
Abstract
The free radical theory of aging proposes a causal relationship between reactive oxygen species (ROS) and aging. While it is clear that oxidative damage increases with age, its role in the aging process is uncertain. Testing the free radical theory of aging requires experimentally manipulating ROS production or detoxification and examining the resulting effects on lifespan. In this review, we examine the relationship between ROS and aging in the genetic model organism Caenorhabditis elegans, summarizing experiments using long-lived mutants, mutants with altered mitochondrial function, mutants with decreased antioxidant defenses, worms treated with antioxidant compounds, and worms exposed to different environmental conditions. While there is frequently a negative correlation between oxidative damage and lifespan, there are many examples in which they are uncoupled. Neither is resistance to oxidative stress sufficient for a long life nor are all long-lived mutants more resistant to oxidative stress. Similarly, sensitivity to oxidative stress does not necessarily shorten lifespan and is in fact compatible with long life. Overall, the data in C. elegans indicate that oxidative damage can be dissociated from aging in experimental situations. Antioxid. Redox Signal. 13, 1911–1953.
Long-Lived Mutants Show Increased Resistance to Oxidative Stress
Screening for Mutants with Altered Sensitivity to Oxidative Stress
I. Introduction
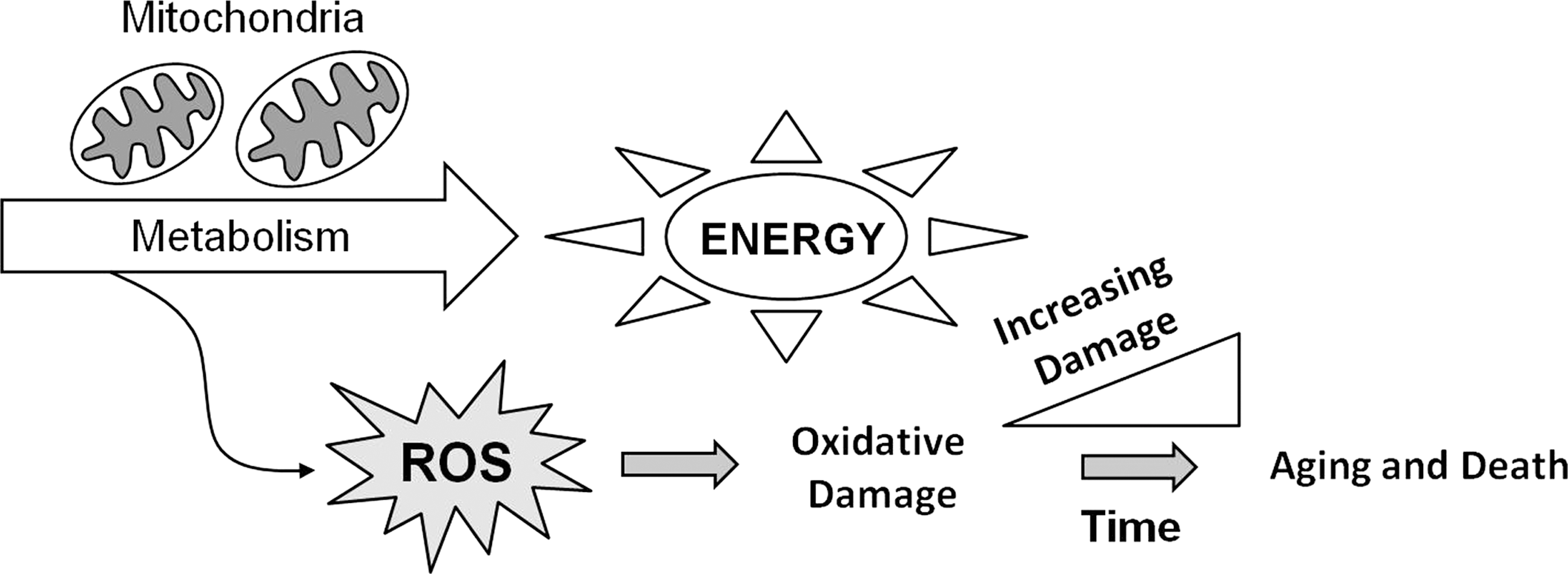
A possible mechanism that would account for the observations of Rubner and Pearl was suggested by the free radical theory of aging. This theory, which was first proposed by Denham Harman in 1956, suggests that aging results from the accumulation of molecular damage caused by by-products of normal metabolism called reactive oxygen species (ROS) (defined below) (Fig. 1) (96). Combining the two theories results in the idea that an individual, or a species, with a higher metabolic rate will produce more ROS (assuming that a fixed percentage of electrons undergoing electron transport are leaked to form ROS), accumulate oxidative damage more rapidly, age faster, and thereby have a decreased lifespan. When it was suggested that mitochondria are the main producers of ROS within the cell, Harman refined his theory to highlight the role of the mitochondria in what is called the mitochondrial theory of aging (97). This theory is also referred to as the oxidative stress theory of aging (251). Here, these theories will collectively be referred to as the free radical theory of aging. Since its formulation, the free radical theory of aging has been the most widely tested theory of aging with numerous experiments both supporting and refuting its claims [reviewed in refs. (168, 207)]. Thus far, a consensus on whether or not oxidative damage is responsible for aging has not yet been achieved. This review will provide a summary of experiments in the worm Caenorhabditis elegans that have examined the relationship between ROS and aging.
II. Reactive Oxygen Species
ROS are highly reactive molecules that contain at least one atom of oxygen. While it is generally assumed that the majority of ROS production occurs in the ETC of the mitochondria, this has not been directly shown in vivo due to technical limitations (166, 212). Attempts to estimate ROS production in isolated mitochondria have yielded results suggesting that between 0.12% and 2% of all oxygen consumed is converted to ROS (24, 39, 95, 162, 253), though these results may overestimate the amounts of ROS generated in vivo under normal conditions (212). Nonetheless, it is clear that, at least under certain conditions, some of the electrons transferred in the ETC escape to form ROS.
While the production of ROS has typically been considered an unwanted side reaction in the production of energy by the ETC, the fact that ROS have been demonstrated to have a role in signaling (see below) suggests the possibility that generation of ROS could be used as a signal to detect the functional status of the ETC. Nonetheless, ROS have the potential to damage all parts of the cell. To limit this damage, cells also have numerous antioxidant enzymes and nonenzymatic antioxidants, which detoxify ROS. This section provides a general description of the generation of ROS, elimination of ROS, and damage caused by ROS. Note that much of this work has been elucidated in species other than C. elegans.
A. Generation of ROS
The ETC uses electrons, primarily generated from the citric acid cycle in the mitochondrial matrix, to generate ATP through a series of electron transfers that are coupled to the translocation of protons across the mitochondrial inner membrane (Fig. 2). Electrons from nicotinamide adenine dinucleotide (NADH) are removed by complex I (NADH dehydrogenase/NADH:ubiquinone oxidoreductase) and transferred to the lipid-soluble electron carrier ubiquinone (coenzyme Q). Electrons are transferred to ubiquinone one at a time, yielding first a semiquinone and then ubiquinol. During the electron transfer, protons are transferred from the matrix to the intermembrane space contributing to the electrochemical gradient that exists across the inner mitochondrial membrane.
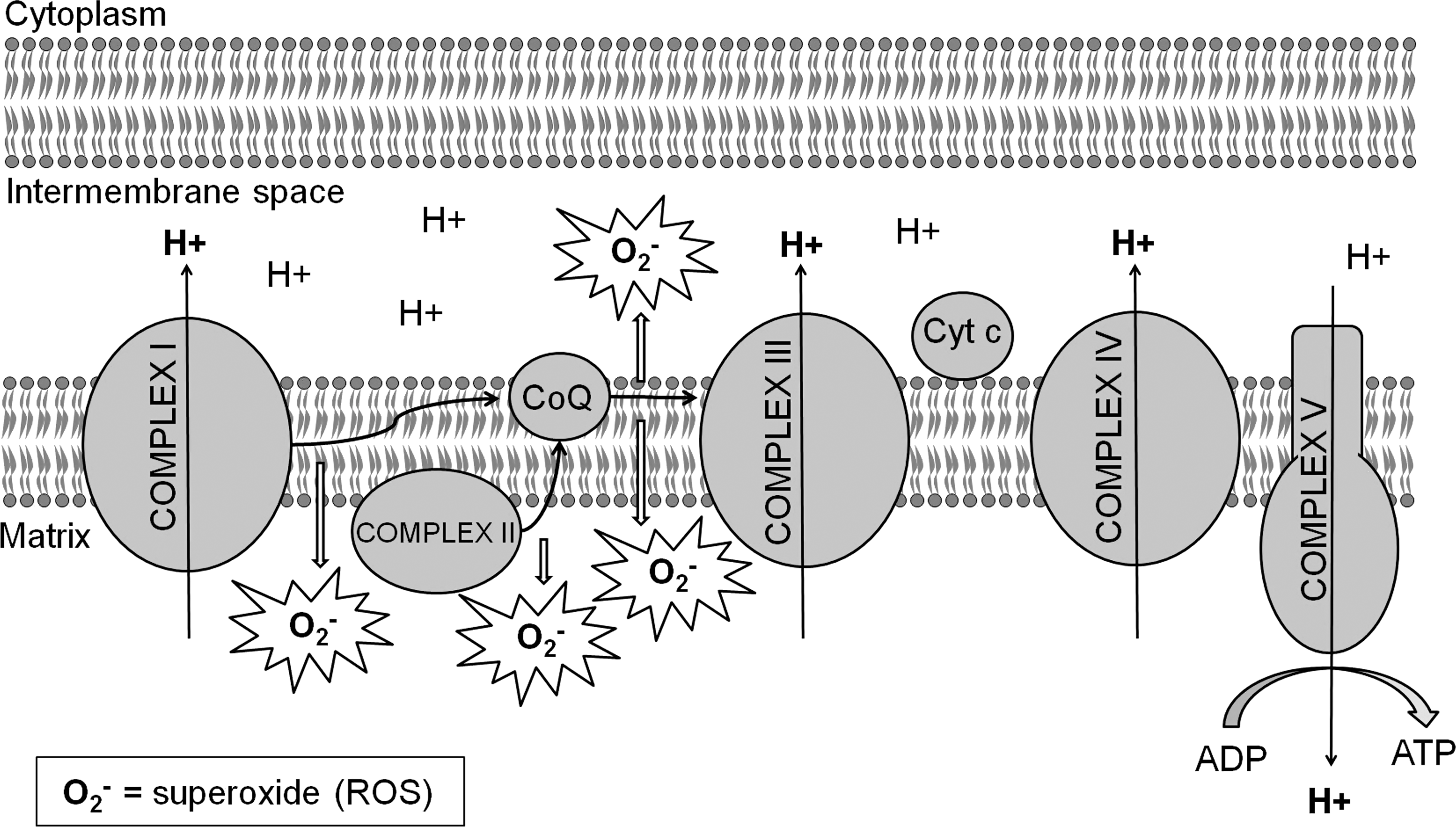
Electrons can also be transferred to ubiquinone from complex II (succinate dehydrogenase), which is located on the inner mitochondrial membrane and receives electrons from succinate in the citric acid cycle. In addition, electrons derived from fatty acid oxidation can be transferred to ubiquinone via the electron transfer flavoprotein (ETF), ETF-oxidoreductase, and flavin adenine dinucleotide. Complex III (cytochrome bc1 ) passes electrons from ubiquinone to cytochrome c transferring protons from the matrix to the inner membrane space in the process. Finally, complex IV (cytochrome c oxidase) transfers electrons from cytochrome c to oxygen to generate water, also moving protons to the intermembrane space. The electrochemical proton gradient produced by electron transfer at complexes I, III, and IV is used by ATP synthase (complex V) to convert ADP into ATP energy.
During the process of electron transport, some electrons are leaked directly to oxygen to generate the ROS superoxide (O2 −). This is thought to occur primarily as electrons are transferred from complex I to ubiquinone and from ubiquinone to complex III (Fig. 2). However, ROS is also thought to be generated inside of complex I (16, 28, 107, 262), at complex II (242), and by ETF and ETF-Q (253). Superoxide from complex I is released into the matrix, whereas that generated from complex III is primarily released into the intermembrane space with some being released into the matrix (42, 225, 253). The rate of superoxide production in the ETC is dependent on oxygen concentration and the degree to which the complexes of the ETC are reduced. Increases in either oxygen concentration or the degree to which the ETC complexes are reduced result in increased generation of superoxide (262). As a result, active mitochondria produce less ROS than resting mitochondria, in which ETC complexes are highly reduced. In addition, mitochondrial ROS generation has been found to be increased in the presence of ETC complex inhibitors (185, 253, 287), leading to the idea that damage to components of the ETC will lead to increased ROS production. However, decreased ROS production is observed in the presence of ETC complex inhibitors under certain conditions [e.g., when succinate is used as a substrate (256)], and this suggests that damage to components of the ETC has the potential to increase or decrease ROS generation.
Superoxide generated from the ETC is thought to remain within the compartment in which it is generated. This conclusion was based on the observation that superoxide is unable to cross membranes (93). While recent work has identified chloride channels that are capable of transporting superoxide across the cell or endosome membrane (69, 101, 208), it is uncertain whether channels exist in the mitochondria that are capable of transporting superoxide, and functional experiments support the conclusion that the bulk of mitochondrial and cytoplasmic superoxide tends to be confined to the compartment in which it is generated (199). In addition, superoxide is rapidly converted to hydrogen peroxide (H2O2) by the enzyme superoxide dismutase (SOD) (superoxide can also undergo spontaneous dismutation albeit at a much slower rate) (Fig. 3).
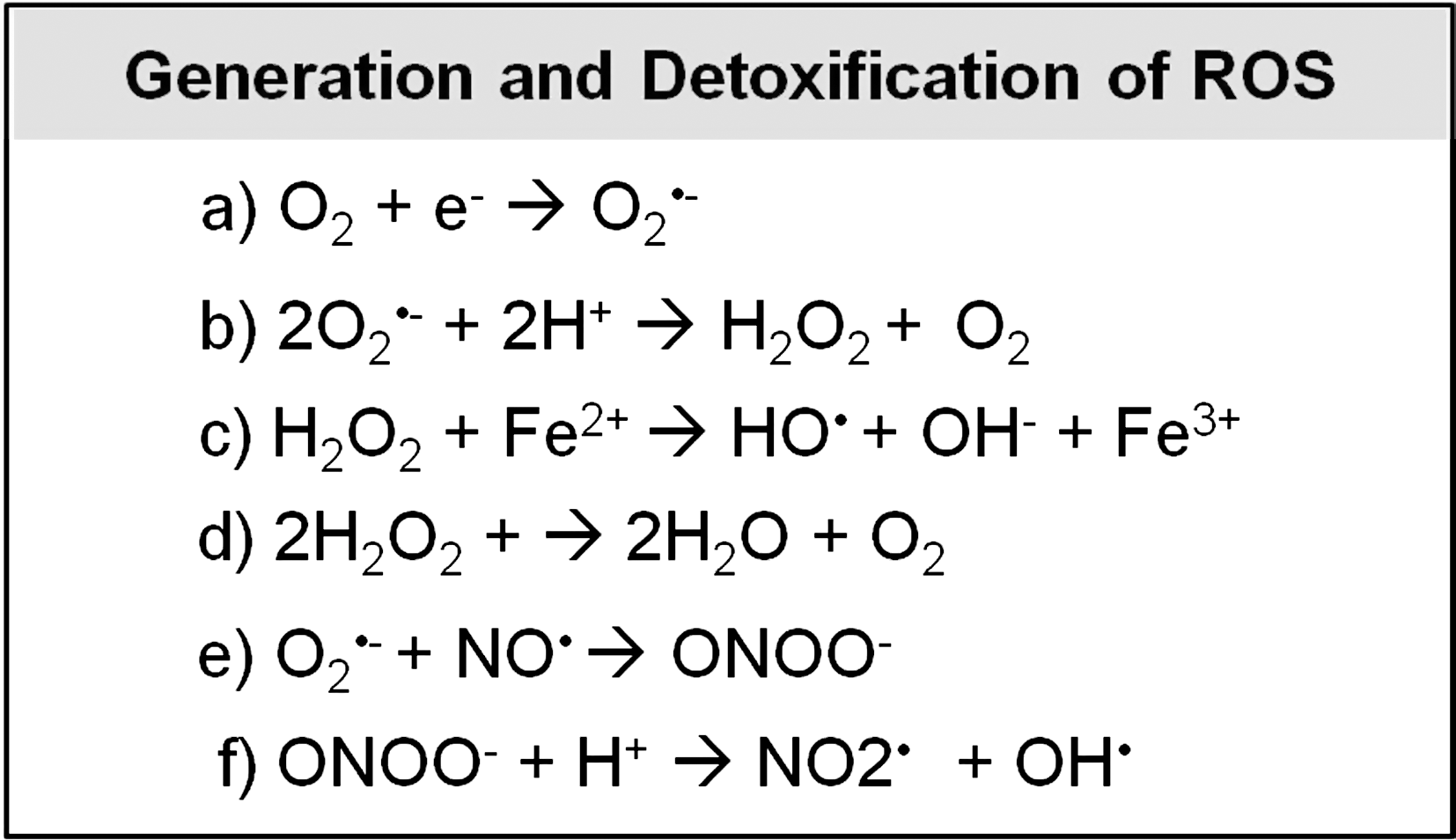
Unlike superoxide, H2O2 is able to travel easily between different compartments of the mitochondria and cytoplasm. While it has generally been assumed that this occurs by simple diffusion through the membrane, it appears that at least some transport of H2O2 across membranes occurs through aquaporin channels (20, 21, 105). H2O2 can be partially reduced to form two other forms of ROS: hydroxyl radicals (OH•) and hydroxide ions (OH−) (Fig. 3). Alternatively, H2O2 can be fully reduced to water (Fig. 3). Superoxide can also react with nitric oxide (NO•) to generate the reactive nitrogen species (RNS) peroxynitrite (ONOO−), which can subsequently be decomposed into two other reactive radicals: nitrogen dioxide (NO2•) and hydroxyl radicals (OH•) (Fig. 3).
In addition to ROS generated by electron transport, a number of other enzymes within the cell are able to generate ROS, either inadvertently or for a particular purpose. P450 oxidases catalyze the oxidation of various molecules within the cell (RH + O2 + 2H+ + 2e− → ROH + H2O) but can generate superoxide as a side reaction (195). Xanthine oxidase catalyzes the conversion of hypoxanthine to xanthine and xanthine to uric acid, producing H2O2 during both conversions (hypoxanthine + H2O + O2 → xanthine + H2O2; xanthine + H2O + O2 → uric acid + H2O2). Similarly, monoamine oxidases in the outer mitochondrial membrane catalyze the oxidation of monoamines and generate H2O2 in the process (RH2NH2 + H2O + O2 → ROH + NH3 + H2O2). Finally, the function of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase is to produce superoxide from oxygen using electrons from NADPH (NADPH + 2O2 → NADP+ + 2O2 − + H+). The existence of enzymes whose function is to generate ROS indicates that in addition to causing oxidative damage, at least some of the ROS within a cell are intentionally generated (e.g., to fight pathogenic bacterial infections; see below).
B. Detoxification of ROS
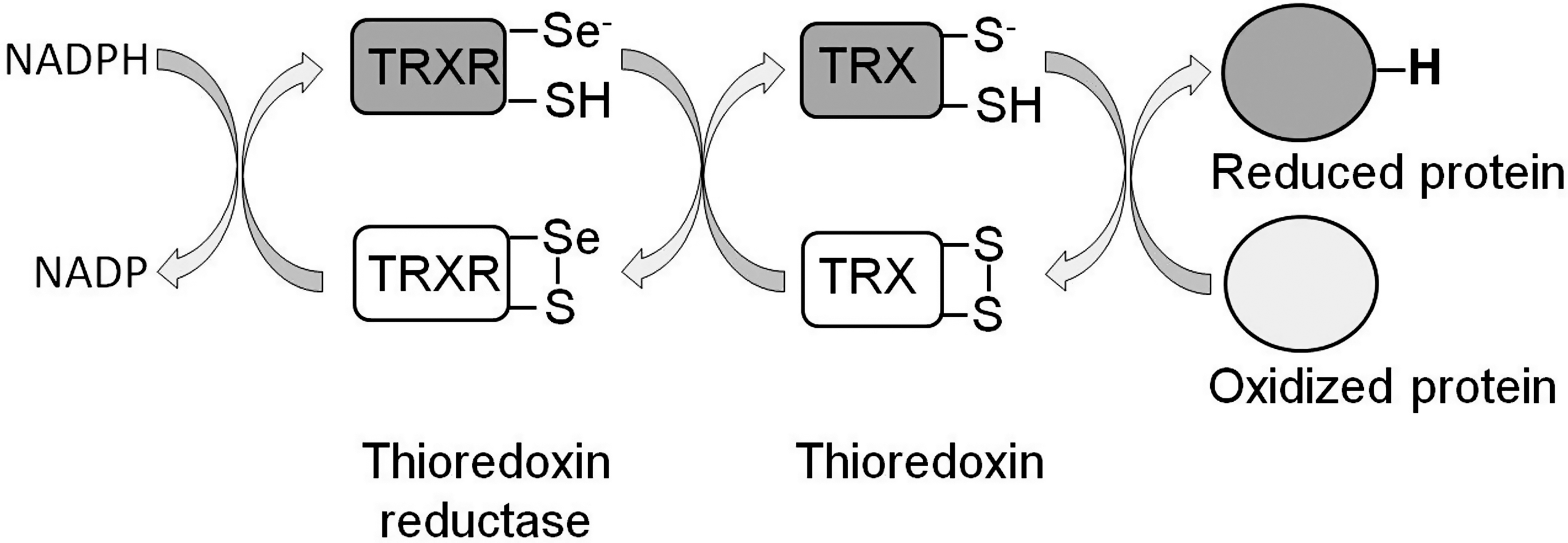
To limit the cellular damage caused by ROS, cells have a number of antioxidant defenses. The first line of defense is SOD, an enzyme that converts superoxide to H2O2. This enzyme is present in the mitochondria, cytoplasm, and extra-cellular space since superoxide detoxification is required at the site of generation. H2O2 can be converted to water by either a catalase or peroxidase (e.g., glutathione peroxidase or peroxiredoxin). In the case of peroxiredoxin, the reduction of H2O2 to water results in the inactivation of the enzyme [see ref. (275) for review]. However, its peroxidase activity can be subsequently restored through reduction by either thioredoxin (TRX) or glutaredoxin (Fig. 4), which are active components of the TRX and glutathione systems, respectively.
