
Abstract
Aims:
Cationic manganese (Mn)-substituted N-pyridylporphyrin-based potent mimics of the family of superoxide dismutases (SODs) protect normal tissues from injury related to ionizing radiation (IR) by reducing levels of reactive oxygen and nitrogen species (ROS/RNS). Furthermore, Mn-porphyrins have demonstrated antitumor and radiosensitizing effects on cancer cells by promoting IR-induced tumor vasculature damage and apoptotic processes. In this study, we explored the underlying mechanisms of Mn-porphyrin-mediated tumor radiosensitization using murine mammary carcinoma 4T1 and melanoma B16 cells in vitro and in vivo.
Results:
Combination treatment with MnTnHex-2-PyP and IR substantially reduced cell viability, clonogenic cell survival, and DNA damage repair and synergistically increased IR-induced apoptosis of 4T1 and B16 cells. MnTnHex-2-PyP in combination with IR caused a significant delay in growth of 4T1 and B16 xenograft tumors. MnTnHex-2-PyP dose-dependently enhanced IR-mediated production of H2O2-derived species, but not superoxide. Catalase overexpression reversed MnTnHex-2-PyP-enhanced ROS production and apoptosis. Demonstrated suppression of phosphorylation of several mitogen-activated protein (MAP) kinases and activation of NF-κB by MnTnHex-2-PyP/IR, which presumably inhibited activation of the antiapoptotic pathway, are in agreement with our other data on the apoptosis of cancer cells.
Innovation and Conclusions:
MnTnHex-2-PyP exerted a radiosensitizing effect on 4T1 and B16 tumor models in vitro and in vivo via pro-oxidative actions and therefore bears a large therapeutic potential. When combined with IR, it attenuated DNA damage repair and triggered a shift from prosurvival pathways to apoptotic cell death, likely due to increased ROS production and disturbed cellular redox balance, acting at the level of nuclear factor κB (NF-κB). Antioxid. Redox Signal. 27, 1067–1082.
Introduction
R
Normal tissue damage during radiation therapy is triggered by free radicals from water radiolysis. Based on these reactions, manganese superoxide dismutase (Mn-SOD), an antioxidant enzyme-catalyzing superoxide, has been tested as a potential endogenous radioprotector (24). Although direct application of Mn-SOD in rodents mitigates radiation-induced lung damage (14, 29), successful delivery of Mn-SOD to target tissues remains technically challenging. To overcome this challenge, a variety of Mn-SOD mimics such as Mn-porphyrins have been developed and are being tested in preclinical studies for the protection of normal tissues from radiation-induced injury (2, 4).
Manganese (Mn)-porphyrin-based drugs have been suggested as redox-modulating agents for diseases, including cancer. Mn-porphyrin-based superoxide dismutase (SOD) mimics are believed to broaden the therapeutic window for radiotherapy via radioprotection in normal tissues and radiosensitization in cancer cells. This report demonstrated mechanisms of synergistic action of MnTnHex-2-PyP with radiation therapy in vitro and in vivo. Combination therapy with MnTnHex-2-PyP and radiation resulted in accumulation of reactive oxygen species (ROS), primarily long-lived H2O2, leading to attenuated DNA damage repair and increased apoptosis. S-glutathionylation, as a major oxidative modification of protein cysteines, catalyzed by MnTnHex-2-PyP in the presence of H2O2 and glutathione (GSH) in a glutathione peroxidase (GPx)-like manner, is presumably a main action of Mn-porphyrins resulting in promotion of proapoptotic pathways. Data of ours (27, 28, 54) and others (7, 43, 46) demonstrate that such oxidative damage occurs with nuclear factor κB (NF-κB) and p38 mitogen-activated protein kinase (MAPK) and is likely involved in the Mn-porphyrin-driven inhibition of other MAPKs found in this work, giving rise to strong radiosensitization. The findings of this study increase our understanding of the molecular pathways behind the effects of Mn-porphyrin-based SOD mimics involved in sensitization of cancer cells to radiation. Our studies could also help with design of new therapeutic strategies for radiation therapy. The findings have particular relevance due to the progress of Mn-porphyrins toward multiple clinical trials.
While being radioprotectors of normal tissues, Mn-porphyrins also exhibit antitumor and radio- and chemosensitizing effects on cancer cells. In a 4T1 mouse breast tumor xenograft model, Mn(III) meso-tetrakis(N-ethylpyridinium-2-yl) porphyrin (MnTE-2-PyP, AEOL10113, BMX-010), when injected as a single drug, showed anticancer effect only at a very high level of 15 mg/kg given subcutaneously (sc) daily throughout the study (48). Such treatment downregulated angiogenesis factors, including hypoxia-inducible factor 1α (HIF-1α) and vascular endothelial growth factor (VEGF), essential for tumor growth (48). Yet, much stronger effect was observed when MnTE-2-PyP was coupled with either radiation or ascorbate (53, 55). The strongest effect was observed with both sources of reactive oxygen species (ROS), ionizing radiation (IR), and ascorbate combined (53, 55). In a D245-MG glioblastoma multiforme xenograft mouse model with nu/nu Balb/c mice, Mn(III) meso-tetrakis(N-n-hexylpyridinium-2-yl) porphyrin, MnTnHex-2-PyP, showed significant tumor radiosensitization (3). MnTnHex-2-PyP was injected sc twice daily at 1.6 mg/kg, starting 24 h before radiation and continued throughout the study. Radiation was given for 3 days at 1 Gy per day. In subsequent study, Mn(III) meso-tetrakis(N-n-butoxyethylpyridinium-2-yl) porphyrin (MnTnBuOE-2-PyP, BMX-001) was tested under identical conditions and demonstrated tumor radiosensitizing effect (61). Under same conditions, MnTnBuOE-2-PyP was also shown to chemosensitize D245-MG tumor; temozolimide was given intraperitoneally for 5 days at 5 mg/kg/day (60).
With strong evidence that MnTnBuOE-2-PyP does not protect a tumor, additional studies demonstrated that it protects normal tissue against radiation (60, 61). Such data enabled the start of a clinical trial at Duke University where it is now tested as a radioprotector of normal brain (NCT02655601) with high-grade glioma patients. Another preclinical study on a head and neck cancer model showed that MnTnBuOE-2-PyP mitigated radiation-induced normal tissue damage, including mucositis, xerostomia, and fibrosis, and augmented the antitumor effect of radiation (1). Such study has enabled another clinical trial, with head and neck cancer patients on radioprotection of salivary glands and mouth mucosa, to be jointly conducted at Duke University and University of Colorado in early 2017 (NCT02990468). Taken together, such results imply that Mn-porphyrins may be ideal for broadening the therapeutic window of radiotherapy. Given the complex cellular biology and complex chemistry and biology of Mn-porphyrins, we are just starting to gain insight into the differential actions of Mn-porphyrins whereby they induce apoptotic processes in the tumor, but preserve normal tissue (3, 5, 55, 62). Nonetheless, we are far away from fully comprehending such differences.
The very first indication provided by us showed that in inflammatory breast cancer cell line, MnTE-2-PyP suppressed phosphorylation of extracellular signal-regulated kinase (ERK), which in turn suppressed activation of NF-κB (15). Herein, we went one step further and looked into the effect of Mn-porphyrins on different mitogen-activated protein (MAP) kinases, which are critical for cellular apoptotic and proliferative pathways, ERK, c-Jun N-terminal kinase (JNK), p38, and AKT (57), with the ultimate goal to understand underlying mechanisms of Mn-porphyrin-mediated tumor radiosensitization using murine mammary carcinoma 4T1 and melanoma B16 cells. We have chosen to work with lipophilic analog, which has shown impressive efficacy in numerous studies, including central nervous system injuries, cancer, diabetes, and radiation injury (3, 5). Due to its high lipophilicity, it accumulates at high level in all tissues, brain and brain mitochondria included (3, 5, 51). Our in vitro data revealed that MnTnHex-2-PyP attenuated DNA damage repair after irradiation, suppressed phosphorylation of several MAP kinases, and enhanced radiation-induced apoptosis. The same radiosensitizing effect of MnTnHex-2-PyP was observed in vivo using xenograft mouse models. Taken together, our findings support the potential of MnTnHex-2-PyP as a therapeutic agent for radiation therapy.
