
Abstract
Aims:
Peroxiredoxin5 (Prdx5), a thioredoxin peroxidase, is an antioxidant enzyme that is widely studied for its antioxidant properties and protective roles in neurological and cardiovascular disorders. This study is aimed at investigating the functional significance of Prdx5 in mitochondria and at analyzing its roles in ciliogenesis during the process of vertebrate development.
Results:
We found that several Prdx genes were strongly expressed in multiciliated cells in developing Xenopus embryos, and their peroxidatic functions were crucial for normal cilia development. Depletion of Prdx5 increased levels of cellular reactive oxygen species (ROS), consequently leading to mitochondrial dysfunction and abnormal cilia formation. Proteomic and transcriptomic approaches revealed that excessive ROS accumulation on Prdx5 depletion subsequently reduced the expression level of pyruvate kinase (PK), a key metabolic enzyme in energy production. We further confirmed that the promotor activity of PK was significantly reduced on Prdx5 depletion and that the reduction in PK expression and its promoter activity led to ciliary defects observed in Prdx5-depleted cells.
Innovation:
Our data revealed the novel relationship between ROS and Prdx5 and the consequent effects of this interaction on vertebrate ciliogenesis. The normal process of ciliogenesis is interrupted by the Prdx5 depletion, resulting in excessive ROS levels and suggesting cilia as vulnerable targets of ROS.
Conclusion:
Prdx5 plays protective roles in mitochondria and is critical for normal cilia development by regulating the levels of ROS. The loss of Prdx5 is associated with excessive production of ROS, resulting in mitochondrial dysfunction and aberrant ciliogenesis.
Introduction
O
Prdxs are known to be widely expressed in human tissues. Currently, six Prdx genes have been identified in mammals, including humans. There are three different types of Prdxs, which are classified according to their catalytic active sites. Prdx6 is a one-cysteine Prdx that uses a single cysteine residue for catalytic activity. Prdx1–4 are two-cysteine Prdxs and use two cysteine residues for their peroxidatic and resolving reactions. Finally, Prdx5 is an atypical two-cysteine Prdx that uses two active cysteine residues in atypical locations. Prdx5 is known to protect cells from ROS by preferentially removing hydrogen peroxide (H2O2) and alkyl hydroperoxides (12, 16, 28). Other studies reported the localization of Prdx5 to different cellular organelles, that is, peroxisomes and mitochondria, and play a role in protecting them against neurological damage induced by excessive ROS (4, 17, 23, 38).
Our study reveals the novel interaction between peroxiredoxin5 (Prdx5) and reactive oxygen species (ROS) that are involved in the process of ciliogenesis, thoroughly characterizes the ciliary defects induced by Prdx5 depletion, and reveals the relationship between ciliogenesis and ROS levels. Our data show that cilia are key and vulnerable cellular targets that can be damaged by excessive ROS. Further, we show conclusive evidence that elevated ROS levels affect cilia formation by reducing pyruvate kinase levels, which has not been previously demonstrated.
We believe that our study is the first intensive report on ROS-mediated ciliary defects and their underlying mechanisms and that it will likely be of significant interest for future prospects.
Cilia are highly conserved cellular structures that are widely distributed from unicellular organisms to complex multicellular organisms, including humans, and they carry out a variety of functions (27). In mammals, there are mainly two types of cilia, which are categorized based on their structure and function.
Most cells in resting stages possess a single cilium stemming from the mother centriole. These monocilia are often known as primary cilia and are mainly involved in signal reception or sensing in various tissues. Multicilia constitute another type of cilia that are responsible for motility. A single multiciliated cell may produce hundreds of cilia on its apical surface. Multicilia mainly exist in mucociliary epithelia, such as the airway epithelium and fallopian tubes, in which they generate a mucus fluid-flow over the epithelium, either to protect the tissues of the airway or to facilitate the migration of fertilized eggs through the fallopian tubes, respectively (7). In addition to their functional differences, primary cilia and multicilia also possess structural differences. For example, the axonemes of primary cilia have nine doublets of microtubules (9 + 0), whereas those of multicilia have an extra pair of central microtubules (9 + 2).
Recent studies have shown that ROS such as H2O2 function not only as agents that cause damage to cellular components, that is, DNA, proteins, and lipids, but also as intracellular messengers in multiple signaling cascades (28). Although it is considerably well understood how excessive ROS transduce a damage response to cellular organelles such as the endoplasmic reticulum (ER), peroxisome, and mitochondria, the consequences of excessive ROS levels in cilia require more detailed investigations.
In this study, we investigated the roles of Prdxs in ciliogenesis during the process of embryonic development. We observed that several Prdx genes were highly expressed in multiciliated cells and that Prdx5 depletion resulted in excessive ROS production. Consequently, excessive ROS levels disrupted both normal mitochondrial function and the process of ciliogenesis. In addition, silencing of Prdx5 resulted in considerable upregulation of phosphorylation of several signaling molecules such as AKT, ERK, JNK, eIF2α, and p38, revealing that the depletion of Prdx5 induces mitochondrial dysfunctions and ROS-mediated cellular responses. Finally, our results provided compelling evidences that a reduction in pyruvate kinase (PK) levels might be the direct cause of ciliary defects in Prdx5-depleted cells.
Results
Prdx5 is predominantly expressed in multiciliated cells
To investigate the functions of Prdx genes during vertebrate development, we examined the expression profiles of all six members of the Prdx family in developing Xenopus embryos. Prdx genes displayed dynamic expression patterns in developing Xenopus embryos, as revealed by whole-mount in situ hybridization (WISH; Supplementary Fig. S1. Prdx1 and Prdx3 were mainly expressed in the brain and developing kidney (Supplementary Fig. S1) (8), whereas Prdx2 was expressed in the ventral blood island and brain (Supplementary Fig. S1). Prdx4 displayed a low level of expression in the kidney and eyes (Supplementary Fig. S1), and Prdx5 was predominantly expressed in multiciliated cells (Fig. 1A), whereas Prdx6 was expressed in migrating myeloid cells (Supplementary Fig. S1).
