
Abstract
Aims:
NCF1, a subunit of the NADPH oxidase 2 (NOX2), first described the expression in neutrophils and macrophages and participated in the pathogenesis from various systems. However, there are controversial findings on the role of NCF1 in different kinds of kidney diseases. In this study, we aim to pinpoint the specific role of NCF1 in the progression of renal fibrosis induced by obstruction.
Results:
In this study, NCF1 expression was upregulated in kidney biopsies of chronic kidney disease patients. The expression level of all subunits of the NOX2 complex was also significantly increased in the unilateral ureteral obstruction (UUO) kidney. Then, we used wild-type mice and Ncf1 mutant mice (Ncf1m1j mice) to perform UUO-induced renal fibrosis. Results demonstrated that Ncf1m1j mice exhibited mild renal fibrosis but increased macrophages count and CD11b+Ly6Chi macrophage proportion. Next, we compared the renal fibrosis degree between Ncf1m1j mice and Ncf1 macrophage-rescued mice (Ncf1m1j .Ncf1 Tg-CD68 mice). We found that rescuing NCF1 expression in macrophages further alleviated renal fibrosis and decreased macrophage infiltration in the UUO kidney. In addition, flow cytometry data showed fewer CD11b+Ly6Chi macrophages in the kidney of the Ncf1m1j .Ncf1 Tg-CD68 group than the Ncf1m1j group.
Innovation:
We first used the Ncf1m1j mice and Ncf1m1j .Ncf1 Tg-CD68 mice to detect the role of NCF1 in the pathological process of renal fibrosis induced by obstruction. Also, we found that NCF1 expressed in different cell types exerts opposing effects on obstructive nephropathy.
Conclusion:
Taken together, our findings support that systemic mutation of Ncf1 ameliorates renal fibrosis induced by obstruction, and rescuing NCF1 in macrophages further alleviates renal fibrosis.
Introduction
Chronic kidney disease (CKD) is a major public health issue that can bring enormous economic and social burdens. The global prevalence of CKD is estimated to be 3%–18% (Bruck et al., 2016; Ebert et al., 2017; Mills et al., 2015; Okparavero et al., 2016; Stanifer et al., 2014; Zhang et al., 2012), and CKD has a high prevalence in the elderly (Mills et al., 2015). Regardless of the different causes of the disease, such as diabetes, hypertension, glomerulonephritis, renal vasculitis, ureteral obstruction, genetic alterations, autoimmune diseases, and drugs, the severity of CKD is mainly related to tubulointerstitial fibrosis (Wehrmann et al., 1990). Therefore, exploring the molecular mechanism for controlling renal fibrosis is compulsory.
Innovation
NCF1 is a subunit of the NADPH oxidase 2 (NOX2), mainly expressed in phagocytes, including macrophages. In addition, NOX2 was also identified in the kidney. Although there are research on the role of NCF1 and NOX2 in kidney disease, their results are controversial. Hence, we first used both the Ncf1m1j mice and Ncf1m1j.Ncf1 Tg-CD68 mice to detect the role of NCF1 in the pathological process of renal fibrosis. In addition, we found that NCF1 expressed in different cell types exerts opposing effects on renal fibrosis. These results could provide new strategies for chronic kidney disease targeting therapy.
Renal fibrosis is characterized by excessive deposition of extracellular matrix (ECM). ECM in the tubulointerstitium space is generated mainly by tissue-resident cells, including fibroblast, pericytes, and tubular epithelial cells (Djudjaj and Boor, 2019). Another characteristic of renal fibrosis is the secondary inflammatory response. Various immune cells, including macrophages, dendritic cells, mast cells, and T cells, can promote inflammation in the kidney. Still, the progression of fibrosis is driven by the macrophage transition from a proinflammatory phenotype to a profibrotic phenotype (Meng et al., 2014). A large number of macrophages accumulate in the glomerulus and interstitium in the process of renal fibrosis, and the degree of macrophage infiltration correlates with both the severity of renal damage and the extent of renal fibrosis (Eardley et al., 2008; Meng et al., 2014; Yang et al., 1998).
Macrophages can be polarized into different phenotypes and contribute to the progression of renal fibrosis (Wynn and Vannella, 2016). The heterogeneity of monocytes and macrophages can be defined according to their expression levels of the cell surface marker Ly6C (Menezes et al., 2016). The CD11b+Ly6Chi population is associated with the onset of renal injury and an increase in proinflammatory cytokines (Clements et al., 2016), whereas the CD11b+Ly6Clow population is more mature and transcribes genes consistent with a selective profibrotic function (Lin et al., 2009; Rodero et al., 2013). Therefore, pinpointing the molecular mechanism that regulates the macrophage phenotype transition in the progression of renal fibrosis is necessary and valuable in offering novel therapeutic targets for CKD.
NADPH oxidase 2 (NOX2), which could transfer electrons across biological membranes and then generate reactive oxygen species (ROS), is mainly expressed in phagocytes, including macrophages. In addition, NOX2 was also identified in the kidney (Bedard and Krause, 2007). Hence, it is attractive for us to analyze whether NOX2 participates in the progression of renal fibrosis by regulating macrophage phenotype transition. NCF1, also named P47phox, is a subunit of NOX2, and NOX2 activation depends on the recruitment and phosphorylation of NCF1. Therefore, pinpointing the function of NOX2 by regulating the expression of NCF1 is reasonable. However, the studies on NCF1s effect on kidney disease are controversial. Several research revealed that Ncf1-null mice showed reduced renal fibrosis and glomerular injury in mice with diabetic nephropathy (Liu et al., 2012), Adriamycin-mediated or partial renal ablation–mediated glomerular injury (Wang et al., 2015). On the contrary, there are still reports showing that mutation of Ncf1 aggravates pristane-induced lupus nephritis (Geng et al., 2022; Kienhofer et al., 2017).
Given the current controversial findings on the role of NCF1 in different kinds of kidney diseases and on the issue of whether NCF1 participates in the progression of renal fibrosis by regulating macrophage phenotype transition, we used wild-type mice (WT mice), Ncf1 mutant mice (Ncf1m1j mice), and Ncf1 macrophage-rescued mice (Ncf1m1j.Ncf1 Tg-CD68 mice) to establish unilateral ureteral obstruction (UUO)-induced renal fibrosis model. Compared with the WT group, Ncf1m1j mice exhibit minor fibrosis and decreased oxidative stress but more serious macrophage infiltration. However, the Ncf1m1j.Ncf1 Tg-CD68 mice tend to have less pathological manifestation than the Ncf1m1j group. Furthermore, fewer macrophages and fewer proinflammatory CD11b+Ly6Chi macrophages infiltrated the kidney of the Ncf1m1j.Ncf1 Tg-CD68 mice than the Ncf1m1j mice. These findings indicate that NCF1 expressed in macrophages has protected effects on renal fibrosis induced by obstruction via reducing macrophage infiltration and proinflammatory macrophage phenotype transition, but NCF1 produced by kidney resident cells may have opposite effects.
Results
NCF1 expression is upregulated in kidney biopsies of CKD patients
Compared with control samples, we found that kidney biopsies from IgA nephropathy, mesangial proliferative glomerulonephritis, membranoproliferative glomerulonephritis, lupus nephritis, and crescentic glomerulonephritis patients showed increased immunoreactivity of NCF1 in the tubulointerstitium space (Fig. 1A). However, the glomerular area of both CKD patients' renal biopsy and kidney tumor patients' tumor-adjacent tissues barely expressed NCF1 (Fig. 1A). The quantitation of NCF1 was measured by the percentage of NCF1 positive area, and the positive area of NCF1 was significantly increased in the CKD group compared with the control group (Fig. 1B). In addition to the increased expression of NCF1 in the tubular of the patients' biopsy, strong NCF1 expression is also detected in certain inflammatory cells (Fig. 1C). To confirm the cell type of these inflammatory cells, immunofluorescence was performed to detect the expression of NCF1 and CD68 in the patient's biopsy. Both NCF1 and CD68 were strongly expressed in the patient's section, and NCF1 colocalized with CD68, which is a well-known macrophage marker (Fig. 1D).