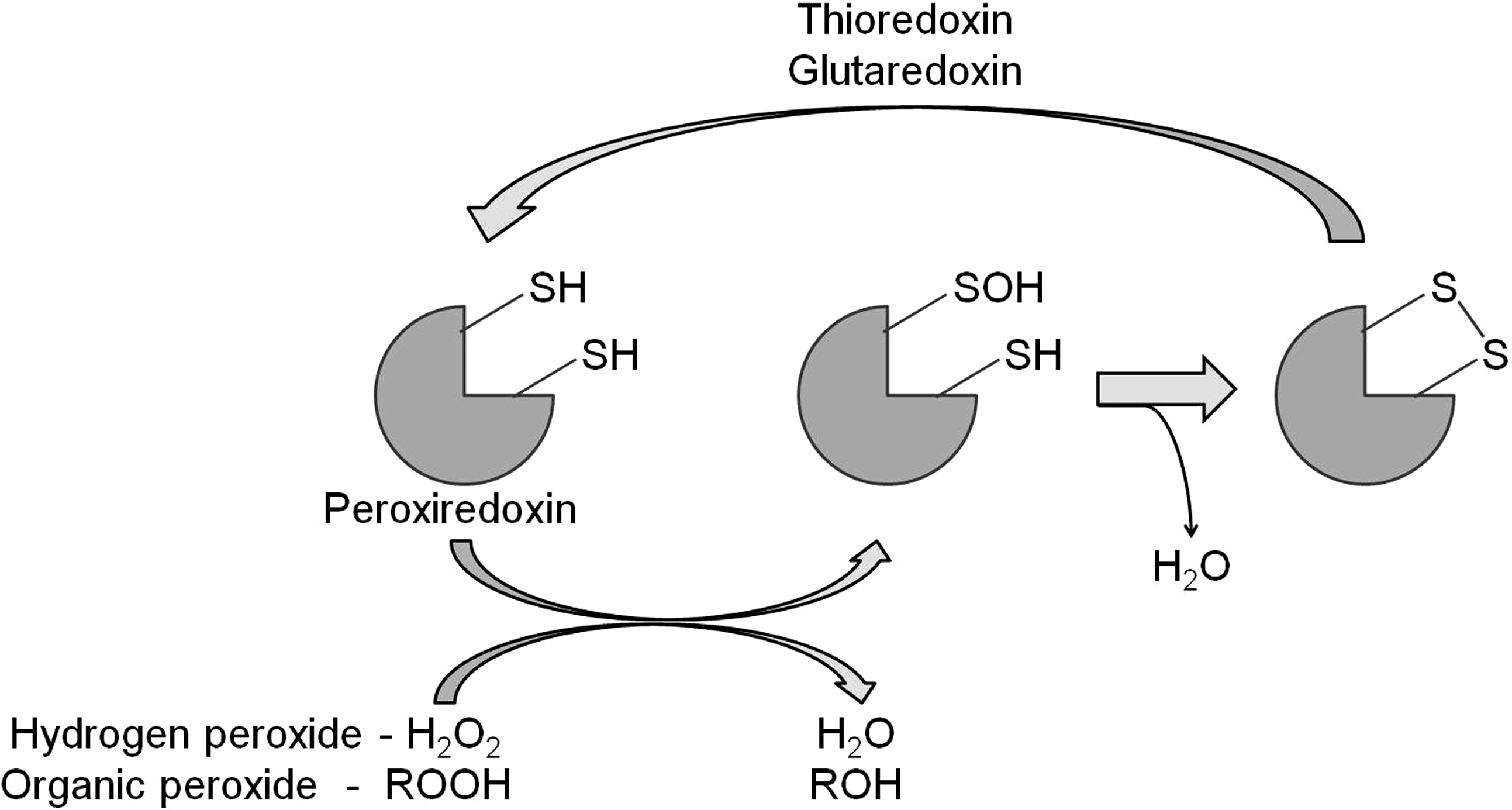
Both the TRX and glutathione systems act by providing reducing equivalents to both ROS and oxidized proteins. Reduced TRX (active) can subsequently be regenerated by TRX reductase (TRXR) using NADPH as a donor of reducing equivalents (Fig. 5) [see ref. (178) for review]. Similarly, glutaredoxin activity is restored through its reduction by glutathione (a readily oxidized tripeptide; see below), which in turn is reduced by glutathione reductase and NADPH (Fig. 6) [see ref. (67) for review]. Finally, glutathione S-transferases act through conjugating glutathione to a variety of toxic substrates, including oxidatively damaged lipids and proteins, thereby decreasing their reactivity and making them more water soluble for removal [see refs. (243, 244) for review] (Fig. 6).
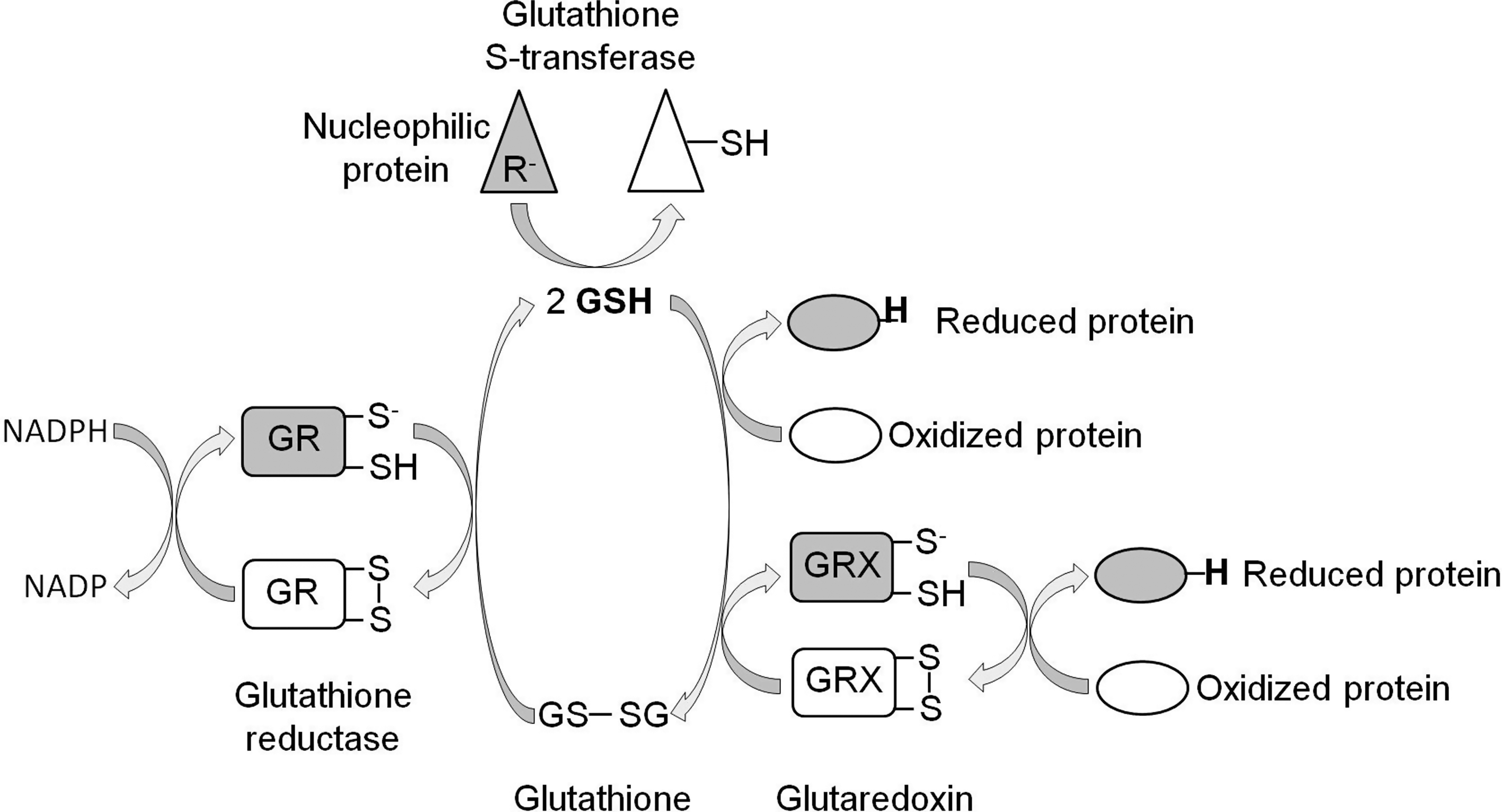
In addition to enzymatic antioxidant defense mechanisms, there are also numerous nonenzymatic defenses. Many of these defenses are simply molecules that are more readily oxidized than nearby essential functional components of the cell. One of the most abundant and important of these is glutathione, a tripeptide composed of glutamate, cysteine, and glycine [see ref. (187) for review]. In its reduced form glutathione can directly reduce substrates, indirectly reduce substrates through glutaredoxin, or act in detoxification and repair in combination with glutathione S-transferase (Fig. 6). Vitamin E is a lipid-soluble nonenzymatic antioxidant that serves to protect membranes from damage. Vitamin C is a water-soluble antioxidant that is important in protecting against lipid peroxidation and protein oxidation. In addition to these three antioxidants, there are also numerous other molecules in the cell that can act as antioxidants, including vitamin A, ubiquinol, and cytochrome c.
C. Molecular damage caused by ROS
Despite the wide array of enzymatic and nonenzymatic antioxidants, at least some of the ROS produced in cells escapes the antioxidant defenses to cause oxidative damage. ROS is able to damage all of the major building blocks of the cell, including DNA, lipids, and protein (Fig. 7). The most commonly quantified form of oxidative damage to DNA is 8-hydroxy-2′-deoxyguanosine, which is formed by the addition of a hydroxyl group to the guanine base of the guanosine nucleotide. Lipid peroxidation is caused by free radical attack on either free lipids or those contained in membranes. Initially, ROS, such as the hydroxyl radical, removes hydrogen from a lipid to generate a lipid radical (R•). The lipid radical can then react with oxygen to form a lipid peroxyl radical (ROO•). This reactive radical can then remove a hydrogen from another lipid to form a lipid peroxide (ROOH). In some cases, such as 4-hydroxynonenal (4-HNE), the lipid peroxidation product can subsequently react with DNA or proteins.
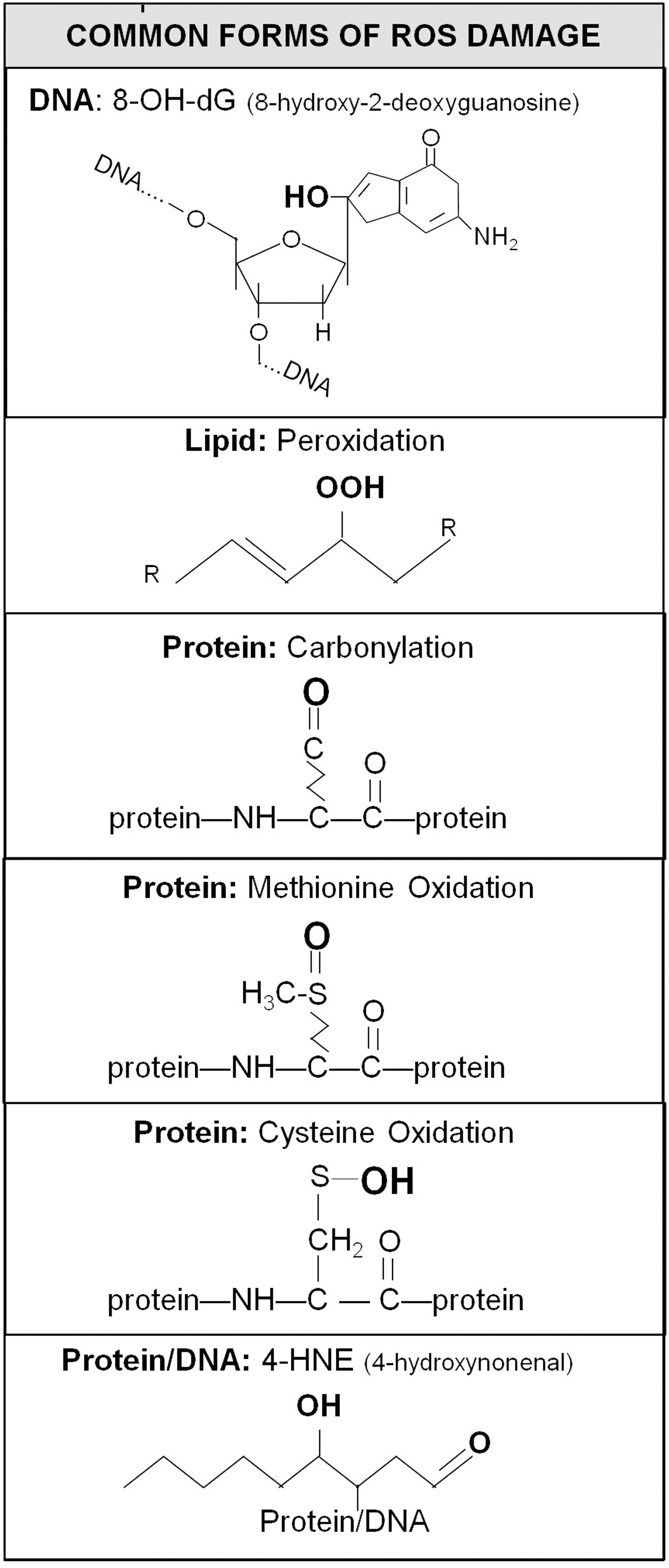
Oxidative damage to proteins can result in carbonylation—the addition of a double-bonded oxygen atom. Protein carbonylation affects specific amino acid side chains, including arginine, lysine, proline, and threonine. In addition, methionine residues can be oxidized to form methionine sulfoxide, which can be further oxidized to methionine sulfone. Similarly, cysteine residues can undergo multiple oxidations to form cysteine sulfenic acid, cysteine sulfinic acid, and cysteine sulfonic acid. Alternatively, cysteine sulfenic acid can form disulfide bonds with other cysteine residues.
III. Caenorhabditis elegans
A. C. elegans as a model organism
C. elegans is a soil-dwelling roundworm (nematode) of about 1 mm in length that feeds primarily on bacteria. As adult worms are just visible to the naked eye, maintaining cultures of C. elegans and experimental manipulations require the use of dissecting microscopes. The idea to use C. elegans as a genetic model organism for the study of developmental biology, particularly the development and function of the nervous system, was first proposed by Sydney Brenner (30). As a genetic model organism, C. elegans has many advantages (Fig. 8). The small size of the worm allows for easy and inexpensive storage and culturing on Petri dishes seeded with bacteria as a food source. The vast majority of C. elegans are hermaphrodites (109), which greatly facilitates genetic manipulations. However, genetic crosses are also possible as a small number of males are present in each population (as a result of nondisjunction to yield XO individuals).
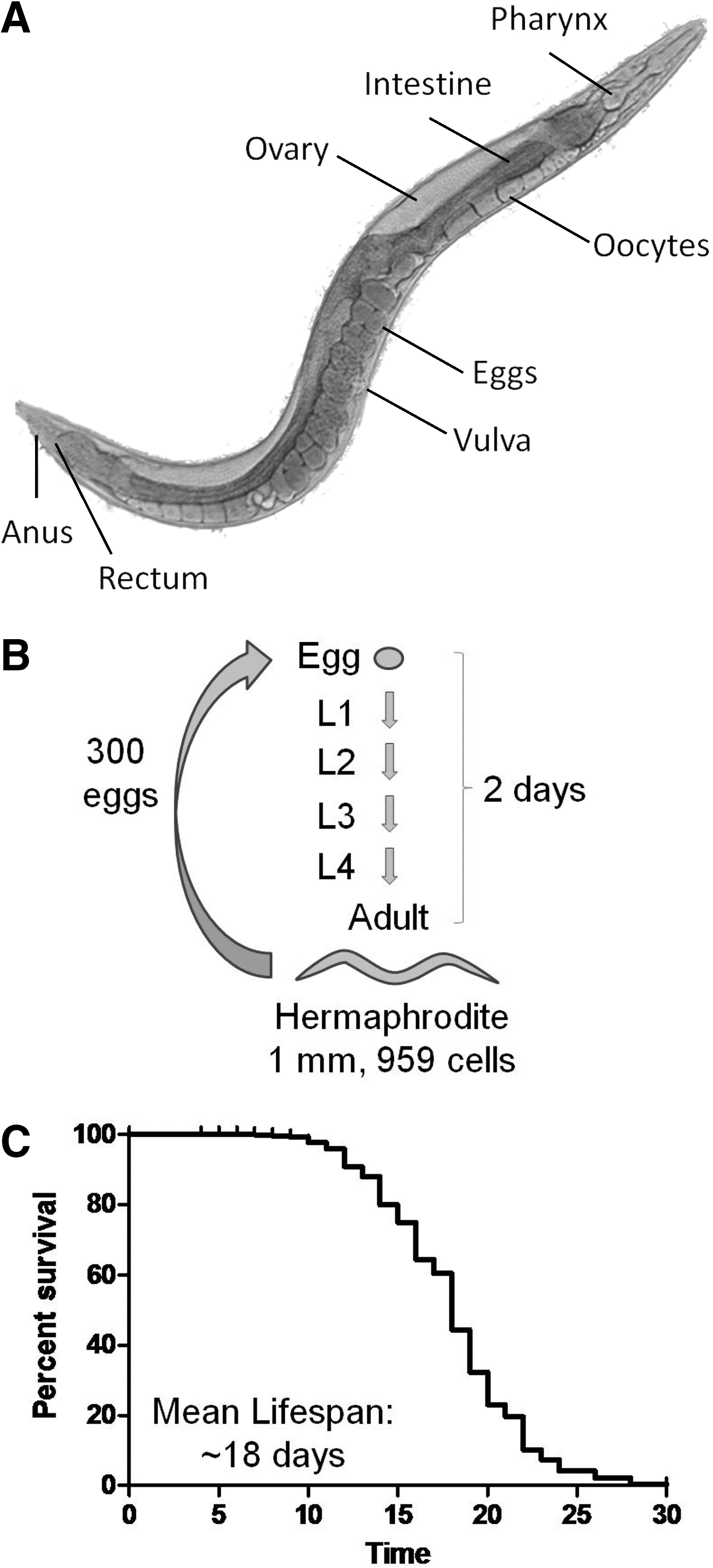
Each worm is able to produce in the order of 300 offspring and this provides more than ample numbers of subjects to power each experiment. In addition, these worms develop from egg to adult in 2 days and complete the egg-to-egg cycle in ∼3 days, thus making it possible to rapidly grow up large populations of worms. C. elegans hermaphrodites have exactly 959 cells (excluding germ cells) of which 302 cells are neurons. Aside from the germ line, all of the cells in the worm are postmitotic. This has allowed researchers to completely map the lineage of each cell in C. elegans, as well as all of the interconnections of the nervous system. Another advantage of C. elegans is the fact that these worms are transparent. This has enabled researchers to determine with particular ease in vivo where and when a particular gene is expressed using fluorescently labeled reporter constructs.
The entire C. elegans genome has been sequenced and was found to contain roughly 20,000 genes [C. elegans sequencing consortium (1)]. A number of resources are available for C. elegans research. The C. elegans Genetics Center (
One of the main advantages of C. elegans is the ability to perform large-scale genetic screens (142). In the classical approach, worms are mutagenized and screened for a particular phenotype, such as increased lifespan (57). While genetic mapping has traditionally been used to identify the gene responsible for the phenotype in question, recent improvements in the speed and cost of whole genome sequencing now provide the opportunity to identify novel mutants by sequencing (108, 236).
