
Abstract
Introduction
Changes in cellular redox status are a prominent feature of immune responses in higher eukaryotes. In this context, a hallmark of plant defense activation is the rapid production of reactive oxygen intermediates (ROIs), known as the oxidative burst and is primarily characterized by the production of superoxide (O2 −) and hydrogen peroxide (H2O2). In addition, nitric oxide (NO) levels are also dramatically increased during the HR, constituting a parallel nitrosative burst. ROIs and NO, either alone or in combination, have been proposed to function as key orchestrators of the different branches of plant immunity. Here, we review the most recent findings in both the synthesis and signaling of ROIs and NO, and their molecular roles in regulating plant immunity. Further, we discuss downstream signaling by NO focusing on the post-translational regulation of defense-related proteins.
Production of ROIs by NADPH Oxidases
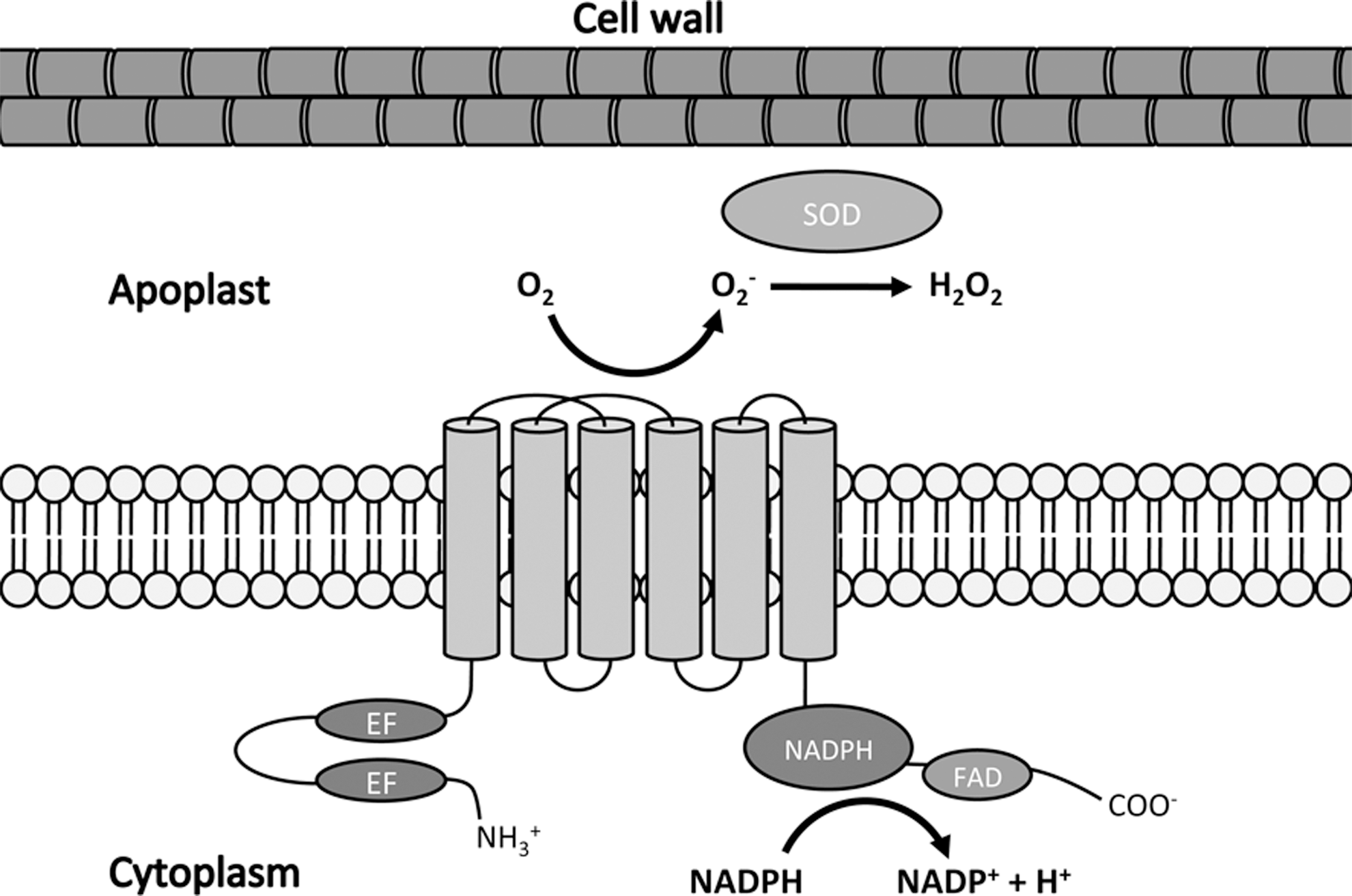
The production of apoplastic ROIs by plants after pathogen recognition has long been attributed to nicotinamide adenine dinucleotide phosphate (NADPH) oxidase enzymes related to those that perform the same role in mammalian phagocytes, termed respiratory burst oxidase homologues (RBOHs). The first report of a plant RBOH revealed that unlike its mammalian homologue gp91phox, it possesses additional potential regulatory EF-hand domains within an N-terminal extension (20). Like mammalian NADPH oxidases, plant RBOHs are transmembrane proteins that transfer electrons from intracellular NADPH across the plasma membrane before they are coupled to molecular oxygen in the apoplast to produce O2 − and its subsequent dismutation product, H2O2 (Fig. 1). This dismutation reaction can ensue spontaneously but can be enhanced by the activity of superoxide dismutase. NADPH and the cofactor flavin adenine dinucleotide (FAD) both bind at regions within the C-terminal. The Arabidopsis genome contains ten predicted RBOH genes (RBOHA-RBOHJ) (42) and through genetic studies using loss-of-function mutants, RBOHD and RBOHF were shown to be the principle generators of ROIs after recognition of PAMPs and avirulent pathogens (43).