Results
In vitro radiation-sensitizing effect of MnTnHex-2-PyP
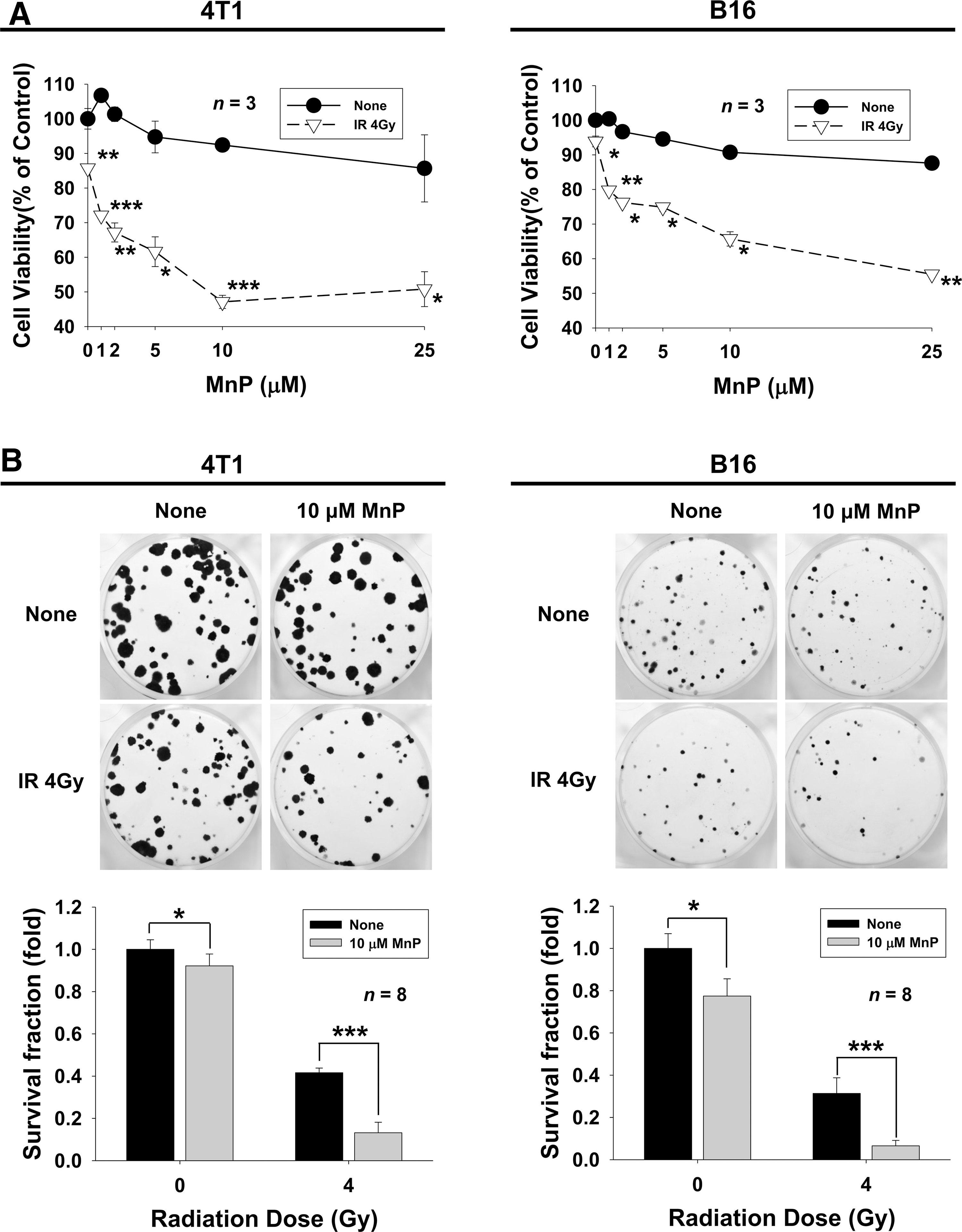
Cytotoxic effects of the SOD mimetic MnTnHex-2-PyP in combination with IR were determined in 4T1 mouse mammary carcinoma cells and B16 mouse melanoma cells by Cell Counting Kit-8 (CCK-8) assays (Fig. 1A). Treatment with 25 μM MnTnHex-2-PyP decreased cell viability of both 4T1 and B16 cells with 14.3% and 12.4% reduction, respectively. Treatment with 4 Gy IR alone decreased 14.3% and 6.1% of cell viability in 4T1 and B16 cells compared with control. The combination of 4 Gy IR and 10 μM MnTnHex-2-PyP resulted in a 52.9% reduction in cell viability of 4T1 cells compared with control and 34.3% reduction of B16 cells compared with 7.6% and 9.3% with 10 μM MnTnHex-2-PyP alone. The radiosensitizing effects of MnTnHex-2-PyP were also assessed by clonogenic assays (Fig. 1B). Combined treatment with 10 μM MnTnHex-2-PyP and 4 Gy IR resulted in a significant reduction (86.8% and 93.6%, p < 0.001) in clonogenic cell survival in 4T1 and B16 cells compared with MnTnHex-2-PyP (8.9% and 22.6%) or IR alone (58.4% and 68.6%). Similar results were seen in human hepatoma cell line Huh7 (Supplementary Fig. S1A; Supplementary Data are available online at
MnTnHex-2-PyP inhibits repair of radiation-induced double-strand breaks
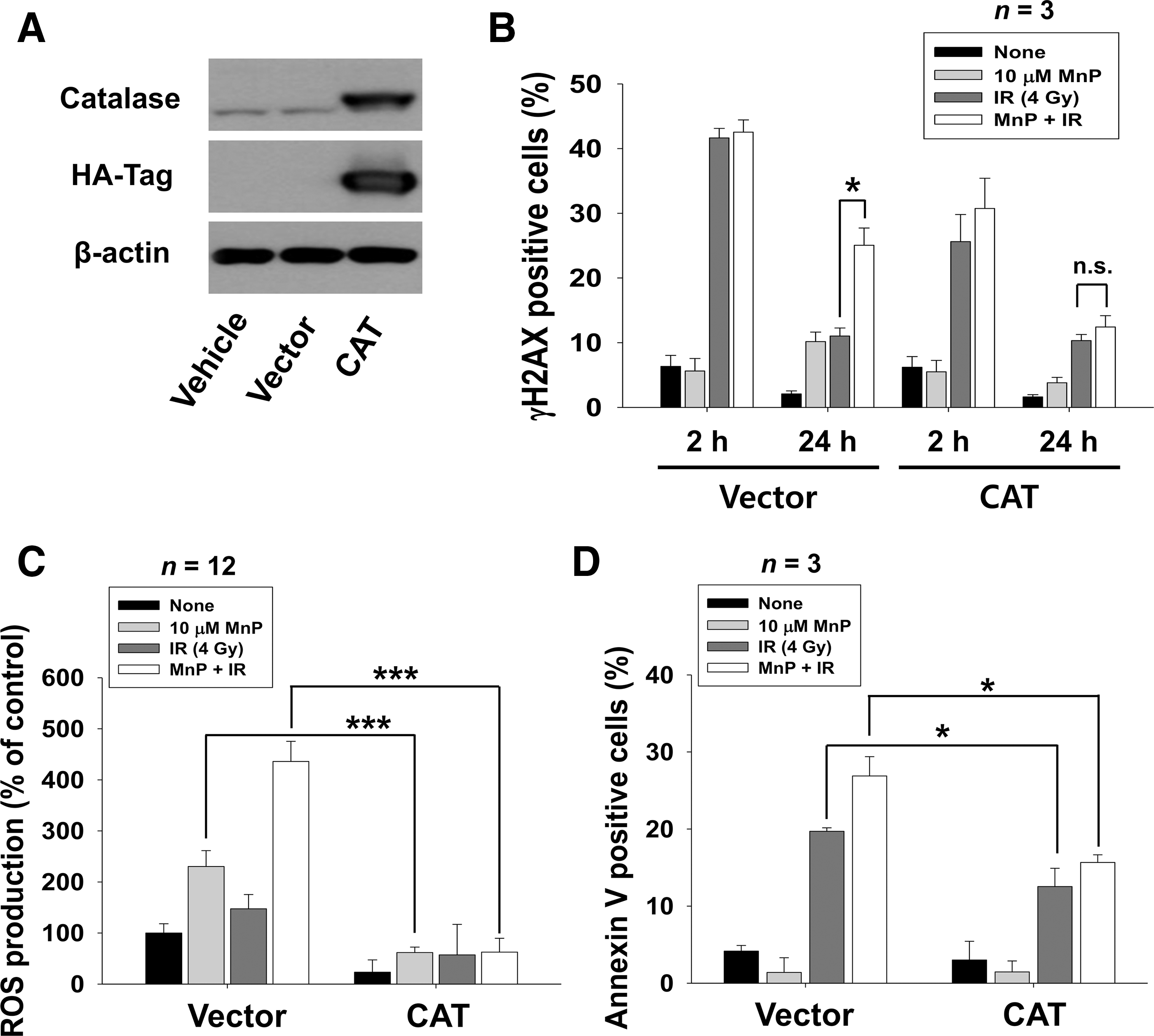
To elucidate the molecular mechanism of the radiosensitizing effects of MnTnHex-2-PyP, we determined the ability to repair DNA double-strand breaks (DSBs) by evaluating foci of phosphorylated histone H2AX (γ-H2AX), a sensitive indicator of DSB formation. Immunostaining with anti-γ-H2AX antibodies showed γ-H2AX foci in the nucleus in both 4T1 and B16 cells at 2 h of irradiation; these mostly disappeared at 24 h (Fig. 2A). In contrast, γ-H2AX foci persisted until 24 h after irradiation with MnTnHex-2-PyP cotreatment. Quantification of γ-H2AX focus number per nucleus showed that cells treated with MnTnHex-2-PyP retained significant amount of γ-H2AX foci at 24 h postirradiation compared with no treatment control (p < 0.001) (Fig. 2B). We also assessed the effect of MnTnHex-2-PyP on DNA repair efficacy using flow cytometry. The percentage of γH2AX-positive cells at 24 h after irradiation was greater in cells pretreated with MnTnHex-2-PyP compared with radiation alone (Fig. 2C). Prolonged detection of γ-H2AX foci was also seen in human Huh7 cells cotreated with IR and MnTnHex-2-PyP (Supplementary Fig. S1B, C). The persistent existence of γH2AX-positive cells at 24 h after irradiation reflects inefficient repair of DSBs.