An alternative screening approach, which has gained much popularity based on its ease of use and rapid results, is to perform RNA interference (RNAi) screens (23). RNAi is an approach for reducing expression of a particular gene by specifically targeting its mRNA for degradation. Unlike in other organisms, in C. elegans it is possible to decrease expression of a gene simply by feeding the worms bacteria that have been engineered to express a double-stranded RNA (dsRNA) that targets the gene of interest (dsRNA can also be injected or worms can be soaked in a dsRNA solution). This fact greatly facilitates performing RNAi screens in the worm. Accordingly, RNAi bacteria libraries are available containing most of the genes present in the genome [16,757 clones (71)], and many researchers have completed RNAi screens for genes that affect a phenotype of interest [e.g., lifespan (94, 173)]. In addition, the RNAi approach allows control over the extent to which gene expression is decreased. While undiluted RNAi can almost completely eliminate expression of a particular gene, diluting the RNAi bacteria with normal bacteria allows for a milder inhibition of gene expression [e.g., refs. (230, 282)].
RNAi in C. elegans is not without its limitations, and these must be kept in mind when interpreting the results of any RNAi experiment. First, RNAi does not affect all tissues equally. Most notably, certain cell types, such as neurons, exhibit low sensitivity to RNAi (68, 258). In some cases this limitation can be overcome by using strains with increased sensitivity to RNAi such as eri-1 or rrf-3 (150, 249). Further, it is not possible to target mitochondrial genes using RNAi. Second, when using the feeding technique, it has not been possible to simply mix two RNAi strains together to achieve knockdown of two genes. Instead, it is necessary to either inject the two different RNAi constructs or generate a single construct that targets the two separate genes (197), both of which are somewhat labor intensive. Third, it is important to note at which stage the RNAi treatment was begun. Especially in the case of RNAi targeting mRNA encoding mitochondrial proteins, RNAi treatment should begin a generation before experimentally tested animals to avoid maternal rescue, especially if the most severe phenotype possible is what is being sought. Finally, it is important to consider how much a gene's expression is decreased and if there is variability in knockdown between tissues within a worm and between worms in a population. In many cases the degree of knockdown has not been measured in RNAi experiments in C. elegans, and thus the possibility cannot be excluded that a gene is not knocked down sufficiently to observe an effect.
While working with C. elegans has many advantages, there are also disadvantages to working with such a small organism. To perform any molecular analysis, large numbers of worms must be collected, particularly if subcellular organelles such as the mitochondria are to be examined. This can be quite challenging when studying mutant strains of worm that have decreased brood sizes and slow development times. Further, while it is possible to express a transgene or RNAi construct in a particular tissue using tissue-specific promoters or by controlling which tissues are sensitive to RNAi (227), it is difficult or impossible to use molecular techniques to examine the effects of a specific manipulation on a particular tissue. Accordingly, changes are normally examined in the whole worm. In addition, worms lack many of the tissue types that are found in higher organisms (e.g., heart, liver, and kidney). Finally, as with any model organism, it is unclear to what extent conclusions drawn from experiments in worms will hold in other species and in humans. Nonetheless, previous studies with this organism suggest that in most cases similar results are obtained (17).
B. Studying aging in C. elegans
C. elegans has been utilized as a model to study aging because of its relatively short lifespan and the other advantages of this organism mentioned above. At 20°C, the average lifespan of a wild-type worm is just under 3 weeks (Fig. 8C). While stochastic effects lead to a range of lifespans within a genetically identical population in the same environment (106, 158, 231), mean lifespan is subject to low variability and differences between strains are highly reproducible. The validity of studying aging in the worm has been supported by experiments demonstrating that genes or interventions that extend lifespan in the worm also extend lifespan in other organisms and vice versa. For example, dietary restriction, which was first demonstrated to extend lifespan in rats (192) and mice (272), was later shown to increase lifespan in worms (159, 165) and flies (43). Conversely, decreasing expression of clk-1, which affects mitochondrial function (see below), was originally found to increase lifespan in the worm (164, 274) and later shown to result in extended longevity in mice (184). Finally, reduced insulin/insulin-like growth factor 1 (IGF-1) signaling is proposed to be a universal mechanism of lifespan extension (151, 152).
While assessing whether or not a worm is alive or dead is relatively straightforward with a bit of practice, care must be taken in performing lifespan studies as variations in protocol can impact the results of a particular study (92, 219). Standard conditions for lifespan assessment in worms is generally 20°C on nematode growth medium plates seeded with OP50 bacteria. In some cases, lifespan studies have been performed in a liquid medium. Under these conditions, some reports have observed increased lifespan compared to worms grown on a solid medium (48), whereas others have not (120). The discrepancy may result from the use of different concentrations of bacteria, as this has been shown to influence lifespan in liquid culture (159), likely through a mechanism of dietary restriction. Importantly, the degree to which lifespan is increased in long-lived mutants in liquid media has been shown to be highly correlated with lifespan increase on solid media (140). One of the main benefits of liquid culture in small wells is the ability to perform high-throughput screens for genes or compounds that influence lifespan (70, 222).
In addition, lifespan studies are frequently performed at 25°C since worms have been found to exhibit a shorter lifespan at higher temperatures (159). This approach is commonly used when studying long-lived strains, as this will reduce the length of time required to complete the experiment. It is also used in cases where a temperature-sensitive mutation is under study. However, performing studies at this temperature may hide differences that would be apparent at lower temperatures. For example, the long-lived mutant clk-1 (described in detail below) has a clearly increased lifespan at 20°C, but shows little or no increase in lifespan at 25°C (164).
One problem with performing lifespan studies in worms is keeping track of which worms are being studied since each worm lays about 300 eggs, each of which will become an adult in about 2 days. Thus, under standard conditions worms are typically transferred to a new plate every 1–2 days during their egg-laying period, which lasts around 1 week. However, this approach can be very labor intensive. Each transfer has the potential to injure the worms and may have unknown effects on lifespan. Two approaches have been adopted to circumvent this problem: the use of strains with temperature-induced sterility (62) or growing worms on plates containing fluorodeoxyuridine, a compound that prevents the worms from producing offspring by inhibiting DNA replication (78, 200). These approaches, however, may also have unknown effects on lifespan and may interact with the gene or intervention under study (5).
Another consideration is how to treat worms that die prematurely of causes that are likely not related to aging. These deaths include internal hatching of progeny (also known as bagging), external exposure of internal organs (also known as exploding or prolapsed gut/gonad), and worms that crawl onto the side of the Petri dish and become desiccated. It is uncertain whether worms that succumb to these early deaths would have a different lifespan than the population of worms that remain healthy. Two general approaches have been utilized to account for these worms. In one approach, the worms are censored—that is, these worms are included in the dataset until the point in time that they die, but their deaths are not recorded. In the other approach, worms that die early by one of these ways are simply removed from the study and replaced with another worm from a group generated as part of the initial cohort of worms but whose survival was not being monitored. In this way, the sample size is constant across strains and trials. Bagging and exploding normally occur before any worms in that population die of a natural death and thus should not affect the survival curve or statistical comparison by either approach. It is only in the rare circumstance that censoring occurs after the first natural death that the censored worms will affect the survival plot and statistical significance.
Finally, it is important to consider the bacterial food source. If the concentration of bacteria on the plates is low, the worms may be calorically restricted, which will extend their lifespan (159). In addition, high concentrations of bacteria can be toxic to worms resulting from bacterial proliferation in the pharynx and intestine (79). Accordingly, to eliminate the possibility that an altered bacterial density is affecting lifespan, some researchers perform lifespan studies on killed bacteria (normally by ultraviolet [UV] irradiation or heat), which has been shown to increase worm lifespan (79, 84).
In addition to the quantity of food, the quality of the food can also influence worm lifespan. Worms exhibit a clear preference for bacteria that are better able to sustain worm growth (18, 248) and avoid bacteria that are pathogenic (291). Feeding worms bacteria with impaired respiratory metabolism has, in fact, been shown to increase worm lifespan (234). It has also been shown that the metabolism of worms is affected by their food source and this may have an impact on lifespan (232).
While much of the research on aging in C. elegans has used lifespan as a surrogate measure, it is also important to consider how an individual ages. Accordingly, a variety of age-related changes have been measured to assess worm health during aging. As worms age, a number of physiologic changes take place, including decreased pharyngeal pumping, decreased movement, cessation of reproduction, slow defecation rate, accumulation of bacteria in the pharynx and intestine, muscle wasting, and accumulation of aging pigments (45). In fact, many of these phenotypes have been examined for their use as biomarkers of aging (224).
A number of groups have demonstrated that body movement is predictive of lifespan (106, 117, 123, 139). Recently, it has been shown that this phenotypic outcome measure can be quantified automatically with a computer and video camera, thereby allowing an objective, quantitative measure of worm health span (the period that worm remains healthy) (122). Similarly, the rate of accumulation of age pigments has been proposed as a predictor of lifespan. Age pigments include lipofuscin, which is composed of oxidized and cross-linked molecules that cannot be degraded, and advanced glycosylation end products. The levels of age pigments in the intestine increase as worms age (159), are inversely correlated with mobility (85) and are found to accumulate less rapidly in long-lived mutant strains (85).
C. Assessing the role of ROS in determining C. elegans lifespan
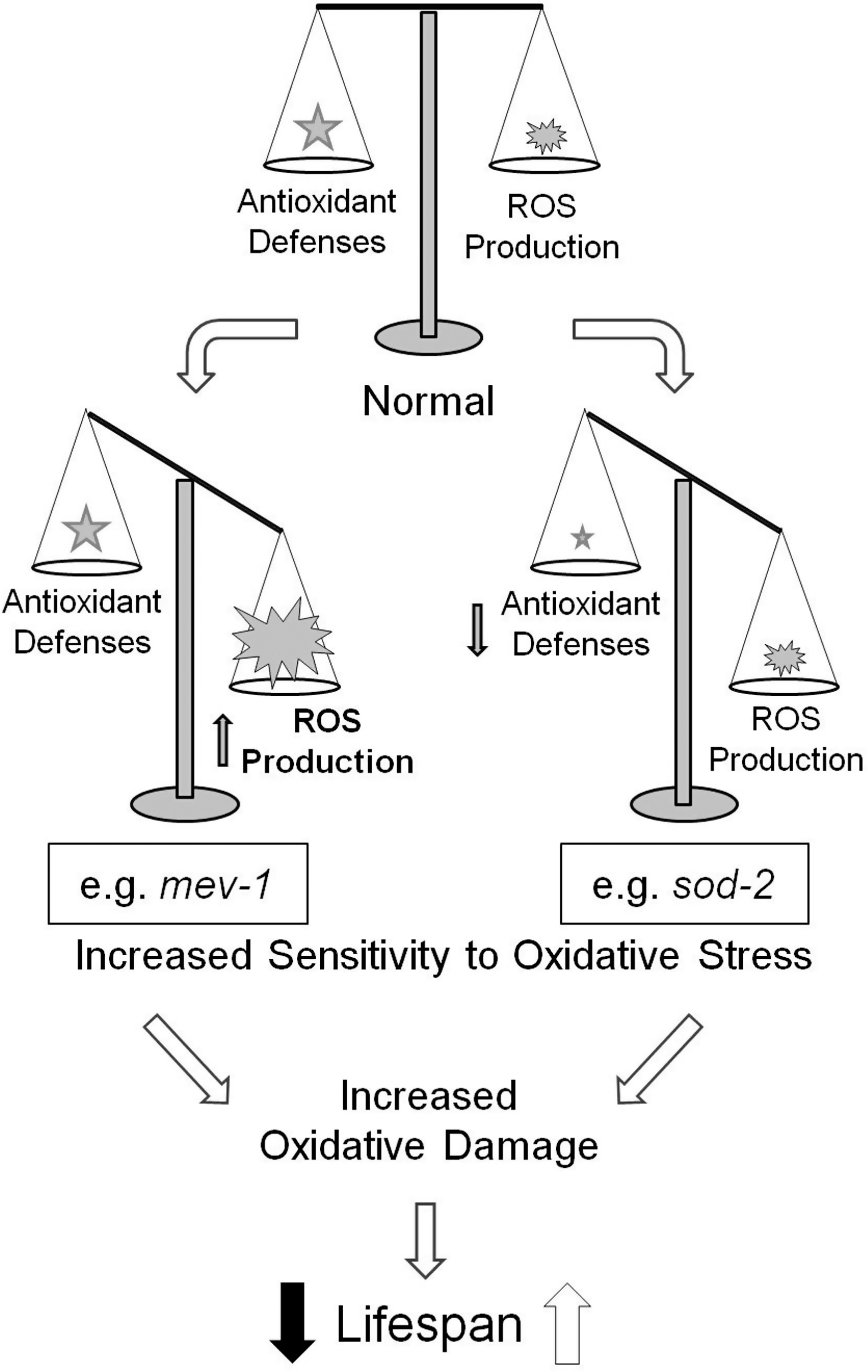
The free radical theory of aging proposes that aging results from the accumulation of oxidative damage over time, which eventually leads to cellular dysfunction and organismal death (96). C. elegans provides an ideal model with which to test the predictions of this theory (Fig. 9). If the free radical theory of aging is correct then increasing oxidative damage should decrease lifespan while decreasing oxidative damage should increase lifespan. To test this theory, researchers have used both genetic and pharmacological approaches to alter the levels of ROS, through affecting either ROS production or ROS detoxification, and examined the effect altered ROS levels on oxidative damage and lifespan.

One of the major limitations of testing the free radical theory of aging is the measurement of ROS and oxidative damage. The measurement of ROS and interpretation of the resulting data is complicated by the short-lived and reactive nature of these molecules, the fact that ROS can be rapidly eliminated by antioxidants and the fact that at least some forms of ROS, such as superoxide, appear to be mostly limited to the compartment in which they are generated.
To measure the levels of ROS in worms, researchers have used a variety of fluorescent or chemiluminescent dyes such as Amplex red, 2,7-dichlorodihydrofluorescein-diacetate (DCF), MitoSOX, MCLA (methyl-cypridine-luciferin analogue), dihydroethidium, and MPEC (2-methyl-p-methoxyphenylethynyl-imidazopyrazinone), either in whole worms or in isolated mitochondria (54, 160, 239, 268, 279, 284). While measuring H2O2 levels in isolated mitochondria is the most common and most reliable approach to measuring ROS, this does not necessarily reflect the levels of ROS in vivo (166, 212). However, measurement of ROS in vivo is technically difficult and less reliable than measurements on isolated mitochondria. In both cases, the dyes used to detect ROS are thought to be nonspecific for a particular form of ROS, and thus it is often uncertain exactly what is being measured (166). In addition, it is also not always clear whether it is ROS production or steady-state ROS levels that is being measured.
Given the technical difficulty in attempting to measure ROS using dyes, many researchers have instead examined sensitivity to oxidative stress. While this may result from either an increase in ROS production or a decrease in antioxidant defenses, increased sensitivity to oxidative stress suggests an increased steady state level of ROS or a decreased ability to respond to ROS, both of which could result in increased oxidative damage (Fig. 10). A number of compounds have been used to assess the sensitivity to oxidative stress with the most common being paraquat (134, 156, 266) and juglone (52, 140, 263). Both of these compounds lead to intracellular generation of superoxide, thereby inducing oxidative stress (22, 55). In addition, direct exposure to H2O2 (57, 169, 214) or high concentrations of oxygen (hyperoxia) (3, 100, 277) are also used to assess sensitivity to oxidative stress (as mentioned above, increasing the concentration of oxygen will lead to increased ROS production). This approach, in addition to providing only an indirect measurement of ROS or oxidative damage, also exposes worms to conditions that would not normally be encountered during normal life. Thus, it is possible that a worm with increased ROS production is more sensitive to paraquat, but that under normal conditions the worms antioxidant defenses are more than sufficient to detoxify all of the ROS produced. Also, while all of these methods for inducing oxidative stress are thought to act through ROS, in some instances worms have been found to be sensitive to one form of oxidative stress but not sensitive or resistant to another (173, 270, 277).
Testing sensitivity to oxidative stress has typically been done either during development or in young adult worms. Assays performed during development examine either the percentage of worms that are able to develop to adulthood under stress conditions or the furthest developmental stage obtained. However, the interpretation of results from this approach becomes complicated if the development time differs between the strains being compared (66). In this case, a slower developing strain will be exposed to the oxidative stress for a longer period, and thus its failure to develop could result from either increased sensitivity to the oxidative stress or the increased exposure to the stress. In addition, assays performed during development and adulthood may yield different results for the above-mentioned reason or it is possible that high ROS production during development results in increased sensitivity to oxidative stress, which induces the upregulation of antioxidant defense genes and leads to decreased sensitivity to oxidative stress during adulthood. Accordingly, it would be more informative to examine sensitivity to oxidative stress at multiple time points.
Altered sensitivity to artificially induced oxidative stress (e.g., through exposure to paraquat) may result from factors that are independent of ROS production and antioxidant defenses. For example, increased resistance to paraquat could potentially result from increased thickness of the cuticle, decreased ability to take up paraquat, or increased ability to metabolize paraquat. In these cases, altered sensitivity to oxidative would not necessarily be expected to influence lifespan and could lead to spurious conclusions with regard to the contribution of oxidative stress to aging.
Finally, it is also possible to examine the net effect of ROS and ROS scavenging ability by quantifying oxidative damage. Measuring oxidative damage provides direct data for testing the free radical theory of aging. To measure oxidative damage, most researchers have focused on damage to proteins either by measuring protein carbonylation by derivatization with dinitrophenylhydrazine and detection with antibodies (3, 223, 281) or by measuring the levels of 4-HNE-modified proteins using antibodies (146, 284). Oxidative damage to DNA has been assessed by measuring the levels of 8-hydroxy-2′-deoxyguanosine (99), whereas few reports have examined lipid peroxidation (213). A summary of experimental techniques used to assess ROS, antioxidant defense, and lifespan is provided in Table 1. In the sections that follow, this review will examine the support for the free radical theory of aging from experiments in C. elegans using environmental, genetic, or pharmacologic interventions.
H2O2, hydrogen peroxide; ROS, reactive oxygen species.
D. Hormesis
In examining the relationship between oxidative stress and lifespan, it is important to consider the potential influence of hormesis. Hormesis may be defined as a beneficial response to a sub-threshold dose of a stimulus that at a higher dose is toxic or detrimental. Using a variety of stress conditions, Cypser and Johnson examined whether hormesis could lead to extended longevity in the worm (49). Without any pretreatment, exposing worms to 100% oxygen for 24 h resulted in 70%–90% death. However, by pretreating worms with hyperoxic conditions (100% oxygen, 8 h), it was found that not only were the worms better able to survive a subsequent semilethal exposure to oxygen (death reduced to 10%–50%), but they were also more resistant to juglone-induced oxidative stress and showed increased lifespan (49).
Similarly, Yanase et al. demonstrated that the lifespan of the long-lived age-1 worms (discussed below) could be increased by treatment with 90% oxygen for 3 h per day (281). Investigation into potential mechanisms leading to the extension of lifespan resulting from hyperoxia revealed that hyperoxia-treated age-1 worms showed increased levels of sod-1, sod-2, sod-3, and sod-5 mRNA (279, 281). This increase in antioxidant defense resulted in a decrease in mitochondrial ROS levels and decreased protein carbonylation (279). Interestingly, the lifespan of wild-type worms was not increased by intermittent exposure to hyperoxia, suggesting the possibility that age-1's ability to increase antioxidant defenses in response to oxidative stress may contribute to their longevity.
As with hyperoxia, exposing worms to chemicals that induce oxidative stress can be either lethal or extend lifespan depending on the dose. While high concentrations of the superoxide generating compound juglone (e.g., 250 μM) results in early death, exposure of worms to lower levels of juglone (40 μM) results in slightly extended longevity (102). Interestingly, exposing worms to low concentrations of juglone does not extend lifespan in daf-16 worms. As daf-16 encodes a transcription factor that mediates a stress response through translocation to the nucleus and activation of transcriptional targets, this result suggests that the mechanism by which the lower level of juglone increases lifespan is acting through a transcriptional response mediated by daf-16. In support of this idea is the fact that exposure to low concentrations of juglone results in nuclear localization of daf-16 and increased expression of the daf-16 transcriptional target sod-3 (102). Worms exposed to low concentrations of juglone exhibit increased expression of hsp-16.2, increased levels of glutathione, increased SOD activity, and increased catalase activity, all of which may contribute to the lifespan extension through an increase in resistance to subsequent oxidative stress (102).