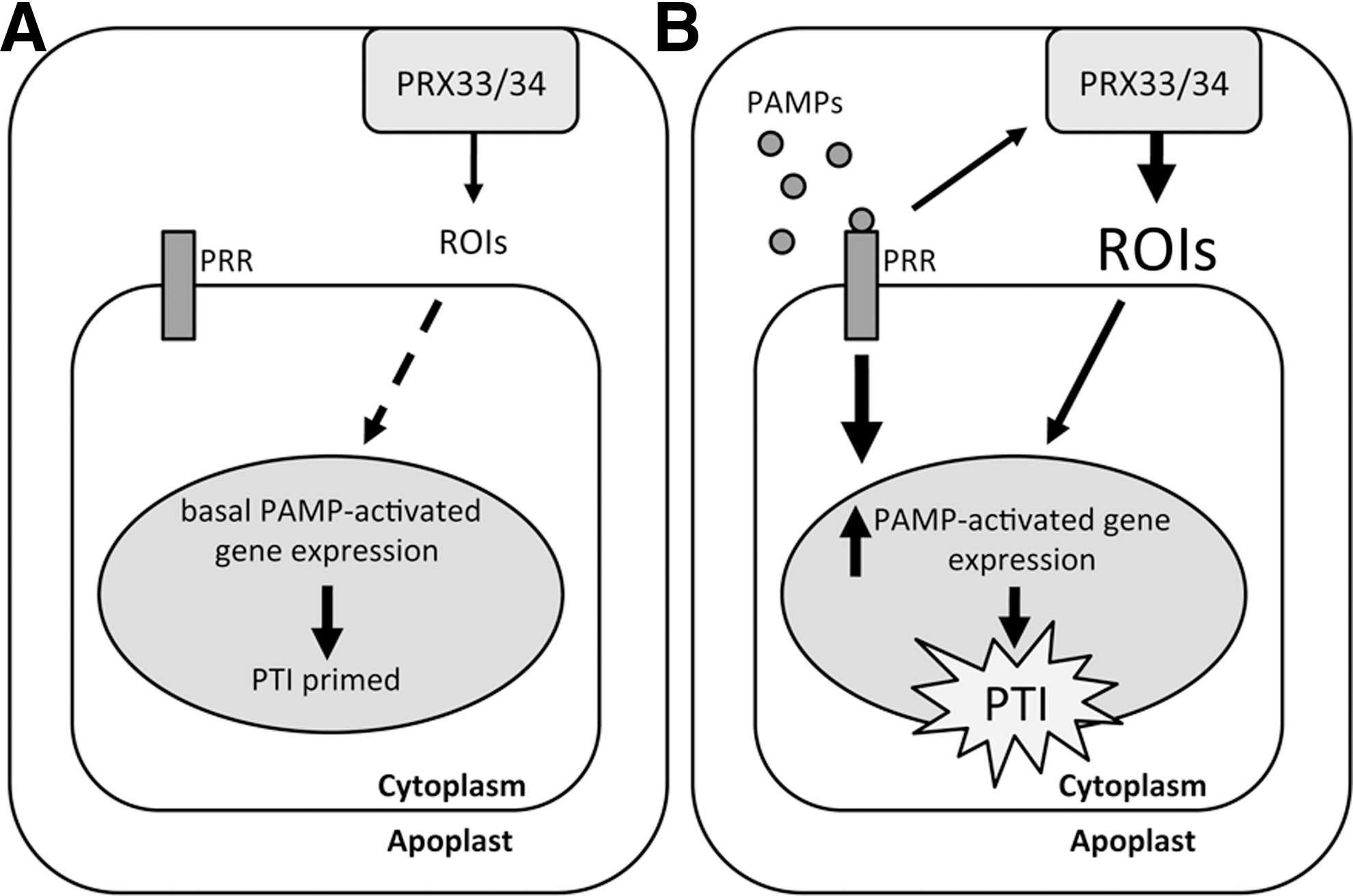
Since plants possess so many potential RBOH genes, this begs the question of whether these are functionally redundant or if there is in fact specificity for each RBOH to different cellular cues or requirements? The answer to this question remains largely enigmatic although recent work has begun to unravel these complex signaling networks. There is indeed evidence that RBOH proteins at least in part share redundancy as some phenotypes of atrbohD and atrbohF mutant plants, including reduced HR, are enhanced in the atrbohD atrbohF double mutant (43). Further study of these mutants surprisingly revealed that both RBOHD and RBOHF may actually suppress the spread of cell death during the HR in a lesion mimic mutant (lsd1) that exhibits “runaway” cell death in infected leaves (44). Most work on these enzymes has focused on their role in generation of ROIs after pathogen recognition and their effect on cell death during the HR. However, a recent study (7) investigated the effect of atrbohD and atrbohF mutations on defense-associated metabolic responses. The authors reported that both mutant lines showed similar resistance to Pseudomonas syringae pv. tomato DC3000 carrying the avirulence gene avrRpm1 but only atrbohF plants were more susceptible to the virulent form of the same pathogen. The Arabidopsis catalase 2 (cat2) mutant has increased intracellular H2O2 levels, which causes constitutive HR-like cell death and defense signaling when grown under long days in air (8), and these plants were used as a tool to uncover the effects of atrboh mutations on intracellular oxidative stress. This work revealed that RBOHF, but not RBOHD, was required to maintain various downstream responses emanating from high intracellular ROI levels in the cat2 mutant (7). These responses included the accumulation of salicylic acid (SA), an important signaling molecule essential for the establishment of disease resistance. Thus, although RBOHD and RBOHF have overlapping functions, there appears to be specificity in some instances. The complexity of these signaling networks makes it likely that the same set of proteins can induce different responses depending on the context of their requirement and multiple layers of regulation must exist for plants to fine-tune these both temporally and spatially.