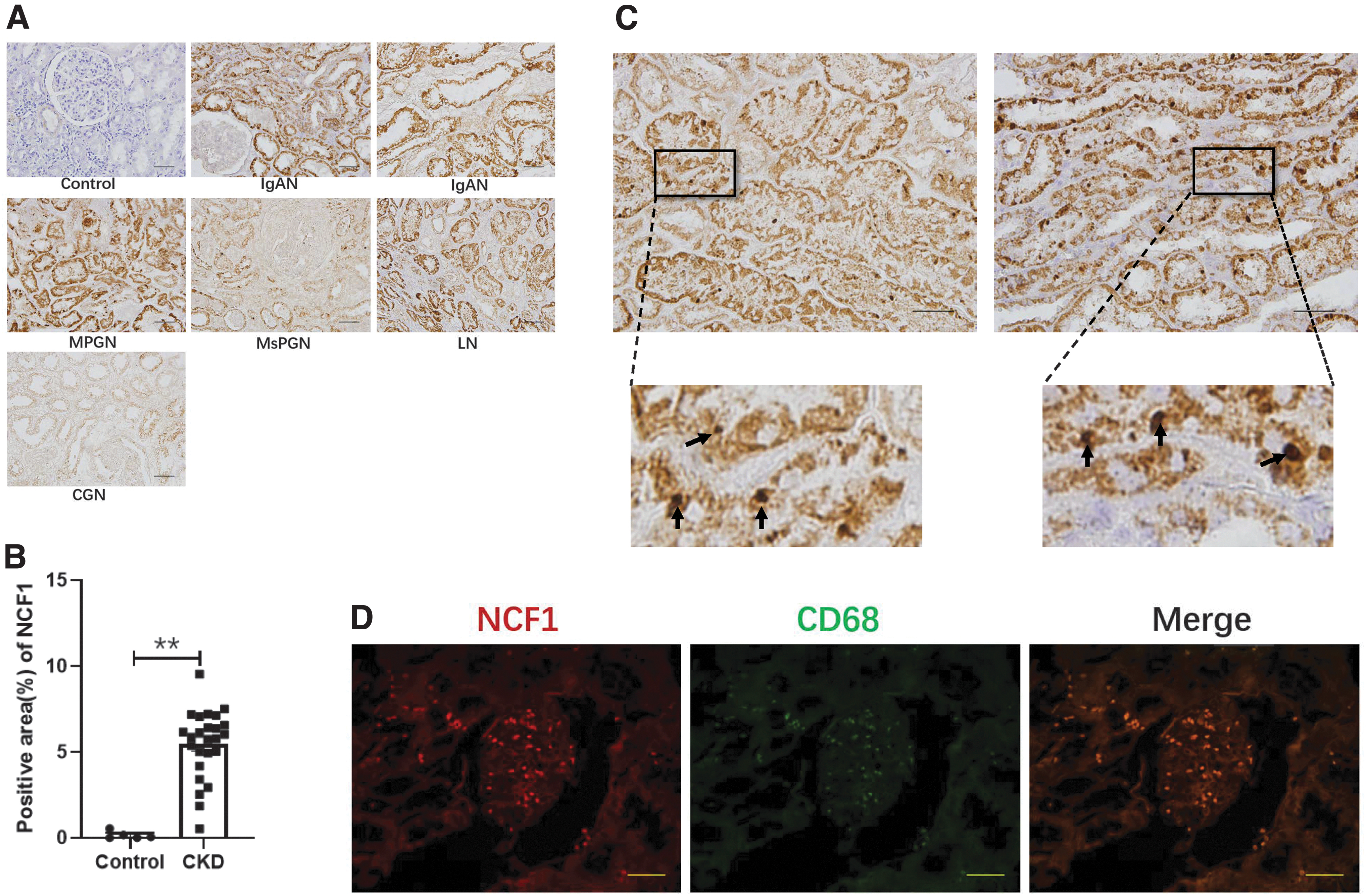
NCF1 expression is elevated in the UUO kidney
The RNA expression level of all subunits of the NOX2 complex, including Ncf1, Nox2, P22phox, P67phox, and P40phox, was significantly increased in the UUO kidney (Fig. 2A). In line with the mRNA expression result, NCF1 protein expression was upregulated in the kidney of UUO mice of 3, 7, and 14 days compared with the contralateral kidney (Fig. 2B). Immunohistochemical analysis was also used to detect the expression abundance and tissue localization of NCF1. Compared with the contralateral kidney, the expression of NCF1 was significantly upregulated in the UUO kidney (Fig. 2C), both in renal tubular cells and in tubulointerstitium space. Flow cytometry was used to analyze the expression of NCF1 in macrophages. Compared with the contralateral kidneys, more CD11b+CD68+ macrophages gathered in the UUO kidneys (Fig. 2D, E). Moreover, macrophages from the UUO kidneys had increased NCF1 expression than those from the contralateral kidneys (Fig. 2D, F). These data indicate that NCF1 is attributed to the progression of renal fibrosis via regulating macrophage function.
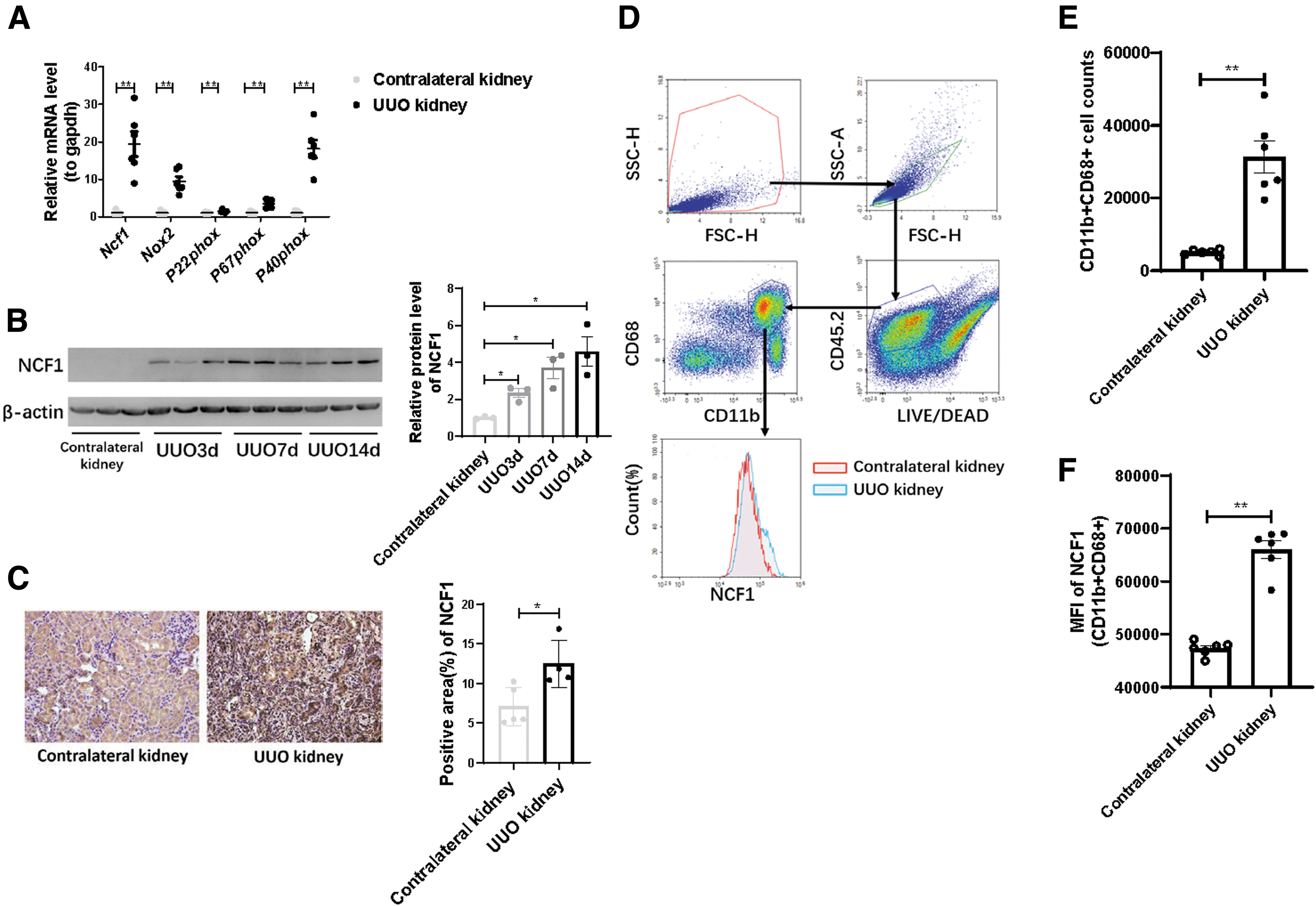
Ncf1 mutation alleviates renal fibrosis induced by obstruction
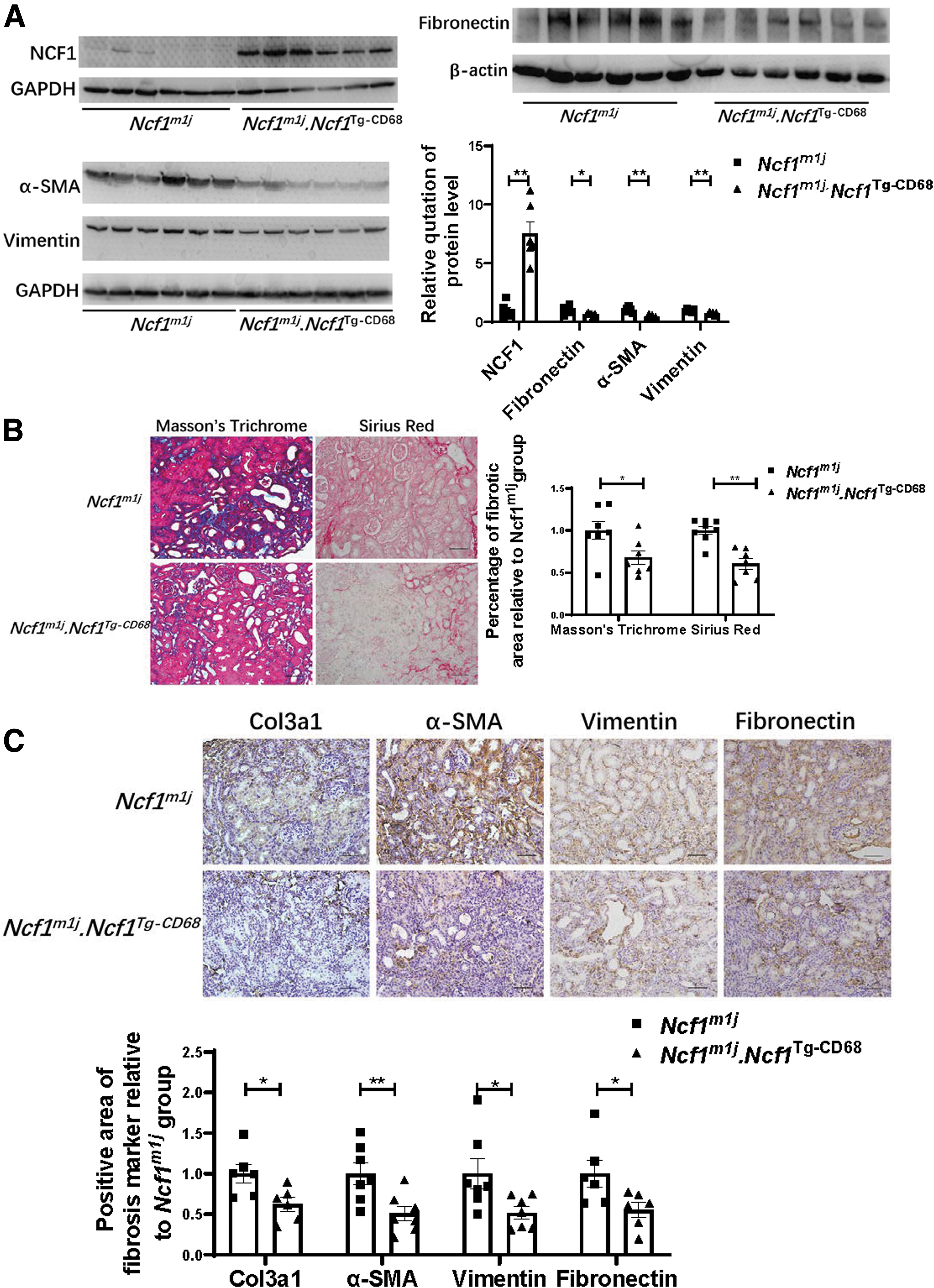
To further reveal the role of NCF1 in the progression of renal fibrosis, we primarily compare the degree of renal fibrosis between the WT group and the Ncf1m1j group. WT mice and Ncf1m1j mice were subjected to UUO surgery. Fourteen days after the operation, the kidneys were obtained and sent to the subsequent detection. The expression of NCF1 can barely be detected in the contralateral