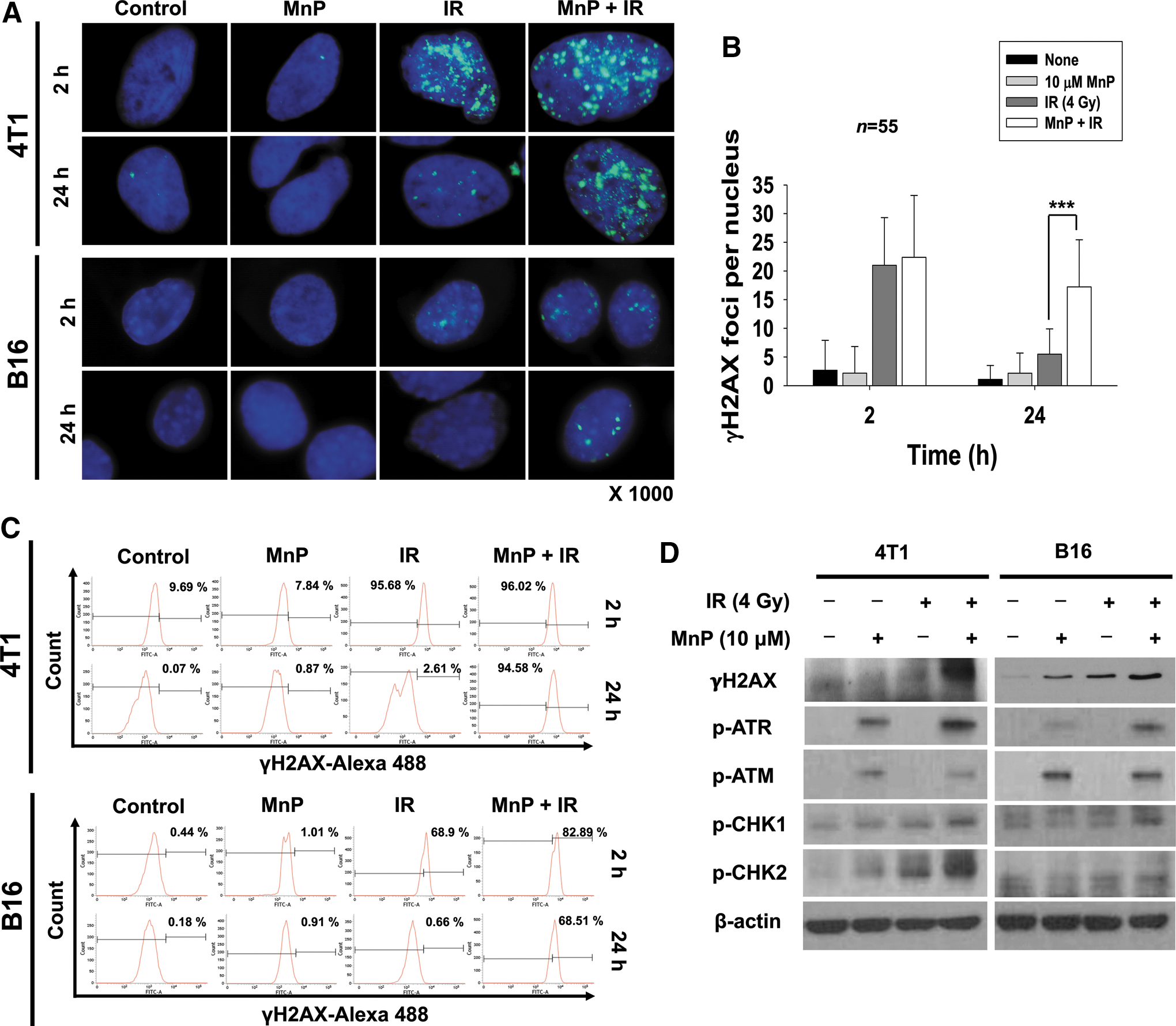
Next, we examined the effect of MnTnHex-2-PyP on DNA damage signaling in response to IR. γ-H2AX expression remained high at 48 h of irradiation in 4T1 and B16 cells cotreated with MnTnHex-2-PyP (Fig. 2D), which is consistent with the immunostaining and flow cytometry data of γ-H2AX focus formation as shown in Figure 2A and B. Cotreatment of MnTnHex-2-PyP and IR also increased phosphorylation of ATR and ATM, DNA damage sensors. In addition, cotreatment led to prolonged activation of CHK1 and CHK2 that are downstream of ATR and ATM, respectively.
MnTnHex-2-PyP enhances radiation-induced apoptosis
We examined the effects of MnTnHex-2-PyP on IR-mediated cell cycle redistribution using flow cytometry (Fig. 3A). DNA content analysis with propidium iodide (PI) staining revealed that 10 Gy IR increased the sub-G1 and G2/M populations in 4T1 and B16 cells after 72 h. The effect was further enhanced by combination with MnTnHex-2-PyP (Fig. 3A). Induction of the sub-G1 peak via cotreatment with IR and MnTnHex-2-PyP may represent DNA fragmentation during apoptosis.
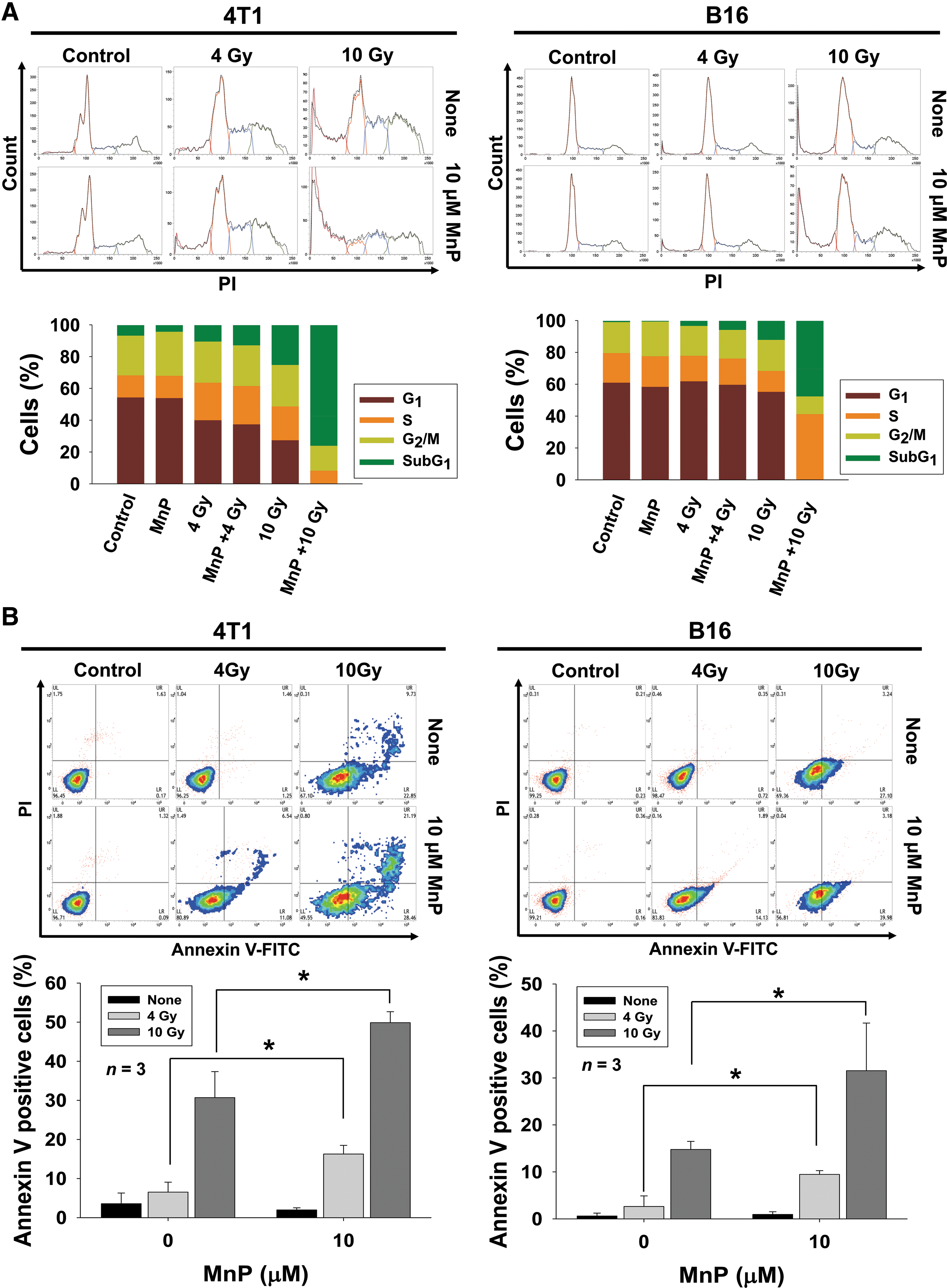
Given that the induction of the sub-G1 peak may be due to DNA fragmentation during apoptosis, we determined the impact of MnTnHex-2-PyP on radiation-induced apoptosis using flow cytometry with Annexin V and PI double staining. Annexin V has a strong affinity to phosphatidylserines exposed on the outside of cell membranes due to disruption of membrane integrity during apoptosis. Compared with IR alone, combination treatment with MnTnHex-2-PyP and IR significantly increased the fraction of Annexin V-positive cells in 4T1 (30.7% vs. 49.9% at 8 Gy, p < 0.05) and B16 cells (14.8% vs. 31.5% at 8 Gy, p < 0.05), proportional to the radiation dose (Fig. 3B). Enhancement of IR-induced apoptosis by MnTnHex-2-PyP was also seen in Huh7 cells (Supplementary Fig. S1D).