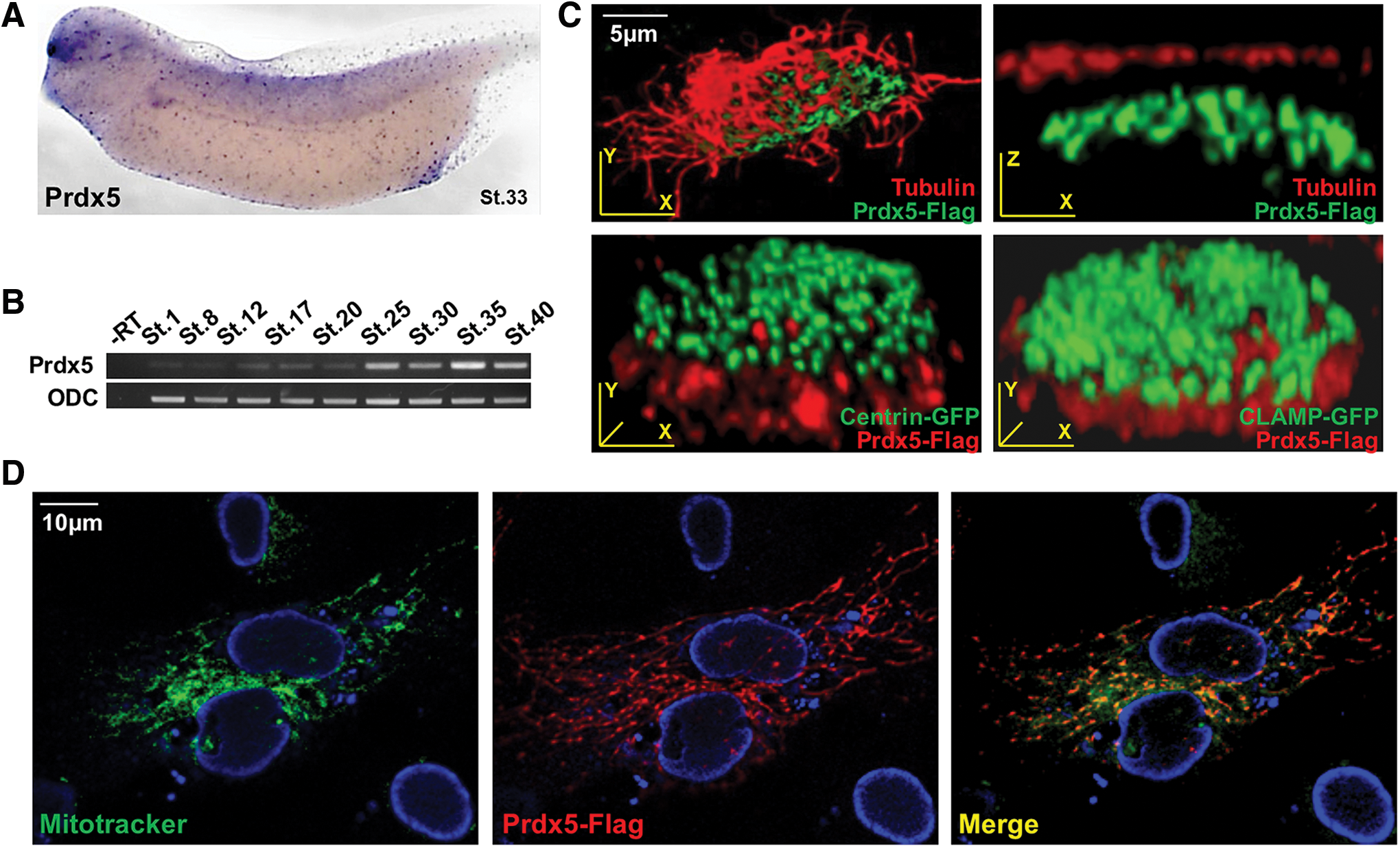
The mucociliary epithelium functions as the first line of defense against external contaminants or pathogens. In the airway epithelium of mammals, mucociliary cells line the airway to protect the respiratory system (37). In Xenopus, the mucociliary epithelium exists in the embryonic epidermis to protect developing embryos.
The Xenopus mucociliary epidermis is an emerging model system for studying the physiology of the mucociliary epithelium (7, 34). Therefore, we exploited the Xenopus embryonic epidermis to study the biological functions of Prdxs in multiciliated cells. Among the Prdx genes, Prdx1 and Prdx5 were strongly expressed in multiciliated cells. Then, we conducted knockdown studies to investigate the significance of Prdx1 and Prdx5 in cilia formation. Among Prdx1 and Prdx5, the knockdown of Prdx1 did not significantly affect multicilia formation (data not shown); therefore, Prdx5 was selected for further studies to evaluate its roles in multicilia formation. Prdx5 expression increased sharply at embryonic stage 25, at which multicilia began to form (Fig. 1B). The subcellular localization of Prdx5 in multiciliated cells was investigated by expressing Flag-tagged Prdx5 (Prdx5-Flag). Prdx5 localized to the mitochondria-rich sub-apical region (i.e., beneath the basal bodies) of multiciliated cells (Fig. 1C). Moreover, when Prdx5-Flag was expressed in hTERT-RPE1 cells that were subsequently stained with Mito-Tracker, Prdx5 localized to the mitochondria (Fig. 1D), consistent with the role of Prdx5 in mitochondrial function (6, 17, 20).
Prdx5 depletion causes severe malformation of multicilia, and the Prdx5 catalytic cysteines are required for cilia formation
We further investigated Prdx5 function in multiciliated cells by inhibiting Prdx5 expression via an injection of antisense morpholino oligonucleotides (Prdx5-MO) into two-cell-stage Xenopus embryos. Prdx5-MO effectively inhibited the translation of wild type (WT) Prdx5 messenger RNA (mRNA), whereas mRNAs with a four-base pair mismatch (Prdx5-4MT) were not affected by Prdx5-MO, indicating the specificity of Prdx5-MO for Prdx5 transcripts (Fig. 2A).
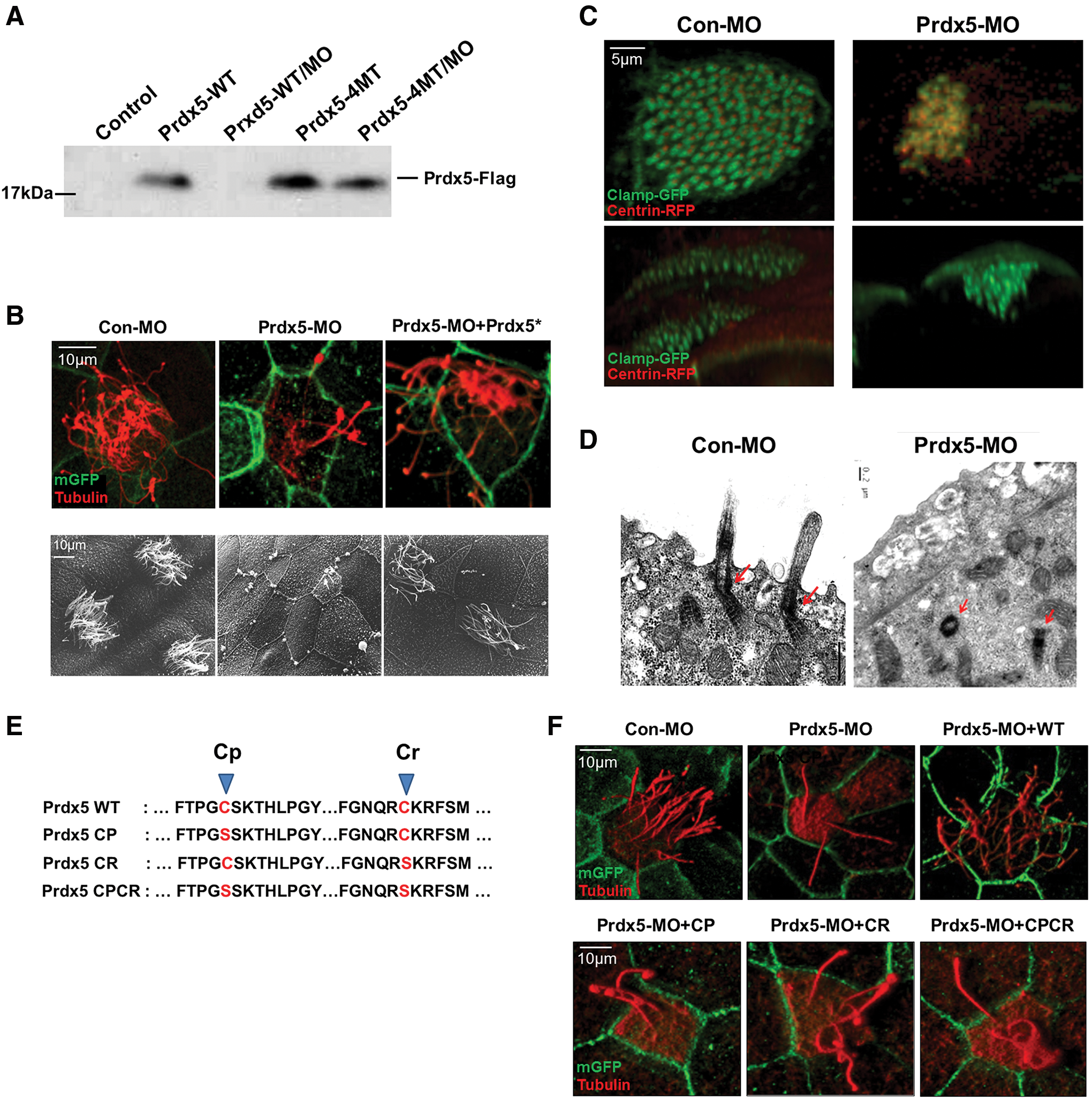
Next, we observed multicilia formation in Prdx5-MO-injected embryos (Prdx5 morphants) by immunofluorescent and electron microscopy (EM) imaging. Consistent with the results showing multiciliated cell-specific expression, MO-mediated depletion of Prdx5 in developing Xenopus embryos almost completely abolished multicilia formation in the embryonic mucociliary epidermis (Fig. 2B and Supplementary Fig. S5A). Moreover, the cilia-driven fluid-flow assay revealed that the velocity of the fluid-flow on the surface of the epidermis was significantly reduced in Prdx5 morphant embryos (Supplementary Fig. S2A, B; Supplementary Video S1A, B).
