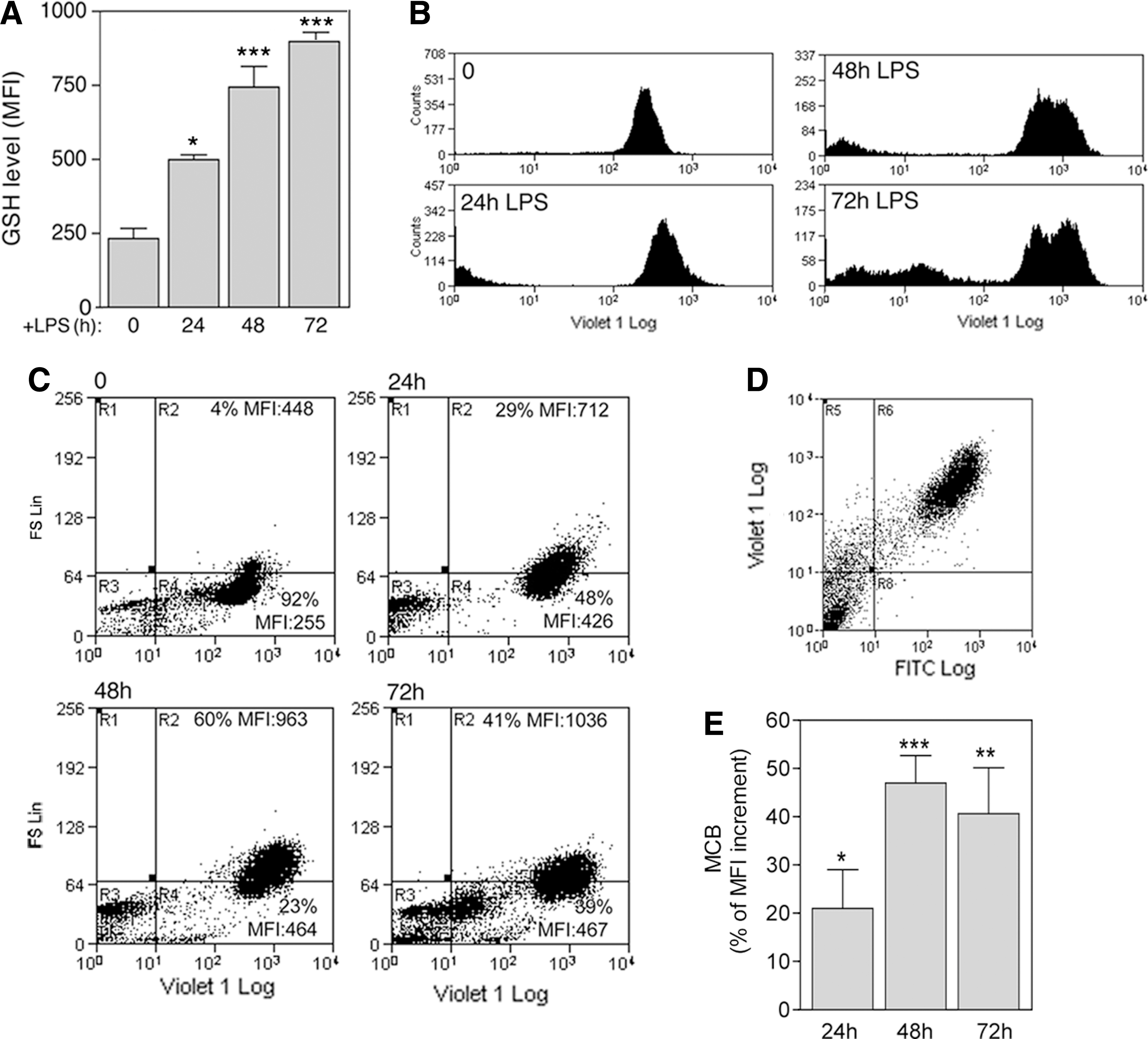
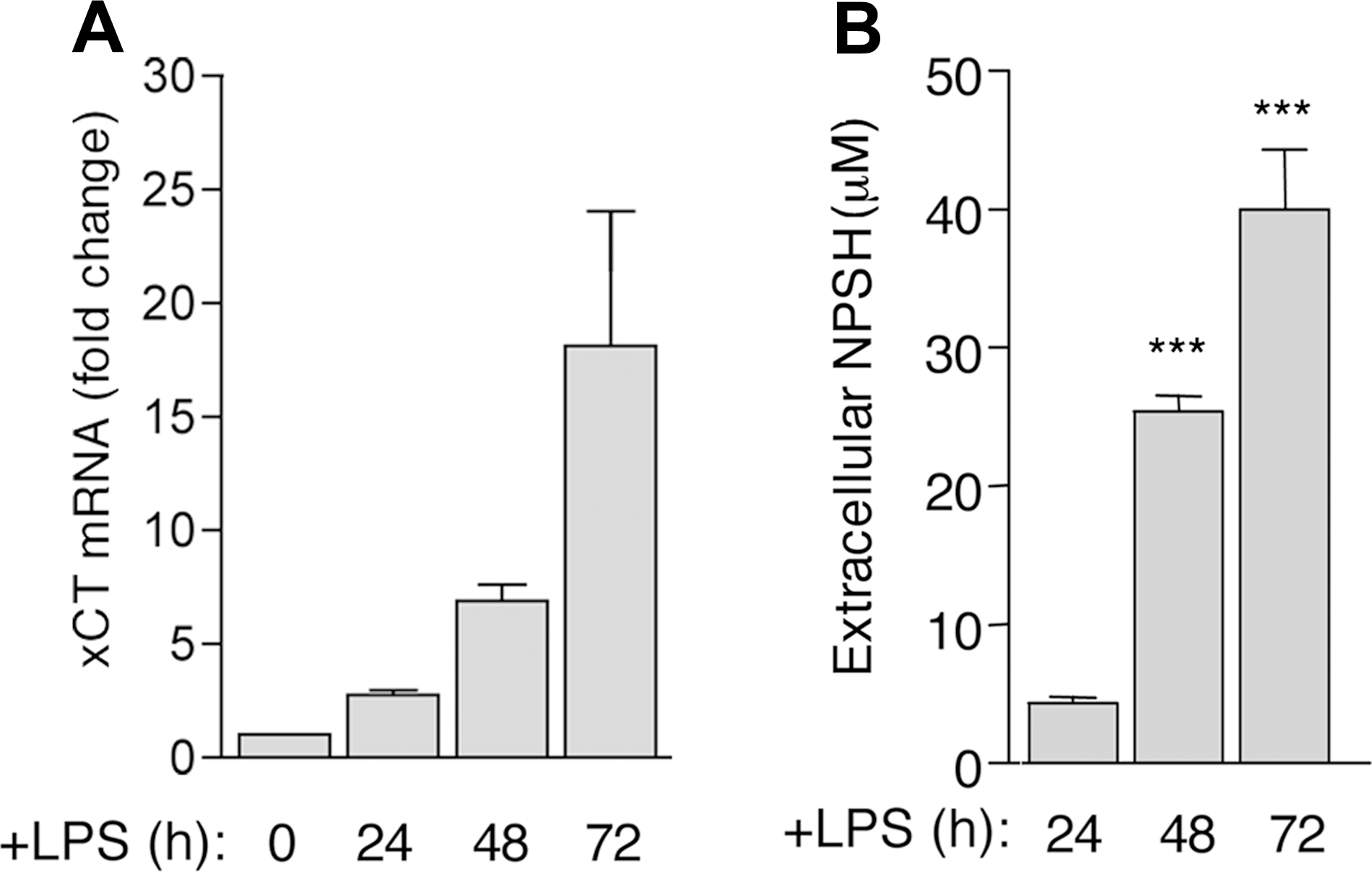
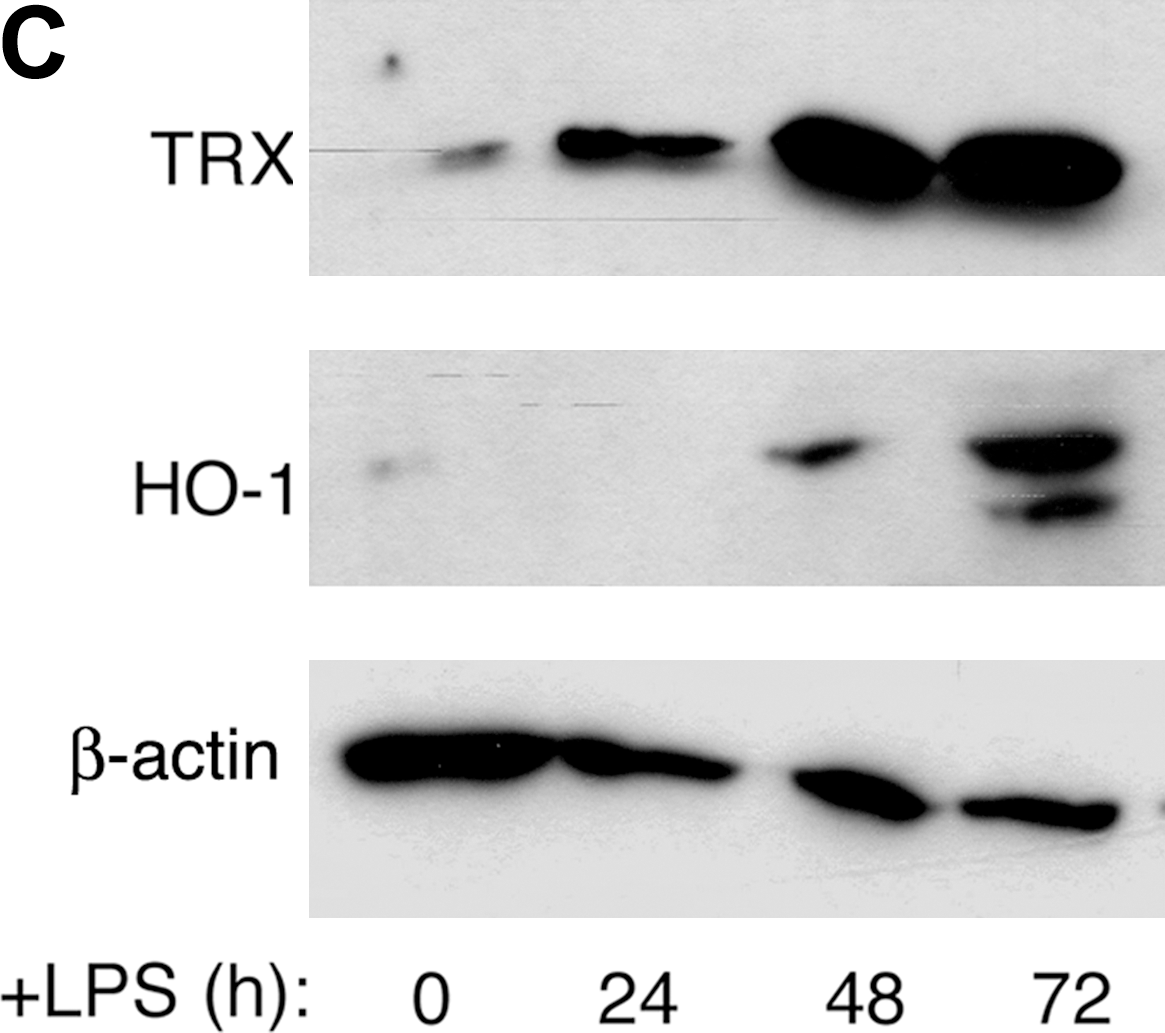
In addition to GSH, the cystine/cysteine redox cycle was upregulated in differentiating B cells, with a delay compared with ROS induction. Expression of xCT, very low or absent in resting B cells (7), was induced by LPS and increased along with B-cell differentiation (Fig. 5A). Likewise, extracellular cysteine accumulated significantly 48 h from exposure to LPS and increased thereafter (Fig. 5B). Intracellular Trx was also strongly increased in differentiating B lymphocytes (Fig. 5C), with kinetics that paralleled cysteine release (panel B). The oxidant-responsive enzyme HO-1 (34) was upregulated in LPS-stimulated B cells, and its expression was delayed compared with that of thioredoxin.
Different redox requirements in the early and late phases of terminal B-cell differentiation
NADPH-oxidase–dependent ROS generation has been proposed as a crucial event in B-cell activation (31). Our assays, however, did not reveal significant ROS increases early after LPS stimulation (from 5 min to 2 h; Fig. 2A and data not shown), possibly because of insufficient sensitivity of H2DCF-DA or rapid ROS scavenging. To investigate further the potential signaling role of ROS in the early activation phases, B cells were exposed to LPS in the presence of DPI, to block NADPH-oxidase–dependent ROS production. DPI was highly toxic for freshly isolated resting B cells that died at DPI doses not affecting the viability of different white blood cells (38; data not shown). Therefore, lower concentrations (100 and 200 nM) were used to limit cytotoxicity. DPI-treated B cells displayed a mild decrease in ROS levels both at 24 and 48 h (Fig. 6A). Moreover, the amount of secreted IgM, normalized on a “per viable cell” basis, was reduced in DPI-treated B-cell culture (Fig. 6B).