MnTnHex-2-PyP enhances delayed tumor growth in mice receiving radiotherapy
The in vitro radiosensitization effect of MnTnHex-2-PyP was verified using in vivo mouse models. We implanted 4T1 into syngeneic BALB/c mice and B16 cells into syngeneic C57BL/6 mice and evaluated the ability of MnTnHex-2-PyP to inhibit tumor growth in combination with radiotherapy. Tumor-bearing mice were treated with 5 Gy for 3 consecutive days for a total of 15 Gy. Mice received daily intraperitoneal injections of MnTnHex-2-PyP (2 mg/kg/day), starting 3 days before irradiation. Consistent with in vitro results, the in vivo data showed that treatment with MnTnHex-2-PyP alone did not change 4T1 and B16 tumor growth. Combination with radiation therapy reduced tumor growth compared with radiation therapy alone (Fig. 4A, B). At 15 days, the average volume of tumors in the 4T1 xenograft mice cotreated with MnTnHex-2-PyP and radiation was reduced by 78.9% and 48.4% (p < 0.05) compared with the control group and radiation-alone group, respectively. Similar results were seen in the B16 xenograft mice. We did not observe unusual symptoms such as sickness or body weight loss in mice treated with MnTnHex-2-PyP.
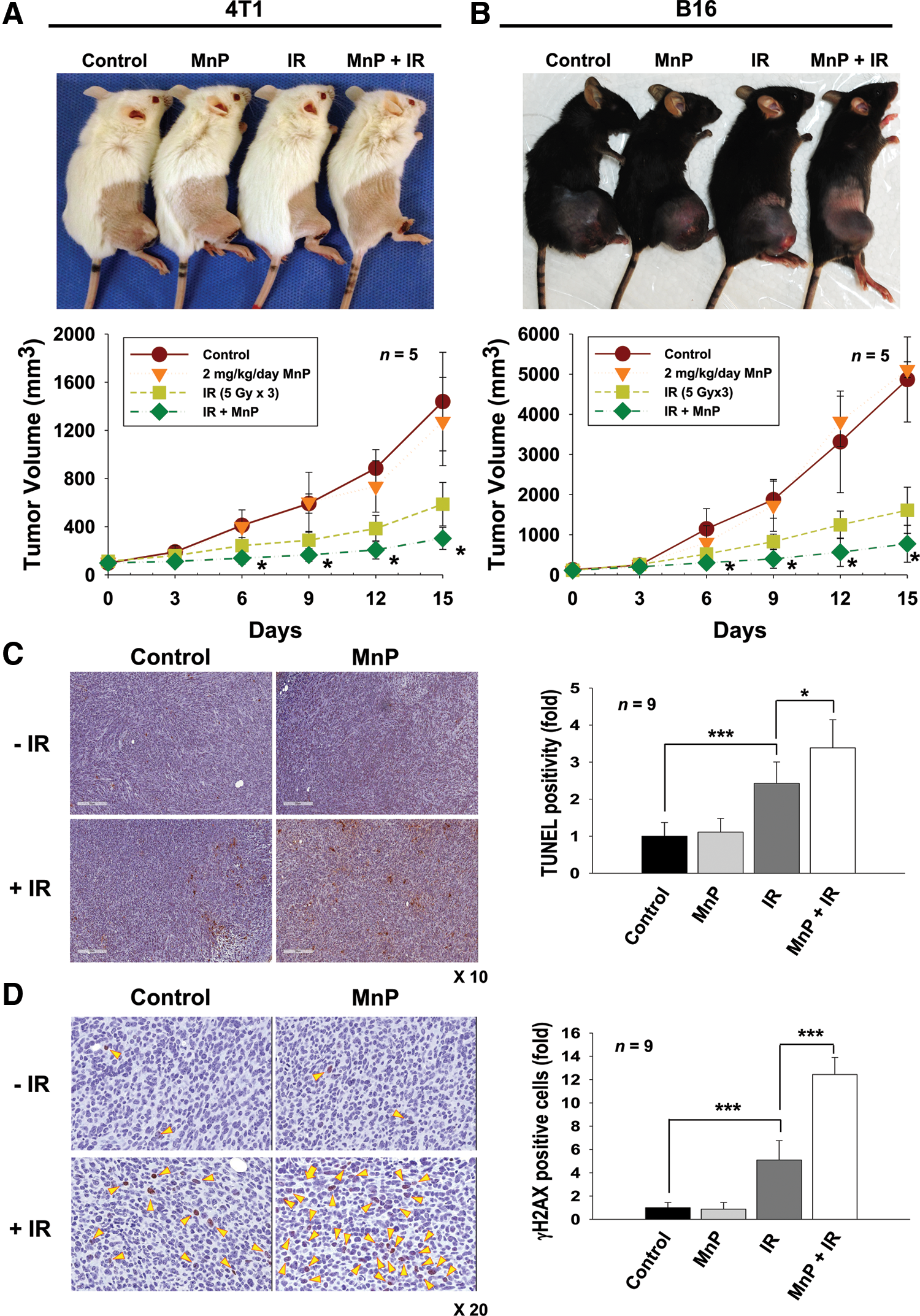
Based on our in vitro results that MnTnHex-2-PyP enhanced IR-induced apoptosis in cells, we performed terminal deoxynucleotidyl transferase dUTP nick end-labeling (TUNEL) assays to determine the apoptosis level in tumor tissues from mice treated with MnTnHex-2-PyP. Histological analysis and quantification data on TUNEL-stained tumor sections revealed that irradiation significantly increased TUNEL-positive cells (p < 0.001), which was further enhanced by cotreatment with MnTnHex-2-PyP (p < 0.05), indicating increased apoptosis (Fig. 4C). Immunohistochemical assessment of γ-H2AX expression on tumor tissues at 24 h postirradiation showed that significant increase in γ-H2AX-positive cells was seen in tissue sections cotreated with MnTnHex-2-PyP and IR compared with those treated with IR alone (p < 0.001) (Fig. 4D). These data suggested a strong relationship between prolonged DNA damage and radiosensitization in MnTnHex-2-PyP-treated cells or tissues.
MnTnHex-2-PyP acts as a pro-oxidant in cancer cells
Given that MnTnHex-2-PyP-mediated enhancement of sensitivity of 4T1 and B16 cancer cells to radiation would be related to disturbance of cellular redox balance (4), we measured intracellular ROS levels in cells treated with radiation and MnTnHex-2-PyP using the redox-sensitive dye carboxy-H2DCFDA (DCFDA). Fluorescence measurements showed that treatment of 4T1 cells with MnTnHex-2-PyP dose-dependently increased ROS levels (Fig. 5A), suggesting MnTnHex-2-PyP was a pro-oxidant. MnTnHex-2-PyP further dose-dependently increased intracellular ROS levels when combined with IR.
