
Abstract
Aims:
Intervertebral disc degeneration (IDD) is closely related to low back pain, which is a prevalent age-related problem worldwide; however, the mechanism underlying IDD is unknown. Glutamine, a free amino acid prevalent in plasma, is recognized for its anti-inflammatory and antioxidant properties in various diseases, and the current study aims to clarify the effect and mechanism of glutamine in IDD.
Results:
A synergistic interplay was observed between pyroptosis and ferroptosis within degenerated human disc specimens. Glutamine significantly mitigated IDD in both ex vivo and in vivo experimental models. Moreover, glutamine protected nucleus pulposus (NP) cells after tert-butyl hydroperoxide (TBHP)-induced pyroptosis, ferroptosis, and extracellular matrix (ECM) degradation in vitro. Glutamine protected NP cells from TBHP-induced ferroptosis by promoting the nuclear factor erythroid 2-related factor 2 (Nrf2) accumulation by inhibiting its ubiquitin–proteasome degradation and inhibiting lipid oxidation.
Innovation and Conclusions:
A direct correlation is evident in the progression of IDD between the processes of pyroptosis and ferroptosis. Glutamine suppressed oxidative stress-induced cellular processes, including pyroptosis, ferroptosis, and ECM degradation through deubiquitinating Nrf2 and inhibiting lipid oxidation in NP cells. Glutamine is a promising novel therapeutic target for the management of IDD.
Introduction
Low back pain (LBP) is a highly prevalent musculoskeletal disease with multiple contributing factors, and intervertebral disc degeneration (IDD) is a closely related condition (Knezevic et al., 2021). The prevalence of LBP within a person’s lifetime is estimated to be almost 70%, resulting in significant social and economic burden (Gianola et al., 2022, Hoy et al., 2014). IDD is characterized by the progressive degeneration and breakdown of an intervertebral disc (IVD), which are key contributors to inhibited spine movement (Adams et al., 2000). An IVD comprises three interconnected components: the outer annulus fibrosus (AF), cartilaginous endplates (CEPs), and NP. NP is a gel-like tissue located in the central region of an IVD and comprises a high concentration of proteoglycans (Urban and Roberts, 2003). The vulnerability of IVDs to inflammatory factors and reactive oxygen species (ROS) is primarily attributed to the inadequate nutrient level available to NP cells resulting from the absence of blood flow. These factors lead to the disruption of ECM component synthesis and loss of NP cells and then play a crucial role in the progression of IDD (Zhang et al., 2021). However, whether this factor also further compromises the structural integrity and biomechanical stability of an IVD needs to be determined (Adams and Roughley, 2006).
Innovation
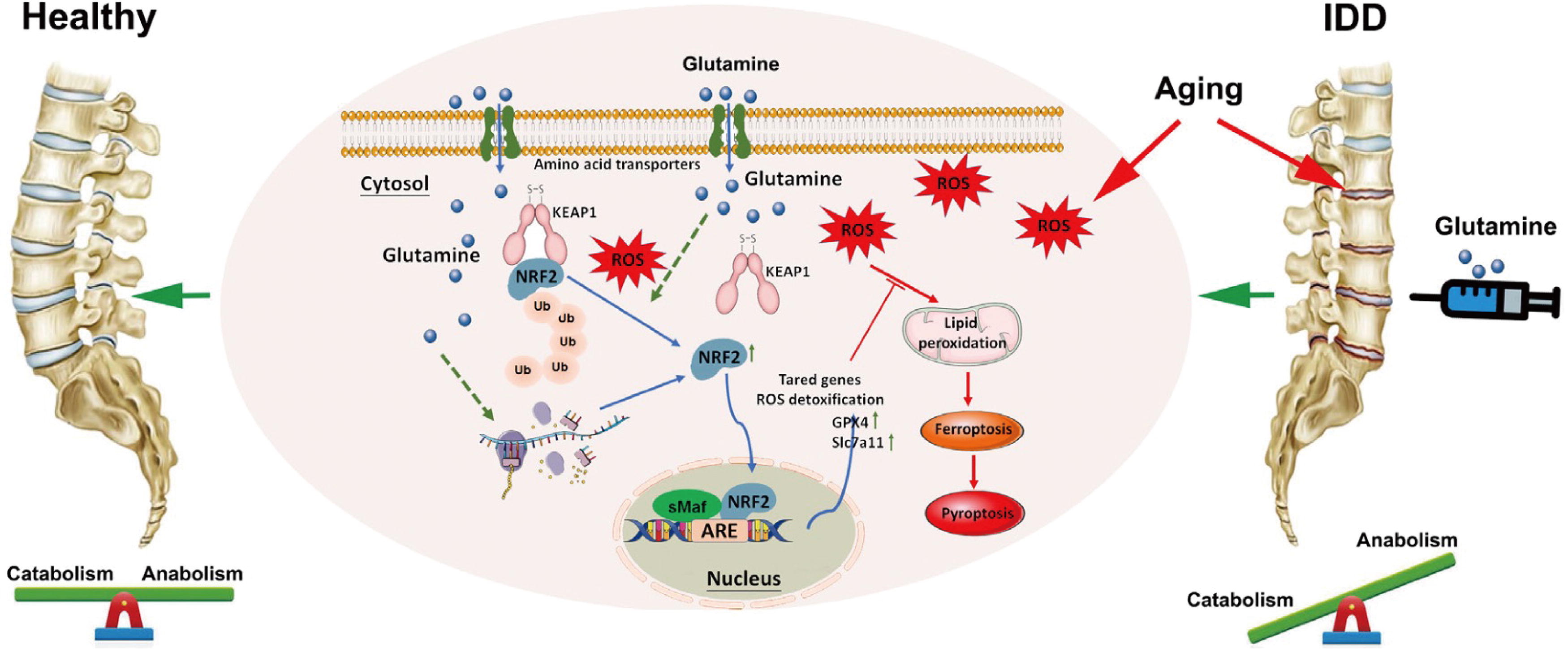
The present study identified glutamine as a promising therapeutic agent for slowing the progression of IDD. Through analysis of clinical samples and ex vivo assays, the current study reports the potential involvement of ferroptosis and pyroptosis in NP cells with increased oxidative stress. Further in vitro investigations confirmed that glutamine can mitigate ferroptosis, pyroptosis, and ECM degradation in NP cells. In addition, the findings revealed that glutamine mediates the deubiquitination of Nrf2 and subsequent activation of GPX4 to reduce ROS and NLRP3 levels. Hence, glutamine effectively mitigates ECM degradation and pyroptosis (Fig. 1).