Ciliogenesis in multiciliated cells begins with the de novo formation of basal bodies, with hundreds of basal bodies docking to the apical surface of cells and initiating axoneme assembly (9, 25). Therefore, we analyzed the number and localization of basal bodies in multiciliated cells. Both confocal and EM imaging revealed that the numbers of basal bodies were similar between control and Prdx5 morphant embryos (data not shown); however, most of the basal bodies, which are marked by Centrin-GMP and CLAMP-RFP, in the cells of Prdx5 morphants failed to move apically for axoneme assembly (Fig. 2C, D).
Prdx5 catalyzes the reduction of H2O2, and two cysteine residues in the catalytic active site are critical for its reductase activity (16). To determine whether Prdx5 catalytic activity is necessary for ciliogenesis, we co-injected embryos with Prdx5-MO and Prdx5 catalytic mutants (in which the peroxidatic cysteine, resolving cysteine, or both cysteines were changed to serine; Fig. 2E). The embryos injected with Prdx5 mutants failed to rescue the ciliary defects (Fig. 2F) as compared with the embryos injected with WT Prdx5, which affectively reversed the ciliary defects. These data indicate that Prdx5 is critical for multicilia formation, presumably by affecting ciliogenesis after de novo formation of basal bodies. In addition, our data indicate that the catalytic activity of Prdx5 is necessary for cilia formation.
Prdx5 is necessary for primary cilia formation
We next examined the role of Prdx5 in primary cilia formation during the embryonic development. Xenopus embryos are known to generate primary cilia in the luminal surface of the neural tube. We observed that Prdx5 knockdown in Xenopus embryos disrupted primary cilia formation in the neural tube (Fig. 3A). In addition, Prdx5 depletion inhibited the convergent extension of neural tissue and resulted in a shortened body axis in neurula stage embryos (Fig. 3D, E), which was a previously reported consequence of defective cilia formation during embryonic development (24, 29).
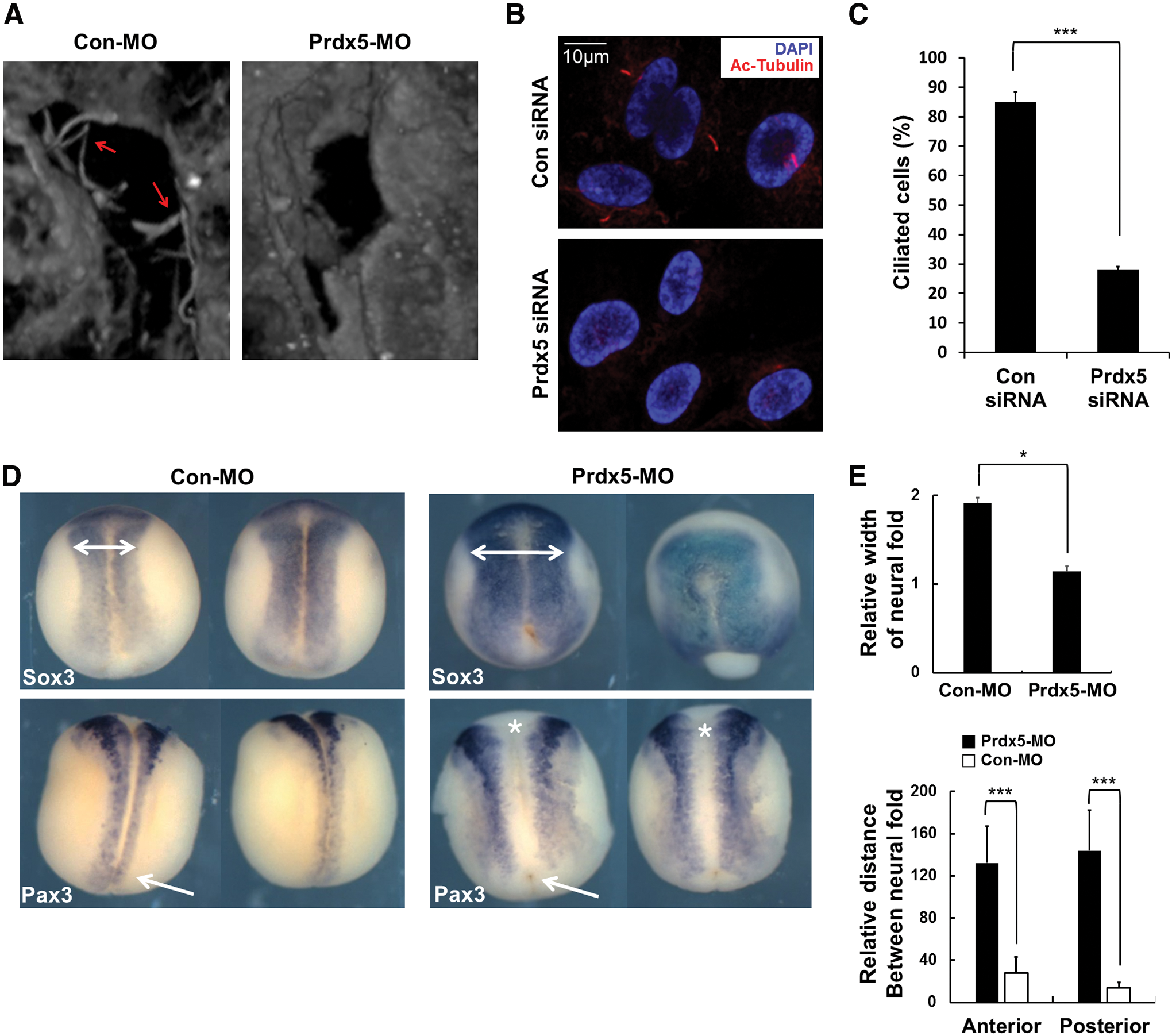
We further investigated the role of Prdx5 in primary cilia formation in hTERT-RPE1 cells after transfection with Prdx5 small interfering RNA (siRNA). Prdx5 knockdown in hTERT-RPE1 cells effectively decreased Prdx5 expression (Supplementary Fig. S3A), and Prdx5 depletion significantly reduced the number of ciliated cells (Fig. 3B, C). These data indicate that Prdx5 is necessary for primary cilia formation and that the function of Prdx5 in ciliogenesis is conserved among vertebrates.