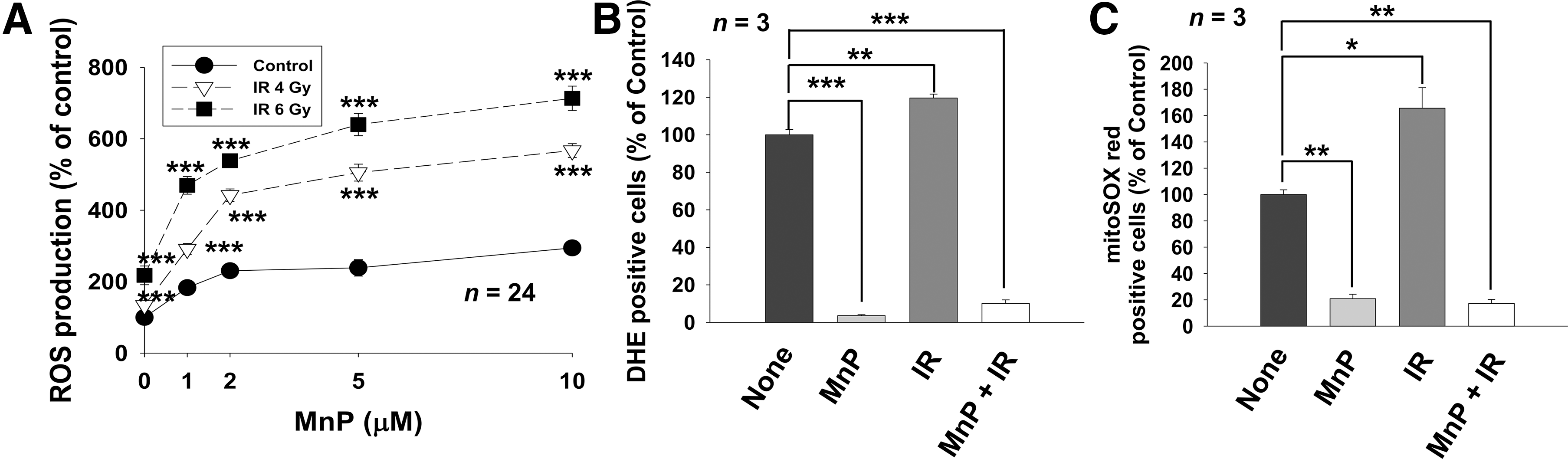
Among Mn-porphyrin-based SOD mimics, MnTnHex-2-PyP is a highly effective catalyst of superoxide dismutation that produces molecular oxygen and hydrogen peroxide (3, 4). We monitored superoxide production after treatment with IR and MnTnHex-2-PyP using fluorescent dyes sensitive to superoxide. Flow cytometry analysis on cells stained with dihydroethidium (DHE) showed that 4 Gy IR increased intracellular superoxide production, which was suppressed by treatment with MnTnHex-2-PyP (Fig. 5B). Staining with MitoSOX Red, a mitochondrial superoxide indicator, showed similar results (Fig. 5C), indicating that MnTnHex-2-PyP acts as a potent SOD mimic scavenging superoxide from both cytosol and mitochondria.
Since superoxide levels were significantly decreased by treatment with MnTnHex-2-PyP, it is likely that increased ROS levels by MnTnHex-2-PyP may be attributed to increased H2O2 production, a by-product of superoxide dismutation. We assessed the effect of catalase overexpression in 4T1 cells to determine whether MnTnHex-2-PyP acts as a pro-oxidant by generating H2O2. HA-tagged catalase overexpression was confirmed by Western blot analysis using HA-tag and catalase antibodies (Fig. 6A). Flow cytometry analysis on cells stained with γ-H2AX antibody showed that attenuated IR-induced DNA damage repair by MnTnHex-2-PyP was partially restored in 4T1 cells expressing catalase, but not in cells expressing a control vector (Fig. 6B). DCFDA staining showed that catalase overexpression significantly reduced total ROS levels increased by either MnTnHex-2-PyP or a combination of IR and MnTnHex-2-PyP compared with cells transfected with a control vector (p < 0.001) (Fig. 6C). Further analysis on apoptosis showed that increased apoptosis by either IR alone or a combination of IR and MnTnHex-2-PyP was suppressed by catalase overexpression (p < 0.05) (Fig. 6D). Together, our data suggested that MnTnHex-2-PyP may act as a pro-oxidant in cancer cells and increased H2O2-caused radiosensitization.
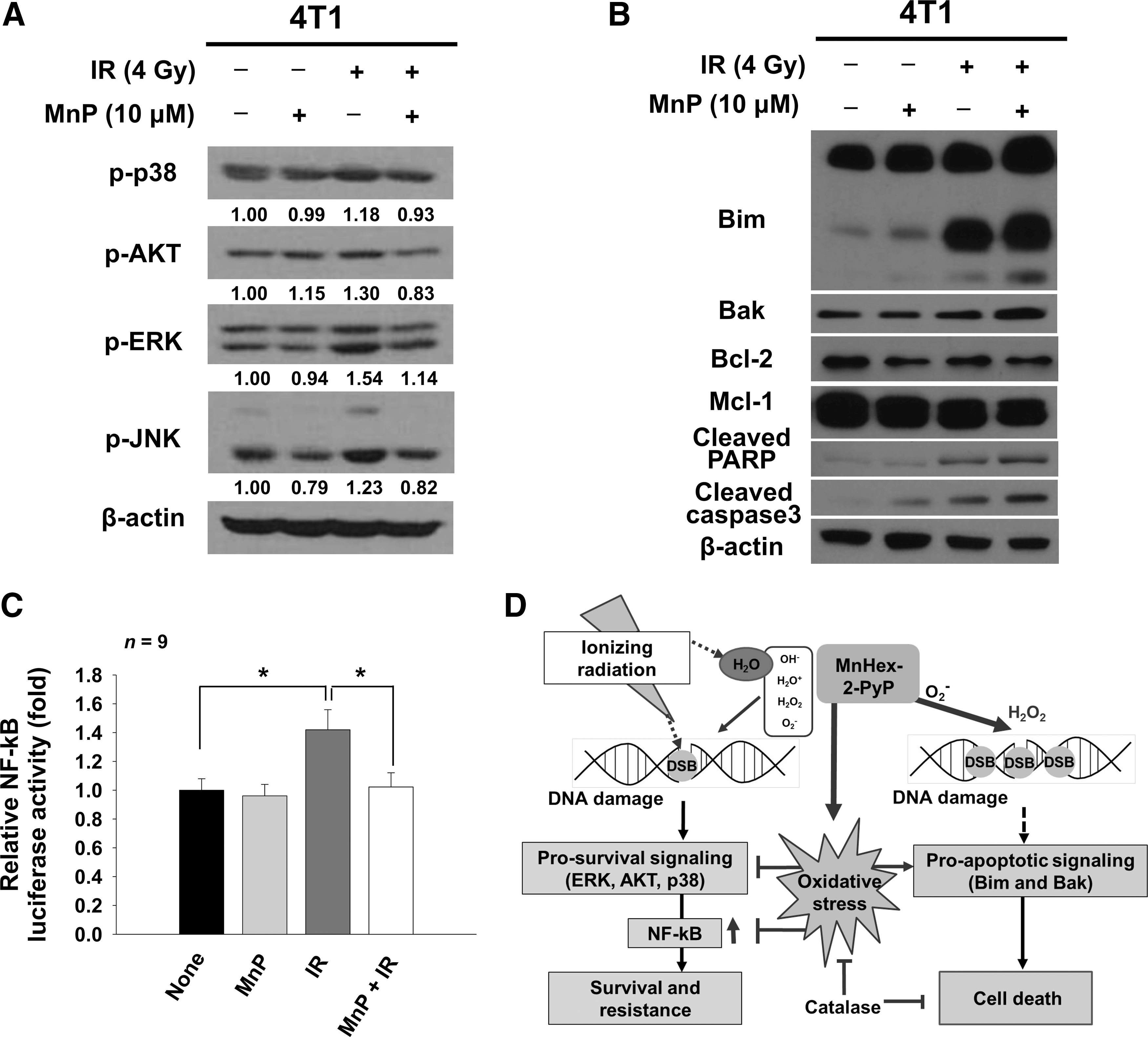
MnTnHex-2-PyP triggers the shift from prosurvival signaling to cell death
IR activates prosurvival signaling pathways, which contributes to tumor radiation resistance and limits the efficacy of radiation therapy (23, 42, 57). To assess how MnTnHex-2-PyP increased the efficacy of radiation therapy, we determined effects of MnTnHex-2-PyP on IR-activated mitogen-activated protein kinase (MAPK) signaling in 4T1 cells. Western blots showed that MnTnHex-2-PyP decreased expression of phosphorylated p38 MAPK, AKT, ERK, and JNK that were induced by IR treatment, suggesting inhibition by MnTnHex-2-PyP of IR-induced activation of prosurvival signaling (Fig. 6A).
We also determined effects of MnTnHex-2-PyP on antiapoptotic and proapoptotic effectors in 4T1 cells. Upregulation of the proapoptotic proteins, Bim and Bax, was seen in 4T1 cells cotreated with MnTnHex-2-PyP and IR. Levels of the antiapoptotic proteins, B cell lymphoma 2 (Bcl-2) and myeloid cell leukemia 1 (Mcl-1), were not changed (Fig. 6B). These data suggested that increased expression of proapoptotic proteins may be closely correlated with increased apoptosis after combination therapy of IR and MnTnHex-2-PyP. This hypothesis was confirmed by increased expression of cleaved poly ADP ribose polymerase (PARP) and caspase-3, which are surrogate markers for apoptosis (Fig. 6B). IR-activated survival signaling leads to subsequent activation of the downstream nuclear factor κB (NF-κB) transcription factor, resulting in antiapoptotic gene expression. Luciferase-based NF-κB promoter activity assay revealed that increased NF-κB activity by 4 Gy IR returned to basal levels when MnTnHex-2-PyP was cotreated (Fig. 6C).