kidneys of both the WT mice and the Ncf1m1 j mice (Supplementary Fig. S1). Hence, before evaluating the severity of illness of these mice, we first verified the expression of NCF1 in the UUO kidneys of the WT mice and the Ncf1m1 j mice (Fig. 3A). Compared with the WT mice, Ncf1m1j mice's UUO kidney contained extremely small amount of NCF1 protein (Fig. 3A). Then, the protein expression level of fibronectin, α-SMA, and vimentin was detected by Western blotting. The results showed that these fibrosis markers all dramatically decreased in the Ncf1m1j group (Fig. 3A).
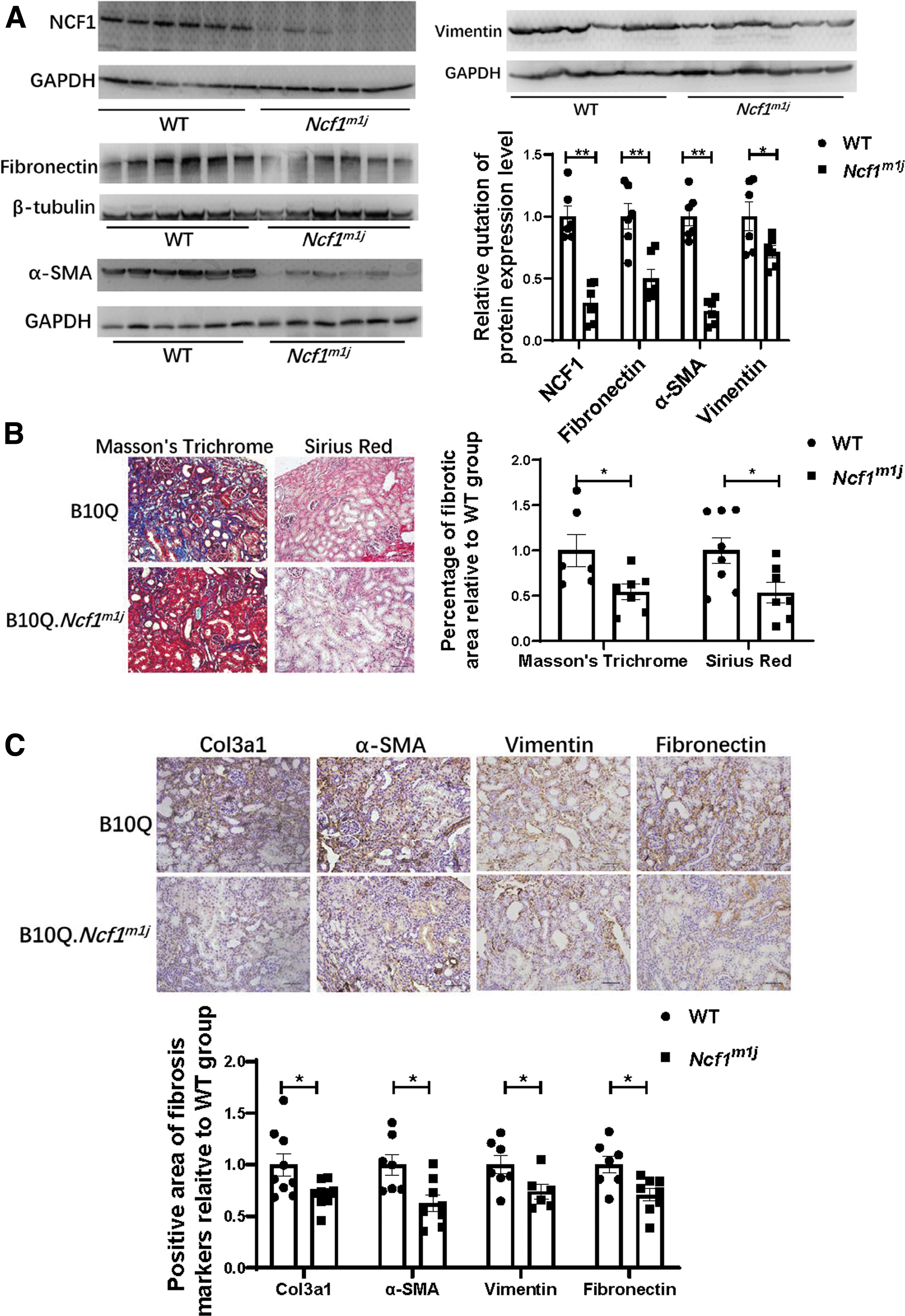
The degree of collagen deposition was evaluated by Sirius Red staining and modified Masson's trichrome staining (Fig. 3B). Similar to the Western blotting results, collagen deposition amount in the UUO kidneys of the Ncf1m1j mice was significantly less than that in the UUO kidneys of the WT mice (Fig. 3B). Immunohistochemical staining of Col3a1, α-SMA, vimentin, and fibronectin also proved less renal fibrosis level in the Ncf1m1j group than in the WT group (Fig. 3C). HK2 cell, a human kidney proximal tubular epithelial cell line, was used to further prove the role of NCF1 expressed at the cell in renal fibrosis. After knockdown of the NCF1 expression in HK2 cells by small interfering RNA (siRNA), TGF-β1 (10 ng/mL) was used to induce ECM production (Supplementary Fig. S2A–C). The mRNA expression level of NCF1 was significantly decreased after siRNA transfection (Supplementary Fig. S2A). NCF1 gene silencing significantly reduced the expression of ECM-related proteins, including fibronectin and Col1a1, induced by TGF-β1 in HK2 cells (Supplementary Fig. S2B, C).