The programmed death of NP cells is one of the reasons for IDD. Among the diverse modalities of programmed cell death, pyroptosis has emerged as a crucial player in the death of NP cells. Pyroptosis is a tightly regulated form of cell death characterized by the formation of plasma membrane pores. Importantly, pyroptosis triggers the activation and release of inflammatory cytokines, which further amplify inflammation and promote ECM degradation (Chao-Yang et al., 2021). Notably, the NOX1/ROS/NF-κB axis mitigates pyroptosis in NP cells (Deng et al., 2023). ROS governs the process of pyroptosis by oxidizing the cysteine residues within Gasdermin D (GSDMD) (Cuevas and Pelegrín, 2021). In addition to pyroptosis, ferroptosis has been found to cause the death of NP cells (Li et al., 2022, Yang et al., 2021). Ferroptosis is a multifaceted cellular process characterized by lipid peroxidation and ferrous iron overload (Dixon et al., 2012). This intricate death modality has been implicated in arthritis with notable alterations in the expression of pivotal genes such as glutathione peroxidase 4 (GPX4) (Wu et al., 2022). Yang et al (Yang et al., 2023b) investigated the effectiveness of antiferroptosis therapy in a rat model of IDD by employing polydopamine nanoparticles to inhibit ferroptosis. The contribution of ferroptosis to IDD resulted in partial deterioration of the disc microenvironment. Hence, elucidating the mechanisms underlying ferroptosis is crucial for comprehending the pathogenesis of IDD.
In addition, NLR family pyrin domain containing 3 (NLRP3) can detect and respond to a broad spectrum of agonists. Several notable examples of stimuli that activate NLRP3 include ATP, ROS, and factors released from mitochondria due to organelle damage (Horng, 2014, Ma et al., 2022a, Zeng et al., 2023). Notably, the lipid ROS can add fuel to pyroptosis (Kang et al., 2018). Moreover, enhancing the expression of GPX4 eliminates lipid oxidation, thereby inhibiting pyroptosis (Gu et al., 2022, Russo and Rathinam, 2018). The expression of GPX4 is upregulated by Nrf2, and it remains at a high level under normal physiological conditions (Yang et al., 2023a). The lipid ROS is hypothesized to be the link between these cell death pathways, as Nrf2 and GPX4 play critical roles in eliminating lipid ROS, and the lipid ROS is involved in pyroptosis activation.
Glutamine, a prominent nonessential amino acid in plasma, exhibits a multifaceted role in diverse cellular processes (Biswas et al., 2012, DeBerardinis and Cheng, 2010). Notably, glutamine plays a vital role in the production of various nonessential amino acids, which are fundamental building blocks of functional cellular proteins (Ahluwalia et al., 1990, Fan et al., 2013, Young and Ajami, 2001). Additionally, glutamine is a precursor in the synthesis of glutathione, a potent antioxidant molecule that facilitates ROS clearance and contributes to cellular homeostasis (Welbourne, 1979). In addition, glutamine regulates the progression of osteoarthritis (Ma et al., 2022b). Despite previous research, the potential impact of glutamine and the molecular mechanism underlying IDD remain unclear.