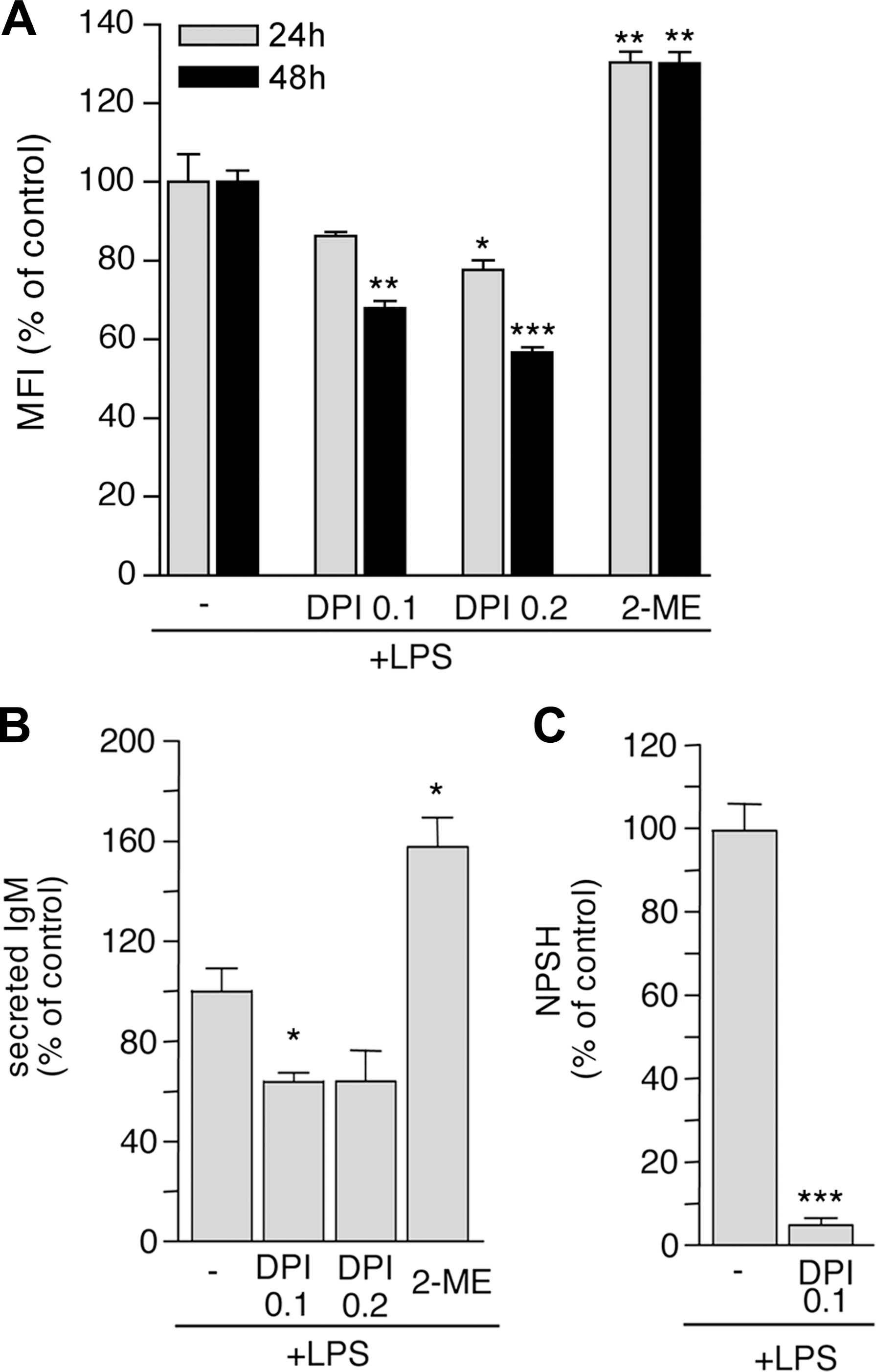
Exposure to DPI also resulted in a decreased cysteine release by LPS-treated B lymphocytes (Fig. 6C), supporting the hypothesis that the upregulation of antioxidant systems occurring in differentiating B lymphocytes represents a response to an LPS-induced oxidative hit. To assess whether the antioxidant response affects differentiation, B lymphocytes were stimulated with LPS in the presence of 0.8 mM 2-ME. Under these conditions, the percentage of cells dying at 24 h of culture shows increases of 15–25% in the different experiments (data not shown), so that the number of differentiating B cells is smaller. However, Ab-secreting cells generated in the presence of 2-ME displayed a higher IgM secretion rate than did controls (Fig. 6B).