Ncf1 mutation aggravates macrophage infiltration in the UUO kidney
The expression of NCF1 in the peritoneal macrophages of the WT mice and Ncf1m1j mice was detected by Western blotting. Significantly, NCF1 was barely expressed in Ncf1m1j mice's macrophages but strongly expressed in WT mice's macrophages (Fig. 4A). The macrophage counts in the UUO kidneys of the WT group and of the Ncf1m1j group were detected by immunohistochemical staining of F4/80. Contrary to the finding that Ncf1m1j mice showed a lighter renal fibrosis degree, the percentage of F4/80 positive area in the UUO kidney was significantly upregulated in the Ncf1m1j group compared with the WT group (Fig. 4B).
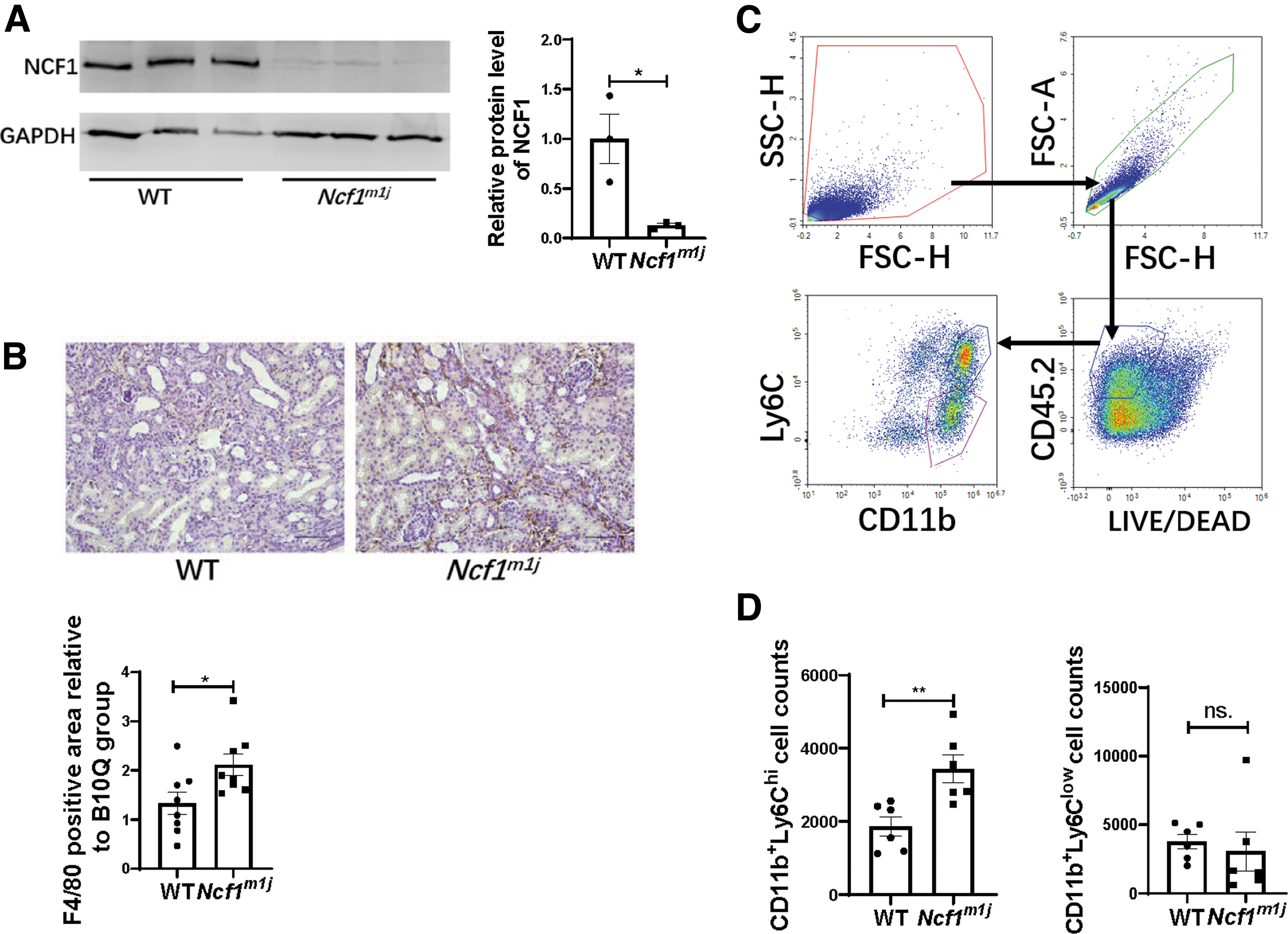
Because differential Ly6C expression was widely used to distinct monocyte/macrophage populations in various tissues (Clements et al., 2016; Gordon and Taylor, 2005; Lin et al., 2009; Movahedi et al., 2010; Ramachandran et al., 2012), we used flow cytometry to analyze the cell surface marker Ly6C to identify distinct monocyte/macrophage subsets in the kidneys of the WT mice and Ncf1m1j mice (Fig. 4C). Compared with the WT group, Ncf1m1j mice showed more CD11b+Ly6Chi macrophages gathering in the UUO kidneys (Fig. 4D).
Rescuing the expression of NCF1 in macrophages reduces the severity of renal fibrosis induced by obstruction
Ncf1 mutant mice and NCF1 macrophage-rescued mice were used to construct the UUO model to investigate the NCF1 role in macrophages during renal fibrosis. Before disease evaluation, NCF1 protein expression was first measured in the UUO kidneys of those two groups of mice, and the Ncf1m1j.Ncf1 Tg-CD68 mice were proved to be re-expression of NCF1 protein in their UUO kidneys (Fig. 5A). Then, the fibrosis degree of kidneys was assessed by Western blotting and histological assessment. The protein expression level of fibronectin, α-SMA, and vimentin decreased significantly in the Ncf1m1j.Ncf1 Tg-CD68 group compared with the Ncf1m1j group (Fig. 5A). Sirius Red staining (Fig. 5B), modified Masson's trichrome staining (Fig. 5C), and immunohistochemical staining of Col3a1 (Fig. 5C), α-SMA (Fig. 5C), vimentin (Fig. 5C), and fibronectin (Fig. 5C) results proved that Ncf1m1j.Ncf1 Tg-CD68 mice showed less renal fibrosis than Ncf1m1j mice.
Rescuing the expression of NCF1 in macrophage reduces the infiltration of macrophages
As demonstrated by Western blotting assay, the peritoneal macrophages of Ncf1m1j.Ncf1 Tg-CD68 mice expressed abundant NCF1 protein (Fig. 6A). Similar to the WT mice, the Ncf1m1j.Ncf1 Tg-CD68 mice showed significantly elevated NCF1 protein expression in the UUO kidneys compared with that in the contralateral kidneys (Fig. 6B). To prove that the elevated NCF1 expression in the UUO kidneys of Ncf1m1j.Ncf1 Tg-CD68 mice was owing to the infiltrating macrophages, flow cytometry assay was performed to assess the expression of NCF1 in the CD11b+CD68+ macrophages and other lived cells in the contralateral kidneys and UUO kidneys of Ncf1m1j.Ncf1 Tg-CD68 mice (Fig. 6C).