The ability of increased ROS to contribute to extended longevity was also studied by Schulz et al., who examined the mechanism by which decreased glucose metabolism could increase lifespan (239). They showed that inhibition of glucose metabolism through treatment with 2-deoxy-D-glucose (DOG) resulted in decreased ATP production, which lead to a compensatory increase in mitochondrial respiration. This increase in the level of mitochondrial respiration resulted in increased production of ROS (as measured by DCF), which induced a stress response exhibited by an increase in catalase activity (though SOD and GPX activity were unchanged) (239). As a result, DOG-treated worms are resistant to paraquat-induced oxidative stress and exhibit extended lifespan. Finally, to demonstrate that increased ROS is essential in mediating the extended longevity of DOG-treated worms, Schulz et al. showed that exposing DOG-treated worms to antioxidants (N-acetyl cysteine, ascorbic acid, or trolox) eliminates the observed increase in lifespan (239).
These examples clearly demonstrate that it is important to consider the levels of ROS and whether the effect on lifespan is direct or a result of compensatory mechanisms. While acute exposure to high levels of ROS is directly toxic, exposure to low levels of ROS might increase lifespan indirectly through upregulation of stress response genes.
IV. Long-Lived Mutants Show Increased Resistance to Oxidative Stress
One way in which researchers have investigated the relationship between ROS and lifespan is to examine resistance to oxidative stress of long-lived strains. Generally, it has been observed that mutations that increase lifespan also result in increased resistance to at least some forms of stress (141). While resistance to other forms of stress, such as heat stress (182, 183), UV stress (209), and heavy metals (15), has been observed in long-lived strains and may contribute to their longevity, this section will focus on resistance to oxidative stress.
A. age-1–phosphoinositol 3-kinase
age-1 (PIK3CA) was the first long-lived strain to be studied in C. elegans (138) and it was found later to encode phosphoinositol 3-kinase, a protein involved in insulin/IGF-1 signaling (205). A number of mutations in the insulin/IGF-1 signaling pathway, which impacts growth, metabolism, and dauer formation (see below) in the worm, have been shown to extend lifespan. While age-1 worms were originally found to live 1.5–2 times longer than wild-type worms (73), some mutations in the age-1 gene have now been shown to increase lifespan almost 10-fold (10).
Examination of resistance to oxidative stress revealed that age-1 worms exhibited increased resistance to paraquat (266), H2O2 (169), hyperoxia (3), and juglone (140). Consistent with their increased resistance to oxidative stress, age-1 worms have been found to exhibit increased SOD and catalase activity (169, 266, 267). In fact, Larsen et al. showed that the resistance of age-1 worms to H2O2 only occurred as the worms aged and corresponded to an age-dependent increase in SOD and catalase activity (169). In line with these findings, Adachi et al. found that age-1 worms have decreased protein carbonylation and that the levels of protein carbonylation did not increase with age until very late in life, in contrast to the clear increases observed in the wild-type strain (3). Unlike wild-type worms, age-1 worms did not show increased levels of protein carbonylation under hyperoxic conditions (3). age-1 worms also exhibit a hyperoxia-induced hormesis that is not observed in wild-type worms (281). age-1 worms that are preconditioned with 3 h exposures to 90% oxygen daily from day 5 to 15 exhibited increased expression of SOD and catalase mRNA, decreased levels of ROS, decreased levels of protein carbonylation, increased resistance to paraquat, and a 15% increase in lifespan (279, 281).
While these results suggest that the extended longevity of age-1 worms may result from increased resistance to oxidative stress, the association does not indicate causality. In addition to increased resistance to oxidative stress, age-1 worms are also resistant to other stresses, such as heat stress (183), UV stress (209), and heavy metals (15), which may contribute to their longevity. Thus, it is possible that multiple forms of stress and damage contribute to aging, of which oxidative damage is only one. Further, in examining the lifespan and sensitivity to H2O2 of three different age-1 alleles, it was found that the allele conferring the longest lifespan did not increase resistance to H2O2, thereby indicating that the effects of age-1 on oxidative stress resistance could be dissociated from its effects on lifespan (57). In addition, age-1 worms actually have increased levels of superoxide compared with wild-type worms (279) as well as an increased metabolic rate potential (rate at which superoxide is produced in a lucigenin assay) (267). While the increased longevity of age-1 worms is associated with increased resistance to oxidative stress and decreased oxidative damage, it is uncertain to what extent oxidative stress resistance contributes to their long life.
B. daf-2–insulin/IGF-1 receptor
daf-2 (IR) worms have impaired insulin/IGF-1 signaling resulting from a mutation in the insulin/IGF-1 receptor (157) and live at least twice as long as wild-type worms (153). In assays of resistance to oxidative stress, daf-2 worms have been shown to be resistant to the superoxide generator menadione (112), paraquat (114, 118), and hyperoxia (114). In line with this finding, daf-2 worms have increased mRNA expression of sod genes (sod-1, sod-3, and sod-5) and catalase (ctl-3), as well as 16 glutathione S-transferases (56, 112, 190, 191, 211, 217, 281). Antioxidant activity measurements show that SOD and catalase activity are also increased (118, 267). The increased antioxidant defense of daf-2 worms is also indicated by a more gradual decline in glutathione levels compared with wild-type worms (32) and decreased levels of protein carbonylation (283, 285).
Attempts to measure ROS levels in daf-2 worms have yielded varied results depending on the technique employed. Measurement of the rate of superoxide production using lucigenin [the authors refer to this as metabolic rate potential (267)] and measurement of H2O2 levels from isolated mitochondria (32) indicated increased levels of ROS in daf-2 worms. In contrast, examining MitoSOX staining in the pharyngeal bulb (54) or whole-worm DCF staining (8) suggested that ROS levels are not increased in daf-2 worms. In addition, Complex I-dependent oxygen consumption was significantly increased in daf-2 worms (63), which would suggest increased ROS production if the percentage of electrons leaked to form ROS is similar to wild-type worms.
The long life of daf-2 worms is dependent on daf-16, which encodes a transcription factor in the insulin/IGF-1 signaling pathway (179). Among the numerous targets of daf-16-mediated transcription are several antioxidant defense genes, including sod-3, ctl-1, ctl-2, and gst-4 (177, 189, 211). Accordingly, daf-16 mutants have decreased SOD expression and activity, decreased catalase expression and activity, increased sensitivity to paraquat, increased levels of protein carbonylation, and decreased lifespan (118, 281).
To test whether increased antioxidant defenses contributed to the extended lifespan of daf-2 mutants, researchers have decreased expression of antioxidant defense genes and examined its impact on daf-2 lifespan. Decreasing expression of either sod-3, ctl-1, or ctl-2 by RNAi all resulted in some shortening of the lifespan of daf-2 worms (211), suggesting that each of these genes partially contributes to the long life of daf-2 mutants. However, the contribution of increased antioxidant defenses to daf-2 lifespan is not supported by experiments using genetic mutants. Two groups have demonstrated that deletion of individual sod genes (including sod-3) did not decrease the lifespan of daf-2 worms (56, 114). In fact, deletion of sod-3, or sod-2 and sod-3 in combination was found to significantly increase the lifespan of daf-2 worms (114). Similarly, RNAi targeted against sod-1 was also shown to increase the lifespan of daf-2 worms (283). Deletion of sod-2 and sod-3 together eliminated daf-2 worms' resistance to oxidative stress but increased their lifespan; this suggests that resistance to oxidative stress is not necessary for the long life of daf-2 worms (114). Further support for this conclusion comes from the observation that a mutation in mev-1, which encodes succinate dehydrogenase subunit c (discussed below), eliminates daf-2 mutants' resistance to oxidative stress but does not affect their lifespan (114). Thus, while daf-2 worms have increased lifespan and increased resistance to oxidative stress, their resistance to oxidative stress can be experimentally dissociated from their increased longevity.
C. eat-2–dietary restriction
eat-2 (CHRNA7) encodes a subunit of a ligand-gated ion channel located postsynaptically in pharyngeal muscle, and mutations in this gene result in reduced food intake caused by decreased pharyngeal pumping (9, 193, 229). The decrease in food intake in eat-2 worms results in dietary restriction, which has been shown to increase the lifespan of eat-2 worms (165). As with daf-2 worms, eat-2 worms have increased sod mRNA expression (sod-1, sod-2, sod-4, and sod-5 are increased), increased SOD activity, and increased catalase activity (121, 217). These worms also have increased resistance to juglone-induced oxidative stress (Van Raamsdonk and Hekimi, unpublished observations). However, deletion of individual sod genes does not reduce the increased lifespan resulting from dietary restriction induced by growth in axenic media (286). This suggests the possibility that the long life of eat-2 worms may not be dependent on their enhanced antioxidant defense.
D. Dauer larvae
Under unfavorable environmental conditions, C. elegans can arrest development before adulthood to form dauer larvae, which are designed to survive until conditions are more suitable for growth and reproduction (37). Accordingly, dauer larvae do not feed, have decreased metabolism, and can survive for multiple months (35). The increased longevity of dauer larvae is accompanied by increased resistance to a variety of stresses, including oxidative stress. Resistance to oxidative stress likely results from the observed increases in SOD and catalase levels and activity (56, 119, 268). In addition, the decreased metabolism of dauer larvae is predicted to result in decreased production of ROS, although this has not been measured. While increased resistance to oxidative stress is associated with increased lifespan in dauer larvae, it is uncertain to what extent dauer larvae's resistance to oxidative stress contributes to their survival, since these larvae are resistant to a wide variety of stresses.
E. Summary
These three long-lived strains all demonstrate increased resistance to oxidative stress and upregulation of antioxidant defenses; however, in each case there is evidence suggesting that the resistance to oxidative stress can be dissociated from the increase in lifespan (Table 2). As these long-lived strains and dauer larvae are also resistant to other stresses, such as heat stress, it is possible that increased resistance to a variety of stresses contribute to the lifespan of these long-lived mutants and that the individual contribution of oxidative stress resistance has only a minor impact on lifespan.
IGF-1, insulin-like growth factor 1.
V. Screening for Mutants with Altered Sensitivity to Oxidative Stress
To examine the relationship between oxidative stress and lifespan, some researchers have performed screens for strains of worms that have altered sensitivity to oxidative stress and then determined whether alterations in stress resistance were associated with changes in lifespan. If ROS are a key contributor to aging, then it would be predicted that those strains that are resistant to oxidative stress would be long lived, whereas those that are sensitive to ROS would be short lived.
A. Genetic mutants with altered sensitivity to paraquat
Ishii et al. performed a screen to identify worms with altered sensitivity to paraquat (methyl viologen), a compound that induces oxidative stress (the strains identified were called mev to indicate their altered sensitivity to methyl viologen) (134). While both mev-1 and mev-2 worms were found to be sensitive to paraquat, only mev-1 worms exhibited decreased lifespan (134, 277). mev-2 worms were not sensitive to hyperoxia, indicating a dissociation between sensitivity to paraquat and to hyperoxia, despite the fact that both are thought to be mediated by increased ROS (277). This point is emphasized by the fact that mev-3 worms, which exhibit a normal lifespan, are resistant to paraquat but sensitive to hyperoxia (277).
mev-4 worms were found to be resistant to paraquat and long lived (76). Interestingly, the lifespan extension in mev-4 worms was daf-16-dependent, whereas the paraquat resistance was not, suggesting that different mechanisms are responsible for increased lifespan and resistance to paraquat. Further investigation revealed that sod mRNA levels were unaffected in mev-4 worms and that the causative mutation occurred in che-11, which affects the chemosensory cilia (76). It had previously been shown that che-11/mev-4 mutants, along with many other mutants with affected sensory cilia, exhibit extended longevity, which is partially dependent on daf-16 (6). Fujii et al. extended these findings to show that, as with daf-2 and age-1, which also act through the insulin/IGF-1 signaling pathway, the majority of the chemosensory cilia mutants were resistant to paraquat (76).
Finally, mev-5, mev-6, and mev-7 mutants were all found to exhibit increased resistance to paraquat, but only mev-5 showed an increase in lifespan, which was observed at 26°C and not 20°C (77). The mev mutants demonstrate that sensitivity to oxidative stress does not necessarily result in decreased lifespan, nor does resistance to oxidative stress result in long lifespan.
B. Genetic mutants with increased sensitivity to juglone
Johnson et al. examined the hypothesis that mutants with increased resistance to stress would exhibit long lifespan. To do this, a collection of long-lived strains and wild-type worms were used to determine the correlation between lifespan and resistance to juglone, heat, or UV irradiation. In fact, resistance to all three stressors was correlated with increased lifespan with correlation coefficients of between 0.51 and 0.65 (140). This suggests the possibility that general stress resistance, not just resistance to oxidative stress, may contribute to long life. However, this conclusion is complicated by the fact that UV irradiation and heat stress may act, at least partially, through the generation of ROS.
To further examine the relationship between resistance to juglone-induced oxidative stress and lifespan, De Castro et al. screened 50,000 worms that had undergone transposon-mediated mutagenesis to identify six strains that were resistant to juglone (52). The rationale for this experiment was that if ROS causes aging, then mutants that are resistant to increased oxidative stress should exhibit extended longevity. Examining the lifespan of the six juglone-resistant strains revealed that four out of six had increased lifespan. While this suggests a correlation between lifespan and resistance to oxidative stress, the fact that the strain that was most resistant to juglone-induced oxidative stress had the shortest lifespan weakens this conclusion.
C. Genetic mutants with increased sensitivity to hyperoxia and paraquat
In a screen for mutants that are sensitive to oxidative stress (hyperoxia and paraquat), Fujii et al. identified oxy-4, which encodes an [FeFe] hydrogenase-like protein (74). These mutants were sensitive to oxidative stress and showed decreased lifespan at 20°C. Interestingly, they were resistant to heat and UV stress and exhibited a trend toward increased lifespan at 26°C. Examination of antioxidant defenses revealed no significant changes in the level of sod-1, sod-2, sod-3, or sod-4 but increases in both ctl-1 and ctl-2 (74).
D. RNA interference screen for increased resistance to paraquat
In an experiment trying to link resistance to oxidative stress with extended lifespan, Kim and Sun screened the ∼6000 RNAi clones from chromosomes III and IV to identify clones that resulted in resistance to paraquat and then assessed whether these clones also resulted in increased lifespan (156). Of the 608 genes that conferred paraquat resistance, 84 were found to increase lifespan by at least 10%. While screening for paraquat resistance certainly enriches the number of clones that result in long life, the fact that 524 of the paraquat resistant clones do not increase lifespan indicates that paraquat resistance is clearly not sufficient for increased longevity. In addition, the authors note that the degree of paraquat resistance and lifespan extension showed little correlation (156).
E. Summary
Combining the results of the screens described above reveals an association between oxidative stress resistance and long life. However, the fact that many strains or RNAi-treated worms that have increased resistance to oxidative stress are not long lived suggests that resistance to oxidative stress alone is not sufficient for extended longevity. Similarly, while some strains with increased sensitivity to oxidative stress are short lived, others were shown to have a normal lifespan.
VI. Mitochondrial Mutants Have Divergent Effects on Lifespan
As the mitochondrion is one of the primary sites of ROS production in the cell, mutations that affect mitochondrial function are likely to have an impact on ROS generation, either increasing or decreasing ROS production. For example, if a fixed percentage of electrons undergoing electron transport are leaked to produce ROS, then a mutation that decreases the level of electron transport (i.e., the number of electrons that are transported in a given amount of time) should result in decreased production of ROS. Conversely, mutations affecting the mitochondria may increase ROS production by decreasing the efficiency of the ETC (i.e., increasing the percentage of electrons undergoing electron transport that are leaked to produce ROS).
In general, deletion mutations in genes encoding subunits of the ETC are lethal [e.g., refs. (124, 261)], as may be expected if energy production is prevented. Similarly, greatly decreasing expression of specific subunits of the ETC by undiluted RNAi can also lead to lethality (64, 230). In contrast, point mutations in multiple genes that directly affect ETC function (clk-1, isp-1, and nuo-6) can actually result in increased lifespan. This was first observed in the genetic mutant clk-1, where both deletions and point mutations have been found to increase lifespan (61, 274). However, as clk-1 affects electron transport somewhat indirectly (see below), the clearest demonstration of a lifespan-lengthening effect of modifying the ETC came from the characterization of isp-1(qm150), a single amino acid change in a catalytic subunit of mitochondrial complex III (66).
Subsequently, it has been found in RNAi screens for genes that extend lifespan that the most highly represented group of genes identified are those that affect mitochondrial function (94, 173). While these large scale screens were performed under conditions that prevent reproduction, thereby decreasing the overall energy demands of the worms (94, 173), other studies have demonstrated that RNAi against genes that affect mitochondrial function can increase lifespan under normal reproductive conditions (53, 230, 282). RNAi against genes encoding subunits of the ETC was found to exert its effect on lifespan only during development (53) and is dependent on the extent to which mitochondrial function is inhibited (230).
Increasing the concentration of RNAi directed against genes encoding subunits of the ETC resulted in a dose-dependent increase in lifespan only until a certain threshold was reached, after which further inhibition of mitochondrial function resulted in a sharp decline in longevity (230). Examining the lifespan and levels of carbonylated proteins in whole worms treated with different dilutions of RNAi against atp-3, which encodes a subunit of ATP synthase, revealed no correlation between levels of oxidized protein and lifespan (230). At high RNAi dilutions, protein carbonyls were increased and lifespan was unaffected. At intermediate RNAi dilutions, protein carbonyls were normal and lifespan was increased. At low RNAi dilutions, protein carbonyls were also normal while lifespan was decreased. Thus, although the technique of quantifying oxidatively damaged proteins is not precise, these findings suggest that the degree of oxidative damage may be unrelated to lifespan.
On the basis of phenotypic differences between point mutants and worms treated with RNAi, it was proposed that the mechanism underlying lifespan extension by the two types of intervention might be distinct (104). Phenotypic differences between point mutants and worms treated with RNAi were also noted by Dillin et al., who attributed the difference in size between isp-1 point mutants and worms treated with RNAi against ETC subunits to the degree to which mitochondrial function was inhibited (53). To distinguish between these two possibilities, Yang and Hekimi compared worms with point mutations in two mitochondrial genes, isp-1 and nuo-6 (see below for gene descriptions), with worms treated with RNAi against the same genes (282). Although both the point mutants and RNAi-treated worms were found to exhibit a similar increase in lifespan, the phenotype of these worms was markedly different, as was their mechanism of lifespan extension. For example, nuo-6(qm200) point mutants exhibit normal body size, a decreased rate of thrashing, decreased pharyngeal pumping, decreased oxygen consumption, increased ATP levels, paraquat resistance, and increased expression of SOD, whereas worms treated with nuo-6 RNAi exhibit decreased body size, an increased rate of thrashing, normal pharyngeal pumping, normal oxygen consumption, decreased ATP levels, normal sensitivity to paraquat, and normal expression of SOD (282).
As further evidence for separate lifespan-extending mechanisms, isp-1(qm150);nuo-6(qm200) double-point mutants exhibit a similar lifespan to the individual mutants isp-1(qm150) and nuo-6(qm200), suggesting that the two mutations affect the same processes and extend lifespan by the same mechanism (since lifespan is not further extended). In contrast, nuo-6 RNAi increases isp-1(qm150) lifespan and isp-1 RNAi increases nuo-6(qm200) lifespan, both by more than 10 days, suggesting that RNAi against these mitochondrial genes extends lifespan through a different mechanism than the point mutations (282). The authors propose that this difference may stem from the way in which RNAi and point mutations are likely to affect the ETC. In the case of the point mutation, it would be predicted that there would be a normal number of ETC complexes but that these complexes would have incorporated a subunit with a single amino acid error. In the case of RNAi, the amount of a particular subunit will be decreased. In some cases, such as nuo-6, this can result in an overall decrease in the amount of a particular mitochondrial complex, and possibly the whole ETC [a 46% knockdown of nuo-6 expression decreased the quantity of complex I and complex V by 39% and 42% respectively (64)]. Thus, the difference between point mutation and RNAi could be envisioned as a normal amount of abnormal complexes compared to a decreased amount of normal complexes, respectively. Accordingly, while both types of alteration result in increased lifespan, the underlying mechanism between point mutation and RNAi is different.
A. clk-1-hydroxylase involved in synthesis of ubiquinone
clk-1 (COQ7) was originally discovered in a screen for maternally rescued worms with altered physiology or development (103). As such, it was unexpected that these worms would also exhibit increased lifespan (274). The clk-1 gene encodes a hydroxylase involved in the synthesis of ubiquinone, which is involved in transferring electrons from complex I and complex II to complex III in the ETC, among other functions (61). To determine whether the long life of clk-1 mutants resulted from a decrease in electron transport and decreased ROS, a number of studies have examined clk-1 metabolism and oxidative stress. Felkai et al. observed decreased electron transport in isolated mitochondria (65), whereas others, using oxygen consumption as a measurement of mitochondrial function, have found that oxygen consumption is decreased in clk-1 worms (26, 283). In line with these findings, clk-1 mitochondria were found to have decreased oxidative phosphorylation with NADH-linked electron donors (63, 147). While ETC complex activity was found to be normal, electron transfer from complex I to complex III, but not from complex II to complex III, was found to be decreased in clk-1 worms, as measured spectrophotometrically using enzyme assays (147). Defects in ETC function in clk-1 worms has not been observed by all researchers. Miyadera et al. reported normal NADH cytochrome c reductase and succinate cytochrome c reductase activity (202), whereas Braeckman et al. observed normal or increased oxygen consumption, heat production, and superoxide generation potential (25, 26). Despite the decrease in mitochondrial function, clk-1 worms have increased levels of ATP (26), which presumably results from decreased energy expenditure.