Alternative Sources of ROIs
ROI synthesis by plants after pathogen recognition has also been attributed to a number of alternative mechanisms, including the activity of peroxidase enzymes. Transgenic Arabidopsis plants expressing an antisense cDNA against a French bean class III peroxidase were compromised in resistance against fungal and bacterial pathogens and showed a decreased oxidative burst, which was attributed to the silencing of two Arabidopsis peroxidase genes, PRX33 and PRX34 (4). A more recent study from the same laboratory (12) demonstrated that PRX33 and PRX34 knockdown Arabidopsis plants exhibit a reduced oxidative burst in response to PAMPs, including Flg22 and Elf26, synthetic peptides based on bacterial flagellin and elongation factor Tu, both well-established PAMPs recognized by plants. Further, these knockdown lines were impaired in the induction of several PAMP-activated genes after Flg22 and Elf26 treatment. Interestingly, these genes were also downregulated before PAMP treatment in the PRX knockdown lines compared to wild-type plants, suggesting that basal ROI production by PRX33 and/or PRX34 may constitutively prime plant defenses for attempted pathogen attack (12) (Fig. 2). Further evidence for peroxidases as a source of pathogen-induced ROIs emerged from a recent study in the moss Physcomitrella patens, where it was shown that Prx34 not only catalyzes ROI production but is also required for antifungal resistance (23). It must be noted that this peroxidase is unrelated to the previously mentioned Arabidopsis PRX33/34 (24) and the same gene nomenclature is purely coincidental.
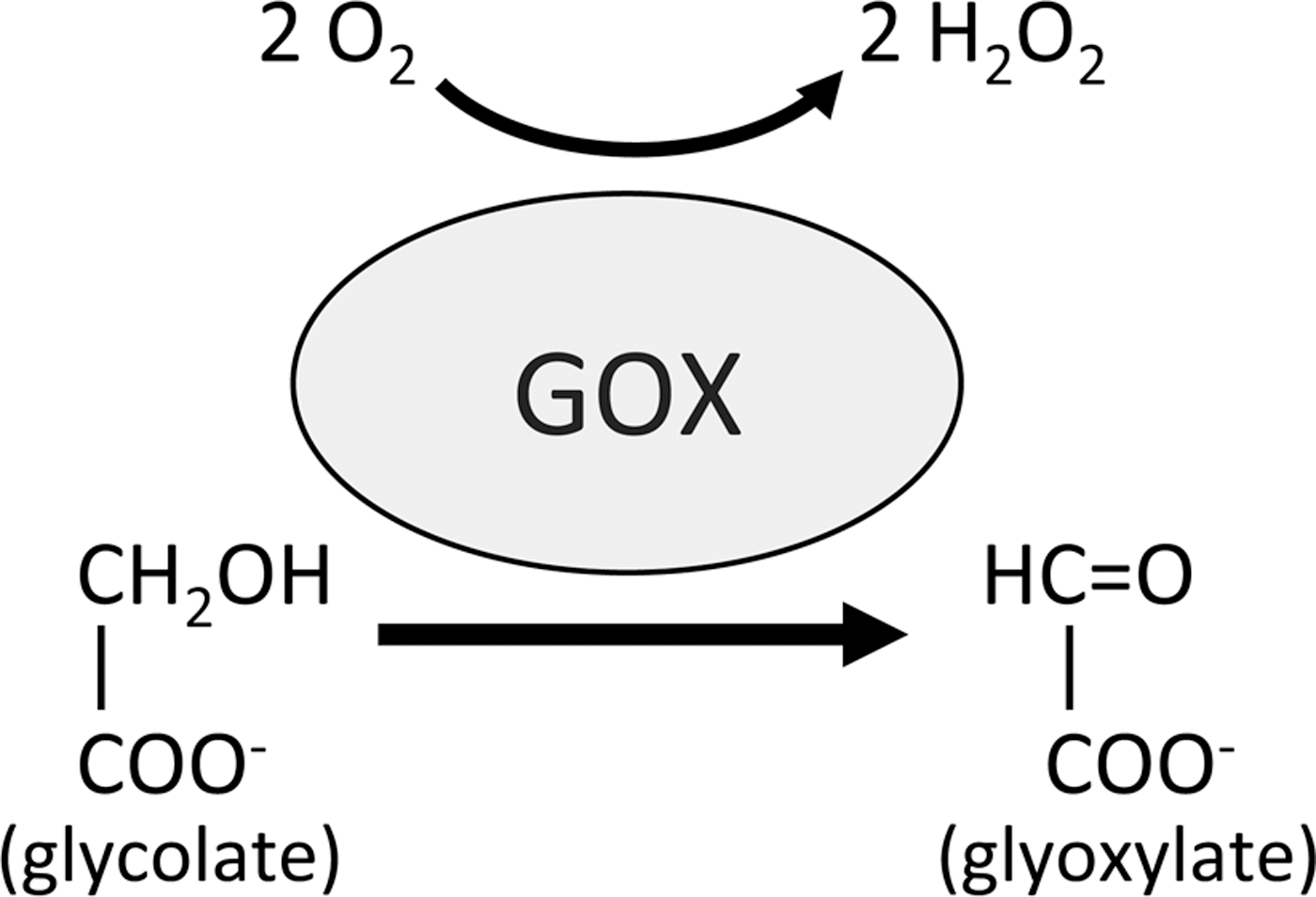
Another potential source of ROIs in plants is through the activity of glycolate oxidases (GOX). These enzymes are located in peroxisomes and catalyze the conversion of glycolate into glyoxylate during photorespiration, producing H2O2 in the process (Fig. 3). GOX has been proposed to be involved in disease resistance by various studies (5, 35, 41) although until recently its role has remained unclear. A recent study identified GOX in a screen for genes involved in non-host resistance (NHR) signaling in Nicotiana benthamiana (31). This work revealed that both GOX-silenced N. benthamiana and Arabidopsis gox mutant plants exhibit decreased NHR and gene-for-gene resistance. Further, these plants accumulated less H2O2 independent of NADPH oxidase activity and silencing RBOHD in gox plants did not affect their susceptibility to non-host pathogens. Thus, this study suggests that GOX is an important alternative source of ROIs during both NHR- and R-gene-mediated defense responses.