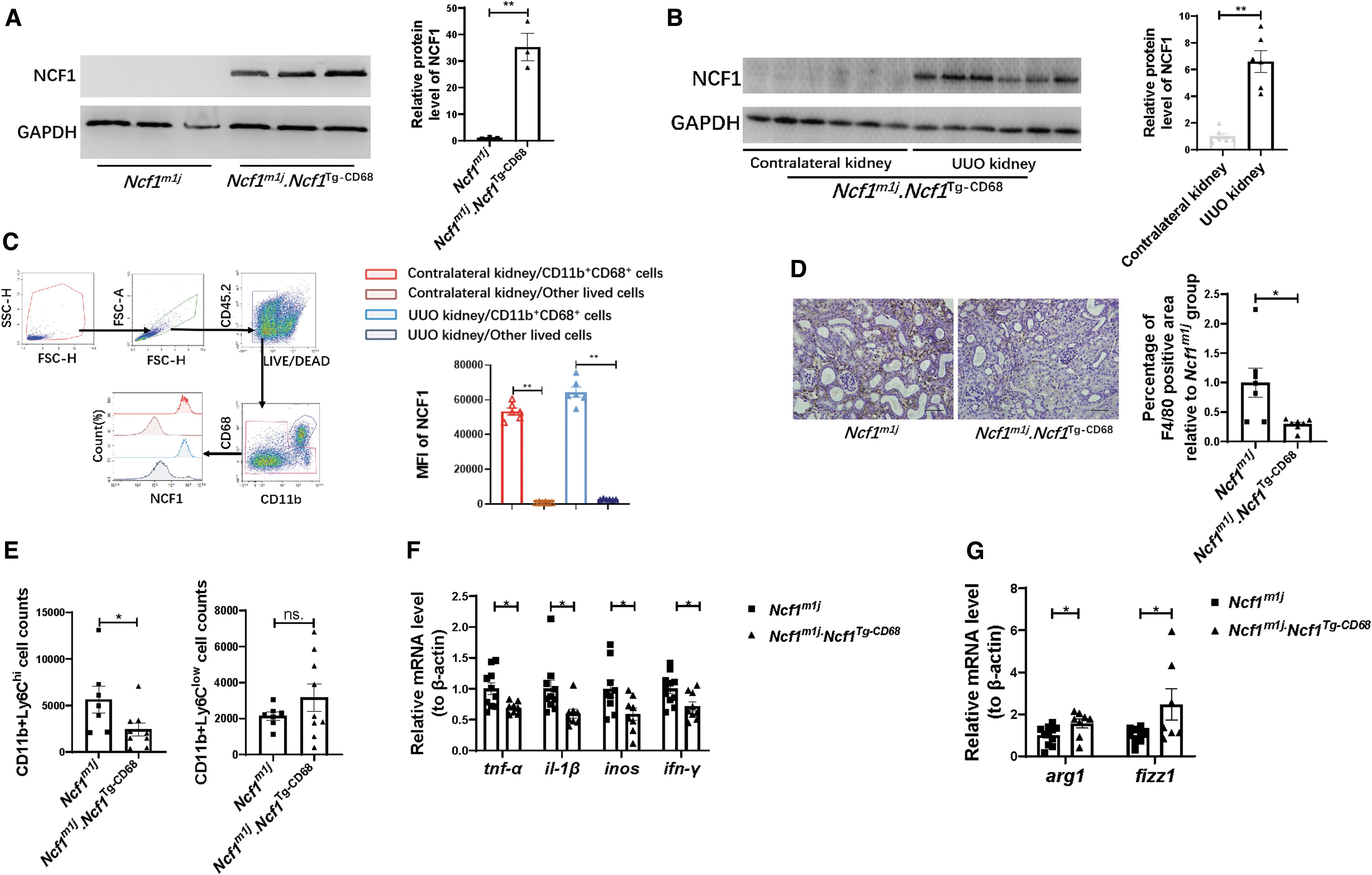
The results showed that in both the contralateral kidneys and UUO kidneys of the Ncf1m1j.Ncf1 Tg-CD68 mice, NCF1 is robustly expressed in CD11b+CD68+ macrophages but barely expressed in other lived cells (Fig. 6C). Immunohistochemistry staining demonstrated decreased infiltration of F4/80-positive macrophages in the interstitium of the Ncf1m1j.Ncf1 Tg-CD68 mice's UUO kidneys compared with Ncf1m1j mice's UUO kidneys (Fig. 6D). Flow cytometry was also used to analyze the Ly6C expression in monocyte/macrophage subsets in the kidneys of Ncf1m1j mice and Ncf1m1j.Ncf1 Tg-CD68 mice. Compared with the Ncf1m1j group, the UUO kidney of Ncf1m1j.Ncf1 Tg-CD68 mice contained lesser numbers of CD11b+Ly6Chi macrophages (Fig. 6E).
The Ly6Chi macrophages from the kidneys exhibited classical (M1) macrophages activation characteristics with high levels of proinflammatory cytokine production, such as, TNF-α, IL-1β (Lin et al., 2009). We next detected the mRNA expression level of M1 markers, including tnf-α, il-1β, inos, and ifn-γ, in the UUO kidneys of the Ncf1m1j mice and Ncf1m1j.Ncf1 Tg-CD68 mice. Ncf1m1j.Ncf1 Tg-CD68 mice's UUO kidney exhibited minor tnf-α, il-1β, inos, and ifn-γ expression (Fig. 6F). Different from the expression of markers of M1 macrophages, the expression of markers of M2 macrophages (arg1 and fizz1) in the UUO kidney significantly upregulated in the Ncf1m1j.Ncf1 Tg-CD68 group than in the Ncf1m1j group (Fig. 6G).
NCF1 promotes ROS production in UUO kidneys and macrophages
The expression of 4-hydroxynonenal (4-HNE) was used to assess the oxidative stress state of the UUO kidneys. Compared with the UUO kidney of the WT mice, the UUO kidney of the Ncf1m1j mice exhibited decreased 4-HNE staining (Fig. 7A). However, the positive area of 4-HNE staining in the UUO kidneys of Ncf1m1j mice and Ncf1m1j.Ncf1 Tg-CD68 mice had no significant difference (Fig. 7C). The NOX activity was significantly activated in the UUO kidneys of those three strains of mice compared with their contralateral kidneys (Fig. 7B, D). Similar to the result of 4-HNE assay, the NOX activity decreased significantly in both the contralateral kidney and the UUO kidney of the Ncf1m1j mice compared with that in the WT mice (Fig. 7B). But the NOX activity in either the contralateral kidney or the UUO kidney of the Ncf1m1j mice and Ncf1m1j.Ncf1 Tg-CD68 mice had no significant difference (Fig. 7D).