Oxidative and reducing events in B cell follicles of naïve or immunized spleen
To investigate the sites of ROS and NPSH accumulation in lymphoid organs in vivo, spleens from naïve or SRBC-immunized mice were stained with H2DCF-DA (that reacts mainly with H2O2 and peroxynitrite), PG1 (which specifically stains H2O2), or with the free thiol-binding dye Mercury Orange. Nonimmunized spleens did not stain with PG1 (Fig. 7A) or H2DCF-DA (not shown), and displayed a weak staining with Mercury Orange (Fig. 7A). In contrast, spleens from immunized animals showed a diffuse staining with PG1 (Fig. 7B) or with H2DCF-DA (Fig. 7C), which was stronger in clusters of cells within germinal centers (Fig. 7G). Remarkably, cells highly stained with the ROS probes were also the most positive to Mercury Orange (Fig. 7B and C). The specificity of the ROS and NPSH staining was confirmed by the control experiments shown in Fig. 7D–F. ROS staining by PG1 and H2DCF-DA was prevented by incubating spleen sections with the reducing agent DTT (Fig. 7D and E), whereas Mercury Orange staining was prevented by treatment of spleen sections with NEM, which blocks free thiols (Fig. 7F).
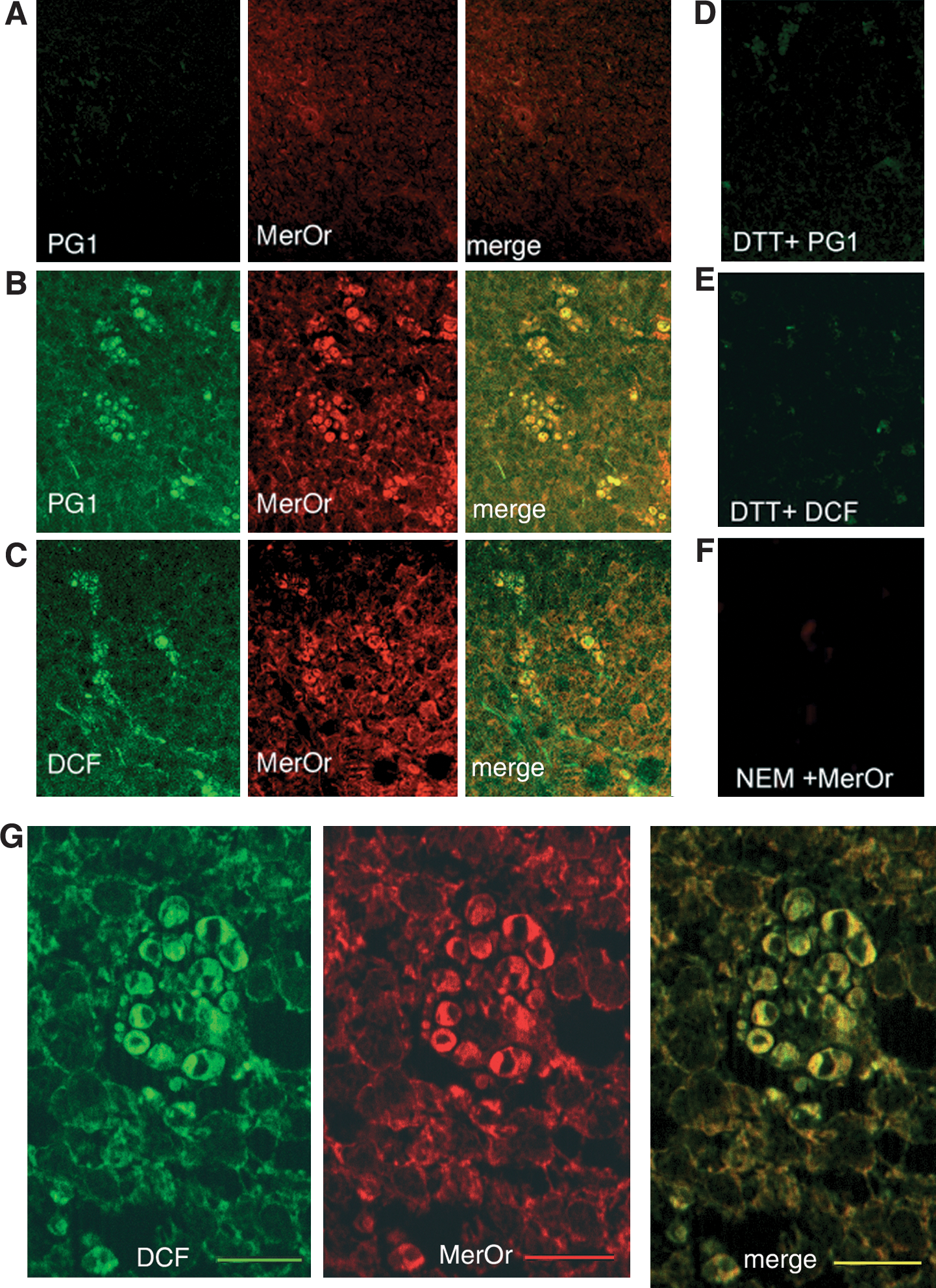
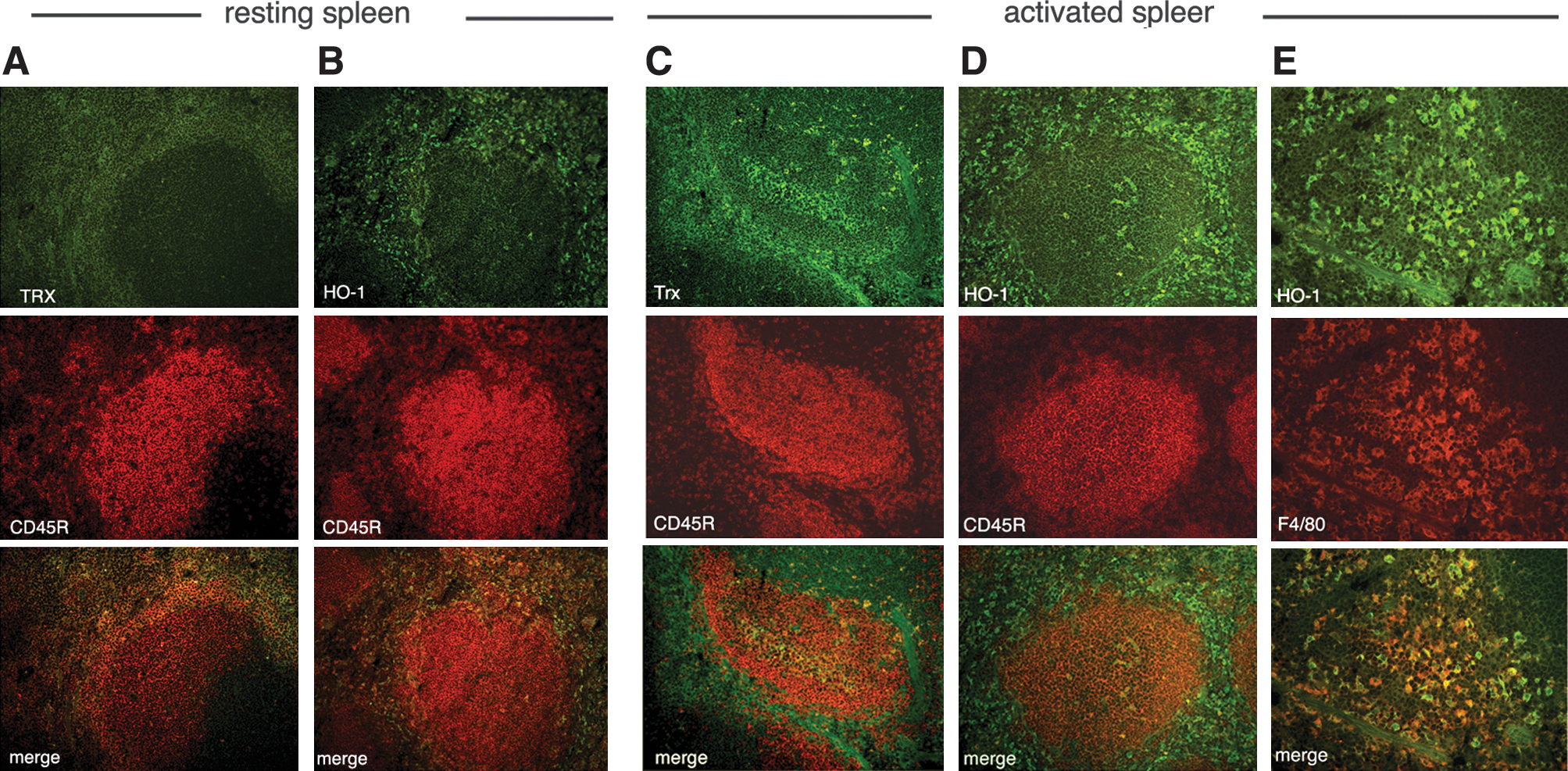
Spleen sections from nonimmunized (Fig. 8A and B) and immunized (Fig. 8C–E) mice were then analyzed with double immunofluorescence with anti-Trx (Fig. 8A and C) or anti-HO-1 (Fig. 8B, D, and E) and CD45R (Fig. 8A–D) or F480 (Fig. 8E) antibodies. A mild positivity for Trx was found in nonimmunized spleens, restricted to the interfollicular areas, but absent from the B-cell follicle (Fig. 8A). In immunized spleens, Trx staining was increased in the interfollicular areas and was clearly strongly detected also in groups of differentiating B cells within the germinal centers (Fig. 8C). Trx staining appeared slightly diffuse, probably because of its secretion and extracellular diffusion (33). HO-1 was mildly detected in the interfollicular areas of nonimmunized spleens (Fig. 8B), whereas it was strongly positive both in interfollicular cells and in a few germinal-center B cells (Fig. 8D). Interfollicular HO-1–positive cells were not stained by markers of B cells, T cells, granulocytes, DCs, and macrophages of the T-cell areas (not shown) but were strongly positive for the red pulp macrophage marker F4/80 (Fig. 8E).