Production of NO
Despite accumulating evidence implicating a central role for NO in plant growth, development, and environmental responses, a nitric oxide synthase (NOS), structurally similar to those found in animals, has not been uncovered in higher plants. These NADP(H)-dependent enzymes catalyze the formation of NO and citrulline from
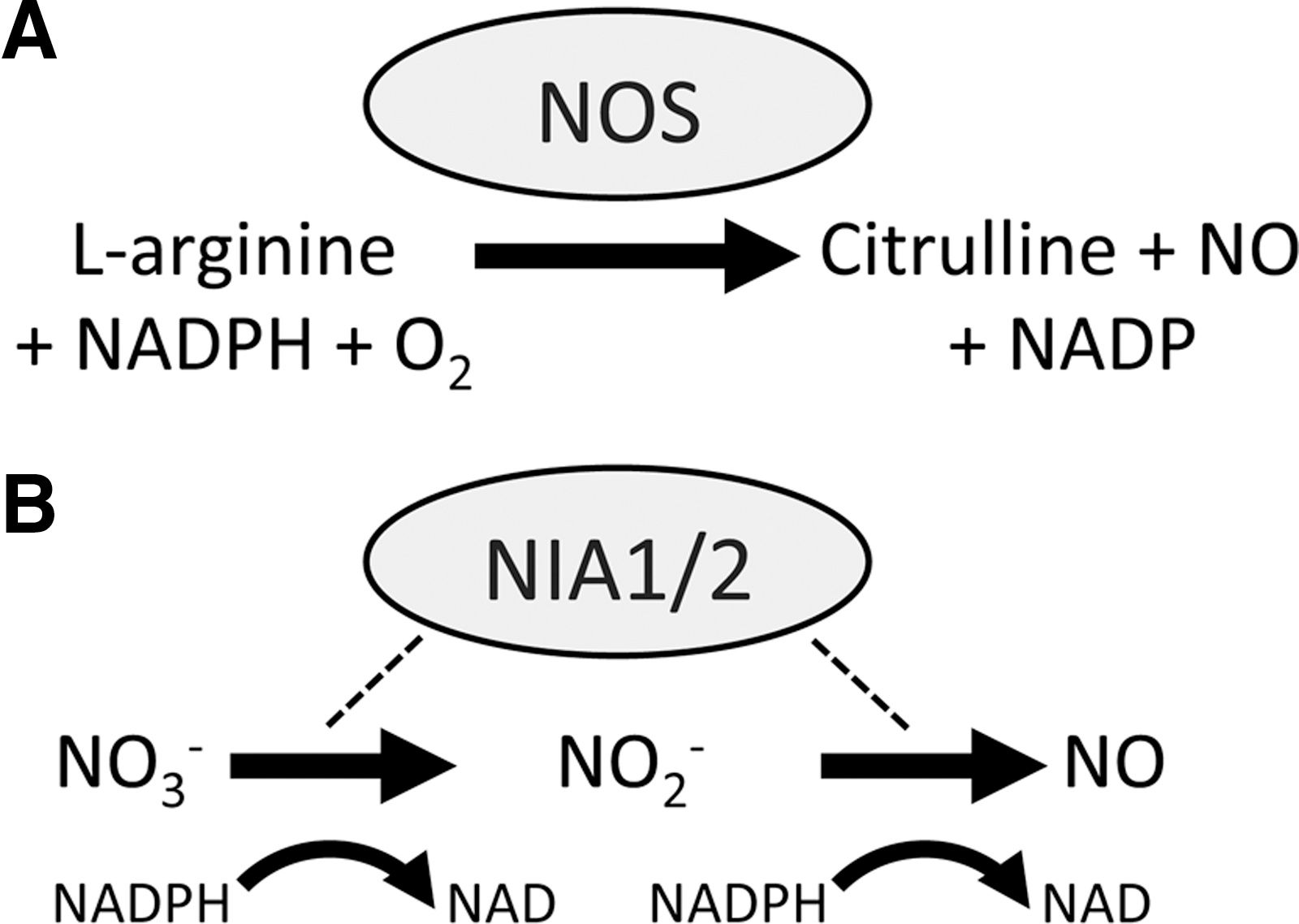
While the hunt for an NOS in higher plants continues, other mechanisms for NO synthesis after pathogen recognition have been explored. A possible source of NO is through the action of cytosolic nitrate reductase (NR). The Arabidopsis genome encodes two of these genes, named NIA1 and NIA2, with NIA2 being responsible for the bulk of NR activity (47). These enzymes are involved in reducing nitrate, the main nitrogen source of plants, to nitrite (NO2 −) in a NADPH-dependent fashion (Fig. 4B). However, NR appears to also be capable of producing NO in vitro and in vivo and this was unaffected by NOS inhibitors (30). The in vivo data from this study did rely on artificial chemical stimulation of NR and so may not be physiologically relevant. Further, the production of NO by NR is extremely inefficient and depends upon low O2 levels and high concentrations of NO2 − (30). In the context of plant immunity, Arabidopsis nia1 and nia2 mutant plants, and wild-type plants treated with NR inhibitors might be impaired in NO production after recognition of toxins from the fungal pathogen Verticillium dahliae (36).
S-Nitrosoglutathione-Reductase and S-Nitrosylation
It is well established that NO levels increase in plants after pathogen infection but how does this small molecule convey its biochemical activity in plant cells? An emerging redox-based post-translational modification is S-nitrosylation, by which an NO moiety is coupled to a reactive cysteine thiol, forming an S-nitrosothiol (SNO). This form of protein modification was discovered in 1992 (38) and is analogous to the addition of a phosphate group during protein phosphorylation (45). In addition to being implicated in many cellular processes in animals (18), recent evidence has shown that S-nitrosylation is involved in the regulation of various important plant processes including plant defense-related gene expression and hypersensitive cell death (45).
Glutathione (GSH) is an antioxidant tripeptide that can be S-nitrosylated to form S-nitrosoglutathione (GSNO), a molecule thought to function as a reservoir of NO bioactivity. NO can be released from GSNO or can be directly transferred to a target by a process known as transnitrosylation (18). The enzyme responsible for controlling GSNO levels is known as GSNO-reductase (GSNOR) and reduces GSNO to oxidized GSH and NH3. While this enzyme is highly specific for GSNO, it indirectly controls the levels of protein SNOs, by turning over this natural NO donor (27). In Arabidopsis, loss of GSNOR function in the mutant line gsnor1-3 increases SNO levels and compromises R gene-mediated defense and NHR, while a gain-of-function mutant line gsnor1-1 showed enhanced resistance to virulent pathogens (15). This study provided evidence that SNO metabolism controlled by GSNOR had a key role in plant disease resistance. Further studies have since revealed that GSNOR appears to be a key regulator of cell death (9) and is also required for thermotolerance (22). Various studies discussed in the following section have also shown that S-nitrosylation of specific proteins is increased in gsnor1-3 mutant plants.