Ciliary defects on Prdx5 depletion results from excessive ROS
Next, we sought to understand the molecular mechanisms underlying the ciliary defects observed after Prdx5 depletion. Prdx5 is known to eliminate mitochondrial ROS by scavenging peroxides, such as H2O2, organic hydroperoxides, and peroxynitrite, all of which can irreversibly harm cellular organelles and cells (3, 12). We speculated that an excess of ROS resulting from Prdx5 depletion may impair the ciliogenesis process. Indeed, Prdx5 knockdown significantly increased ROS levels in hTERT-RPE1 cells, and MitoQ, which is known to reduce ROS levels (15), effectively reduced the corresponding elevated ROS levels (Fig. 4A).
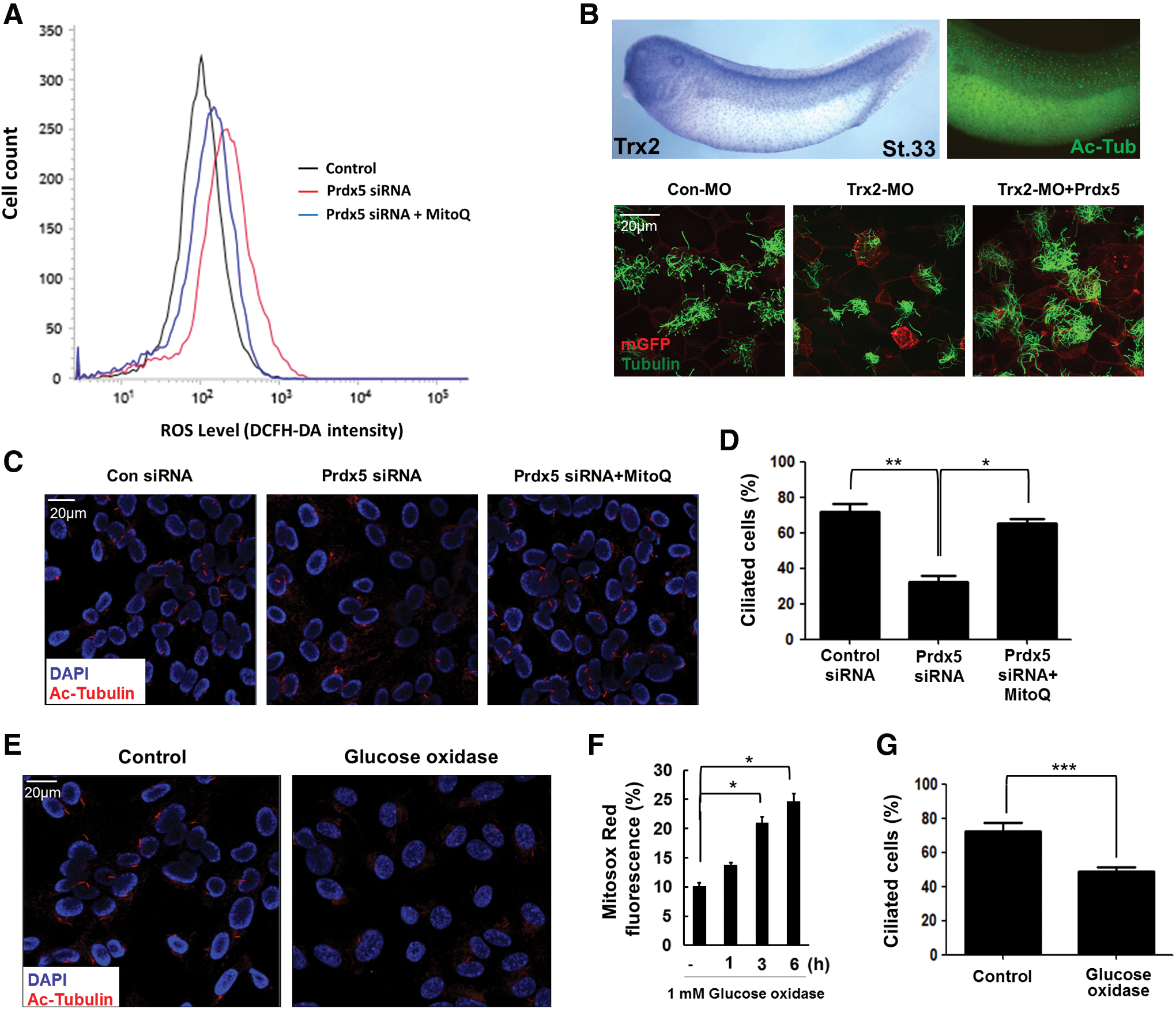
Since Prdxs function in Prdx/Trx (thioredoxin) systems to remove excessive H2O2 and need to be reduced by Trx for the next round of antioxidant reactions (2), we speculated that depletion of Trx would also cause ciliary defects and that these ciliary defects would be rescued by the overexpression of Prdx5. To test this speculation, we observed the expression of Trx2 in multiciliated cells and found that it was highly expressed in multiciliated cells. Moreover, depletion of Trx2 caused similar ciliary defects as Prdx5 depletion (Fig. 4B). We also observed that these defects were rescued by the overexpression of Prdx5, induced by injecting Prdx5 mRNA (Fig. 4B). Our findings suggest that Trx, in conjunction with Prdx5, has a significant role in cilia formation.
To confirm that the enhanced ROS levels were the major cause of ciliary dysfunction, we artificially quenched ROS by treating hTERT-RPE1 cells with MitoQ (Fig. 4A).We found that the percentage of ciliated cells was restored by MitoQ treatment of Prdx5-depleted hTERT-RPE1 cells (Fig. 4C, D). Further, artificial induction of excessive ROS levels via treatment with glucose oxidase (GOX) (1) inhibited primary cilia formation in hTERT-RPE1 cells (Fig. 4E–G). Taken together, our results indicate that excessive ROS are the major underlying causes of the ciliary defects observed after Prdx5 depletion. Thus, we suggest that Prdx5 regulates mitochondrial ROS levels and plays a significant role in cilia formation.
Prdx5 depletion causes mitochondrial dysfunction
We further investigated the underlying mechanism by which excessive ROS disrupt ciliogenesis in hTERT-RPE1 cells. Prdx5 mainly localizes to the mitochondria and protects them from oxidative damage (3). This prompted us to examine mitochondrial function after Prdx5 depletion. Using the membrane potential indicator JC-1, we observed that the mitochondrial membrane potential and adenosine triphosphate (ATP) production were dramatically reduced after Prdx5 knockdown in hTERT-RPE1 cells (Fig. 5A, B). Prdx5 knockdown also induced outer mitochondrial membrane blebbing (Fig. 5C, D) and reduced the number of mitochondria in Prdx5 morphant embryos (Fig. 5E). Further, the mitochondrial defects observed in Prdx5 morphants were recovered by a co-injection of Prdx5 mRNA with Prdx5-MO (Fig. 5C–E).