Discussion
We followed the oxidant and antioxidant events occurring in B lymphocytes during their transition to plasma cells and found that ROS generation accompanies the differentiation process and is counterbalanced by a strong upregulation of antioxidant defense mechanisms.
The increase in ROS, detected since 24 h from LPS stimulation, was hindered by inhibitors of protein synthesis and of different ROS-generating systems, including NADPH oxidases and the mitochondrial electron-transport chain. The degree of inhibition observed at all times with the single drugs ranged between 15% (with CHX) and 35% (with DPI or rotenone), indicating that ROS are simultaneously produced by different intracellular sources throughout differentiation. In addition, the modest decrease in ROS levels induced by CHX suggests that neither the high rates of Ig biosynthesis nor the process of Ig oxidative folding is responsible for the strong ROS increase. Rather, the observation that ROS are generated at high levels even when Ig production is blocked is in agreement with previous findings that, on activation, B cells expand their metabolic capacity and secretory machinery well before IgM secretion starts (42). The increased energetic metabolism and the massive expansion of the secretory apparatus may account per se for ROS production.
ROS increase is followed by upregulation of various antioxidant systems (6). We focused on three of these systems, different although related: intracellular glutathione, antioxidant enzymes (Trx and HO-1), and the cystine/cysteine redox cycle. Glutathione, the principal redox buffering system in living cells (26), seems to act as a first line of defense against oxidative stress in LPS-activated B lymphocytes, because it increases early, almost simultaneous with ROS. The increase