Discussion
Mn-porphyrin complexes have been extensively studied as potent redox-active therapeutics. Their ability to radioprotect normal tissue has been well demonstrated (1 –4, 26, 60, 61). As SOD mimics, Mn-porphyrins exert their protective effects either by direct scavenging of ROS and reactive nitrogen species (RNS) generated by water radiolysis during radiation therapy or by direct interaction with cellular proteins bearing redox-active cysteine (3, 5, 27, 28). Furthermore, most potent cationic Mn(III)-substituted N-pyridylporphyrins have tumor radiosensitization activity (1, 3, 33, 41, 60, 61), but the mechanism by which Mn-porphyrins sensitize cancer cells to radiation remains largely unexplored. A previous study with MnTE-2-PyP showed that it dramatically attenuated in vivo tumor growth when combined with radiation therapy. Initially, it was speculated that MnTE-2-PyP scavenges ROS/RNS that are necessary for activation of HIF-1, which further blocks secretion of proangiogenic cytokines such as VEGF, thereby promoting tumor devascularization (41). A recent study in a head and neck cancer model revealed that MnBuOE-2-PyP suppresses FaDu xenograft tumor growth by increasing infiltration of tumor-associated macrophages with M1 phenotype in the tumors, although the mechanism is not known (1). The effect on M1 may also be NF-κB controlled (22). Furthermore, the effect of MnTnBuOE-2-PyP on NF-κB pathway was seen in the D245-MG glioma xenograft study (3). In support of the effects at the level of NF-κB, results came from the breakthrough data on in vitro and in vivo anticancer effects of Mn-porphyrins provided by Tome's group study on lymphoma cells. The data demonstrate that potent Mn-porphyrin-based SOD mimic, MnTE-2-PyP— and most so when combined with dexamethasone—in the presence of H2O2 and glutathione (GSH), interacts with cysteines of different proteins, including master transcription factor NF-κB, inducing their S-glutathionylation via glutathione peroxidase (GPx)-like actions (3 –5, 8, 28). The impact of Mn-porphyrin on caspases 3 was also seen.
In addition to NF-κB, MnTE-2-PyP S-glutathionylated complexes I and III caused their inactivation, which in turn resulted in loss of ATP (17). The effect on glycolysis was also seen (27) and similarly it was seen with MnTnBuOE-2-PyP (Tome, unpublished data). Such cytotoxic effects of MnTE-2-PyP were only seen with lymphoma cells, but not with normal lymphocytes (27). Studies on several other cancer and normal cell lines support such conclusion on toxicity of Mn-porphyrins when combined with source of ROS (radiation, coupling with ascorbate, or chemotherapy) toward cancer, but not normal cells (3, 9, 63). Detailed studies on normal cells await further exploration. However, even the understanding of anticancer effects of Mn-porphyrins, with regard to hypoxia, angiogenesis, and immune responses that control tumor growth, is in infancy. We aimed here to further our understanding on pathways involved in the actions of MnHex-2-PyP in enhancing radiation-induced killing effects at the molecular and cellular levels.
The cytotoxicity of MnHex-2-PyP in murine 4T1 and B16 cells was low, but was enhanced by combination with IR. Treatment with 10 μM MnHex-2-PyP resulted in 10–20% reduction in the colony formation ability of 4T1 and B16 cells and IR treatment reduced colony formation up to 60–70% in both cell types. Combination treatment with 10 μM MnHex-2-PyP and IR further reduced colony formation, suggesting a synergistic effect. Similar effects were seen with human hepatoma Huh7 cells. Our in vivo animal experiments showed that administration of MnTnHex-2-PyP (2 mg/kg/day) did not affect 4T1 and B16 tumor growth compared with a control group, but inhibited tumor growth when combined with IR. This finding indicated a radiosensitizing effect of MnTnHex-2-PyP in vivo, consistent with previous studies on Mn-porphyrins (1, 41, 60, 61). Thus, Mn-porphyrins have potential to sensitize tumor cells to radiation therapy regardless of tumor type.
Lethal damage to DNA such as DSBs occurs in normal and tumor cells during radiation therapy. Their capacity for repair determines radiation sensitivity. To determine the effects of MnTnHex-2-PyP on DNA damage repair processes in cancer cells, we performed γ-H2AX focus assays. Analysis of DSB repair kinetics using immunofluorescence showed that 4T1 and B16 cells treated with MnTnHex-2-PyP had comparable levels of γ-H2AX focus formation 2 h after irradiation, but retained the foci until 24 h. This result indicated that unrepaired DSBs remained. Flow cytometry confirmed that in cells cotreated with MnTnHex-2-PyP and IR, the population of γ-H2AX-positive cells persisted after 24 h, whereas the positive population returned to basal levels in cells treated with IR alone. Immunohistochemistry (IHC) analysis on tumor tissues at 24 h postirradiation exhibited enhanced γ-H2AX expression by administration of MnTnHex-2-PyP. Our data suggested that MnTnHex-2-PyP attenuated the process of repairing radiation-induced DNA damage in cancer cells in vitro and in vivo. The opposite was found in a previous study with normal cell type—ataxia telangiectasia—showing that MnTnHex-2-PyP reduces radiation-induced DNA damage (47). Such data are in agreement with studies on normal tissue radioprotection and are explained on the basis of different redox environments of cancer versus normal tissue (3, 5, 55, 62).
Although the main form of cell death induced by IR is mitotic death, enhancement of IR-induced apoptosis by therapeutics is important in increasing the efficacy of radiation therapy. Our results showed that MnTnHex-2-PyP increased apoptosis in combination with a radiation dose of 10 Gy, as assessed by flow cytometry with Annexin V/PI staining. Immunoblots showed that levels of cleaved caspase 3 and PARP increased with cotreatment with MnTnHex-2-PyP and IR. Consistent with this result, increased apoptosis was also observed in tissue sections from xenograft tumors cotreated with MnTnHex-2-PyP and IR. Antiapoptotic proteins, Bcl-2 and Mcl-1, were unaffected, whereas proapoptotic proteins, Bim and Bcl2-homologous antagonist/killer (Bak), increased when MnTnHex-2-PyP and IR were applied together. This finding suggested that MnTnHex-2 may increase tumor radiation sensitivity by inducing proapoptotic signaling.
Activation of prosurvival signaling pathways by IR confers tumor resistance to radiation therapy (23, 42, 57). Our data showed that IR-triggered activation of prosurvival proteins, AKT and ERK, was hindered by MnTnHex-2-PyP. Since pharmacological inhibition of AKT and ERK increases radiosensitization via inhibiting ATM-mediated DNA damage repair (23, 42), MnTnHex-2-PyP-mediated inhibition of DNA damage repair may be due to inhibition of AKT and ERK signaling. MnTnHex-2-PyP also decreased IR-mediated activation of JNK and p38-MAPK, suggesting implication of prosurvival role rather than proapoptotic role. These MAPKs have redox-active cysteines, and S-glutathionylation affects their signaling (7, 19, 43, 45, 46). Such data agree well with our redox proteomics data on 4T1 cell line exposed to Mn-porphyrin/ascorbate-derived source of H2O2 where cysteine S-glutathionylation was found with numerous proteins, including NF-κB and p38MAPK (54). Such action of Mn-porphyrins, detailed in Batinic-Haberle et al. (3, 4, 5), in essence represents GPx-like activity (8). Collectively, these data suggested that MnTnHex-2-PyP may potentiate radiation-induced tumor cell killing by shifting from prosurvival pathways, which are operative with normal cells/tissues with MnTnHex-2-PyP and its analogs (1, 3, 17, 18, 60, 61), to apoptotic cell death. The relevance of these prosurvival pathways in oncogenesis and tumor maintenance provides a possibility that downregulation of these pathways is involved in the chemotherapeutic action of combined treatment with IR and MnTnHex-2-PyP (37, 38, 50).
Beyond DNA damage-mediated apoptosis, other relevant pathways may be involved. IR-induced ROS production leads to mitochondrial dysfunction and endoplasmic reticulum stress, resulting in induction of autophagy (10). IR-induced autophagy involves the mTOR pathway via activation of PI3K and AKT, and a similar strategy to target mTOR with pharmacological inhibitors such as rapamycin (11, 44) has been applied to radiation therapy (10). We did not see any additive effect of MnTnHex-2-PyP on IR-induced autophagy, which was determined by LC3 expression (data not shown). It suggests that apoptosis may be a more relevant pathway than autophagy. In tumor microenvironment, accumulation of ROS inducers such as NO-derivatives by IR may promote immunosensitization by inducing TNF-related apoptosis-inducing ligand (TRAIL) (36). Thus, the enhanced apoptosis seen with tissues in 4T1 tumors cotreated with MnTnHex-2-PyP and IR may be related to TRAIL induction. Indeed, in a cellular brain tumor study with several glioma cell lines, we have shown that analogous lipophilic redox-active MnTnBuOE-2-PyP enhanced the effect of TRAIL and more so in the presence of another source of H2O2-ascorbate (63). A recently characterized NO donor, GIT-27/NO, exerts anticancer effects by generation of ROS and induction of apoptotic cell death (35, 39). This mode of action resembles that of combined use of MnTnHex-2-PyP and IR.