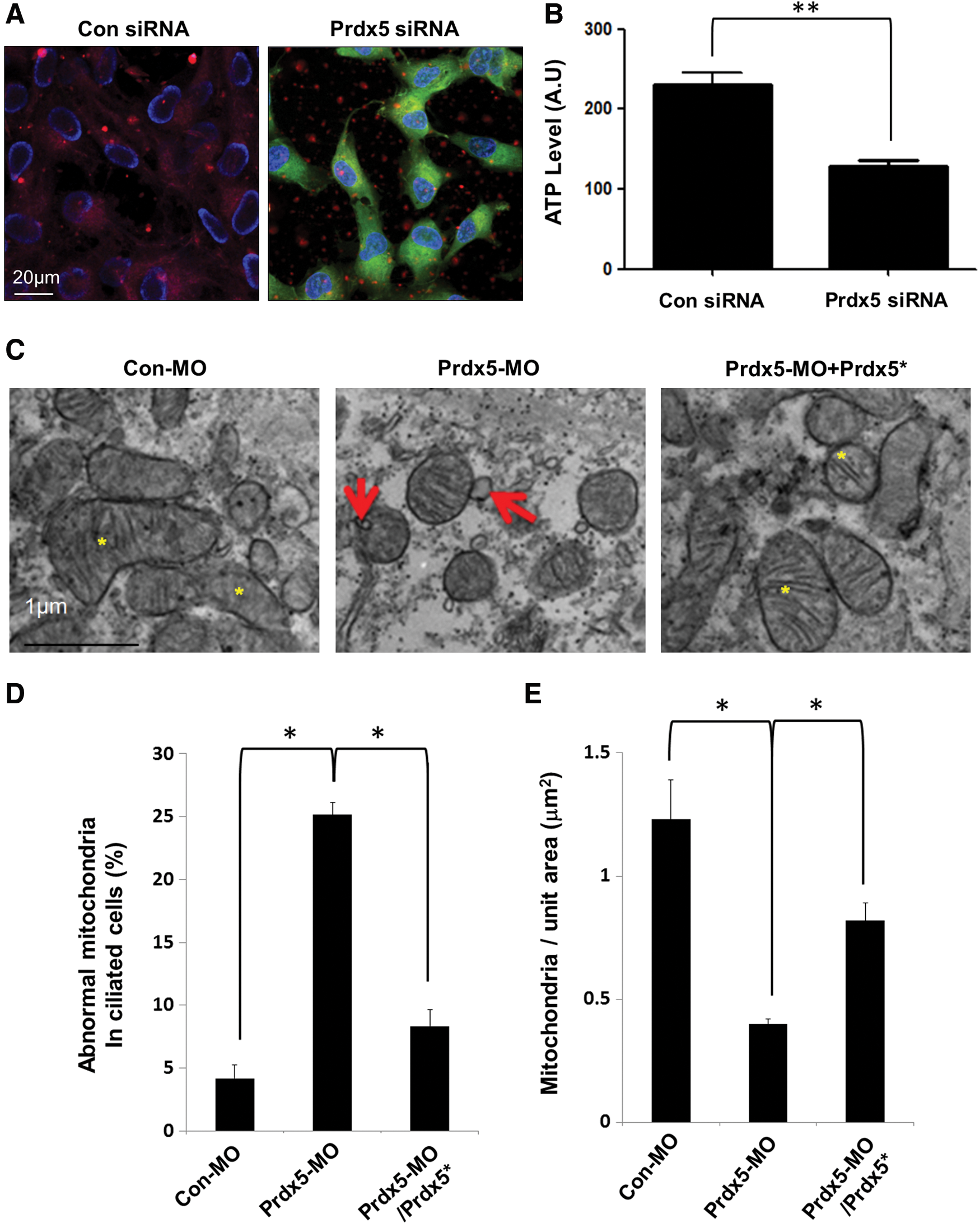
The induction of mitochondrial dysfunction by excessive ROS is known to trigger several downstream signaling cascades, such as apoptosis, autophagy, and stress responses (14, 31). Therefore, we examined several downstream signaling pathways affected by mitochondrial dysfunction in Prdx5-depleted cells. Prdx5 depletion did not affect the activation of the cell death signaling, such as AMPK, caspase-3, or RIP1/3 (Fig. 6).
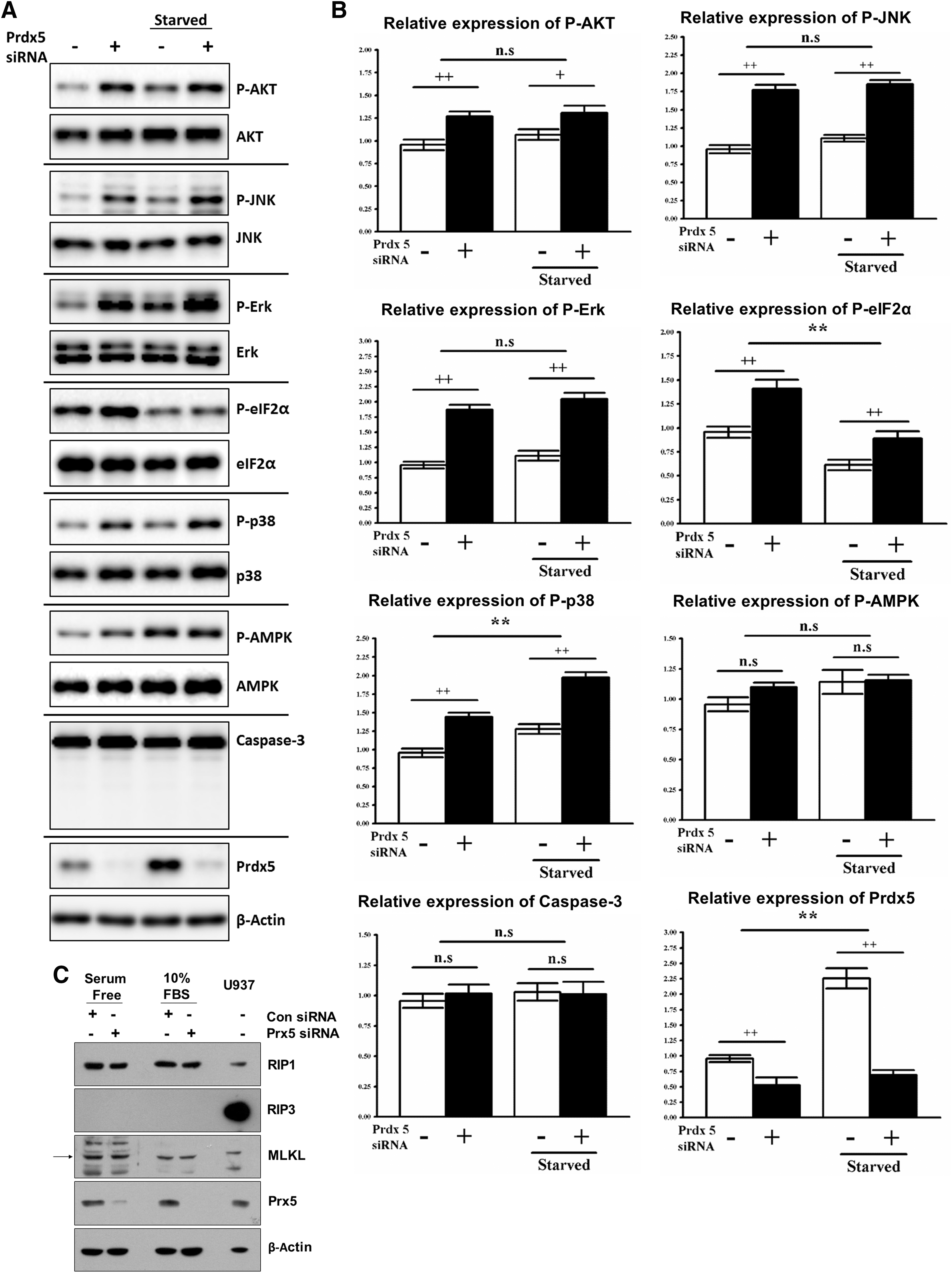
However, Prdx5 siRNA transfection significantly upregulated the phosphorylation of several signaling molecules, such as AKT, ERK, JNK, eIF2α, and p38 (Fig. 6A, B, and Supplementary Fig. S5D). Among these changes, eIF2α phosphorylation was previously shown to globally reduce general translation as a mitochondrial stress response (40), and this is consistent with our finding that loss of Prdx5 induces significant mitochondrial dysfunction. In addition, AKT, ERK, JNK, and p38 have been shown to be activated by excessive ROS such as H2O2 (36, 39). Overall, these data indicate that Prdx5 depletion significantly disrupts mitochondrial function and induces ROS-mediated cellular responses.