S-Nitrosylation of Plant Defense-Related Proteins
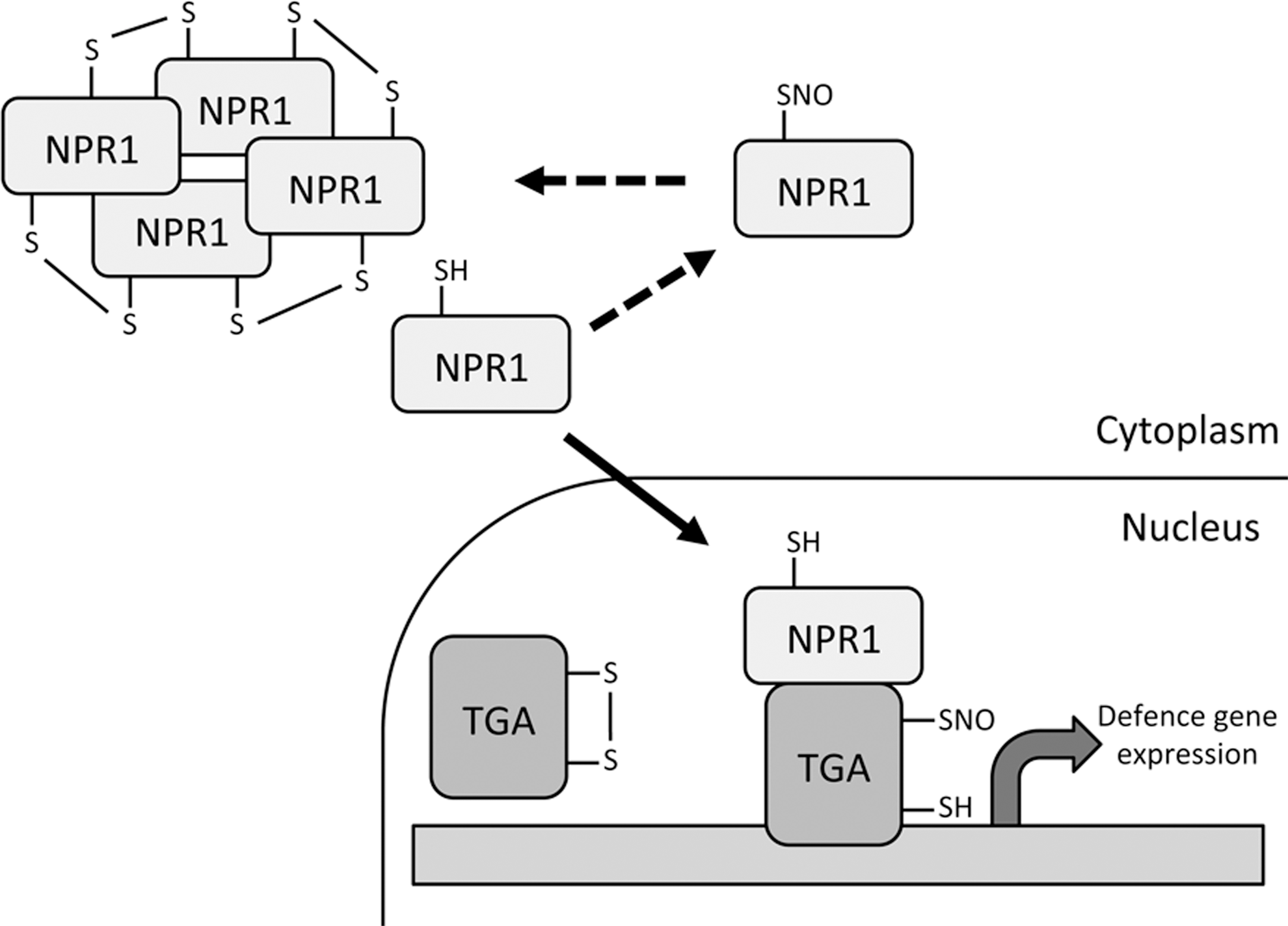
The defense-related transcription co-activator nonexpressor of pathogenesis-related genes 1 (NPR1) possesses many cysteine residues that form intermolecular disulfide bonds forming a high molecular-weight oligomer that is localized in the cytoplasm. After pathogen infection, these disulfide bonds are reduced allowing monomeric NPR1 to translocate to the nucleus where it exerts its co-activator function on many target genes (29). The complexity of this process was further revealed in a subsequent study where the authors showed that S-nitrosylation of NPR1 promoted its oligomerization and thus blunted its transcriptional activity (40). S-nitrosylation of NPR1 was increased in the gsnor1-3 mutant thus providing a link between high global SNO levels and the compromised disease resistance of this line. Cellular redox status not only affects NPR1 but also its binding partners TGA1 and TGA4. These transcription factors have been shown to contain intramolecular disulfide bridges at key cysteine residues that prevent NPR1 interaction (13). The authors of this study showed that reduction of these disulfide bonds allows NPR1-TGA1/4 interaction and NPR1-stimulated DNA binding of these transcription factors. A recent study revealed that these same cysteine residues can be S-nitrosylated and this may block further oxidative modification of TGA1/4, enhancing their DNA-binding capacity (26). There is now a wealth of evidence suggesting the NPR1-TGA transcriptional regulation system is under complex redox control with S-nitrosylation appearing to play a key role (37) (Fig. 5).