Unlike other long-lived strains, clk-1 worms are not resistant to oxidative stress (112). While it was originally reported that they do not exhibit increased expression of sod-3 mRNA (112, 264), this has been observed in another report (283), and might therefore be sensitive to the conditions of growth. clk-1 worms have increased catalase activity (223, 257), which may contribute to their long lifespan, though loss of peroxisomal catalase (ctl-2) does not affect clk-1 lifespan (223). Measurements of mitochondrial ROS production in clk-1 mitochondria also suggest that clk-1 worms have an upregulated antioxidant defense. Intact clk-1 mitochondria were found to produce the same amount of ROS (H2O2) as wild-type mitochondria, while sub-mitochondrial particles from clk-1 worms produce significantly more ROS than wild type (284). Since sub-mitochondrial particles are thought to be lacking the antioxidant defenses present in the intact mitochondria, the authors concluded that clk-1 mitochondria produce more ROS than wild type but also have an increased antioxidant defense that counterbalances the elevated ROS.
Measurements of oxidative damage in clk-1 worms also suggest that these worms have increased antioxidant defenses, as clk-1 worms have been shown to have decreased levels of both carbonylated proteins and 4-HNE modified proteins (146, 283, 284). However, the decrease in oxidative damage does not appear to mediate the long lifespan of clk-1 worms as increasing oxidative damage (by using RNAi to knockdown sod-1 expression) did not decrease clk-1 lifespan (283).
The contribution of altered ROS to the phenotype of clk-1 worms is suggested by the modulation of the clk-1 phenotype that results from altering the levels of SOD. Decreasing sod-1 expression by RNAi suppresses the delayed egg production and slow germline development of clk-1 worms, suggesting that decreased cytoplasmic ROS may contribute to these phenotypes (246). Similarly, decreased expression of either sod-1 or sod-4 reduces clk-1's ability to suppress the multivulva phenotype caused by gain-of-function mutations in ras (let-60) (246).
Finally, RNAi knockdown or deletion of sod-2 results in an extension of clk-1 lifespan (263, 283). This suggests the possibility that increased mitochondrial ROS contributes to the longevity of clk-1 worms. Overall, decreased mitochondrial function in clk-1 worms results in increased production of ROS, a compensatory increase in antioxidant defenses and decreased levels of oxidatively damaged proteins. However, decreased oxidative damage in clk-1 worms can be dissociated from their long life, and increasing ROS, at least in some cellular compartments, can actually increase clk-1 lifespan.
B. isp-1-Rieske iron sulfur protein subunit of complex III
isp-1 (UQCRFS1) encodes the Rieske iron sulfur protein, which is part of complex III of the ETC (66). isp-1(qm150) point mutants are very long-lived mutants and were originally identified in a screen in which slow-growing, slow-defecating mutants were examined for lifespan [isp-1(gk267) deletion mutants were also found to be long lived (230)]. isp-1(qm150) worms have decreased whole worm oxygen consumption (66) and decreased oxidative phosphorylation in isolated mitochondria when using either malate or succinate as an electron donor (63). On the basis of the fact that the mitochondrial function of isp-1(qm150) worms was found to be decreased, it was initially suggested that the long life of isp-1(qm150) mutants resulted from decreased ROS production. To assess whether isp-1(qm150) worms did in fact have increased ROS production, Feng et al. examined their sensitivity to paraquat during development with the prediction that if ROS production was decreased then isp-1(qm150) worms would be less sensitive to paraquat. However, the experiment was complicated as isp-1(qm150) worms normally take twice as long to develop as wild-type worms. As a result, it was found that isp-1(qm150) worms failed to develop when exposed to paraquat. While this result is consistent with increased ROS levels in isp-1(qm150) worms, the characterization of isp-1(qm150);ctb-1 double mutants suggests that this may not be the case.
ctb-1 encodes cytochrome b, which is the only subunit of complex III encoded by a gene of the mitochondrial genome (mtDNA). A missense mutation in ctb-1 was found to restore the slow development of isp-1(qm150) worms toward wild type without affecting their lifespan (66). In contrast to isp-1(qm150) worms, isp-1(qm150);ctb-1 double mutants were found to be more resistant to paraquat during development than wild-type N2 worms. This suggests the possibility that isp-1(qm150) worms are more resistant to oxidative stress than wild-type worms but that this effect is masked by their long development time in this assay (66). Alternatively, the mutation in ctb-1 could increase the resistance of isp-1(qm150) worms to oxidative stress. Consistent with the former possibility, sod-3 expression was found to be increased in isp-1(qm150) worms (66). Deletion of daf-16 prevents the increase in sod-3 levels in isp-1(qm150) worms but does not decrease their lifespan, and this suggests that the increased levels of sod-3 do not contribute to the long life of isp-1(qm150) worms (66). Attempts to directly measure ROS levels in isp-1(qm150) worms through MitoSOX staining in the pharynx revealed no differences from wild-type worms (54).
Examination of oxidative damage to protein revealed a trend toward decreased levels of carbonylated proteins in isp-1(qm150) worms compared with wild-type worms, but the difference was not significant (283). To eliminate the possibility that decreased oxidative damage contributed to isp-1(qm150)'s long lifespan, Yang et al. treated isp-1(qm150) worms with RNAi against sod-2, the primary mitochondrial sod gene. isp-1(qm150) worms treated with RNAi against sod-2 had significantly more carbonylated proteins than wild-type worms but still exhibited the same long life of isp-1(qm150) worms (283). This suggests that the long lifespan of isp-1(qm150) worms does not result from a decrease in oxidative damage. This is supported by the observation that 4-HNE modified proteins are increased in isp-1(qm150) worms (54). Thus, as with clk-1, the long life of isp-1(qm150) mutants cannot be explained by a decrease in oxidative damage.
C. nuo-6–NADH ubiquinone oxidoreductase subunit 6 (complex I)
The nuo-6(qm200) mutation was originally identified in a screen for slow development and slow defecation (282). nuo-6 (NDUFB4) encodes a subunit of complex I of the ETC and accordingly complex I activity as well as whole worm oxygen consumption are both reduced in nuo-6(qm200) mutant worms. Similar to clk-1 worms, nuo-6(qm200) worms have increased levels at ATP in spite of their decreased mitochondrial function (282). Examination of lifespan and sensitivity to oxidative stress revealed that nuo-6(qm200) worms are very resistant to paraquat-induced oxidative stress and are long lived (282). Resistance to paraquat likely results from the observed increase in both SOD-1 and SOD-2 protein levels in nuo-6(qm200) worms. As mentioned above, RNAi against nuo-6 also increases lifespan but by a mechanism that is distinct from that of the nuo-6(qm200) point mutant (282). RNAi against nuo-6 (or isp-1) does not result in decreased oxygen consumption, altered SOD levels, or resistance to paraquat but instead appears to be acting by triggering a mitochondrial stress response (282).
D. lrs-2–mitochondrial leucyl-tRNA synthetase
lrs-2 (LARS) was identified in a screen for mutations that extended the lifespan of the short-lived daf-16 mutant (173). This gene was found to encode the mitochondrial leucyl-tRNA synthetase, which is responsible for joining the amino acid leucine to its corresponding tRNA to be used in translation. While lrs-2 is a nuclear-encoded gene, mutations in this gene would be expected to affect genes expressed from the mitochondrial genome, which encodes 12 proteins that are involved in mitochondrial function. Despite the fact that lrs-2 mutants displayed abnormal mitochondrial morphology, these worms exhibited a marked increase in lifespan (173).
To determine whether the increase in lifespan resulted from a decrease in ROS production stemming from impaired mitochondrial function, Lee et al. examined sensitivity to paraquat and H2O2. Surprisingly, opposite results were obtained for the two inducers of oxidative stress: lrs-2 worms showed increased sensitivity to paraquat, but dramatically increased resistance to H2O2 (173). This phenomenon did not result from a specific peculiarity of lrs-2 as increased sensitivity to paraquat in combination with increased resistance to H2O2 was also observed in worms treated with RNAi against genes involved in mitochondrial function (173). One possible explanation for this observation would be that there was a compensatory upregulation of catalase or peroxiredoxin without a change in SOD levels, but this was not examined. Nonetheless, this finding illustrates the point that not all means of inducing oxidative stress are equal. As both lrs-2 point mutants and worms treated with RNAi against genes encoding proteins that act in the mitochondria were long lived, this indicates an association between H2O2 resistance and lifespan but that paraquat sensitivity can be dissociated from decreased lifespan.
E. mev-1–succinate dehydrogenase subunit C (complex II)
As described above the mev-1 mutation was identified in a screen for mutants with altered sensitivity to paraquat (134). mev-1 (SDH-C) was later found to encode succinate dehydrogenase subunit C (sdhc-1) (131) [see ref. (132) for review]. Accordingly, mev-1 worms have decreased complex II activity but normal ATP levels, suggesting decreased utilization of energy (242). It is also possible that the defect in complex II activity is compensated for by an increase in electron transport from complex I to complex III, as electron transport does not function at its maximum rate under normal conditions. In contrast to the long life exhibited by clk-1, isp-1, and nuo-6 worms, mev-1 worms are short lived (134, 281). In addition, no level of mev-1 inhibition by RNAi was found to increase lifespan (126, 230). The fact that RNAi inhibition of another component of the ETC (cyc-1; cytochrome c reductase) was able to increase the lifespan of mev-1 suggests that decreased mitochondrial function may not be the cause of mev-1's short lifespan (47). However, measurement of mitochondrial function would be necessary to confirm this conclusion, as mutations in ETC components and RNAi against these components appear to affect lifespan independently (282).
mev-1 worms are sensitive to paraquat and hyperoxia but not heat stress, suggesting a specific sensitivity to oxidative stress (100, 134, 281). Congruent with an increased sensitivity to oxidative stress, mev-1 worms have decreased levels of sod mRNA, and decreased SOD activity but exhibit an increase in the levels of catalase mRNA (134, 281). mev-1 worms have been shown to have increased levels of superoxide in the mitochondria (160, 242, 279) and this ROS was shown to be produced from complex II (133, 242). mev-1 worms also show increased oxidative damage to proteins (54, 146, 281) and DNA (99). While some researchers have observed increased accumulation of the age pigment lipofuscin compared to wild type (115), others have reported decreased accumulation (85).
mev-1 worms also show an increased rate of mutation that was exponentially increased by increasing oxygen levels and may contribute to their decreased lifespan (99). The fact that daf-16 is found to be translocated to the nucleus in mev-1 worms suggests that their increased oxidative stress initiates a stress response (160). Decreased levels of glutathione are also indicative of increased oxidative stress (242). A role for increased oxidative stress in causing the short lifespan of mev-1 worms is suggested by the fact that multiple antioxidants have been shown to increase the lifespan of mev-1 worms, including the SOD/catalase mimetics Euk-8 and Euk-124 (194), coenzyme Q10 (133), and epigallocatechin gallate (EGCG) (2). It should be noted, however, that some other antioxidants, such as Vitamin E (133), fail to improve mev-1 lifespan, suggesting the possibility that other properties of the antioxidants may be improving lifespan in mev-1 worms. For example, in addition to its ability to act as an antioxidant, coenzyme Q10 also acts as an electron carrier in the ETC and thus may exert its effects through increasing the efficiency of electron transfer in the ETC.
Interestingly, not all groups have observed detrimental phenotypes in mev-1 worms. For example, in some cases paraquat sensitivity was not observed during adulthood (235). In another example, mev-1 worms were found to have a normal lifespan despite markedly increased oxidative damage, as measured by 4-HNE levels in the mitochondria (54). In this experiment, measurement of ROS levels by MitoSOX staining in the pharyngeal bulb failed to observe a difference between mev-1 and wild-type worms (54). In addition, SOD mRNA, protein and activity were found to be increased, although the increase in SOD activity was not significant (54). This suggests the possibility that under certain conditions, mev-1 worms can upregulate their antioxidant defenses and live a normal lifespan. Of note, however, is the fact that the normal lifespan is achieved despite a 50% increase in oxidative damage to protein (54).
In addition to the putative role of increased ROS levels in determining mev-1 lifespan, some evidence suggests that abnormal apoptosis contributes to the short lifespan of mev-1 worms (241). The number of apoptotic cells in developing embryos was found to be increased in mev-1 worms. In addition, the protein levels of CED-9, which encodes the C. elegans homolog of the cell death inhibitor Bcl-2, were not increased under hyperoxia as they are in wild-type worms (241). The authors propose that increased ROS may directly damage CED-9 protein and indirectly affect CED-9 binding to the mitochondrial membrane through oxidative damage to the lipid bilayer. However, it is also possible that the mev-1 mutation more directly affects expression of ced-9 as these two genes are found on the same operon (218). In support of a role for apoptosis in causing mev-1's short lifespan, deletion of ced-3, which acts downstream of CED-9, partially restored the decreased lifespan of mev-1 worms toward wild type (241). Overall, the mev-1 mutation decreases complex II activity, affecting mitochondrial function, which results in increased production of ROS and abnormal apoptosis, both of which may contribute to their decreased lifespan.
F. gas-1 and nduf-2.2–iron sulfur protein subunit of complex I
gas-1 (NDUFS2) was originally identified in a screen for hypersensitivity to gas anesthetics (204) and was later found to encode an iron protein subunit of complex I of the ETC (145). Accordingly, gas-1 worms have decreased electron transport through complex I but increased activity of complex II (144). Like mev-1 worms, gas-1 worms are short lived and sensitive to both paraquat and hyperoxia during development and adulthood (100). In addition, gas-1 worms have increased oxidative damage to mitochondrial proteins (54, 146), but decreased age pigment accumulation compared to wild-type worms (85). In examining superoxide levels, it was found that gas-1 worms have increased superoxide in submitochondrial particles but decreased superoxide in intact mitochondria (160). The authors proposed that this discrepancy resulted from increased expression of SOD in the mitochondria, which could compensate for increased ROS production. In line with this suggestion, daf-16 is found in the nucleus in gas-1 worms, suggesting that increased ROS production triggers a stress response that would then upregulate mitochondrial SOD expression (160). This conclusion has subsequently been supported by the observation of increased levels of mitochondrial sod mRNA and SOD protein in gas-1 worms (54).
The gas-1 paralog nduf-2.2 (T26A5.3) is able to fully rescue the gas-1 mutation when expressed under the gas-1 promoter (144). This suggests that nduf-2.2 encodes another complex I iron sulfur protein. The fact that expression of nduf-2.2 is very low indicates that gas-1 encodes the primary iron sulfur protein for complex I, though it is possible that nduf-2.2 exhibits increased expression under certain conditions or in certain tissues. Despite its low level of expression, deletion of nduf-2.2 results in decreased lifespan (146). The fact that nduf-2.2 worms do not exhibit sensitivity to oxygen suggests that gas-1's sensitivity to oxidative stress may not cause its shortened lifespan (146). Interestingly, gas-1;nduf-2.2 double mutants are sterile but live about four times longer than gas-1 despite maintaining gas-1's sensitivity to hyperoxic conditions. Similarly, the gas-1 mutation has been found to markedly increase the already long lifespan of clk-1 worms despite the fact that gas-1 shortens lifespan by itself (146). Thus, while gas-1 worms have increased sensitivity to oxidative stress and decreased lifespan, data from nduf-2.2 worms and gas-1;nduf-2.2 double mutants suggest that these two phenotypes can be dissociated.
G. nuo-1–NADH ubiquinone oxidoreductase subunit 1 (complex I)
nuo-1 (NDUFV1) encodes a subunit of complex I. Homozygous deletions in nuo-1 are lethal, as nuo-1 mutant worms fail to develop past the third larval stage (261). Taking advantage of the fact the nuo-1 deletion mutants could be rescued by nuo-1 transgenes, Grad et al. generated specific nuo-1 point mutants by expressing nuo-1 with various point mutations from a transgene in homozygous nuo-1 deletion worms (91). The rescued worms show decreased oxygen consumption, decreased ETC function (especially complex I), decreased lifespan, and increased sensitivity to hyperoxia and paraquat (91). These worms show an association between sensitivity to oxidative stress and decreased lifespan.
H. sdhb-1–succinate dehydrogenase subunit b (complex II)
Huang et al. examined a series of mutations in sdhb-1 (SDHB), which encodes succinate dehydrogenase subunit B (iron sulfur subunit of complex II). Worms that are homozygous for this mutation arrest as L3 larvae. As with nuo-1, this phenotype could be rescued by various point mutants, most of which showed decreased lifespan as well as increased sensitivity to hyperoxia and paraquat (124). These mutants were shown to have elevated production of superoxide, and their survival under hyperoxia was rescued by treating with antioxidants (either N-acetyl cysteine or ascorbate/Vitamin C). Treatment with ascorbate increased the lifespan of the various sdhb-1 mutants but did not significantly increase the lifespan of wild-type worms, suggesting that increased ROS is responsible for their decreased lifespan (124). Thus, mutations in sdhb-1 lead to increased ROS production that appears to contribute to their decreased lifespan.
I. phb-1- and phb-2-prohibitins
Prohibitins (PHB and PHB2) are mitochondrial proteins that are essential for development (215). These proteins form a ring structure in the inner mitochondrial membrane composed of prohibitin-1 and prohibitin-2 subunits. While the function of this complex is currently unknown, it is proposed to act as a protein scaffold and appears to be important in maintaining mitochondrial morphology. Artal-Sanz et al. examined the effect of prohibitin depletion by RNAi knockdown during adulthood, as mutations in phb-1 result in sterility and RNAi against phb-1 or phb-2 leads to embryonic lethality (8). They found that RNAi knockdown of either phb-1 or phb-2 resulted in decreased lifespan in wild-type worms at 20°C (but increased lifespan at 25°C). In contrast, decreased expression of prohibitin increased the lifespan of both short-lived (mev-1, gas-1) and long-lived (clk-1, isp-1) mitochondrial mutants, as well as mutants with disrupted dauer formation (e.g., daf-2). Examination of ROS production in whole worms by DCF staining and resistance to paraquat revealed that depletion of prohibitin increased ROS production and decreased paraquat resistance in both wild-type and daf-2 worms (8). This suggests either that increasing ROS levels had opposite effects on wild-type (decreased) and daf-2 (increased) lifespan or that the observed increase in ROS levels in these two strains did not affect their longevity.
J. frh-1-frataxin
Frataxin is a mitochondrial matrix protein that acts as an iron donor in the formation of iron–sulfur clusters. A trinucleotide GAA repeat expansion in intron 1 of FXN, the human frataxin gene, causes an autosomal recessive neurodegenerative disorder called Friedreich ataxia, which among other symptoms results in premature death. Mutations in the FXN gene result in decreased expression of frataxin, which can subsequently affect multiple subunits of the ETC that contain iron–sulfur clusters (complexes I, II, and III). As with mutations that encode mitochondrial ETC subunits, null mutations in frh-1 are sterile and do not develop past the L3 stage (270).
Decreasing levels of frh-1 by RNAi can either increase lifespan (270, 271) or decrease lifespan (90, 269, 289) depending on the extent to which frataxin levels are reduced (230, 271). Under frh-1 RNAi knockdown conditions that lead to decreased lifespan (presumably more severe knockdown though this was not measured), worms were found have increased sensitivity to paraquat (269). Under frh-1 RNAi knockdown conditions that resulted in increased lifespan (30%–70% reduction in frh-1 mRNA), worms were found to be more resistant to H2O2 but more sensitive to juglone (270). Thus, increased sensitivity to oxidative stress is observed under conditions that lead to both decreased and increased lifespan. Interestingly, the different effects of frh-1 knockdown on lifespan may both be mediated by cep-1, the p53 homolog in C. elegans [though only its role in increasing lifespan was demonstrated (271)]. In the case of increased lifespan, the effect of frh-1 knockdown on longevity does not require daf-16 or skn-1 (271) but may be partially due to observed increases in expression of gst-4 and other stress response genes (260).
K. Uncoupling increases lifespan
Uncoupling proteins uncouple the ETC from ATP synthesis by providing an alternate route for reducing the mitochondrial inner membrane potential (58). Accordingly, uncoupling proteins have been proposed to have a role in reducing ROS levels since more ROS are produced when the membrane potential is high (161, 201). In fact, uncoupling proteins have been shown to be activated by superoxide (59) and have been examined for their role in aging (29). A decreased mitochondrial membrane potential has been associated with increased lifespan in C. elegans (175). Under conditions where the ETC is inhibited, uncouplers can result in increased rather than decreased production of ROS (262).