Reduction in PK levels induced by excessive ROS results in ciliary defects
Mitochondrial dysfunction and the consequent elevation in ROS have been shown to alter the global metabolic status and transcriptome (19). We further investigated direct causes of cilia loss in Prdx5-depleted cells. To this end, we assessed the transcriptomic and proteomic profiles of Prdx5 morphant embryos (Fig. 7A). We focused on differentially expressed genes involved in mitochondrial ATP production and identified dozens of genes downregulated in multiciliated cells after Prdx5 knockdown (Supplementary Table S1). We further analyzed the expression patterns of candidate genes by performing WISH analysis.
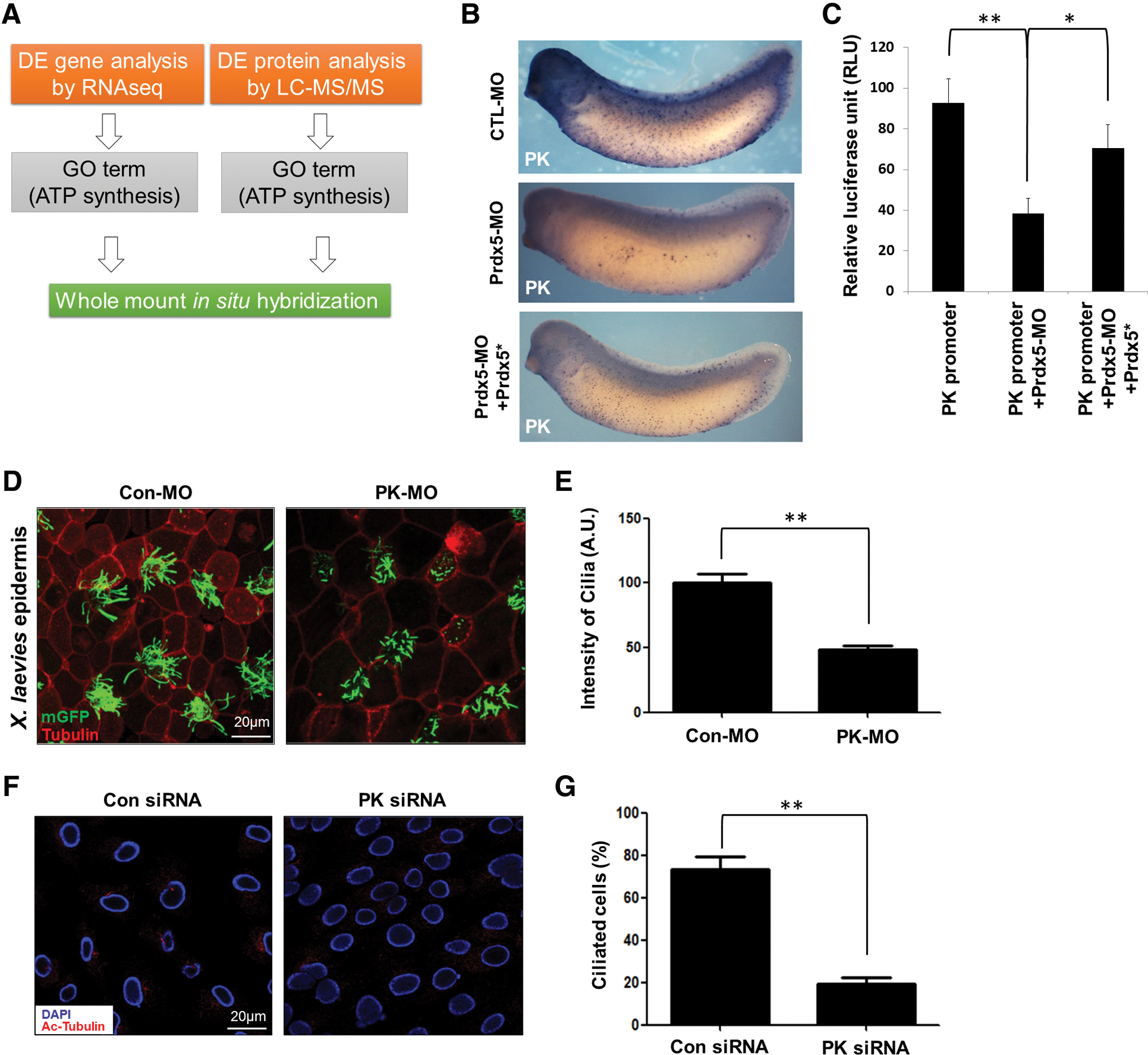
The muscle-specific PK gene, which is predominantly expressed in multiciliated cells, was among those downregulated in the morphants (Fig. 7B). PK is a critical enzyme that converts phosphoenolpyruvate to pyruvate, thereby generating ATP in the glycolytic pathway. WISH analysis further confirmed that PK expression, indeed, decreased sharply in the multiciliated cells of Prdx5 morphants (Fig. 7B). To confirm the reduction in PK transcription in Prdx5 morphants, we cloned the promoter region of PK and performed a promoter assay. Consistent with the WISH analysis, the activity of the PK promoter was significantly reduced in the Prdx5 morphants (Fig. 7C). The reduction in PK expression and its promotor activity was rescued by a co-injection of Prdx5 mRNA with Prdx5-MO (Fig. 7B, C). Further, the knockdown of PK in Xenopus embryos and hTERT-RPE1 cells severely disrupted multicilia (Fig. 7D, E) and primary cilia formation, respectively (Fig. 7F, G), indicating that PK is critical for ciliogenesis. We also confirmed that MitoQ treatment did not rescue defects in ciliogenesis in PK-depleted cells (Supplementary Fig. S4C, D), suggesting that PK acts downstream of ROS signaling in ciliogenesis.
A reduction in PK expression causes a reduction in pyruvate levels, and this not only affects ATP production through mitochondrial oxidative phosphorylation but may also affect the process of glycolysis by increasing NADH levels in the cytoplasm, which is necessary for the lactate formation. Thus, we speculated that impairments in glycolysis may cause additional energy crises in Prdx5-depleted cells. To rule out this possibility, we assessed the levels of NADH in Prdx5-depleted cells and found that NADH levels were not significantly different from those in controls (Supplementary Fig. S3C). These data indicate that glycolysis is not significantly affected by the reduction in PK and that the mitochondrial dysfunction and consequent increase in ROS is the primary cause of abnormal ciliogenesis, rather than impairment in glycolysis.
Discussion
Prdx5 is well known for its antioxidant effects, which are exerted by scavenging H2O2 and alkyl hydroperoxides in mammalian cells and Xenopus embryos (12). It plays significant protective roles in neurological disorders and in balancing the amount of ROS produced in cells, but its roles in vertebrate development have not yet been fully elucidated.
In this study, we systematically investigated the expression patterns of Prdx genes in developing Xenopus embryos (Supplementary Fig. S1) and found that multiciliated cells expressed multiple members of the Prdx family, that is, Prdx1 and Prdx5 (Supplementary Fig. S1). Cilia are important organelles that are required for a variety of developmental processes and are involved in various signaling pathways (22). Among the six Prdx genes, only knockdown of Prdx5 resulted in the failure of multicilia formation (Fig. 2B). Further research into the causes of the ciliary defects observed in Prdx5-depleted embryonic multicilia revealed that the basal bodies in multiciliated cells failed to properly dock to the apical surface to generate ciliary axonemes (Fig. 2C, D).