Herein, we evaluate the ameliorating effect of exogenous glutamine supplementation on NP cellular pyroptosis, ferroptosis, and ECM degradation via deubiquitinating and stabilizing Nrf2 under oxidative stress stimulation.
Discussion
Glutamine, an endogenous multifunctional and conditionally essential amino acid has attracted significant attention from researchers in various fields, including researchers studying infection (Powell-Tuck, 1999), colitis (Jeong et al., 2018), and sickle cell anemia (Anker et al., 2018). Glutamine supplementation has shown promising protective effects (Arra et al., 2022). Notably, the role of amino acid and cholesterol metabolism in IDD is significant (Chen et al., 2021, Yan et al., 2021, Zhang et al., 2020). Interestingly, glutamine is pivotal to the regulation of two aforementioned metabolic processes (Simon et al., 2020, Stegen et al., 2020). The current study revealed that glutamine alleviated IDD. Studies with puncture models have been the predominant approach for studying IDD in animals, as they offer advantages such as repeatability, simplicity of execution, and a short experimental cycle (Keorochana et al., 2010, Qin et al., 2023). The combination of intradiscal administration and a rat acupuncture model is a valuable approach for studying the effectiveness and mechanisms of different interventions for IDD. An ex vivo puncture model was selected for this study, and analysis of the model yielded valuable results (Zieba et al., 2022).
The pathogenesis of IDD involves multiple intricate processes, including oxidative stress, inflammation, ECM degradation, and various forms of cell death. Pyroptosis is distinguished by the activation of inflammatory cascades that exacerbate cell pathology (Chao-Yang et al., 2021). The pivotal event in pyroptosis initiation involves the activation of pro-caspase-1. Once activated, pro-caspase-1 triggers the maturation of GSDMD and IL-1β. The maturation of these molecules ultimately leads to the formation of pores in the cell membrane and the subsequent release of inflammatory cytokines. Although various types of inflammasomes have been described, research on IDD has primarily been focused on the classical NLRP3 inflammasome (Yan et al., 2021). The present study demonstrated that the administration of glutamine inhibited the TBHP-induced expression of NLRP3, GSDMD-NT and pro-caspase-1, and the release of LDH from NP cells. These findings suggest that glutamine can reverse pyroptosis induced by oxidative stress in NP cells.
Ferroptosis has garnered significant attention in studies aimed at the pathogenesis of IDD. Zhu et al expanded on prior research and demonstrated the involvement of SIRT3 and USP11 in IDD through their regulatory effects on ferroptosis (Zhu et al., 2023a). Previous studies have reported that glutamine deprivation induces ferroptosis, and glutamine inhibits ferroptosis through SLC1A5 (Yuan et al., 2022). In the current study, an escalation of ferroptosis was observed with IDD progression, which was effectively inhibited by exogenous glutamine treatment, highlighting the functional role of glutamine in maintaining redox homeostasis.
Notably, oxidative stress can induce pyroptosis, and the interplay between pyroptosis and ferroptosis factors has been previously documented in studies of various diseases, including radiation-induced intestinal injury (Zhang et al., 2022), tumors (Wang et al., 2022), and acute respiratory distress syndrome (Wang et al., 2023). Notably, caspases are activated and cleave GSDMD family members through ROS pathways (Zhou et al., 2018), and the knockout of GPX4, a key regulator of ferroptosis, has been shown to exacerbate the release of lipid peroxidation-dependent GSDMD-N and subsequent pyroptosis (Kang et al., 2018). However, the interrelation between pyroptosis and ferroptosis in IDD has been unexplored to date. Therefore, we utilized ferrostatin-1, a thiol-containing chemical agent known for its ferroptosis-inhibiting properties, to elucidate the relationship between pyroptosis and ferroptosis. The results showed that TBHP, an oxidative stress activator, was critical for inducing ferroptosis, and pretreatment with ferrostatin-1 significantly suppressed pyroptosis compared with the effect of TBHP treatment alone. Furthermore, GPX4 deletion exacerbated TBHP-induced pyroptosis under glutamine deprivation conditions.