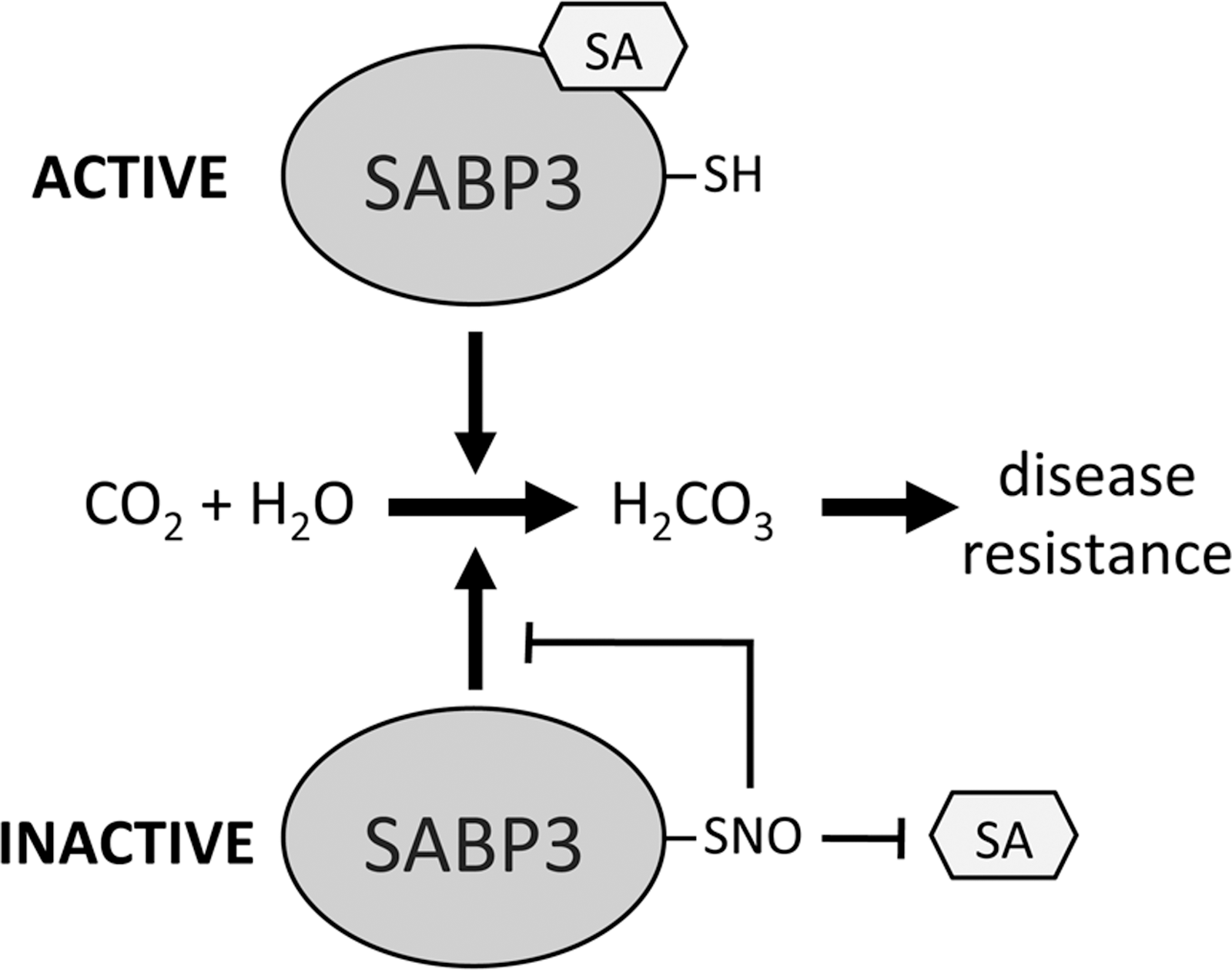
SA-binding protein 3 (SABP3) is a positive regulator of plant immunity that was identified as a target for S-nitrosylation (46). Similar to NPR1, S-nitrosylation of SABP3 is increased in the gsnor1-3 line. SABP3 possesses carbonic anhydrase activity that is required for expression of resistance in plants and this was shown to be inhibited by S-nitrosylation (Fig. 6). This raises a dilemma since a nitrosative burst and subsequent S-nitrosylation of proteins is associated with activation of plant defense responses, yet, S-nitrosylation of SABP3 inhibits its defensive activity. Interestingly, this could represent a negative feedback loop in plant defense signaling to regulate SA-mediated responses or could be a strategy employed by pathogens to suppress host immunity.
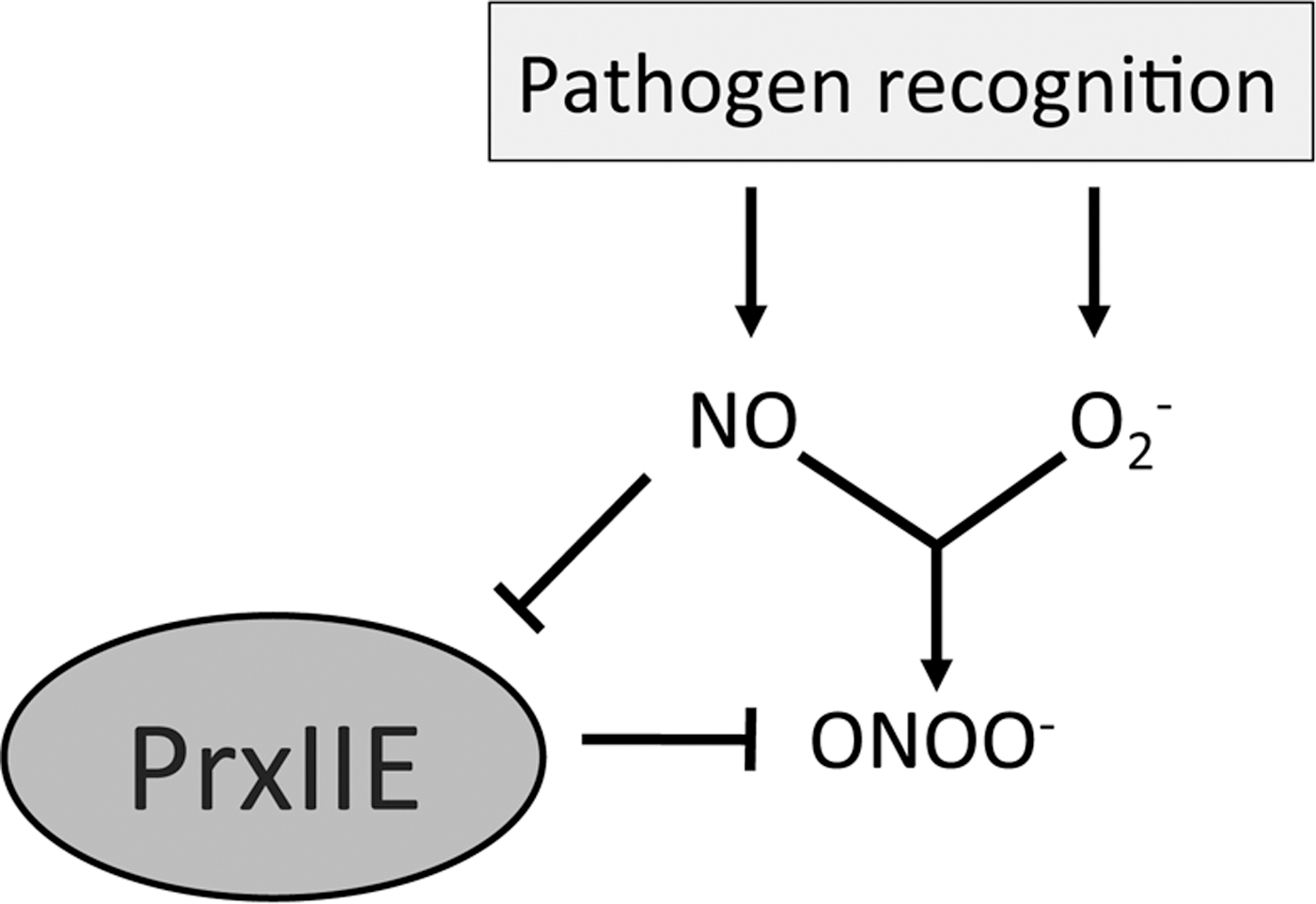
Peroxynitrite (ONOO−) is a reactive toxic agent formed by NO reacting with O2 −. Peroxiredoxin II E (PrxIIE) is responsible for ONOO− detoxification in plants and S-nitrosylation of this enzyme was shown to inhibit this activity (33) (Fig. 7). ONOO− has been shown to induce programmed cell death (PCD) in animals (6) but not in plants, even in a PrxIIE mutant line (33). The S-nitrosylation of PrxIIE may serve as a regulatory mechanism that NO employs to control the levels of ONOO−, but whether this mechanism is implicated in PCD in plants remains an unanswered question. Caspases are animal proteins involved in triggering PCD in humans, and caspase-3 has been shown to be S-nitrosylated at its active-site cysteine in unstimulated human cell lines (28). This S-nitrosylation suppressed the cysteine protease activity of caspase-3, which is required for triggering PCD. Upon induction of the apoptotic pathway, SNO-caspase-3 levels decreased and this was associated with an increase in intracellular caspase activity and subsequent increased PCD. The Arabidopsis relative of caspase-3, metacaspase 9, has also been shown to be S-nitrosylated and similarly this suppresses its proteolytic activity (1); however, the biological relevance of this is not yet as well understood as in animals.
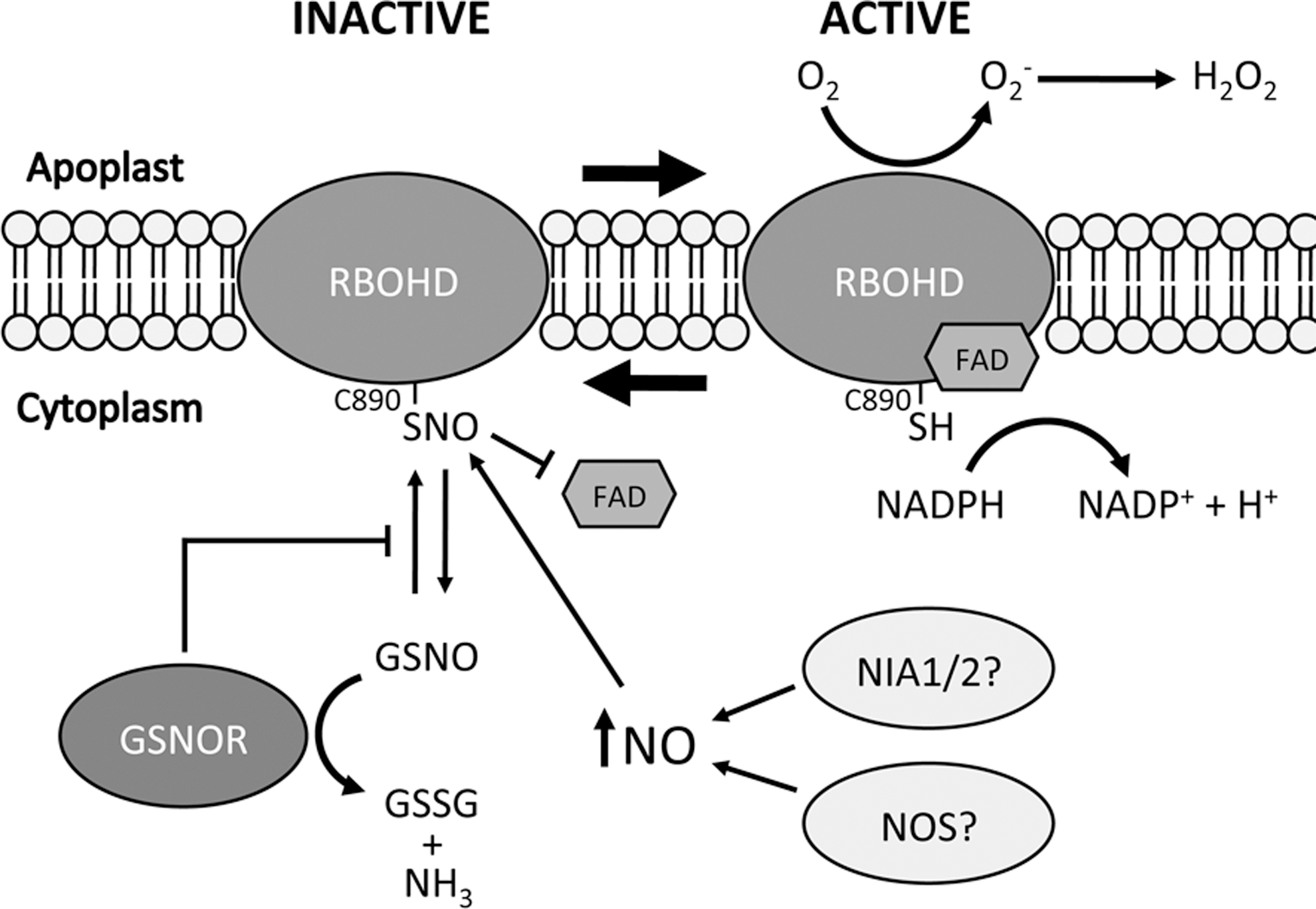
Recent work has identified Arabidopsis RBOHD as a target for S-nitrosylation during the plant immune response (48). The authors found that the production of ROIs after pathogen challenge was reduced in both gsnor1-3 and NO overproducing 1 mutant plants. Since both of these lines have elevated SNO levels it was hypothesized that S-nitrosylation of RBOH proteins could be regulating their ability to produce ROIs. Indeed, RBOHD was shown to be S-nitrosylated at Cys890 both in vitro and in vivo and this modification inhibited its NADPH oxidase activity. Computational modeling and experimental evidence showed that S-nitrosylation