In addition to its role in multicilia, Prdx5 was also required for primary cilia formation. Prdx5 dysfunction in primary cilia formation is known to inhibit convergent extension and result in a shortened body axis (24, 29). Consistent with these previous reports, our results showed that knockdown of Prdx5 resulted in a shortened body axis in neurula stage embryos and blocked their convergent extension (Fig. 3D, E). Moreover, we showed that siRNA-mediated knockdown of Prdx5 significantly reduced the number of cilia in hTERT-RPE1 cells (Fig. 3B, C).
Prdx5 is an atypical two-cysteine Prdx protein, and both the peroxidatic and resolving cysteines are critical for its catalytic activities (16). In this study, we investigated the significance of both the peroxidatic and resolving cysteines in normal cilia formation by co-injecting Prdx5 catalytic mutants with Prdx5-MO. These Prdx5 mutants failed to rescue the ciliary defects induced by Prdx5 knockdown, indicating that the peroxidatic activity of Prdx5 is necessary for multicilia formation (Fig. 2E, F).
Prdx5 regulates the amount of ROS produced in cells and protects them from ROS-induced damage (26). The depletion of Prdx5 leads to increased levels of cellular ROS and ciliary defects. Our data showed that ciliary defects and enhanced ROS levels in Prdx5-depleted cells are directly related, that is, Prdx5 depletion leads to an excess of ROS, which impairs normal ciliogenesis. Removal of ROS by MitoQ treatment efficiently reversed the ciliary defects observed in Prdx5-depleted hTERT-RPE1 cells (Fig. 4C, D). Thus, in addition to the other known targets of ROS, such as the mitochondria, ER, and peroxisome, our results clearly suggest that the cilium is a vulnerable target of excessive ROS.
Prdx5 is widely distributed in the cytosol, mitochondria, nucleus, and peroxisomes, but the long form of Prdx5 is preferentially located in the mitochondria (16). Since the mitochondria are the vital source of intracellular ROS production, we focused on the antioxidant properties of Prdx5 in the mitochondria of multiciliated cells.
Our findings present compelling evidence that the major function of Prdx5 in multiciliated cells is to protect the mitochondrial function from the oxidative damage caused by ROS. The depletion of Prdx5 led to severe malformation of mitochondria and significantly disrupted the mitochondrial membrane potential and ATP production (Fig. 5). Consistent with its role in protecting the mitochondria, we confirmed that Prdx5 depletion further transduces a signal through p38 and eIF2α phosphorylation, which downregulates global translation as a mitochondrial stress response (Fig. 6A, B). Prdx5 depletion not only affected p38 and eIF2α phosphorylation but also significantly increased the phosphorylation of AKT, JNK, and ERK, which were previously shown to be activated by ROS, indicating that Prdx5 depletion causes mitochondrial dysfunction and elevates ROS (Fig. 6A, B).
Ciliogenesis is extremely important in vertebrate development, and abnormalities in ciliary development can lead to serious congenital disorders (11). Since our data showed that increased ROS production is a contributing factor in ciliary dysfunction in Prdx5-depleted cells, we analyzed the proteomic and transcriptomic changes induced by Prdx5 depletion for further insight into the direct causes of ciliary defects on Prdx5 depletion. Based on these analyses, it was clear that levels of the muscle-specific PK were significantly reduced on Prdx5 depletion, and this was further supported by WISH analysis showing no detectable expression in multiciliated cells (Fig. 7B). Indeed, PK expression was almost completely abolished in the multiciliated cells of Prdx5 morphant embryos. The activity of the PK promoter was significantly less responsive under Prdx5-depleted conditions (Fig. 7C). Moreover, PK depletion recapitulated the ciliary defects observed in Prdx5-depleted embryos.
The molecular mechanism by which PK affects cilia formation and function is currently not well understood. However, it has been shown that PK, along with other glycolytic enzymes, is present in the axoneme of the Chlamydomonas flagellum (21). An in-depth analysis is necessary to further elucidate the exact roles and mechanisms of metabolic enzymes involved in cilia formation.
In conclusion, we suggest that in addition to its protective role as an antioxidant in cells, Prdx5 is also involved in vertebrate development by significantly influencing the process of ciliogenesis. Moreover, the intact peroxidatic function of Prdx5 is critical for proper ciliary development.
Materials and Methods
In vitro fertilization
This study was performed in strict accordance with the guidelines of the Animal Care and Use Committee and in agreement with international laws and policies (National Institutes of Health Guide for the Care and Use of Laboratory Animals, publication no. 85-23, 1985). The Institutional Animal Care and Use Committee (IACUC) of the Ulsan National Institute of Science and Technology approved the experimental use of amphibians (approval no. UNISTIACUC-16-14). All members of our laboratory attended educational and training courses on the proper care and use of experimental animals. Adult Xenopus laevis obtained from the Korean Xenopus Resource Center for Research were housed at 18°C under conditions of 12 h light/12 h dark in containers recommended by the IACUC of the Ulsan National Institute of Science and Technology. Ovulation was induced in adult female amphibians by the subcutaneous injection of 1000 IU of human chorionic gonadotropin (Sigma-Aldrich, St. Louis, MO). The next day, the females were allowed to lay eggs in 60-mm plastic dishes. After washing three times in 0.1 × modified Barth's solution (MBS; 88 mM NaCl, 5 mM Hepes, 2.5 mM NaHCO3, 1 mM KCl, 1 mM MgSO4, and 0.7 mM CaCl2, pH 7.8), the eggs were fertilized in the same solution. Thereafter, testes were obtained from adult male amphibians and minced in 3 to 4 mL of ice-cold 1 × MBS. After successful fertilization, the jelly coat was removed by gently swirling the embryos in a 2%
Plasmids and reagents
A complementary DNA clone encoding full-length Prdx5 was obtained from ATCC (GenBank ID: NM_001092111.1). We designed a translation-blocking Prdx5-MO based on the sequence in the Xenbase database. We purchased the Prdx5-MO from Gene Tools (5′-ATGATACCGGGATACGAAGAGCCAT-3′). Flag-tagged Prdx5 fusion constructs (WT, mutant constructs; C47S, C151S, and C47S/C151S) were generated by polymerase chain reaction (PCR) and subcloned into the pCS2+ vector.
Embryo injections
Synthetic capped mRNAs were prepared by using the SP6 mMessage mMachine Kit (Ambion, Carlsbad, CA) and subsequently microinjected into embryos as previously described (18). To rescue the Prdx phenotype, a Prdx5-MO-resistant mRNA containing 4MT was synthesized. MOs (20–60 ng) and/or mRNAs were microinjected into blastomeres of two-cell-stage embryos.
Whole-mount in situ hybridization
To analyze gene expression in developing embryos in situ, WISH was performed as previously described (35). Briefly, embryos at the indicated stage were fixed in MEMFA buffer (4% paraformaldehyde in 0.1 M MOPS, 1 mM MgSO4, and 2 mM EGTA, pH 7.4) for 2 h at room temperature. The antisense-RNA probes were labeled with digoxigenin-UTP by using the mMessage mMachine Kit, and the fixed embryos were incubated overnight with the antisense probes in the hybridization buffer at 55°C. The probes were detected with an alkaline phosphatase-labeled anti-digoxigenin antibody (1:1000; Roche, Basel, Switzerland) and BM purple dye.