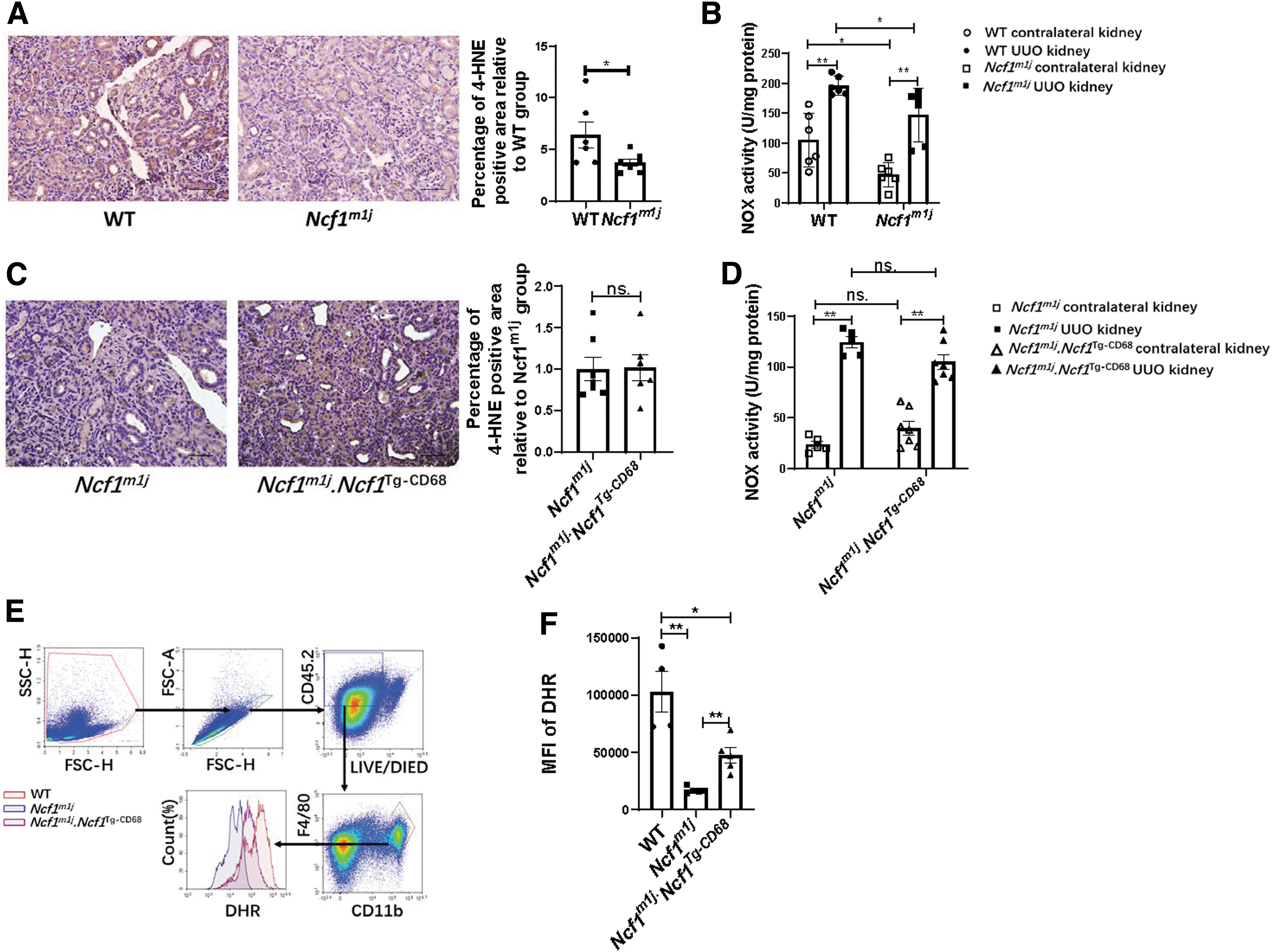
The ROS production capacity by macrophages was tested by stimulating spleen cells with phorbol myristate acetate (PMA), followed by dihydrorhodamine-123 staining. As shown in Figure 7E, the median fluorescence intensity of dihydrorhodamine was measured in the CD11b+F4/80+ macrophages from the spleen of WT mice, Ncf1m1j mice, and Ncf1m1j.Ncf1 Tg-CD68 mice. The level of DHR staining of CD11b+F4/80+ macrophages from Ncf1m1j mice was significantly decreased than that from WT mice, but CD11b+F4/80+ macrophages from Ncf1m1j.Ncf1 Tg-CD68 mice showed increased DHR staining than that from the Ncf1m1j mice (Fig. 7F). In summary, compared with the UUO kidney of the WT mice, the UUO kidney of the Ncf1m1j mice exhibited decreased oxidative stress level and NOX activity. Meanwhile, the oxidative stress level and the NOX activity of the UUO kidney of Ncf1m1j mice and Ncf1m1j.Ncf1 Tg-CD68 mice had no significant difference. However, NCF1 expressed in the macrophages enhanced the capacity of the ROS production of the macrophages.
Discussion
In this study, we focused on the role of the upregulated NCF1 in the progression of renal fibrosis induced by obstruction. We found that NCF1 expression was significantly increased in the kidney biopsies of CKD patients and mice's UUO kidneys. Compared with WT mice, Ncf1m1j mice exhibited mild renal fibrosis and decreased oxidative stress but increased total macrophages and proinflammatory CD11b+Ly6Chi macrophage infiltration. However, rescuing the NCF1 expression in macrophages further alleviates renal fibrosis in UUO mice. Quantitative and phenotypic analyses of macrophages infiltrating the UUO kidneys revealed fewer macrophages and lesser numbers of proinflammatory CD11b+Ly6Chi macrophages gathered in the UUO kidney of Ncf1m1j .Ncf1 Tg-CD68 mice than Ncf1 m1j mice (Fig. 8).
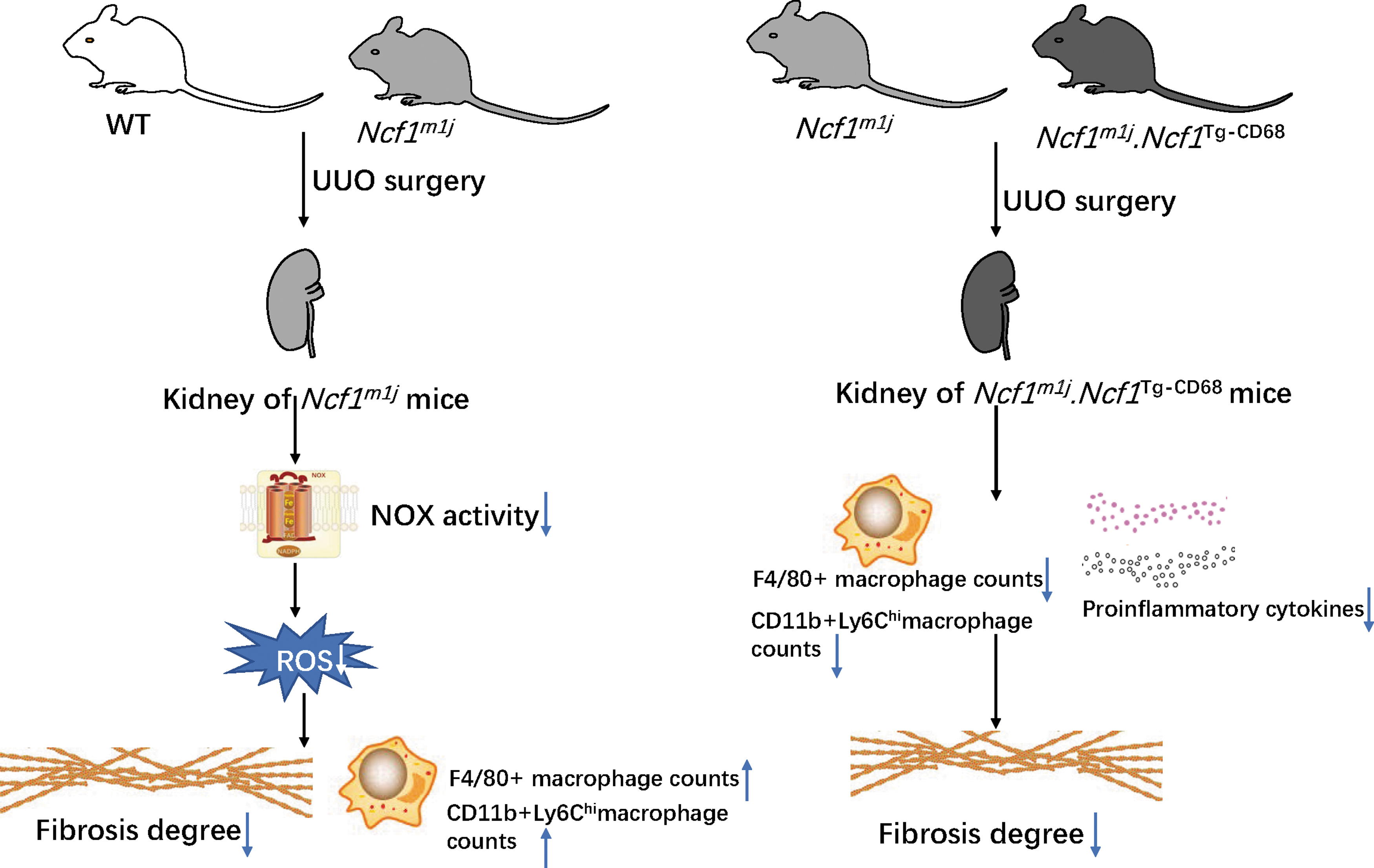
As demonstrated in this study, all subunits of the NOX2 complex were elevated in the UUO kidney. Both UUO kidney and CKD patients' renal biopsy exhibited evaluated NCF1 expression in the space of tubulointerstitium. Similar to our finding, some research showed increased expression of NCF1 and NOX2 in the injured kidney in several kidney disease models, such as Adriamycin-mediated kidney injury (Wang et al., 2015), diabetic nephropathy (Liu et al., 2012; You et al., 2013), and ischemia–reperfusion-induced kidney injury (Karim et al., 2015). However, these research did not resolve the issue of which cell type contributes to the elevated amount of NCF1. As demonstrated in our research, NCF1 expression increased not only in the tubular of fibrosis kidneys but also in the CD68+ macrophages in the kidneys of both the CKD patients and the UUO mice. Hence, it is valuable to explore the effect of NCF1 expressed in the intrinsic renal cells and the infiltrating macrophages in the progression of CKD.
Seven NOX isoforms have been identified. Among these, NOX1, NOX2, and NOX4 are expressed in both mice and human kidney (Nlandu Khodo et al., 2012). NOX4 is the major isoform in the kidney and is an essential source of renal ROS (Sedeek et al., 2013b). NOX4 has been reported to exert a destructive effect on diabetic nephropathy (Fujii et al., 2007; Gorin et al., 2015; Jha et al., 2016; Jha et al., 2014; Sedeek et al., 2013a), acute kidney injury (Meng et al., 2018), and hypertensive nephropathy (Cowley et al., 2016). However, there are also research indicating that NOX4 has protective effects on acute kidney injury (Nlandu-Khodo et al., 2016) and obstructive nephropathy (Nlandu Khodo et al., 2012). Although NOX4 is the predominant form in the kidney, NOX2 is also expressed in both human and mouse kidney (Sedeek et al., 2013b).