As much of the previous work on uncoupling proteins had been examined in mice, Iser et al. sought to determine whether there were uncoupling proteins present in the C. elegans genome and whether these had an impact on aging. They found that in C. elegans there is only one putative uncoupling protein, UCP-4 (SLC25A27). While the ability of UCP-4 to act as an uncoupling protein was not directly assessed, the increased levels of ATP and increased inner membrane potential observed in ucp-4 mutant worms are consistent with an uncoupling function (128). To determine whether these worms produced increased levels of ROS, Iser et al. examined sensitivity to paraquat but found no change from wild type (128). These worms also exhibited a normal lifespan (128).
In addition to experiments examining the genetic mutant ucp-4, multiple groups have examined the effect of chemical uncouplers on lifespan. These studies have found that treatment of worms with chemical uncouplers, such as carbonylcyanide-3-chlorophenylhydrazone (CCCP) and carbonylcyanide-4-trifluoromethoxyphenylhydrazone (FCCP), results in extended longevity (175, 203, 237, 238). The fact that one of these uncouplers, FCCP was shown to reduce the levels of mitochondrial ROS suggests that the positive effects on lifespan may be due to decreased production of ROS (203, 237). Thus, although deletion of the gene encoding the only putative uncoupling protein in C. elegans did not yield the predicted increase is ROS (at least as far as could be measured by paraquat sensitivity), chemical uncouplers were effective at increasing lifespan in association with the predicted decrease in ROS.
L. Mutations affecting mitochondrial function increase lifespan without decreasing ROS
The Clk mutants of C. elegans were identified in screens for maternally rescued worms that exhibit slow development and slow defecation (103, 264, 274). Despite the fact that slow development and physiology were the only screening criteria, all 10 Clk mutants were found to have decreased mitochondrial function, as measured by oxygen consumption, and increased lifespan (247, 264). To determine whether the long life of these mutants resulted from decreased ROS production stemming from decreased levels of electron transport, Van Raamsdonk et al. examined sensitivity to oxidative stress and oxidative damage in the Clk mutants. With the exception of clk-2 worms, all of the Clk mutants exhibited increased sensitivity to oxidative stress either during development, adulthood, or both, and there was no evidence for decreased oxidative damage (264). Thus, the long life of the Clk mutants was found not to result from decreased production of ROS and, in most cases, occurred in spite of increased sensitivity to oxidative stress.
M. Summary
Mutations affecting mitochondrial proteins can have either detrimental or beneficial effects on lifespan depending on how mitochondrial function is altered (Figs. 11 and 12). A number of mitochondrial mutants (mev-1, gas-1, sdhb-1, phb-1 RNAi, and phb-2 RNAi) exhibit increased levels of ROS, decreased resistance to oxidative stress, and decreased lifespan, thereby supporting an association between high ROS levels and decreased lifespan. However, clk-1 worms also exhibit increased levels of ROS and decreased resistance to oxidative stress but are long lived. Similarly, while mev-1 and gas-1 worms exhibit increased oxidative damage and decreased lifespan, it is possible to increase oxidative damage beyond wild-type levels in both clk-1 and isp-1 worms without decreasing lifespan. This conclusion is further supported by the fact that two mutations that have been shown to increase oxidative damage (sod-2 and gas-1) markedly increase the already long lifespan of clk-1 worms (see Table 3 for a summary of lifespan and oxidative stress resistance in mitochondrial mutants).
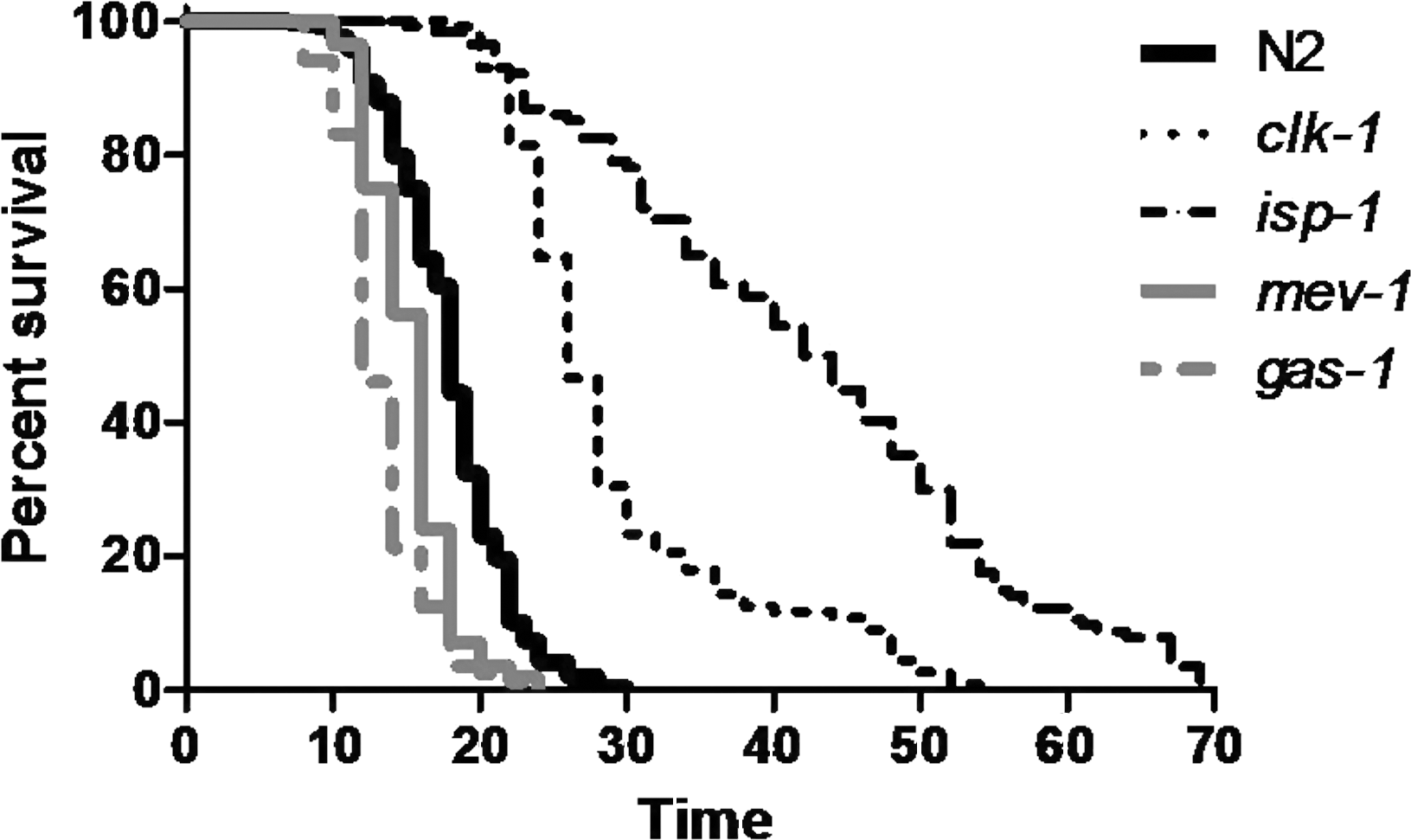
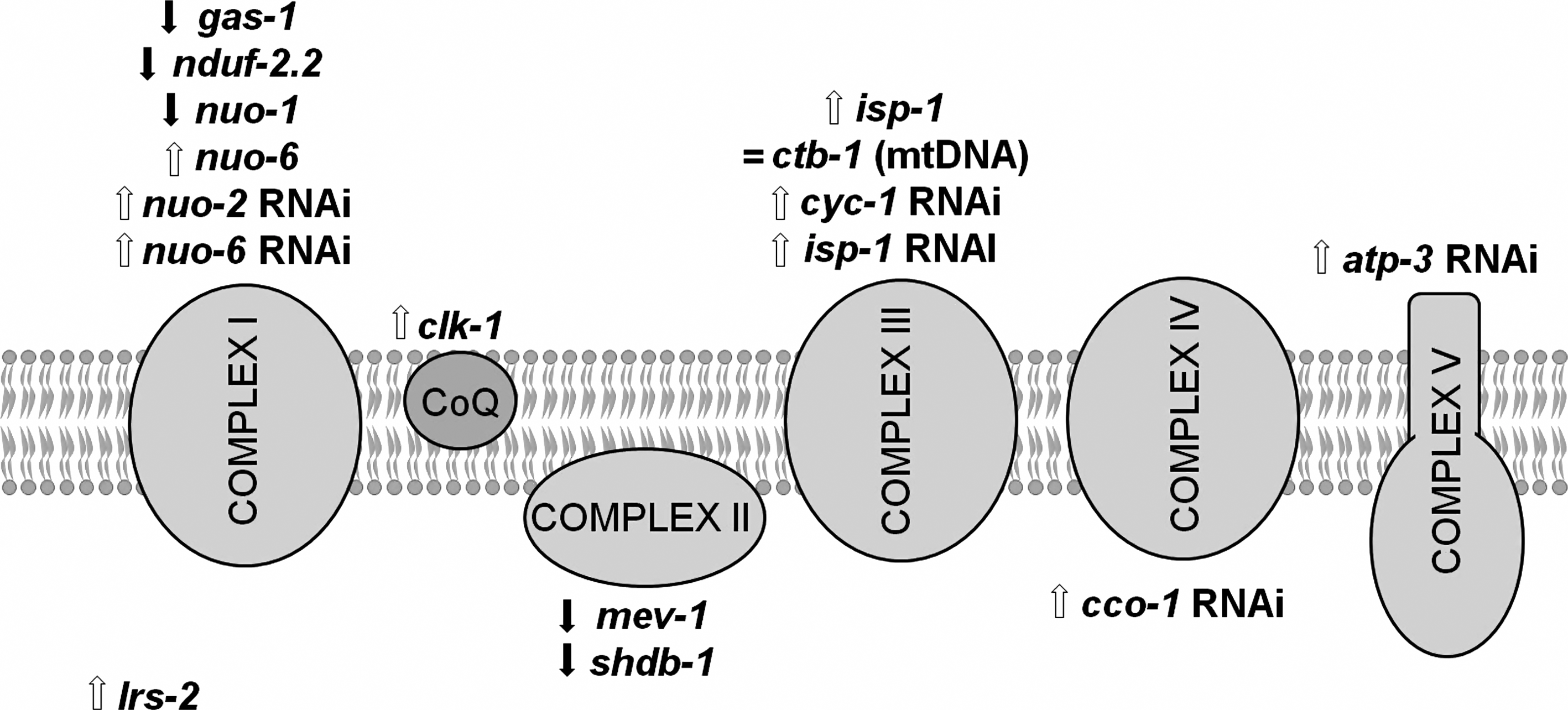
VII. Endogenous Antioxidant Defenses
A more direct approach to examining the relationship between ROS and aging is to experimentally increase ROS and examine its effect on lifespan. Increasing ROS can theoretically be accomplished by decreasing antioxidant defenses. However, care must be taken to ensure that ROS levels are actually increased by this approach as it is possible that other genes may compensate for the function of a deleted gene. C. elegans mutants with deletions in numerous antioxidant enzymes have been characterized with respect to ROS detoxification and lifespan.
A. Superoxide dismutase: sod-1, sod-2, sod-3, sod-4, and sod-5
Superoxide dismutase (SOD) is an antioxidant enzyme that converts superoxide into H2O2, which can subsequently be converted to water. While most organisms have three SODs, C. elegans has five sod genes (167). sod-1, sod-2, and sod-4 encode the primary cytoplasmic, mitochondrial, and extracellular SODs, respectively, whereas sod-3 and sod-5 appear to encode secondary SODs in the mitochondria and cytoplasm that are markedly upregulated under certain conditions such as dauer and oxidative stress (Fig. 13A) (56, 75, 86, 87, 125, 137, 255). In a wild-type adult worm, sod-1, sod-2, sod-3, sod-4, and sod-5 account for 76%, 18%, 1%, 5%, and 0.5% of total sod mRNA, respectively (56). Numerous mutations and interventions that extend lifespan have been shown to increase SOD expression, including decreasing insulin/IGF signaling (112, 172, 189, 217, 283), decreasing mitochondrial function (66, 283), ablation of the germline (177), and dietary restriction (118, 121, 217). It has also been found that sod-3 expression is increased by oxidative stress (278).
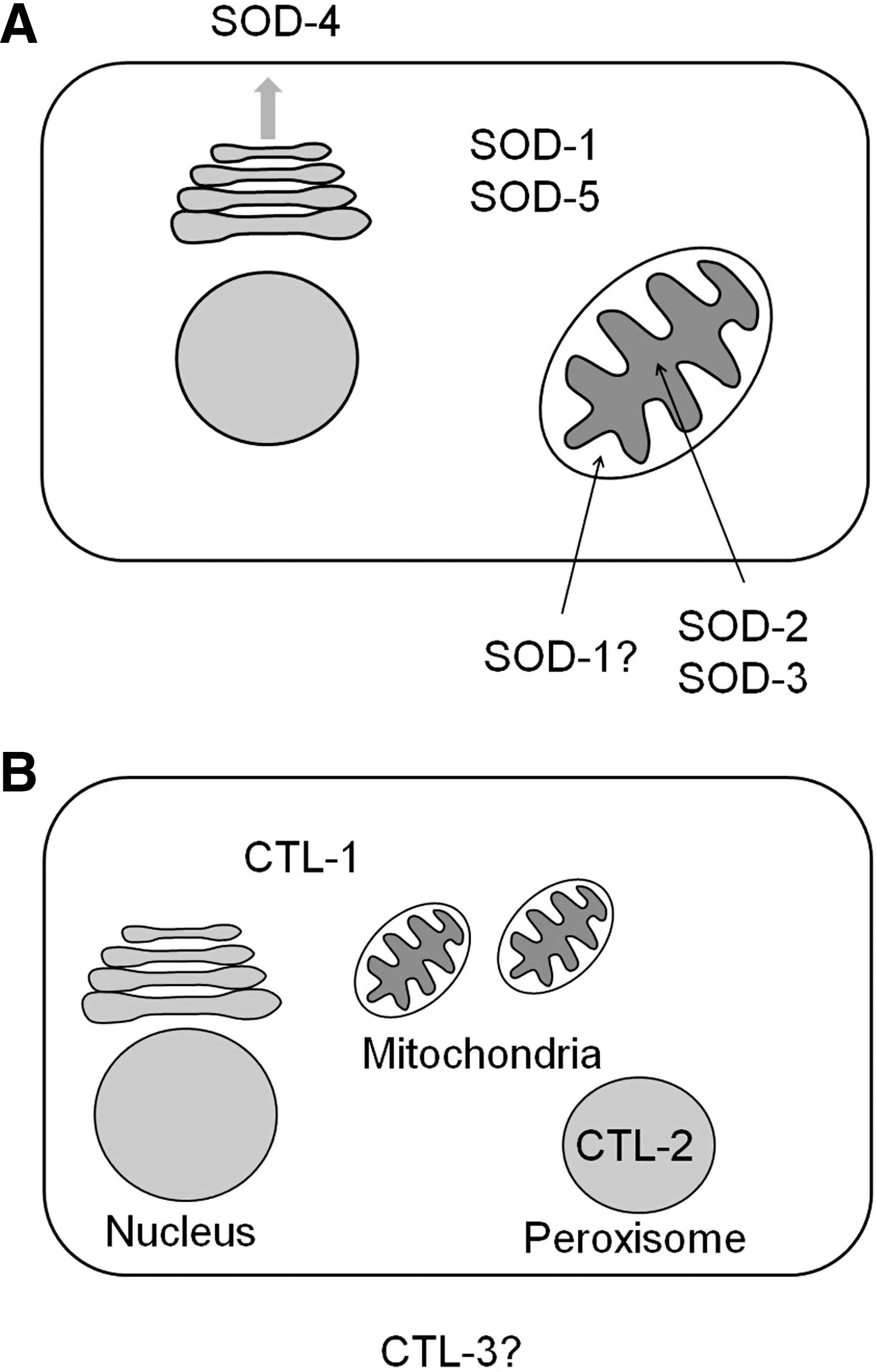
A number of groups have examined the effect of deleting individual or combinations of sod genes on lifespan and oxidative stress in C. elegans. Using an RNAi approach, it was found that knockdown of either sod-1 or sod-2 increased sensitivity to paraquat (150% increase in fraction dead after 72 h on 4 mM paraquat) and increased protein carbonylation (30%–40% increase) but only had a minor effect on lifespan in the case of sod-1 (7% decrease) or no effect on lifespan in the case of sod-2 (283). Honda et al. examined sod-2, sod-3, and sod-4 deletion mutants and found that only sod-2 mutants exhibited increased sensitivity to a combined challenge with paraquat and 98% oxygen and all three mutants had a normal lifespan (114). They also found that sod-2;sod-3 double mutants, which have no mitochondrial SOD, were very sensitive to hyperoxia but had a normal lifespan (114).
Doonan et al. examined deletion mutants affecting all five sod genes and found that sod-1 mutant worms were sensitive to paraquat and hyperoxia, whereas sod-2 worms were only sensitive to paraquat and to a lesser extent than sod-1 (56). In agreement with the increased sensitivity to oxidative stress in sod-1 worms, these worms exhibited an 80% decrease in total SOD activity and were found to have a modest 15%–31% decrease in lifespan. Worms engineered to overexpress sod-1 exhibited a modest increase in lifespan (56). The fact that both sod-1 deletion mutants and sod-1-overexpressing worms exhibited increased sensitivity to paraquat suggests that the decreased lifespan of sod-1 worms does not result from their increased sensitivity to oxidative stress. Similarly, it was found that sod-2;sod-3 double mutants are much more sensitive to hyperoxia than sod-1 mutant worms but do not have decreased lifespan, thereby dissociating sensitivity to hyperoxia from lifespan (56).
Yen et al. also examined deletion mutants affecting all five sod genes and observed no effect of sod deletion on lifespan under normal conditions (286). They observed increased sensitivity to paraquat in adult sod-1 worms as well as sod-2, sod-3, and sod-4 worms (not sod-1) during development (286). In contrast to what others have observed, they observed increased resistance to paraquat in adult sod-3 and sod-4 worms (286).
Van Raamsdonk et al. observed no decrease in lifespan for deletion of any individual sod gene or double mutant containing sod-1 or sod-2, despite increased sensitivity to paraquat and juglone (263). This included sod-1;sod-5 double mutants, which completely lack cytoplasmic SOD, and sod-2;sod-3 double mutants, which are completely missing SOD from the mitochondrial matrix. Although there was a mild compensatory increase of other sod mRNAs, there was no detectable increase in SOD-1 or SOD-2 protein in any of the sod deletion mutants. In addition, the compensatory increase in sod mRNA was not sufficient to prevent the increased sensitivity to oxidative stress. Moreover, even sod-1;sod-2;sod-4 triple mutants, which lack the primary cytoplasmic, mitochondrial, and extracellular sod genes, were found to live at least as long as wild-type worms. A subsequent study examining gene expression changes in the sod deletion mutants found no significant upregulation of sod mRNAs but did observe increased expression of various stress response genes controlled by skn-1 in sod-1 and sod-2;sod-3 mutant worms (14).
Interestingly, sod-2 worms were found to live longer than wild-type worms despite being sensitive to oxidative stress and showing an increase in oxidative damage (263). This finding is congruent with the observation that sod-2 RNAi was found to extend the lifespan of clk-1 worms (283). Long life and increased oxidative damage in sod-2 worms was also observed by Dingley et al. (54). These authors also reported increased ROS levels in both sod-2 and sod-3 mutant worms, as measured by MitoSOX staining in the pharyngeal bulb (54).
Since impacting mitochondrial function can result in increased lifespan, a plausible mechanism for the effect on sod-2 deletion on lifespan would be that (a) sod-2 mutants have increased mitochondrial ROS resulting from decreased detoxification, (b) the increased ROS causes an increase in oxidative damage to proteins (and possibly lipids and DNA), (c) the increased oxidative damage causes decreased mitochondrial function (as measured by oxygen consumption), and (d) the decrease in mitochondrial function leads to increased lifespan by a type of retrograde signaling to the nucleus as has been proposed (47).
Finally, Yanase et. al observed decreased lifespan and increased sensitivity to paraquat in mutants lacking sod-1 (280). In addition, the sod-1 deletion mutants were shown to have increased production of superoxide in both cytosolic and mitochondrial fractions, as well as a twofold compensatory increase in sod-5 mRNA expression (280).
While some of the results differed between these reports, it seems clear that deletion of sod-1 or sod-2, the primary cytoplasmic and mitochondrial sod genes, respectively, results in increased oxidative stress but that the deletion of sod genes results in either little or no decrease in lifespan. The fact that worms with either no mitochondrial SOD (sod-2;sod-3 double mutants) or no cytoplasmic SOD (sod-1;sod-5 double mutants) can live as long as wild-type worms suggests either that the worms can compensate for the complete loss of SOD activity in specific cellular compartments or that SOD activity is not required to live a normal lifespan.