Cilia-driven fluid-flow assay
Control and Prdx5 morphant embryos were anesthetized with benzocaine and placed in agarose-coated plastic dishes. Fluorescently labeled polystyrene beads with a diameter of 10 μm (Molecular Probes, Eugene, OR) were added on top of the embryos, and real-time images were recorded by time-lapse microscopy and cellSens Dimension Software (Olympus, Tokyo, Japan). The directionality and velocity of the beads were measured by merging 10 consecutive images.
Cell culture
hTERT-RPE1 cells were cultured in DMEM-F12 (Welgene, Seoul, Korea) containing 10% fetal bovine serum (Welgene) and 100 U/mL streptomycin/penicillin at 37°C in a humidified atmosphere of 5% carbon dioxide in air.
Western blot analysis
Whole embryos were homogenized in lysis buffer (137 mM NaCl, 20 mM Tris–HCl, 1% Nonidet-P40, and 10% glycerol, pH 8.0) containing 1 mM phenylmethylsulfonyl fluoride, 5 mM sodium orthovandate, and 1 × protease inhibitor cocktail (Sigma-Aldrich). Lysates were heated at 95°C in gel loading buffer for 5 min and electrophoresed on a 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis gel. Fusion proteins were detected with anti-flag-horseradish peroxidase (HRP)-conjugated (Sigma-Aldrich) and anti-HA-HRP conjugated (Roche) antibodies. Immunoreactive bands were detected by using the HyGLO Kit (Denville Scientific, South Plainfield, NJ).
Anti-RIP3 and anti-MLKL antibodies were purchased from Abcam (Cambridge, MA). Anti-RIP1 antibody was purchased from BD Transduction Laboratories (San Jose, CA). Anti-Prx5 antibody was purchased from Ab Frontier (Seoul, Korea). Anti-actin antibody was obtained from Sigma-Aldrich.
Immunofluorescent staining and confocal microscopy
The embryos at stage 26 were stained immunofluorescently according to previously described documented standards (10) by using a polyclonal anti-acetylated tubulin (1:400; Sigma-Aldrich) or a monoclonal anti-flag (1:400; Applied Biological Materials, Richmond, Canada) primary antibody, followed by incubation with a Cy3-conjugated anti-rabbit (1:400; Jackson ImmunoResearch Laboratories, West Grove, PA) or a fluorescein isothiocyanate-conjugated anti-mouse (1:400; Invitrogen, Carlsbad, CA) secondary antibody. hTERT-RPE1 cells were immunostained by using an anti-acetylated tubulin (1:500) or an anti-flag (1:500) antibody. Cells were co-stained with Mito-Tracker, a mitochondria-specific probe (1:1000; Invitrogen), and 4′,6′-diamidino-2-phenylindole dihydrochloride (1:5000; Invitrogen). Images were acquired with the Zeiss LSM7 PASCAL confocal microscope (Oberkochen, Germany) and analyzed with Image J and Adobe Photoshop Software.
Electron microscopy
To take EM imaging of embryos, the embryos were fixed at stage 26 in 2% glutaraldehyde and 2% paraformaldehyde in 0.1 M NaH2PO4, pH 7.4 for 2 h at 4°C. After washing three times, the embryos were postfixed in 1% osmium tetroxide on ice for 2 h and then washed three times. After dehydration in a graded series of ethanol baths and infiltration in a 1:1 ratio of propylene oxide:epon, the embryos were embedded in Epon 812. Polymerization was carried out in resin at 70°C for 24 h. Ultrathin sections with a thickness of ∼70 nm were prepared with an MT-X ultramicrotome (RMC, Tucson, AZ) and collected on 100-mesh copper grids. After staining with 2% uranyl acetate for 7 min and lead citrate for 2 min, images were acquired with conventional transmission electron microscopy (TEM) (JEM-1400 Plus, at 120 kV) and Bio-HVEM (JEM-1000BEF, at 1000 kV) (Jeol, Tokyo, Japan).
Measurement of mitochondrial ROS levels
Mitochondrial ROS levels were determined by using Mitosox Red (Invitrogen). RPE-1 cells were treated with 1 mM GOX for the indicated periods, and then cells were stained with Mitosox Red for an additional 10 min. Then, cells were trypsinized and resuspended in phosphate buffered saline (PBS), and fluorescence was measured at specific time intervals with a flow cytometer (Becton-Dickinson, Franklin Lakes, NJ).
Flow cytometry
hTERT-RPE1 cells were trypsinized, washed with PBS, and fixed in 70% ethanol for 30 min. To analyze the cell cycle by flow cytometry, cells were washed, resuspended, and stained with 10 μg/mL propidium iodide in PBS. The BD FACSVerse System (BD Biosciences, San Jose, CA) was used for flow cytometry. Cells were also stained with DCFH-DA or JC-1 in PBS at 37°C for 30 min in the dark. The stained cells were washed and then analyzed by flow cytometry.
Transcriptome and proteome profiling
For transcriptome profiling, the animal caps of WT and Prdx5 morphant embryos were explanted and cultured until Nieuwkoop and Faber stage 26, and total RNA was extracted by using TRIzol Reagent. The strand-specific library construction with poly-A enrichment was followed by the manufacturer's instruction (Illumina, San Diego, CA), and sequencing was carried out with Illumina HiSeq 2500 with 100PE configuration. The expression analysis was performed as previously described (33).
For proteome profiling, the animal caps of WT and Prdx5 morphant embryos were explanted and cultured until developmental stage 26 and they were then lysed in 100 mM triethylammonium bicarbonate, pH 8.0. The proteins were then reduced in 10 mM dithiothreitol at 60°C for 20 min and alkylated in 55 mM iodoacetamide at room temperature for 30 min, followed by trypsin digestion. The peptide fragments were analyzed by nano-ultra performance liquid chromatography (Waters, Milford, MA) and mass spectrometry using quadrupole-time-of-flight Premier (Waters). Peptides were separated by a 25 cm × 75 μm nanoACQUITY C18 column (Waters) using the LC system. Mobile phase A was composed of water with 0.1% FA (formic acid), and mobile B phase was composed of 0.1% FA in acetonitrile. The mass spectrometer was operated in the data-dependent mode, automatically switching between MS (mass spectrometry) and MS/MS acquisition modes. The method included a full sequential MS scan (m/z 150–1600, 0.6 s) and five MS/MS scans (m/z 100–1990, 0.6 s/scan) of the five most intense ions present in the full-scan mass spectrum. The spectra were analyzed with Mascot Software (version 2.2.1; Matrix Science, Boston, MA), and the identities of the peptides were determined by searching the Swiss-Prot database (version 51.6; 257,964 sequence entries). The search parameters were as follows: X. laevis for taxonomy, fragment ion mass tolerance of 0.5 Da, and parent ion tolerance of 0.1 Da. Two missed cleavages were allowed for the trypsin digestion. Carbamidomethylation of the cysteines and oxidation of methionines were considered as variable modifications. After searching the GO term “GO:0006754,” we selected proteins involved in the production of ATP in mitochondria. The proteins identified by this process are listed in Supplementary Table S1. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD009323.
Statistical analysis
Data from WISH and reverse transcriptase-PCR were analyzed by using ImageJ software (National Institutes of Health * ). Results are presented as mean ± standard error (n = 5 with replicas for each sample). To determine statistical significance, results were analyzed by analysis of variance before Tukey post hoc test. A p-value of <0.05 was considered statistically significant.
Footnotes
Acknowledgments
This study was supported by grants from the National Research Foundation of Korea (NRF) (Grant No. NRF-2015R1A2A1A10053265) and the Ministry of Science, ICT, and Future Planning (MSIP) (Grant No. 2015R1A4A1042271), the Republic of Korea.
Author Disclosure Statement
All the authors declare no competing interests.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.