of Cys890 inhibited binding of the cofactor FAD, and this effect was abolished in a C890A mutant protein. Transgenic plants expressing a C890A mutant RBOHD showed greater pathogen-induced ROI accumulation and increased cell death during the HR. These findings suggest an interesting negative feedback loop by which the increasing NO levels during the HR promote S-nitrosylation of RBOHD at Cys890, attenuating its NADPH oxidase activity to prevent excessive ROI-induced cell death (Fig. 8). Importantly this cysteine reside is conserved from plants to humans and this study also showed that recombinant human and Drosophila melanogaster NADPH oxidases can be S-nitrosylated at this site in vitro, suggesting that this mechanism of inhibiting cell death may also operate in other complex eukaryotes.
Identification of SNO-Proteins in Plants
The highly labile nature of the S–NO bond makes it a technically challenging post-translational modification to detect. Most work to date has relied on a technique known as the biotin-switch (17), where free cysteines are first blocked before SNOs are specifically reduced and “switched” with a biotin label to allow detection with antibodies against biotin or purification with streptavidin. Recent years have seen the emergence of proteome-wide screens for S-nitrosylated proteins in Arabidopsis (25), however, some of these have involved the application of an exogenous NO-donor and thus may not reflect the relevant in vivo catalogue of SNOs. Only one report to date exists describing the SNO-proteome of plants engaged in defense signaling (32) and this only uncovered 16 proteins involved in metabolism, antioxidant defense, and signaling. The fact that other proteins not identified in this study have been shown to be S-nitrosylated during defense signaling suggests that many other targets of this important modification are yet to be uncovered. A more recent study investigated the endogenous SNO-proteome under resting conditions and after salt stress (14) with only 53 proteins being identified as S-nitrosylation targets. Again, many proteins described in the previous section as being regulated by S-nitrosylation were not picked up in this screen although this could be expected as the plants in this study had not been challenged with a pathogen. Thus, a more complete assembly of the SNO-proteome after pathogen recognition and the subsequent nitrosative burst is desirable to identify more defense-related targets regulated by NO.
Denitrosylation by Thioredoxin Proteins in Plant Defense?