The growing evidence points to the predominantly pro-oxidative, rather than antioxidative, actions of Mn-porphyrins in both cancer and normal cells/tissues. Such actions are likely related to the Mn-porphyrin-driven catalysis of oxidative modifications of cysteines of proteins in the presence of H2O2; indeed, our data provide direct evidence that the effect is lost with catalase expression. The magnitude of the effects depends upon the magnitude of oxidative stress of those cells/tissues (5, 27, 28, 55). The differences in the yield of such reactions, controlled by differences in levels of H2O2 and Mn-porphyrins in normal versus tumor tissue (3, 5, 55), give rise to the proapoptotic processes in tumors and antiapoptotic processes in normal tissues. The latter are supposedly related to the ability of Mn-porphyrin to oxidize cysteines of Keap1, thereby activating nuclear factor erythroid 2-related factor 2 (Nrf2), which in turn upregulates endogenous antioxidative defenses (9, 12). The actions of many natural redox-active flavonoids on protection of normal tissue, such as curcumin, have been ascribed also to the activation of Nrf2 presumably via S-glutathionylation of its Keap1 cofactor (20). The same seems to be valid for redox-active nitroxides as well (21). In tumor tissues, IR-mediated activation of NF-κB confers radioresistance. As with MnTE-PyP (28), MnTnHex-2-PyP suppressed activation of NF-κB via pro-oxidative action. It may attenuate DSB repair in cells cotreated with MnTnHex-2-PyP and IR because NF-κB positively regulates homologous recombination DSB repair (58).
In agreement with data obtained on tumor cells/tissues, we found that MnTnHex-2-PyP acted as a pro-oxidant and increased ROS level when combined with IR. MnTnHex-2-PyP is an efficient SOD mimic with a rate constant for catalysis of superoxide dismutation of log k cat (O2 −) = 7.34 (2). This factor could explain why MnTnHex-2-PyP abolished IR-induced superoxide production. In agreement with recent data with MnTE-2-PyP in prostate cancer cells (52), catalase overexpression in 4T1 cells significantly lowered ROS levels, restored IR-induced DNA damage repair capacity, and diminished IR-induced apoptosis. We propose therefore that IR treatment generated ROS, including hydrogen peroxide and superoxide, and MnTnHex-2-PyP further increased ROS levels by acting as an SOD mimic or by pro-oxidant coupling with cellular antioxidants such as ascorbate and GSH. Coupling of Mn-porphyrins with ascorbate or GSH further increases H2O2 levels. It is well known that MnTnHex-2-PyP is an MnSOD mimic that is highly accumulated in mitochondria (3). Our MitoSOX and DHE staining results revealed that superoxide generated from mitochondria was scavenged by MnTnHex-2-PyP. It is likely that enhanced production of H2O2 may inhibit prosurvival protein such as NF-κB via its S-glutathionylation. Redox proteomic data demonstrate that NF-κB was found to be a major signaling protein S-glutathionylated by Mn-porphyrin in cancer cells/tissues (3, 27, 28, 54). In addition, prolonged DNA damage response signal may activate proapoptotic proteins such as Bim and Bax (Fig. 7D). The pro-oxidant effect of MnTnHex-2-PyP is dependent on IR-induced and/or ascorbate and GSH/oxygen-mediated (4) superoxide generation, which may be very low in the hypoxic condition of tumor cells in vivo due to lack of oxygen. Transient tumor reoxygenation following IR treatment may be able to supply oxygen (25). Thus, it is speculated that antiangiogenic drugs normalizing blood flow may be a plausible combination with MnTnHex-2-PyP and IR.
Redox homeostasis is deregulated in transformed cells and its perturbation can be a target for cancer therapy (56). In contrast to normal cells, in which ROS levels are tightly controlled by a variety of antioxidants, cancer cells have increased ROS levels from metabolic alterations and oncogenic signaling. This feature makes cancer cells more vulnerable to ROS-modulating agents (56). As radiation therapy is an ROS-modulating treatment, combination with other agents to target redox systems is a reasonable way for synergistic tumor control with radiation. Despite large number of studies, the complex biological effects of Mn-porphyrin-based drugs make their in vivo actions difficult to comprehend. We provided herein evidence on pro-oxidative actions of MnTnHex-2-PyP, presumably in large part at the level of cysteine-bearing proteins (such as NF-κB and MAPK) in a GPx-like manner, which in turn increase the therapeutic effects of radiation. A newly developed Mn-porphyrin, MnTnBuOE-2-PyP, is currently in clinical trials as a normal tissue radioprotector for high-grade glioma patients and head and neck cancer patients. Our findings provide mechanistic insight into the synergistic effect of MnTnHex-2-PyP/IR and indicate its large therapeutic potential as a radiosensitizer.
Materials and Methods
Cell culture and irradiation
Mouse mammary carcinoma 4T1 cells, melanoma B16 cells, and human hepatocarcinoma Huh7 cells were obtained from the Korean Cell Line Bank (Seoul National University, Seoul, Korea) and cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum (FBS), 100 U/ml penicillin, 100 μg/ml streptomycin, and 25 mM HEPES (Gibco, Carlsbad, CA). Cultures were maintained in a humidified atmosphere of 95% air/5% CO2 at 37°C. For IR treatment, cell monolayers were irradiated with various doses of 6 MV photons at a dose rate of 3.96 Gy/min from a Varian Clinac 6EX accelerator (Varian Medical Systems, Palo Alto, CA).
Reagents and antibodies
The Mn-SOD mimic, MnTnHex-2-PyP (λmax 454.5 nm, log ɛ = 5.21), was obtained from Professor Ines Batinic-Haberle at Duke University. Annexin V-fluorescein isothiocyanate (FITC) apoptosis detection kits were from BD Biosciences (San Jose, CA) and 5(6)-carboxy-2B,7′-dichlorofluorescein diacetate (carboxy-H2DCFDA) and anti-rabbit Alexa488-conjugated secondary antibodies were from Molecular Probes (Carlsbad, CA). PI, N-acetyl cysteine (NAC), and 10% neutral buffered formalin (NBF) were from Sigma-Aldrich (St. Louis, MO). Anti-phospho-H2AX(Ser139) antibodies were from Cell Signaling Technology (Beverly, MA) and Millipore (Billerica, MA). Antibodies against Bim, Bak, Bcl-2, Mcl-1, cleaved PARP, and active caspase-3 were from Cell Signaling Technology. Anti-β-actin, anti-rabbit IgG, and anti-mouse IgG secondary antibodies were from Sigma-Aldrich. Unless otherwise noted, all other chemicals were from Sigma (St. Louis, MO).
Clonogenic survival assays
Radiosensitivity was determined by clonogenic survival assays (16). Single-cell suspensions were seeded onto a six-well plate (200–1000 cells/well) 1 day before irradiation. Cells were pretreated with MnTnHex-2-PyP (10 μM) for 3 h and then exposed to single dose of 4 Gy IR. After 7–14 days, cells were fixed in 98% ethanol and stained with 0.5% crystal violet (Sigma-Aldrich) and colonies were counted using an inverted microscope (Zeiss primovert; Carl Zeiss Co., Ltd., Jena, Germany). Colonies with >50 cells were considered viable. Plating efficiency was calculated by dividing the numbers of viable colonies by the number of plated cells. Survival fraction was determined by dividing the plating efficiency of the irradiated cells by the plating efficiency of untreated control.
Cell viability assays
Cell viability was measured by Cell Counting Kit-8 (CCK-8; Dojindo). Cells were seeded in 96-well plates at 3 × 103 cells/well 1 day before irradiation. Cells were pretreated with various concentrations of MnTnHex-2-PyP for 3 h, followed by exposure to 4 Gy IR. At 72 h postirradiation, 10 μl of CCK-8 solution was added to each well and incubated for 2 h. Absorbance was measured at 450 nm using a SpectraMax i3 microplate reader (Molecular Devices, Sunnyvale, CA).
Cell cycle analysis
Cell cycle analysis was performed by flow cytometry using PI staining; 2 × 105 cells were plated in six-well plates and allowed to attach overnight. Cells were pretreated with MnTnHex-2-PyP for 3 h and exposed to IR. At 72 h postirradiation, cells were collected by trypsinization, fixed in cold 70% ethanol, washed in phosphate-buffered saline (PBS), resuspended in 1 ml of PBS containing 1 mg/ml RNase and 50 μg/ml PI, incubated in the dark for 30 min at 37°C, and analyzed by FACSVerse flow cytometer (Becton-Dickinson, San Jose, CA).
Apoptosis assays
The extent of apoptosis was evaluated by Annexin V-FITC and flow cytometry; 1 × 105 cells were plated in six-well plates and allowed to attach overnight. Cells were pretreated with MnTnHex-2-PyP for 3 h and subjected to 4 or 8 Gy IR. After 72 h of incubation, cells were treated with trypsin, washed with PBS (pH 7.4), and stained with Annexin V-FITC (BD Pharmingen, San Diego, CA) and 2 μg/ml PI in 100 μl Annexin V binding buffer (10 mM HEPES, pH 7.4, 140 mM NaCl, 2.5 mM CaCl2) for 15 min at 37°C in the dark. Samples were analyzed by flow cytometry using a BD FACSVerse flow cytometer. Data were acquired using BD FACSuite software.