of the key redox-regulating protein Trx (22) is dramatic but delayed with respect to GSH, confirming also in primary B lymphocytes the observation that Trx levels are lower in B-cell than in plasmacytic tumor cell lines (30). The increased expression of the HO-1 (35) late along the differentiation process of B cells is a novel finding. Indeed, although HO-1 is emerging as a key regulator of T cell–mediated immunity (43), no data on expression or function in B cells have been reported. Because this enzyme has been proposed as an antiapoptotic factor (5, 37), its expression could be finalized to delay in the death of antibody-secreting cells.
In addition to GSH and oxido-reductases, we identified a third antioxidant system upregulated in differentiating B lymphocytes, the cystine/cysteine redox cycle. The increase in this cycle is a key event in T-cell activation by antigen-presenting DCs (2) and IL-1β secretion by monocytes (38). Its importance in redox homeostasis was recently highlighted also in tumors (3, 8).
Our data show that blocking NADPH-oxidase–dependent ROS production with DPI impairs B-cell activation/differentiation as well as the efficiency of IgM secretion. This result is in line with the previous suggestion (31) that H2O2 may act as a secondary messenger in the initiation and amplification of signaling after BCR triggering. However, we failed to detect ROS increase in the first hours after LPS stimulation. This is possibly due to a low increase of ROS over basal levels, undetectable by standard methods, or to a too-rapid ROS scavenging in this phase. An alternative explanation is that other cells, such as monocytes or granulocytes, that undergo an oxidative burst on activation, provide ROS for activating the neighboring lymphocytes (31). In our culture conditions, however, granulocytes were virtually absent, and macrophages consistently were <1% (not shown). Moreover, LPS stimulation in the presence of splenic macrophages increased neither the number of B cells entering the differentiation program nor the efficiency of IgM secretion (not shown).