Among the five subunits of the NOX2 complex, NCF1 is the “organizer subunit,” which leads to a conformational change allowing its interaction with P22phox and then activating the NOX2 enzyme. The role of NCF1 or NOX2 in the progression of kidney disease has been reported by several groups (Geng et al., 2022; Karim et al., 2015; Kienhofer et al., 2017; Liu et al., 2012; Wang et al., 2015; You et al., 2013), but there is still no research on the role of NCF1 in the progression of the classic renal fibrosis model-UUO. Hence, we first reported that Ncf1m1j mice, which result in a relative deficiency of NCF1, have a relatively light fibrosis degree after the UUO surgery. In vitro experiments also demonstrated that knockdown of NCF1 expression in tubular epithelial cells alleviated TGF-β1-induced ECM production. Meanwhile, Ncf1 mutant mice showed decreased 4-HNE expression and NOX activity than WT mice, indicating that Ncf1 mutant mice had lighter ROS production and less oxidative stress than WT mice.
Therefore, we concluded that NCF1/p47 in intrinsic renal cells, including renal tubular cells, accelerates the cells to suffer fibrosis injury by promoting NOX activity and ROS production. Although there still lack in vivo research that provides direct evidence on the role of NCF1/p47 in renal tubular epithelial cells, the role of NOX-derived ROS in the kidney is clear. Under physiological conditions, nonphagocytic NADPH oxidases in the kidney have very low-level constitutive activity (Li and Shah, 2003). However, enzyme activity can be upregulated both acutely and chronically in response to stimuli, such as growth factors, cytokines, and mechanical stress (Berry et al., 2000; Inoguchi et al., 2000; Jaimes et al., 1998; Li et al., 2002; Yasunari et al., 2002). ROS production by the NOX could serve a signaling role (Kappert et al., 2000; Mollnau et al., 2002; Sundaresan et al., 1995) and also lead to oxidative damage (Geiszt et al., 2000; Nath and Norby, 2000; Shiose et al., 2001).
Hence, we predict that systemic mutation of Ncf1 ameliorates obstruction-induced renal fibrosis via alleviating ROS production and oxidative stress injury in renal intrinsic cells, including renal tubular epithelial cells. Our results align with Liu et al.'s (2012) findings in the mice model of diabetic nephropathy deletion of Ncf1 attenuated high glucose-induced activation of profibrotic gene expression in glomeruli and mesangial cells. And this research indicated that NCF1-dependent NOXs are important sources of superoxide in the diabetic glomerulus. But, You et al. (2013) reported that diabetic glomerular and tubulointerstitial disease was not affected by the deletion of Nox2.
NOX2 is mainly expressed in phagocytes, including macrophages, and macrophages contribute to kidney injury and repair (Han et al., 2011; Lech et al., 2014; Ma et al., 2011; Tan et al., 2010; Wise et al., 2014; Zhang et al., 2014). Hence, it is valuable to define the specific role of NOX2 in regulating macrophage recruitment and polarization in renal fibrosis. Contrary to the protective effect of NCF1 inhibition on kidney injury reported by several groups (Liu et al., 2012; Wang et al., 2015), recent research on lupus nephritis indicated that the mutation of Ncf1 aggravates pristane-induced lupus nephritis (Geng et al., 2022; Kienhofer et al., 2017). In addition, they also found elevated levels of inflammatory cytokines and chemokines, such as IL-12, IL-6, IL-10, IL-17a, IFN-γ, TNF-α, and MCP1, in the Ncf1 mutant mice.
These seemingly contradictory conclusions provoke us to hypothesize that NCF1 expressed in phagocytes such as macrophages reduces inflammatory cell infiltration and inflammatory cytokines secretion and then alleviates the renal fibrosis severity. Hence, we set out to compare the fibrosis and inflammatory condition between the Ncf1 mutant group and the Ncf1m1j.Ncf1 Tg-CD68 group. As we have demonstrated, Ncf1m1j.Ncf1 Tg-CD68 mice showed mild fibrosis and inflammatory infiltration compared with the Ncf1m1j mice. Therefore, we supposed that NCF1 expressed in the infiltrating macrophages of fibrosis kidney alleviates renal fibrosis by reducing inflammatory cell counts and inflammatory cytokine secretion. Similar to our results, several research that used the Ncf1m1j.Ncf1 Tg-CD68 mice reported that NCF1 expressed in the CD68+ macrophages protected against arthritis (Gelderman et al., 2007; Shakya et al., 2016) and chronic granulomatous disease (Deffert et al., 2012; Guerard et al., 2016; Pizzolla et al., 2012).
In addition, these research also showed that Ncf1m1j.Ncf1 Tg-CD68 mice were protected against hyperinflammation and suppressed the long-lasting production of inflammatory cytokines such as IL-1β, IL-6, and TNF-α compared with Ncf1m1j mice (Deffert et al., 2012; Gelderman et al., 2007), which is consistent with our results. As has been demonstrated by Deffert et al. (2012), the amount of ROS production in mononuclear phagocytes is markedly lower than that observed in neutrophils, which is consistent with our finding that there is no difference in 4-HNE between Ncf1m1j mice and Ncf1m1j.Ncf1 Tg-CD68 mice. The DHR staining of the spleen-derived macrophages indicated that NCF1 expressed in the macrophages promotes the ROS production capacity of macrophages. Although the role of NOX2/NCF1 in mononuclear phagocytes is less clear, we supposed that NOX2/NCF1 regulates the function of macrophages through a signaling function of NOX2-derived ROS.
In summary, NCF1 expression is upregulated in fibrosis kidneys, and systemic mutation of Ncf1 ameliorates obstruction-induced renal fibrosis but aggravates macrophage infiltration. Interestingly, macrophage-rescued NCF1 further alleviates renal fibrosis and macrophage infiltration. These results suggest that NCF1 expressed in renal intrinsic cells and infiltrating macrophages exert opposing effects on the progression of renal fibrosis. Hence, a better understanding of the function and mechanisms of NCF1 expressed in renal intrinsic cells and infiltrating inflammatory cells will provide an opportunity to define new therapeutic strategies for managing CKD.
Materials and Methods
Human specimens
Formalin-fixed paraffin-embedded renal biopsies from patients with IgA nephropathy (n = 12), mesangial proliferative glomerulonephritis (n = 5), membranoproliferative glomerulonephritis (n = 3), lupus nephritis (n = 1), and crescentic glomerulonephritis (n = 1) were obtained from the First Affiliated Hospital of Xi'an Jiaotong University. Kidney tumor-adjacent tissues from five patients used as a control group were obtained from the Second Affiliated Hospital of Xi'an Jiaotong University. Informed written consent was given by the patients for using part of the kidney tissue for scientific purposes. The experiment using human kidney tissue was approved by the ethics committee of Xi'an Jiaotong University Biomedical Center (2019-652). Two micrometer–thick, formalin-fixed paraffin-embedded sections were stained by immunohistochemistry with rabbit monoclonal antibody to NCF1 (ab181090; Abcam, Cambridge, United Kingdom).
Animals and UUO model
C57BL/6 mice purchased from Xi'an Jiaotong University animal center were used to detect the expression of NCF1 in the UUO model's kidney. The Ncf1 mutant mice (Ncf1m1j ) have a point mutation in the splice site for exon 8 of Ncf1 gene, which causes a truncated and nonfunctional NCF1 protein (Huang et al., 2000; Hultqvist et al., 2004). The mice were purchased from The Jackson Laboratory and backcrossed to B10.Q for over 12 generations (Gelderman et al., 2007). The Ncf1m1j.Ncf1 Tg-CD68 mice have a transgene expressing functional NCF1 on macrophages using the human CD68 promotor (Gelderman et al., 2007; Khmaladze et al., 2014; Pizzolla et al., 2012). All mice used in our study were 8–12 weeks old. All animal experimental procedures were approved by the ethics committee of Xi'an Jiaotong University Biomedical Center (2020-1095).
Mice were anesthetized with pentobarbital sodium (50 mg/kg) and placed on a heating map to maintain body temperature at 37°C during the operation. Then, a lateral incision was made on the left back, and the left ureter was carefully separated and ligated with the 4-0 silk. Fourteen days after the operation, the mice were euthanized, and the kidneys were collected for further examination.