B. Catalase: ctl-1, ctl-2, and ctl-3
Catalase is an antioxidant enzyme that converts H2O2 into water. C. elegans has three catalase genes (ctl) that exist in tandem at one locus within the genome (223). ctl-1 is expressed in the cytoplasm, whereas ctl-2 is found in the peroxisome (Fig. 13B). There is no catalase present in the mitochondria. Deletion of ctl-1 and ctl-2 led to a 25% and 80% decrease in whole worm catalase activity, respectively, indicating that ctl-2 accounts for the majority of catalase activity in the worm (223). Little is known of the subcellular localization of ctl-3, but based on these activity measurements it would appear to have a very minor contribution to the overall catalase activity, at least under normal conditions. RNAi against ctl-3 has revealed no abnormal phenotypes and no mutant strains have been reported.
As with the sod genes, catalase genes are also found to be upregulated in long-lived mutants, including age-1, daf-2, eat-2, and clk-1 worms (118, 121, 169, 266, 281). Examination of the lifespan of ctl-1 and ctl-2 deletion mutants revealed that the lifespan of ctl-2 mutants was 16% shorter than wild type (223). In contrast to what might be predicted based on decreasing antioxidant activity, protein carbonyl levels in ctl-1 and ctl-2 mutants were normal at day 5 and significantly decreased at day 10 compared with wild-type worms (223). Interestingly, the simultaneous overexpression of all three catalase genes resulted in a 10% decrease in lifespan (56).
C. Peroxiredoxin: prdx-2, prdx-3, and prdx-6
Peroxiredoxins are antioxidant enzymes with peroxidase activity [see ref. (275) for review]. These enzymes can detoxify H2O2, alkyl hydroperoxide, and peroxynitrite and can protect against lipid peroxidation. The antioxidant activity of peroxiredoxins is dependent on either one or two cysteine residues, which reduce its substrates (e.g., reduction of H2O2 to water). The peroxidase activity of peroxiredoxins can subsequently be restored by TRX or glutaredoxin.
C. elegans was found to have three peroxiredoxins (129). Isermann et al. examined the effect of RNAi against each of the three prdx genes and found no differences when either prdx-3 (ceprx-1) or prdx-6 (ceprx-3) was knocked down. In contrast, RNAi knockdown or deletion mutants of prdx-2 (ceprx-2) showed reductions in size and fertility (129). Further study of prdx-2 revealed that prdx-2 is upregulated in response to oxidative stress and is able to detoxify H2O2. Nonetheless, prdx-2 worms were not more sensitive to H2O2 than wild-type worms and exhibited a normal lifespan (129). The authors suggest that this results from the fact that they found prdx-2 expression to be limited to two types of pharyngeal neurons and proposed that prdx-2 may have a role in signaling (129).
In contrast, Olahova et al. found that prdx-2 mutants are hypersensitive to H2O2 and exhibit a 19% decrease in lifespan compared with wild-type worms (214). These differences likely result from differences in the way in which lifespan and H2O2 sensitivity were measured. Isermann et al. measured lifespan at a higher temperature and observed a nonsignificant decrease of 7%, whereas Olahova et al. assessed lifespan at 15 and 20 degrees and observed significant decreases of 26% and 19%, respectively. Similarly, Isermann et al. measured resistance to H2O2 more acutely in a 2 h assay at 5 mM concentration, whereas Olahova et al. examined the effects of 1 mM H2O2 and observed differences only after 12 h. The sensitivity of prdx-2 worms to oxidative stress was also observed with paraquat (214). In addition, Olahova et al. found that expression of prdx-2 also occurred in the intestine as well as the pharyngeal neurons. Interestingly, intestinal expression of peroxiredoxin-2 eliminated the sensitivity of prdx-2 deletion mutant worms to H2O2 but did not restore their lifespan to wild type (214). This suggests that the increased sensitivity to oxidative stress of prdx-2 worms is not responsible for their decreased lifespan.
D. Thioredoxin (trx-1 and trx-2) and thioredoxin reductase (trxr-1 and trxr-2)
Thioredoxins (TRXs) are small disulfide reductases that act by providing reducing equivalents. In addition to their role in reactivating peroxiredoxins, they have numerous other functions within the cell, including preventing cytosolic proteins from aggregating. TRXs are subsequently reactivated through reduction by thioredoxin reductase (TRXR) and NADPH. Analysis of the C. elegans genomic sequence revealed at least eight TRXs and two TRXRs (198), although, to date, only trx-1 and trx-2 have been studied. TRX-1 is found in the cytoplasm and along with TRXR-1 make up the cytoplasmic TRX system. trx-1 deletion mutants are slightly more sensitive to paraquat than wild-type worms and exhibit a 19% decrease in lifespan (136, 198). Similarly, overexpression of trx-1 results in a moderate increase in mean, but not maximum, lifespan (198). TRX-1 was found to be expressed in ASI and ASJ neurons as well as the intestine and expression increased with age (136).
trx-2 and trxr-2 make up the mitochondrial TRX system (Cacho-Valadaez, International Worm Meeting 2009). Against expectation, trx-2 and trx-2;trxr-2 mutants have increased resistance to paraquat. Also, the viability of trx-2;trxr-2;trxr-1 triple mutants suggests that the TRX system is not essential for survival. Although two TRXRs have been identified in C. elegans, there are currently no published reports describing their effect on lifespan or oxidative stress (33, 88, 163).
E. Glutaredoxin: glrx-5, glrx-10, glrx-21, and glrx-22
Glutaredoxin acts by providing reducing equivalents and is part of the glutathione system. The activity of glutaredoxin is restored by the oxidation of glutathione. Similarly, the active reduced form of glutathione is regenerated by glutathione reductase using NADPH as a donor of reducing equivalents. As with TRX, glutaredoxin can restore the activity of peroxiredoxin. Although four glutaredoxin genes appear on Wormbase (glrx-5, glrx-10, glrx-21, and glrx-22), there are no published reports describing these genes.
F. Glutathione S-transferase: gst
Glutathione S-transferases (GSTs) are a family of enzymes that catalyze the conjugation of glutathione (GSH) to a variety of substrates. These enzymes are involved in protecting against oxidative stress and phase II detoxification of other toxic stresses. Consistent with their role in protecting against oxidative stress, gst genes have been found to be upregulated after exposure to oxidative stress (259). C. elegans contains at least 44 gst genes and possibly as many as 57 (12, 174). gst genes have been found to be upregulated in long-lived daf-2 and clk-1 worms (47, 190) and may also contribute to the longevity of isp-1 worms since RNAi against the GST family member cdr-2 decreased their lifespan (47).
On the basis of the observation that gst-4 is markedly upregulated in worms exposed to paraquat (259) or hyperoxia (181), Leiers et al. postulated that gst-4 is involved in protection against oxidative damage. They found that expression from the gst-4 promoter was increased by exposure to paraquat, juglone, or plumbagin, but not heat or UV stress (174), suggesting that the gst-4 promoter responds specifically to oxidative stress. Consistent with the proposed role of gst-4 in defense against oxidative stress, overexpression of gst-4 increased resistance to juglone, whereas homozygous deletion resulted in increased juglone sensitivity (174). Despite exhibiting altered sensitivity to oxidative stress, both worms overexpressing gst-4 and gst-4 deletion mutants lived as long as wild-type worms (174). While the authors suggested that constitutive overexpression of gst-4 may lead to increased lifespan, it is clear that the loss of this gene does not limit normal longevity.
gst-10 catalyzes the detoxification of 4-HNE, a product of ROS-mediated lipid peroxidation that can bind to proteins and DNA causing cross-linking and damage. gst-10 acts by conjugating glutathione to 4-HNE and accounts for ¼–⅓ of the total 4-HNE conjugating activity in the worm (13). Ayyadevara et al. examined the effect of gst-10 on oxidative stress resistance and lifespan. Worms engineered to overexpress gst-10 exhibited a 36% increase in 4-HNE conjugating activity and were resistant to multiple forms of stress, including paraquat, H2O2, heat stress, and UV irradiation (13). This increase in resistance to stress was associated with a 22% increase in lifespan. Similarly, decreasing expression of gst-10 by RNAi resulted in increased sensitivity to 4-HNE, heat stress, and paraquat, as well as decreased lifespan (11). In support of a role for gst-10 in determining lifespan, it was found that increasing 4-HNE conjugating activity was associated with increased lifespan in a series of congenic lines (13). The protective properties of gst-10 also appear to contribute to the longevity of daf-2 mutants. daf-2 worms have increased expression of gst-10 that was shown to be correlated with lifespan among eight different daf-2 alleles. In addition, RNAi targeted against gst-10 was found to decrease the lifespan of daf-2 worms (11).
On the basis of these findings, Ayyadevara et al. surveyed an additional 26 gst genes by RNAi to determine if any possessed 4-HNE conjugating activity. Four additional genes (gst-5, gst-6, gst-8, and gst-24) were identified that possess 4-HNE conjugating activity (12). While the loss of any of the five gst genes resulted in increased damage caused by 4-HNE, only the loss of gst-5 or gst-10 resulted in decreased lifespan. This indicates that increased 4-HNE damage is not sufficient to decrease lifespan. One possibility, suggested by the authors, is that 4-HNE damage has a detrimental impact on lifespan only in specific tissues (12).
gsto-1 is another GST that has been shown to modulate stress resistance in C. elegans. On the basis of the upregulation of gsto-1 during oxidative stress, Burmeister et al. examined the role of gsto-1 in defense against oxidative stress. Worms that overexpress gsto-1 were found to have increased resistance to stress caused by juglone, paraquat, cumene, arsenite, and heat (34). However, the increased stress resistance did not lead to an increase in lifespan (34). Similarly, decreasing expression of gsto-1 by RNAi resulted in increased sensitivity to juglone, paraquat, cumene, arsenite, and heat, but did not affect lifespan (34). Thus, the levels of gsto-1 modulate sensitivity to a variety of stresses, including oxidative stress, but have no effect on lifespan.
While modulation of gst-10 expression altered resistance to oxidative stress and lifespan in a similar manner, altering expression of either gst-4, gst-6, gst-24, or gsto-1 affected oxidative stress resistance without impacting lifespan. This provides another example where resistance to oxidative stress can be modulated independently of lifespan.
G. Nicotinamide nucleotide transhydrogenase: nnt-1
Proton-translocating nicotinamide nucleotide transhydrogenase is found in the inner mitochondrial membrane and catalyzes the reduction of NADP+ by NADH. On the basis of the role of NADPH in providing reducing equivalents to the TRX and glutathione systems, Arkblad et al. examined the lifespan and oxidative stress resistance in nnt-1 worms, which lack nicotinamide nucleotide transhydrogenase (7). nnt-1 worms were shown to have markedly decreased levels of glutathione and showed increased sensitivity to paraquat during development. This suggests that nnt-1 worms have decreased antioxidant defenses and is consistent with NADPH having an important role in reactivating TRX and glutaredoxin. Despite their increased sensitivity to oxidative stress, nnt-1 worms appeared phenotypically normal and their lifespan was not different from wild type (7).
H. Glyoxylase
Glyoxylase is an enzyme that detoxifies methylglyoxal and glyoxal, two glycating agents that cause damage to protein (228). Increased levels of ROS production results in increased formation of methylglyoxal. Glyoxylase-1 activity was found to decrease with age despite constant mRNA levels with a corresponding increase in methylglyoxal (203). Glyoxylase-1 activity could also be reduced by exposure to H2O2, suggesting that ROS may be responsible for the age-dependent decline. Overexpression of glyoxylase-1 resulted in increased lifespan and decreased mitochondrial superoxide production (as measured by whole worm staining with dihydroethidium), whereas silencing glyoxylase-1 resulted in decreased lifespan and markedly increased levels of superoxide (203). These findings support an inverse relationship between superoxide levels and lifespan.
I. Summary
Examining the relationship between lifespan and ROS in mutants with decreased antioxidant defense reveals divergent results (Table 4). In some cases, such as prdx-2, trx-1, and gst-10, lifespan is found to be inversely related to sensitivity to oxidative stress. However, in other cases oxidative stress sensitivity is altered without affecting lifespan. Perhaps the strongest evidence against a role for ROS in causing aging comes from the examination of sod deletion mutants where even the deletion of multiple sod genes has little or no impact on lifespan despite clear increases in sensitivity to oxidative stress and, importantly, oxidative damage. It should be noted that some of these antioxidant defense genes have multiple roles within the cell (e.g., peroxiredoxin, TRX, and glutaredoxin), whereas for other genes the only known function is to detoxifying ROS (e.g., SOD and catalase). Thus, care is needed in interpreting results from the former as any effects on lifespan may not necessarily result from the gene's effect on oxidative stress resistance.
SOD, superoxide dismutase.
VIII. Effect of Exogenous Antioxidants on Lifespan
A number of studies have examined the effects on lifespan of compounds that influence ROS levels [reviewed in ref. (44)]. One of the limitations of this approach is the fact that worms take up drugs poorly, either through exclusion resulting from their thick cuticle, or by metabolizing the drugs. As a result, internal concentrations of compounds delivered to worms in the media are orders of magnitude less than the external concentration (51, 60). In addition, it is difficult or impossible to determine which cells or tissues are exposed to which concentration of drug. Finally, it is always important to consider that any exogenous compound may have effects aside from its intended activity and that these side effects might, in fact, be the cause of the phenotype observed (292). For example, any compound with a mildly toxic side effect could improve oxidative stress resistance and lifespan through hormesis.
A. SOD/catalase mimetics: Euk-8 and Euk-134
Euk-8 and Euk-134 are SOD/catalase mimetics that exhibit SOD activity in vitro and also show low levels of catalase activity. Initial reports found that Euk-8 and Euk-134 extend the lifespan of wild-type worms by up to 54% without affecting development or fertility (194). These compounds were also able to rescue the decreased lifespan of mev-1 worms (194). Both compounds increase resistance to paraquat during development and adulthood (235). Importantly, the protection is not acting through insulin/IGF signaling as the compounds also increased paraquat resistance of both daf-16 and daf-2 worms (235). The fact that an hsp-16:GFP reporter construct was not induced by treatment with Euk-8 or Euk-134 suggests that treatment with Euk-8/Euk-134 is not inducing a stress response.
Not all researchers have observed increased lifespan resulting from treatment with Euk-8 and Euk-134. In agreement with previous studies, Euk-8 and Euk-134 were found to increase SOD activity (up to fivefold) and to improve survival in the presence of oxidative stress induced by either paraquat or plumbagin (149). However, despite this increased resistance to oxidative stress, no increase in lifespan was observed (149). In fact, Euk-8 was found to decrease lifespan at higher doses (148). Thus, Euk-8 and Euk-134 exhibit clear antioxidant activity in vivo leading to increased resistance to oxidative stress, but the increased resistance to oxidative stress only appears to improve lifespan under certain conditions.
B. Platinum nanoparticles
Platinum nanoparticles (Pt-nano) also act as a SOD/catalase mimetic and have been shown to exhibit greater ROS-scavenging abilities than Euk-8 (155). Treatment of wild-type worms with Pt-nano increased survival when exposed to paraquat or H2O2 and was found to increase lifespan by 22% (154, 155). Treatment with Pt-nano also reduced the levels of the aging pigment lipofuscin and was able to increase the lifespan of mev-1 worms (155). A subsequent study has shown that more efficient delivery of Pt-nano can be achieved by linking these antioxidant particles to the TAT-derived peptide from HIV-1 (154). This study also demonstrated that treatment with Pt-nano was able to decrease ROS levels as measured by both DCF and MitoSOX (154). This treatment demonstrates a clear association between increased resistance to oxidative stress and increased lifespan.
C. N-acetyl cysteine
N-acetyl cysteine (NAC) is a precursor of glutathione that also acts as an antioxidant by scavenging ROS. Treatment with NAC was shown to decrease ROS production (239). Worms with mutations affecting mitochondrial complex II were shown to be sensitive to paraquat and hyperoxia, suggesting increased levels of ROS (124). Consistent with the observation that NAC reduces levels of ROS, treatment with NAC was shown to revert survival of the complex II mutants under hyperoxic conditions to wild type (124). Similarly, in the case of worms treated with 2-deoxyglucose, where the lifespan extension is dependent on increased ROS, treatment with NAC reverted the lifespan to wild type (239). In this experiment, NAC did not have an effect on the lifespan of wild-type worms (239). However, a subsequent report was successful in extending worm lifespan with NAC by delivering the NAC encapsulated in liposomes (245). The difference in these results likely stems from the fact that C. elegans is generally poor at taking up chemicals from solution or the nematode growth media. As with NAC, glutathione was also found to increase worm lifespan when delivered in liposomes but not when administered in the medium (245).
D. Blueberry extract
On the basis of the finding that blueberry extract was found to preserve neuronal and motor function in aging rats and its proposed antioxidant properties, Wilson et al. examined the effects of blueberry extract on worm lifespan. Treatment of worms with blueberry extract resulted in a 28% increase in mean lifespan that was accompanied by a slower decline in pharyngeal pumping rate, decreased intestinal lipofuscin accumulation, and decreased levels of 4-HNE (273). Examining the effects of different blueberry components on lifespan revealed that, while each of the components analyzed was shown to have antioxidant activity, only the proanthocyanidins fraction had an impact on lifespan (273). Despite the known antioxidant activities of the blueberry extract, treatment with blueberry extract failed to increase survival after exposure to either paraquat or H2O2, though it did improve thermotolerance. Combined with the fact that the blueberry extract did not increase the lifespan of mev-1 mutants, this suggests that it does not affect lifespan through its antioxidant properties (273). In addition, there was no evidence to support an increase in stress response genes after treatment with blueberry extract.
E. Coenzyme Q10
Coenzyme Q (CoQ), also known as ubiquinone, is involved in transferring electrons in the ETC but also has other functions within the cell. CoQ can affect ROS levels in two opposing ways: it can act as the electron donor for superoxide generation at mitochondrial complex III and it can act as an antioxidant. Consistent with its role as an antioxidant, CoQ10 was shown to increase the lifespan of wild-type worms by 18% and was also found to increase the lifespan of mev-1 worms (133). The fact that CoQ10 treatment resulted in reduced generation of superoxide supports a mechanism of lifespan extension mediated by decreasing oxidative stress (133). Congruent with the beneficial effects of CoQ10 in wild-type worms, examination of lifespan and oxidative damage in clk-1 worms grown on ubiquinone with different side chain lengths (ranging from 6 to 10 isoprenyl units) revealed that clk-1 worms fed bacteria producing CoQ10 exhibited the longest lifespan and least amount of oxidatively damaged proteins (284).
The absence of ubiquinone has also been examined for its effects on lifespan. It was found that feeding worms a diet that lacks ubiquinone results in a 60% increase in lifespan (170). While the increase in lifespan was originally attributed to the lack of ubiquinone and decreased ROS production, the authors subsequently demonstrated that feeding C. elegans respiratory-deficient E. coli that do synthesize ubiquinone also resulted in increased lifespan (234). This clearly indicated that it was not the absence of ubiquinone that increased lifespan in the worm.
F. Ginkgo biloba extract: EGb761
The Ginkgo biloba extract EGb761 is a popular health supplement that is hypothesized to delay cognitive decline. While the mechanism remains unknown, EGb761 has been proposed to increase stress resistance, including resistance to oxidative stress. On the basis of this fact, multiple groups have examined the effect of EGb761 on lifespan and stress resistance in C. elegans. The lifespan of worms treated with EGb761 has been shown to be increased by 8%–10% (143, 276). Further purification of individual components of EGb761 identified the flavonoid tamarixetin, which was able to increase lifespan by 25% (276). Treatment of worms with EGb761 also delayed sarcopenia and resulted in a corresponding increase in the period that the worms remained in a highly active state (36). Pharyngeal pumping was also maintained for a longer period in EGb761 treated worms (36).
In trying to ascertain the mechanism underlying the increased longevity, it was shown that EGb761 possesses antioxidant activity, although this activity was less efficient than the vitamin E derivative TROLOX (143). In stress assays, treatment with EGb761 was found to improve both thermotolerance and resistance to oxidative stress (143, 276). In line with this finding, EGb761 reduced the increase in expression of two different stress reporter genes, gst-4:GFP and hsp-16.2:GFP, after heat stress or exposure to juglone-mediated oxidative stress (143, 254). In addition, treatment with EGb761 was shown to decrease expression of catalase mRNA (143).
G. Epigallocatechin gallate
EGCG is the active extract of green tea and has been reported to protect against oxidative stress. Treating worms with EGCG resulted in decreased levels of ROS and increased resistance to juglone-induced oxidative stress (2, 31, 290). While EGCG was able to delay the age-dependent decline in mobility and pharyngeal pumping, it was initially found to have no effect on lifespan under normal conditions (31, 290). Two subsequent reports did, however, observe a 10% increase in the lifespan of EGCG-treated worms (2, 41). Treatment with EGCG resulted in increased expression of sod-3 and skn-1, indicating that this treatment resulted in upregulation of stress response genes that may have contributed to EGCG's impact on longevity (290).
H. α-lipoic acid
α-lipoic acid is a common dietary supplement with antioxidant activity. Accordingly, worms treated with α-lipoic acid exhibited decreased levels of H2O2 (31). While α-lipoic acid did not delay the decline in mobility or pharyngeal pumping, it was found to increase lifespan by up to 24% (31, 41). α-lipoic acid was also shown to improve an associative learning behavior, which was also improved in the long-lived clk-1 and isp-1 worms but deficient in short-lived mev-1 worms (210).