A novel finding of this study is that not only the oxidative burst but also the antioxidant responses are implicated in the differentiation of B lymphocytes. When the reducing agent 2-ME is added to resting B cells at the beginning of the culture, the number of B cells that survive and differentiate to LPS is severely reduced, suggesting that highly reducing conditions are detrimental in the early phases of B-cell activation.
Conversely, the rate of IgM secretion per cell is increased in the presence of 2-ME, indicating that, later on during differentiation, reduction of the microenvironment promotes IgM secretion. This effect is likely due to the facilitated transport of Ig-assembly intermediates along the exocytoxic pathway. It was previously observed that the addition of reducing agents to the culture medium, at concentrations that do not affect cell viability and do not cause disassembly of IgM monomers or unfolding of individual Ig chains, allows secretion of unpolymerized IgM otherwise retained in the ER by disulfide-interchange reactions (1, 41).
In vitro experimental systems of B-cell activation and differentiation provide many advantages, including the possibility for study of purified cell populations and the determination of the kinetics of the events that follow LPS stimulation. However, many factors may alter the correct development of the Ab response in vitro, such as the unavoidable cell manipulations that stress the cells and may change their redox, and the high oxygen tension that can impair significantly the redox regulation. Furthermore, the redox changes observed in vitro may be restricted to the B-cell population responsive to TLR4 triggering. To circumvent these constraints, we studied spleens from nonimmunized or SRBC-immunized animals. These experiments demonstrated that a dramatic redox reshaping during B-cell differentiation occurs also in response to a specific antigen in the complex choreography of the immune response in vivo. Confocal fluorescence analyses revealed that a minority of B cells in the germinal center produce high levels of ROS, and in all cases, these cells also increased levels of NPSH, thereby confirming the co-staining of ROS and GSH observed in differentiating B cells in vitro. The finding that the same cells concomitantly stain with reagents specific for ROS and antioxidants is only apparently discrepant. In a given cell, as long as the oxidative stress persists, antioxidant responses must continue. Moreover, NPSH, revealed by Mercury Orange, may not neutralize ROS directly but rather may protect targets of ROS from oxidation. Support for this interpretation comes from the in vitro finding that ROS detection is not impaired in the presence of 2-ME.
Interestingly, in addition to the few B cells highly reacting to the dyes specific for ROS and NPSH, all germinal centers were diffusely stained. This observation suggests that ROS and NPSH are not confined within the producing cells but are released in the microenvironment, where they diffuse and generate redox gradients. NPSH is actively secreted by differentiating B lymphocytes through the cystine/cysteine cycle, and ROS can be generated extracellularly by NADPH oxidases and perhaps secreted as well.
In vitro, differentiated B lymphocytes express high levels of TRX and HO-1. Interestingly, in immunized spleen sections, these enzymes are highly expressed, not only in B cells in the germinal centers, but also in red-pulp macrophages. This finding suggests that antioxidant enzymes, besides affording cytoprotection against oxidative injury (20), may represent a barrier to segregate B follicles from each other, preserving the intrafollicular redox from external stress. In the first phases of the soluble immune response, when Ag-specific B cells are triggered, resulting in oxidative stress within the developing germinal center, HO-1 and Trx may block the diffusion of ROS to other follicles. This fence may prevent premature redox responses in other follicles that could impede correct B-cell activation, in keeping with the in vitro observation that the presence of 2-ME reduces the number of B cells that survive and differentiate in response to LPS.
In conclusion, we showed that intra- and intercellular redox remodeling occurs during terminal B-cell differentiation. We propose that ROS production and the consequent antioxidant responses serve signaling and homeostatic functions necessary for full-fledged plasma cell differentiation.
Footnotes
Acknowledgments
We thank Dr. P. Castagnola for help with confocal analyses, Dr. F. Clarke for the generous gift of anti TRX, and Dr. C.J. Chang for kindly providing PeroxyGreen 1. This work was supported in part by grants from Ministero della Salute, Ministero Università e Ricerca, ISS, Cariplo, Compagnia di San Paolo, Telethon.
Author Disclosure Statement
No competing financial interests exist
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.