Although the cellular redox status and GSNOR activity govern global SNO levels, emerging data has revealed that NO can be specifically removed from proteins by enzymes with denitrosylating activity. Thioredoxin (TRX) proteins have been shown to reduce disulfide bonds and are of particular interest. Arabidopsis TRX-h5 (TRX5) has been shown to facilitate the reduction and subsequent monomerization of NPR1 oligomers upon SA-mediated plant defense activation leading to upregulation of defense genes (40). TRX5 has been implicated in plant immunity in response to victorin, produced by Victoria blight (39), and it is upregulated in response to avirulent bacterial pathogens (21). TRX proteins have recently emerged as enzymes with denitrosylating activity in human cells (2) and together with thioredoxin reductase (TRX-reductase) can remove SNO groups from target proteins. Interestingly, some proteins appear to be basally S-nitrosylated with stimulus-induced reduction/denitrosylation by TRX/TRX-reductase providing a cellular signal, while others are constitutively kept in their reduced/denitrosylated form by basal activity of TRX/TRX-reductase. A recent animal proteomic study revealed a list of SNO-proteins with diverse functions to be substrates for TRX/TRX-reductase (3) thus further enhancing the notion that denitrosylation of proteins by TRX may have important roles in cellular signaling (37).
Conclusions and Future Perspectives
A huge effort has been aimed at understanding plant NADPH oxidases and their role in producing the oxidative burst following pathogen recognition. However, the emerging data reveal that this class of enzyme may not provide the only source of ROIs, with new evidence implicating GOX and peroxidases in ROI synthesis after immune activation. It is likely that further contributors to the oxidative burst will be identified in the coming years. In addition to new players in the oxidative burst, the discovery of an NOS in higher plants would significantly move the field forward, although the current data suggest it may be of a structure different from the well-established mammalian NOSs. It is now appreciated that NO exerts its biological effect through the S-nitrosylation of reactive cysteine thiols in proteins and thus it is important to identify new targets of this key post-translational modification. Moreover, recent findings have confirmed that S-nitrosylation is an important regulatory mechanism in immune signaling. Thus, this redox-based modification is a potential target to exploit in breeding or plant design approaches to generate improved disease-resistant crops. Further, as dysregulation of S-nitrosylation is found to underpin the development of an increasing number of human diseases, the molecular machinery associated with this redox modification could represent an attractive drug target for future therapeutics.
Footnotes
Acknowledgments
M.J.S. was the recipient of a Biotechnology and Biological Sciences Research Council (BBSRC) studentship, and work in the lab of G.J.L. is supported by BBSRC grant BB/D0118091/1.