DNA damage and repair analysis
IR-induced DNA damage was quantified by two independent methods, immunofluorescence staining and flow cytometry. For immunofluorescence, cells seeded on cover glasses (Marinfild, Inc., Rochester, NY) were pretreated with MnTnHex-2-PyP for 3 h and irradiated with 4 Gy IR. After 2 or 24 h of incubation, cells were fixed with 4% formaldehyde and permeabilized with 0.01% Triton X-100. After blocking with 2% FBS for 30 min, cells were incubated with anti-phospho-H2AX for 1 h, followed by Alexa Fluor488-conjugated secondary antibodies (Molecular Probes) and DAPI (Sigma) for 30 min. Cells were washed, mounted onto glass slides using glycerol, and imaged using a fluorescence microscope (Zeiss Observer D1; Carl Zeiss Co., Ltd.).
For flow cytometry, 2 × 105 cells were plated in six-well plates and allowed to attach overnight. Cells were pretreated with MnTnHex-2-PyP for 3 h and exposed to 4 Gy IR. At 2 or 24 h after irradiation, cells were collected by trypsinization and fixed in 4% formaldehyde for 10 min, followed by permeabilization with 0.01% Triton X-100 for 3 min. Blocking was performed in 2% FBS in PBS for 30 min at room temperature and incubation with anti-phospho-H2AX for 1 h. Secondary antibodies were added for 30 min. Data acquisition was performed with BD FACSVerse and analysis with BD FACSuite Software. Negative control staining was done with secondary antibodies alone.
Detection of intracellular ROS production
The intracellular total ROS level was quantified using the redox-sensitive dye carboxy-H2DCFDA. Carboxy-H2DCFDA was deacetylated intracellularly by a nonspecific esterase and oxidized by cellular peroxides, yielding the fluorescent compound, 2,7-dichlorofluorescein (DCF, λEX/λEM = 485/535 nm); 1 × 104 cells were plated in 96-well black/clear bottom plates and allowed to attach overnight. Cells were pretreated with MnTnHex-2-PyP for 3 h and then exposed to 4 or 6 Gy IR. Carboxy-H2DCFDA (20 μM) was loaded into cells just before irradiation. After 30 min postirradiation, cells were washed with PBS and DCF fluorescence was measured using a SpectraMax i3 microplate reader.
Intracellular superoxide levels in cytosol and mitochondria were concurrently monitored using flow cytometry with DHE (Molecular Probes) and MitoSOX™ Red Mitochondrial Superoxide Indicator (Molecular Probes) staining. For the assay, 2 × 105 cells were plated in six-well plates and pretreated with MnTnHex-2-PyP for 3 h and exposed to IR. After 30 min of incubation with DHE (10 μM) or MitoSOX (5 μM), cells were washed twice with PBS, trypsinized, and fluorescence was measured by a BD FACSVerse flow cytometer. Data were analyzed using BD FACSuite software.
Animal models
Animal experiments were reviewed and approved by the Institutional Animal Care and Use Committee of Samsung Biomedical Research Institute. Female Balb/c or C57BL/6 mice, 6–7 weeks old, were from Orient Bio (Seongnam, Korea). After injecting 1 × 105 cells sc into the right hind leg, tumor volumes were measured every 3 days with calipers and calculated as follows: volume = DShort 2 × DLong ÷ 2. When the mean tumor volume reached 80–100 mm3, mice were randomized to four groups. MnTnHex-2-PyP was administered by intraperitoneal injection (2 mg/kg) daily after the day of randomization. MnTnHex-2-PyP is a second-generation Mn-porphyrin-based SOD mimic. In comparison with an earlier hydrophilic analog—MnTE-2-PyP—MnTnHex-2-PyP was developed with increased lipophilic, and thus enhanced, bioavailability. Its therapeutic doses used in earlier in vivo animal studies ranged from 0.05 to 3.2 mg/kg/day (59). Based on pharmacokinetic and toxicity studies (59), the 2 mg/kg dose was chosen for this study. Loss of body weight was not seen after injections of 2 mg/kg/day for 15 days. Injections were continued until the day before sacrifice. From the third day of drug treatment, X-ray beam was directed on the tumor-bearing right hind leg over 3 consecutive days at 5 Gy per fraction for a total dose of 15 Gy. During irradiation, mice were anesthetized by intraperitoneal injection of 30 mg/kg zolazepam/tiletamine and 10 mg/kg xylazine under prescription of a veterinarian.
Western blots
Cells were harvested, washed with PBS, and treated with lysis buffer containing 20 mM Tris (pH 8.0), 137 mM NaCl, 10% glycerol, 1% Nonidet P-40, 10 mM EDTA, 100 mM NaF, 1 mM phenylmethylsulfonyl fluoride, and 10 mg/ml leupeptin. Lysates were centrifuged at 13,000 rpm for 15 min and protein concentrations determined using Bio-Rad protein assay reagent (Bio-Rad, Richmond, CA) according to the manufacturer's instructions. Proteins (20 μg) were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose membranes (Bio-Rad) and blots were blocked overnight in 5% skim milk in PBS at 4°C and probed with primary antibody overnight. Blots were probed with a monoclonal anti-β-actin antibody (Sigma) as a relative loading control. Detection of specific proteins was carried out with Amersham enhanced chemiluminescence detection reagents (GE Healthcare, Piscataway, NJ). Blots were cropped and full unedited blots are presented in Supplementary Figure S2.
TUNEL assays
Apoptosis in formalin-fixed, paraffin-embedded (FFPE) tumor tissues was determined using the transferase-mediated deoxyuridine triphosphate (dUTP)-digoxigenin nick end-labeling (TUNEL) assay. Tumor tissues were fixed with 10% NBF for 4 h and embedded in paraffin. Slices were deparaffinized in water and placed in 3% H2O2 for 10 min at room temperature. TUNEL staining was performed with in situ cell death detection kits (Roche). Images were captured using an Aperio ScanScope AT slide scanner (Leica Biosystems, Inc., Buffalo Grove, IL). Numbers of TUNEL-positive cells were determined with ImageScope software (Leica Biosystems, Inc.).
Immunohistochemistry
IHC was performed on 4-μm-thick FFPE sections. Slides were deparaffinized and rehydrated using xylene and a graded series of ethanol (absolute, 95%, 85%, 75%), followed by two 3-min washes in PBS. Antigen retrieval was performed in EDTA (10 mM, pH 9.0), which was microwaved at 90–100°C for 20 min and washed in PBS three times for 3 min. The sections were then incubated for 10 min in 3% (v/v) hydrogen peroxide to block endogenous peroxidase activity and washed in PBS three times for 3 min. Slides were preincubated in 20% normal goat serum and incubated in a humidified chamber overnight at 4°C with the primary anti-phospho-histone H2AX (1:200 dilution; Cell Signaling Technology). The sections were then washed in PBS (3 × 5 min) and incubated at 37°C for 30 min with secondary antibodies (Dako). The slides were then washed in Tris-HCl and detected with horseradish peroxidase-conjugated anti-rabbit EnVision+ kit (Dako). All slides were counterstained with hematoxylin.
Catalase overexpression
4T1 cells were seeded into a six-well plate and transfected at 60% confluency with mouse catalase expression plasmid (MG51898-NY; Sino Biological, Inc., Beijing, China) or empty plasmid using Lipofectamine 3000 (Invitrogen) according to the manufacturer's instructions. To confirm catalase overexpression, cell lysates were harvested 24 h post-transfection and subjected to Western blot analysis with antibodies against HA epitope tag and catalase.
NF-κB reporter assay
For luciferase assays, 4T1 cells (85–90% confluency in six-well plates) were transiently transfected with 2.5 mg of pGL4.32 (luc2P/NF-κB-response element [RE]/Hygro) and pGL4.51 (luc2/CMV/Neo) plasmid (Promega, Madison, WI) using Lipofectamine 3000 reagent (Invitrogen), following the manufacturer's instructions. At 24 h later, 6 × 103 of transfected cells were replated into 96-well plates and allowed to attach overnight. Cells were pretreated with 10 μM MnTnHex-2-PyP for 3 h, followed by exposure to IR. After 24 h, luciferase activity in cell lysates was measured using the Dual-Luciferase Reporter Assay System (Promega).
Statistical analysis
Statistical analysis was performed using SigmaPlot 10 (Systat Software, Inc., San Jose, CA) and Excel software (Microsoft, Redmond, WA). Statistical significance of differences between experimental groups was calculated with unpaired, two-tailed Student's t-test; p < 0.05 was considered statistically significant.
Footnotes
Acknowledgments
The authors would like to thank Dr. Mark W. Dewhirst, Duke University, for his inspiration and guidance. They also thank Dr. Yoonsun Chung, Samsung Medical Center, for providing technical support during irradiation experiments. This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2013R1A1A2010182) and by a Samsung Medical Center grant (GFO02160081).
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.