Peritoneal macrophage generation
Peritoneal macrophages were extracted from mice following injection with 2 mL of 3% thioglycolate broth medium for 72 h. Peritoneal lavage was performed with 5 mL of phosphate-buffered saline (PBS) buffer containing 0.3 mM EDTA. Cells were centrifuged for 5 min at 1500 rpm at 4°C, and then, red cells were lysed by ACK buffer. The cells were cultured in Dulbecco's modified Eagle's medium (DMEM; HyClone, Logan, UT) containing 10% fetal bovine serum (FBS; HyClone), 20 μg/mL penicillin, and 20 μg/mL streptomycin (Sigma–Aldrich, St. Louis, MO) for 24 h and maintained at 37°C in a humidified incubator with 5% CO2. For experiments, cells were seeded at a density of 1.5 × 105 cells/mL in a six-well plate.
Cell culture and transfection
The HK2 cells (Procell, Wuhan, China) were cultured in DMEM/F12 medium with 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin. HK2 cells were cultured in a cell incubator with a 100% humidified atmosphere of 5% CO2 and 95% air (standard culture condition) at 37°C. Cells with 1 × 105 numbers were seeded into each well of a 12-well plate. When cells reached 50%–70% confluence in the plate, negative control sequence (50 nM) or 50 nM siRNA mixture sequence (containing three different kinds of sequences) targeting human NCF1 was transfected into the HK2 cells by Lipofectamine 2000 (Invitrogen, MA) according to the manufacturer's protocol. Their sequences are listed in Table 1. After receiving synchronized treatment (cells were cultured in serum-free medium) for 12 h, TGF-β1 (10 ng/mL; Sino Biological company, Beijing, China) was used to treat the cells for 48 h to detect fibrosis markers.
Small Interfering RNA Sequences Targeting Human NCF1 mRNA
RNA extraction and real-time quantitative polymerase chain reaction
Total RNA was extracted from renal tissues with TRIzol Reagent (Invitrogen, Carlsbad) and reversely transcribed into cDNA with RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Waltham, MA) according to the manufacturer's protocol. The real-time quantitative polymerase chain reaction assays were performed with SYBR-GREEN (Roche, Basel, Switzerland). The primers for Ncf1, Nox2, P22phox, P67phox, P40phox, Tnf-α, Il-1β, iNos, Ifn-γ, Arg1, and Fizz1 are listed in Table 2. Relative expression levels of those mRNA were normalized by β-actin and calculated by the 2−ΔΔCt approach.
Real-Time Polymerase Chain Reaction Primer
Western blotting
Renal tissues were homogenized in RIPA lysis buffer (Beyotime, Shanghai, China) containing the protease inhibitor cocktail (Roche, Indianapolis, IN). Fifty micrograms of total proteins was separated by 7.5% or 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) gels according to the molecular weight and transferred onto polyvinylidene fluoride membranes. After 2 h blocking, the membranes were incubated with primary antibodies, followed by secondary antibodies. Protein bands were visualized on X-ray film (Kodak, Eastman) using an ECL reagent. The relative expression levels of each protein were assessed by densitometric analysis and normalized by glyceraldehyde-3-phosphate dehydrogenase (GAPDH)/β-actin/β-tubulin using the ImageJ software. Target proteins were detected with antibodies against NCF1 (NB100-790; Novus) fibronectin (ab199056; Abcam), fibronectin (ab32419; Abcam), α-SMA (19245; CST, Danvers), vimentin (HN1123; HUABIO, Hangzhou, China), GAPDH (60004-1-Ig; Proteintech, Wuhan, China), β-actin (4970; CST), and β-tubulin (WH267526; ABclonal, Wuhan, China).
Histological and immunohistochemical analysis
Kidney tissue was fixed in 4% paraformaldehyde and then sectioned into 4 μm for Sirius Red staining, modified Masson's trichrome staining, and immunohistochemistry. Sirius Red staining was performed according to the manufacturer's introduction (ST073; HEART, Xi'an, China). Modified Masson's Trichrome Staining Kit (G1346; Solarbio, Beijing, China) was used to assess the degree of collagen deposition. The expression of NCF1, α-SMA, fibronectin, col3a1, and 4-HNE (ab48506; Abcam) was detected by immunohistochemical analysis, respectively. Briefly, renal sections were routinely dewaxed and hydrated, and then, antigens were recovered by 10 mM citrate buffer (pH 6.0) or multiple digestive enzymes. Nonspecific antigens were blocked with 5% bovine serum albumin, and a specific primary antibody was incubated overnight, followed by incubation with a horseradish peroxidase-conjugated secondary antibody and subsequently visualized with 3,3′-diaminobenzidine tetrahydrochloride. Finally, Image-Pro Plus 6.0 software was used to quantify the positive area.
Immunofluorescence
The human renal biopsy section was routinely dewaxed and hydrated, and then, antigens were recovered by multiple digestive enzymes. The sections were incubated with rabbit monoclonal antibody to NCF1 (ab181090; Abcam) overnight, followed by Rhodamine AffiniPure Donkey Anti-Mouse IgG (715-025-150; Jackson Immunoresearch, PA) and Alexa Fluor 488 anti-mouse CD68 antibody (137012; Biolegend, CA) for 1 h. Images were acquired with a fluorescence microscope (BX51; Olympus, Tokyo, Japan).
Flow cytometry
Mice were perfused with cold PBS, and then, kidneys were collected, minced into pieces, and digested in DMEM-F12 containing 2 mg/mL collagenase type I and 100 μg/mL DNase I for 1 h at 37°C. Kidney pieces were passed through a 40 μm mesh, yielding single-cell suspensions. The cells with the number of 1.0 × 105 were collected and first stained with a LIVE/DEAD Fixable Violet Stain Kit (Thermo Fisher, MA) and then treated at 4°C for 30 min in the dark with the following antibodies for the membrane antigen: BV785-conjugated anti-mouse CD45.2 (BD Biosciences, NJ), FITC-conjugated anti-mouse F4/80 (BD Biosciences), APC-conjugated anti-mouse CD11b (BD Biosciences), PB-conjugated anti-mouse ly6G (BD Biosciences), and BV650-conjugated anti-mouse ly6C (BD Biosciences).
For NCF1 and CD68 staining, cells were fixed and permeated by using BD Cytofix/Cytoperm after membrane staining and labeled with monoclonal antibodies for intracellular staining of AF 680-conjugated anti-mouse NCF1 (Santa Cruz) and PB-conjugated anti-mouse CD68 (BD Biosciences). Flow cytometric analysis was performed by Agilent NovoCyte 3005 Flow Cytometer (Agilent, CA) and FlowJo software.
The activity of NADH oxidase
The activity of NADH oxidase was assayed by a UV-2450 UV/Vis spectrophotometer (Shimadzu, Shanghai, China) using the commercially NOX activity assay kit (Solarbio, Beijing, China) following the manufacturer's instructions (Dou et al., 2017). The enzyme activity was expressed as units per milligram protein (U/g pro).
Oxidative burst assay
As described previously, the mice spleen cells with 2.0 × 106 numbers were collected and stained for ROS production (Gelderman et al., 2007). FcR interaction was blocked with an anti-FcR blocker (BD Biosciences) before incubation with the surface markers BV785-conjugated anti-mouse CD45.2, APC-conjugated anti-mouse CD11b, and BV711-conjugated anti-mouse F4/80 (BD Biosciences). Dihydrorhodamine 123 (Sigma, MO) was added to the cells at a concentration of 3 nM, and the cells were stimulated with PMA at a concentration of 200 ng/mL. After washing, cells were analyzed by Agilent NovoCyte 3005 Flow Cytometer (Agilent) and FlowJo software.
Statistics
All data are presented as the mean ± standard error of the mean. Student's t-test was used to analyze the statistical significance between two groups. ANOVA was used for multiple group comparisons, followed by the Kruskal–Wallis test. The statistics analysis was performed by GraphPad Prism 8.0 software, and a p value <0.05 was considered statistically significant.
Footnotes
Authors' Contributions
F.H., X.R., B.Y., W.Y., L.X., J.Z., and H.Z. performed the experiments and analyzed the data. M.G., X.L., J.X., W.Z., L.M., and S.L. contributed to writing the article. S.L., L.M., W.Z., and F.H. designed the study. S.R. collected and provided the CKD patients' biopsy. S.L. and L.M. supervised the study.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of 81970029, 32070913, and 82171784. Originatively, B10Q mice, Ncf1 mutant mice, and Ncf1m1j.Ncf1 Tg-CD68 mice used in this research were kindly donated by Prof. Rikard Holmdahl from Karolinska Institute, Sweden.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.