I. Vitamin C and vitamin E
Vitamin C (ascorbic acid) and vitamin E have both been shown to exhibit antioxidant activity. Treatment with vitamin C was able to decrease levels of H2O2 (254) and to prevent the upregulation of a hsp-16.2:GFP reporter in worms exposed to juglone (180). In addition, Vitamin C increased the survival of mitochondrial complex II mutants under hyperoxia (124). While Vitamin C supplementation of the growth medium was found not to increase worm lifespan (98), delivery of vitamin C in liposomes resulted in a significant increase in lifespan (245).
Vitamin E has also been observed to increase worm lifespan, though only at certain concentrations (98, 294). Interestingly, the beneficial effect of vitamin E with respect to lifespan appeared to occur only during larval development (98, 294). Vitamin E is composed of tocopherols and tocotrienols. Adachi and Ishii examined the effect of both components on lifespan, protein carbonyl content and survival under ultraviolet B (UVB) radiation-induced oxidative stress. On the basis of the fact that tocotrienols are much better antioxidants than tocopherols (216), they predicted that the tocotrienols would have more of an impact on lifespan and resistance to oxidative stress. They found that a mixture of tocotrienols was effective at increasing lifespan, decreasing carbonyls and increasing survival after exposure to UVB radiation, whereas tocopherol had little effect (4). In contrast, others have observed increased lifespan with both α-tocopherol (133) and γ-tocopherol (293). On the basis of the finding that vitamin E reduces brood size and delays reproduction, it has been suggested that vitamin E may be acting by inducing a mild stress leading to hormesis (98).
J. Herbal medicines
To study the benefits of two traditional herbal medicines: Huo Luo Xiao Ling Dan and Shi-Quan-Da-Bu-Tang (SQDB), Yu et al. examined the effects of these compounds on lifespan and oxidative stress resistance. It was found that treatment with SQDB resulted in a modest increase in lifespan, which appeared to result primarily from 2 of its 11 components, Cinnamomum cassia bark (CIN) and Panax ginseng root (GIN), both of which could increase worm lifespan (288). The increase in lifespan resulting from treatment with SQDB, CIN, or GIN was associated with a decrease in ROS levels as measured by DCF. These treatments also resulted in increased hsp-16.2 reporter expression following heat shock, suggesting a more robust heat shock response.
K. Predictive ability of in vitro antioxidant capacity on in vivo antioxidant capacity and lifespan
Pun et al. examined a series of antioxidants to determine whether the effect of an antioxidant in vivo in worms could be predicted by in vitro assays of oxidant scavenging ability. In addition, they sought to determine whether either in vitro or in vivo antioxidant capacity could predict an effect on lifespan. They found that in vivo antioxidant efficacy, as measured by an antioxidant's effect on the levels of protein carbonyls, was not predicted by in vitro performance (226). Further, while all six of the antioxidant compounds tested were able to increase resistance to paraquat-mediated oxidative stress, only two increased lifespan. In both cases the increase in lifespan was modest and did not appear to be related to the compounds effect on oxidative damage, as one compound increased protein carbonyl levels while the other decreased protein carbonyls (226). This study suggests that while antioxidants can increase lifespan, their ability to increase resistance to oxidative stress does not seem sufficient to explain their effects on lifespan. Alternatively, it is possible that the antioxidants that failed to extend lifespan had detrimental effects that masked any beneficial effect of decreasing oxidative stress on lifespan. In addition, this study demonstrates that increased oxidative damage is compatible with long life.
L. Summary
The treatment of worms with exogenous antioxidants results in increased resistance to oxidative stress. While there was some variability in the effect of these antioxidants on lifespan based on dose, experimental conditions, and method of delivery, most of the antioxidants described above were shown to increase lifespan under at least one set of conditions (Table 5). However, in some cases, the effect of the antioxidants on oxidative stress resistance could be dissociated from their effect on lifespan as oxidative stress resistance was observed even when an effect on lifespan was not. In an experiment specifically designed to compare an antioxidants ability to protect against oxidative damage and lifespan, no clear relationship was observed.
NGM, nematode growth medium.
IX. Environmental Effects on ROS and Lifespan
The logic of evolutionary biology dictates that the lifespan of an organism must be dependent on both its genetics and its environment. By using a model organism, such as C. elegans, it is possible to exclusively test the impact of environment on lifespan, since all wild-type worms should be genetically identical.
A. Temperature
Excluding extreme heat or cold temperatures at which worms are unable to survive, there is an inverse relationship between growth temperature and lifespan in worms (116, 159, 164). For example, worms grown at 10°C, 15°C, 20°C, and 25°C exhibited mean lifespans of ∼43, 32, 18, and 11 days, respectively (265). On the basis of the finding that metabolic rate, as measured by carbon dioxide production, increases with temperature over this same range (265), these authors proposed that the underlying mechanism may be an alteration in the rate of ROS production. That is, decreasing temperature results in decreased metabolic rate, which results in less production of ROS, less oxidative damage, and longer life. However, it is quite possible that metabolic alterations extend lifespan through an ROS-independent mechanism. Support for this idea comes from the fact that growing worms at a cold temperature of 16°C increased lifespan but also leads to increased sensitivity to paraquat-induced oxidative stress (286). In addition, it appears that lifespan is at least to some extent regulated by thermosensory neurons, as lifespan at 25°C is further decreased by loss of thermosensory neurons (171). Thus, while modulation of temperature reveals an inverse relationship between lifespan and metabolic rate, it is uncertain if the underlying mechanism is a change in the rate of ROS production.
B. Oxygen concentration
Alterations in oxygen concentration also impact worm lifespan. It has long been known that increased oxygen concentration is toxic to C. elegans (22). Since the production of ROS is believed to be proportional to oxygen concentration, it is thought that the toxic effects of high oxygen concentration results from high levels of ROS (72). In support of this idea, it has been shown that the toxicity of high oxygen concentrations is exacerbated by inhibition of SOD activity, thereby suggesting that the increase in oxygen concentration results in increased production of superoxide to a degree that exceeds the worms antioxidant defense mechanisms leading to toxicity (22). In addition, growing worms under hyperoxic conditions has been shown to increase protein carbonylation (3) and the rate of mutation (99).
Honda et al. examined the survival of worms under different concentrations of oxygen and found that oxygen concentrations of 60% or greater resulted in a dose-dependent decrease in worm lifespan [normoxia is 21% oxygen (111)]. Similarly, under hypoxic conditions of 1% oxygen, worms exhibited increased lifespan, whereas lifespan was relatively constant over the range of 2% to 40% oxygen (110). Interestingly, if worms exposed to hyperoxia were returned to normoxia, the decrease in lifespan that they experienced could be reduced, suggesting the possibility that the oxygen-induced damage can be repaired (111).
Adachi et al. also observed decreased lifespan at >60% oxygen and increased lifespan at 1% oxygen and extended these findings to two additional strains: long-lived age-1 worms and short-lived mev-1 worms (3). Interestingly, the ability to adapt to hyperoxia appears to be lost with age. While young worms upregulate SOD activity in response to hyperoxia, older worms fail to increase antioxidant defenses and exhibit decreased survival (50). Overall, extreme concentrations of oxygen are able to increase (1%) or decrease (>60%) lifespan, while lifespan is relatively constant over the range of oxygen concentrations surrounding normoxia (2%–40%).
C. Axenic medium
While C. elegans is normally grown with bacteria as a food source, some researchers have tried to develop axenic medium (medium that does not contain living organisms), which would provide a more reproducible environment for experiments. It should be noted that at least part of the effects of axenic medium may stem from the fact that this type of medium is normally liquid. In a liquid medium, even with bacteria as a food source, worms can exhibit poor growth and increased lifespan (48, 159). Similarly, worms grow poorly in an axenic medium but exhibit increased lifespan (120). On the basis of their slow development and decreased fertility in the axenic medium, it was proposed that these worms experience severe dietary restriction, which leads to their increased lifespan (120).
Examination of oxidative stress resistance revealed that worms grown in the axenic medium are mildly resistant to paraquat and have increased SOD and catalase activity (118, 120). The effects of the axenic medium do not appear to be mediated through the insulin/IGF-1 signaling pathway, as similar improvements in lifespan and antioxidant activity were observed in daf-16 and daf-2 worms (118). In addition, it was found that a daf-16:GFP reporter construct does not translocate to the nucleus under axenic conditions (118). In looking for possible metabolic changes induced by growth in the axenic medium that could affect lifespan, it was found that growth in the axenic medium did not decrease oxygen consumption but did result in decreased levels of ATP at young ages in these mutants (at later ages ATP levels were greater than in the wild-type strain) (120). Finally, examination of the changes in mRNA expression that result from growth in the axenic medium revealed no evidence for a decreased metabolic rate or shift to anaerobic metabolism in worms grown in the axenic medium (note that metabolism was not directly measured in this study) (38).
D. Summary
Both lifespan and ROS production can be influenced by the environment. Increasing temperature or oxygen concentration are thought to decrease lifespan through an increase in the levels of ROS. However, it will be important to measure ROS levels and oxidative damage under these conditions to support these conclusions more firmly. In the case of altered temperature, the effects of temperature on lifespan could be dissociated from its effects on sensitivity to oxidative stress as worms grown at low temperature were long lived and sensitive to oxidative stress (286). Worms grown in the axenic medium have increased antioxidant defenses and increased resistance to oxidative stress, which may contribute to their long lifespan.
X. Functional Roles for ROS
While ROS are known to cause damage to macromolecules, the reactive nature of these molecules can also be harnessed to perform a particular function such as pathogen resistance. In addition, many forms of ROS have been shown to function in intracellular signaling pathways.
A. A role for ROS in combating pathogens
When C. elegans are exposed to different types of pathogenic bacteria, it was shown that the levels of ROS, oxidatively damaged protein (as measured by lipofuscin), and expression of the antioxidant defense enzyme SOD-3 are all increased (41). The authors propose that the toxicity of the ROS generated to kill the pathogen contributes to the pathogen-induced mortality. This is supported by the fact that sod-3 and ctl-2 mutant worms are more sensitive to pathogens than wild-type worms (ctl-1 has normal sensitivity) and that the addition of exogenous catalase protects worms from pathogen-induced mortality (41).
In a subsequent study, it was found that the dual oxidase (NADPH oxidase and peroxidase) Ce-Duox1/BLI-3 appears to be responsible for ROS production during infection (40). RNAi knockdown of this gene increases susceptibility to infection while decreasing the production of H2O2 in response to pathogen exposure (40). Mutations affecting the peroxidase domain did not affect survival after exposure to pathogen, suggesting that the NADPH oxidase domain is involved in producing ROS. Interestingly, treatment with the antioxidants α-lipoic acid or EGCG extended the lifespan of worms but decreased survival after pathogen exposure, indicating the importance of ROS in fighting off infection (40). Jain et al. also demonstrated an increase in ROS in C. elegans after infection with yeast and were able to associate the ROS generation with the NADPH oxidase activity of bli-3 using a specific inhibitor of NADPH oxidase activity (Diphenyleneiodonium) (135).
B. A role for ROS in intracellular signaling
While much of the research on ROS has focused on its harmful effects, ROS has also been demonstrated to have a significant role in intracellular signaling. ROS can affect signaling either through the modification of kinases, phosphatases, and transcription factors or by directly inducing transcription at specific promoters (46). Despite a growing body of research demonstrating a wide variety of roles for ROS signaling in other organisms, little is known about ROS signaling in C. elegans. Nonetheless, ROS signaling has been found to interact with ras signaling in vulva formation (246).
let-60 encodes ras in C. elegans and a let-60 gain of function allele results in the formation of multiple vulva. This phenotype is suppressed by deletion of clk-1 in clk-1;let-60 double mutants. Decreasing the levels of sod-1 in clk-1;let-60 double mutants by RNAi resulted in an increased frequency of the multivulva phenotype (246). As ras signaling in cell models has been shown to be redox sensitive (127), this suggests that reduced ROS signaling in clk-1 worms suppresses the multivulva phenotype of let-60(gf) worms and that this signaling is restored by knocking down sod-1. This conclusion is supported by the observation of increased levels of SOD-1 in clk-1 worms (264) (Van Raamsdonk and Hekimi, unpublished results).
XI. Executive Summary and Conclusions
C. elegans provides an ideal organism for testing the rate-of-living and the free radical theory of aging, as lifespan experiments can be completed in a matter of weeks and a wide variety of genetic tools are available. Accordingly, the role of oxidative stress in aging in C. elegans has been extensively tested and frequently reviewed (27, 32, 81, 89, 113, 130, 176, 188, 240, 250). As additional data have become available, there appears to be a general trend toward questioning the validity of the free radical theory of aging [e.g., refs. (82, 168, 221)] and there have been suggestions that the focus should be expanded to include other forms of stress (83, 196).
In this review, we have examined the relationship between ROS and aging in long-lived mutants, mutants with altered mitochondrial function, mutants with decreased antioxidant defense, worms treated with antioxidant compounds, and worms exposed to different environmental conditions (summarized in Fig. 14). While there are many examples that are consistent with the proposal that the accumulation of ROS-mediated damage is a major contributor to aging, there is also a great deal of evidence that demonstrates that changes in oxidative damage can be dissociated from the effects on aging. This is demonstrated by the fact that long-lived worms can have either increased, normal, or decreased sensitivity to oxidative stress (Fig. 14).
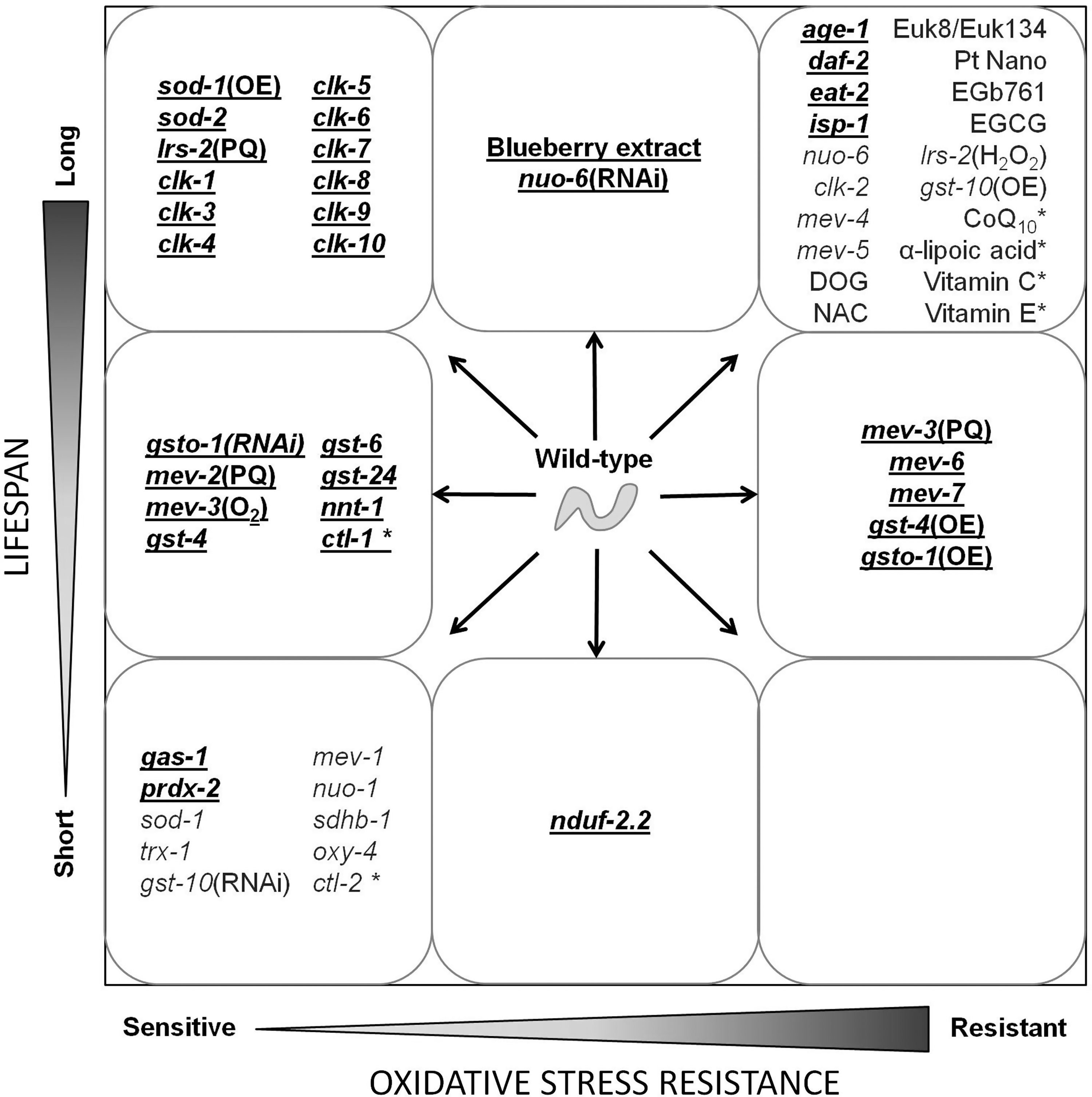
On the basis of the work summarized in this review, it appears that increased resistance to oxidative stress is neither sufficient nor necessary for increased lifespan. Similarly, increased sensitivity to oxidative stress can lead to decreased lifespan but is not incompatible with long life. Despite the fact that exposure to high concentrations of ROS is toxic, the amounts of ROS generated during normal metabolism may not be sufficient to influence aging. Thus, while oxidative damage clearly increases with age (4, 12, 176), experimentally increasing oxidative damage does not necessarily shorten lifespan (54, 263, 283). Oxidative stress may contribute to aging under certain conditions and may be one of multiple factors that influence lifespan. However, oxidative damage does not appear to be the primary cause of aging.
XII. Future Directions
In reviewing the current literature, there are many areas of research that could serve to further define the relationship between the cellular redox state and aging.
A. Develop new tools for measuring ROS
Perhaps the greatest limitation of research in this area to date is the ability to accurately measure ROS generation, ROS levels, and oxidative damage. The experimental tools currently available are inadequate and have the potential to lead to spurious conclusions. The development of new measurement techniques will greatly aid research in this area.
B. Consideration of the role of ROS in signaling
It has been shown in other organisms that ROS has multiple roles in intracellular signaling. As altered ROS signaling may also affect the lifespan of the worm, it will be informative to examine ROS production and removal within the context of signaling.
C. Reactive nitrogen species
Most studies in C. elegans have focused on ROS while few have examined the contribution of reactive nitrogen species (RNS) and nitrosative damage to aging. While the levels of RNS might be minimal in C. elegans due to the absence of nitric oxide synthase (206), RNS generation and the possible contribution of RNS to lifespan has not been explored.
D. Combinatorial mechanisms of aging
While much of the work described in this review has focused on the effect of a single gene or treatment on aging, it is likely that multiple factors influence lifespan. As it is possible that many antioxidant or other defense enzymes each make a small contribution to lifespan, it will be informative to examine worms with mutations affecting multiple defense systems. This could include multiple antioxidant defense systems, such as SOD and peroxiredoxins, or defenses against different forms of stress, such as oxidative stress and heat stress.
E. Cell specificity
The potential for a gene to affect lifespan can be influenced by when the gene is expressed, in which cells it is expressed and where in the cell the product is localized. While the subcellular location of a gene product is generally examined, few studies have examined tissue specificity.
F. Measurement of oxidative damage
Many studies do not report the levels of oxidative damage, and for those that do, in most cases, only oxidative damage to protein was examined. As oxidative damage to DNA has been proposed as a major contributor to aging (80, 186), it will be informative to examine this outcomes in future experiments. It will also be useful to quantify oxidative damage to lipids as lipids have vital roles in membrane biology and signaling.
G. Repair of oxidative damage
While much of the work described in this review has been focused on oxidative damage, much less is known about repair mechanisms. As the accumulation of damage is the result of the balance between the rate of damage and the rate of repair, it will be important to determine what mechanisms are involved in repairing oxidative damage and how they are controlled. While some work has been done on GSTs (13), even within this class of enzyme the genes are largely unexplored [there are 57 predicted GSTs in C. elegans (12)].
Footnotes
Acknowledgments
J.V.R. is supported by the Canadian Institutes of Health Research, the Hereditary Disease Foundation, and Parkinson Society Canada. S.H. is supported by grants from the Canadian Institutes of Health Research (#216377, #216376, and #218649), the Canadian Cancer Society (#89761), and McGill University. S.H. is Campbell Chair of Developmental Biology and Strathcona Chair of Zoology.
Abbreviations Used
Reviewing Editors: Marni J. Falk, Siu Sylvia Lee, Young-Ki Paik, Margaret Sedensky, Federico Sesti, Robert J. Shmookler-Reis, Elizabeth A. Veal, Natascia Ventura, and Qiu-Mei Zhang-Akiyama