Additionally, Nrf2 has been recognized for its essential role in protecting cells against oxidative-induced ferroptosis (Jiang et al., 2023). Previous studies have demonstrated that both MFG-E8 and hesperidin activate Nrf2, inhibiting NLRP3 expression in NP cells and preventing ferroptosis, respectively (Ma et al., 2022a, Zhu et al., 2023b). In the present study, as revealed by GEO analysis, Nrf2 was identified as one of the genes involved in both pyroptosis and ferroptosis, aligning with the role of glutamine in NP cells. Furthermore, posttranslational modifications of the Nrf2 protein, such as ubiquitination and sumoylation, are pivotal in influencing the activity of its downstream molecules (He et al., 2018, Huang et al., 2023). The present investigation demonstrated that glutamine contributes to oxidative stress-related IDD by regulating Nrf2 through modulation of its ubiquitination and stability. Notably, inhibition of Nrf2 activity abrogated the effects of glutamine on GPX4 and NLRP3, thus blocking the mitigating effects of glutamine on pyroptosis and ferroptosis after TBHP stimulation.
In summary, the present study showed the significant contribution of glutamine in attenuating oxidative stress-related IDD, and glutamine exerted a suppressive effect on pyroptosis and ferroptosis in NP cells, an effect that was mediated through Nrf2 ubiquitination and consistency regulation. Therefore, exogenous glutamine supplementation is a promising therapeutic strategy for preventing and treating IDD. However, several limitations need to be acknowledged. The metabolism of glutamine, a nonessential amino acid, in NP cells remains incompletely understood. Glutaminase 1 is the key enzyme critical to the protective effect of glutamine in IDD, and further investigation is needed to elucidate the catabolic processes of glutamine in NP cells.
Data Sharing Statement
No additional data are available.
Methods
Collection of human samples
This study was conducted following ethics guidelines, and approval was obtained from the Medical Ethics Committee of the Sun Yat-sen Memorial Hospital with code SYSKY-2022-217-0121. The study involved the collection of NP specimens from 30 patients who underwent surgical procedures between 2019 and 2023. Among the participants, 19 were females and 11 were males. Their ages ranged from 13 to 84 years. Table 1 provides additional details about the specimens used in the study. Before surgery, T2-weighted MRI scans were performed to evaluate the degree of disc degeneration using the Pfirrmann grading system. Notably, all patients provided informed consent for their voluntary participation in this study.
Details of Clinical Samples
Affected IVD: L, lumbar vertebra; S, sacral vertebra. IVD, intervertebral disc.
Ex vivo IVD organ culture
Fifteen male Sprague–Dawley rats (200–250 g) were euthanized at 12 weeks of age, and their Co7/8 discs were carefully isolated and used for a cell culture experiment performed at 37°C. The location of the Co7/8 discs was determined by palpating the coccygeal vertebrae. In this study, the IVDs were categorized into three distinct groups: a normal control group, a puncture group, and a puncture+ glutamine group. A 25-gauge needle was used to penetrate the Co7/8 discs vertically, and the needle was rotated steadily in a clockwise direction 360° over 30 s before being removed. Glutamine (Sigma-Aldrich, US) was administered via IVD injections every other day with a microsyringe (Hamilton, Switzerland) with a 34-gauge needle. Following a postsurgical period of 7 days, tail discs were harvested for further analysis.
Animals and surgical procedures
All animal experiments conducted in this study received ethics approval from the Institutional Animal Care and Ethics Committee of Sun Yat-sen University (Guangzhou, China). Fifteen male Sprague–Dawley rats, 12 weeks of age and weighing between 200–250 grams, were distributed into three distinct experimental groups: a sham group, an IDD+ PBS group, and a IDD+ glutamine group. Before the surgical procedures, the rats were anesthetized via intraperitoneal injections of 2% pentobarbital. The location of the Co7/8 intervertebral disc was identified through palpation. Following sterilization with 75% ethanol, a 25-gauge needle was rotated steadily in a clockwise direction through a 360° arc over 30 s before being carefully withdrawn. Subsequently, 5 μL of PBS or glutamine solution, with a concentration of 3 mg/mL, was injected using a 34-gauge needle attached to a microsyringe (Hamilton, Switzerland). This injection procedure was repeated every week. After four weeks, all rats underwent MRI scans and were then euthanized, with their tails removed and fixed in 4% paraformaldehyde for further analysis.
Histology and immunohistochemistry
Initially, the NP tissues were fixed with a 4% paraformaldehyde solution for 48 h. Subsequently, ethylenediaminetetraacetic acid (EDTA) was used for 8 weeks to induce rat tissue decalcification. Following decalcification, the samples were sectioned into slices. Finally, the histological changes in the NP tissues were examined after hematoxylin-eosin (H&E) and Safranin O staining. The tissue sections were first exposed to Hematoxylin for 2 min, followed by a 3-minute exposure to Eosin. Other tissue sections were stained with Safranin O staining and then were incubated for 15 min incubation with Safranin dye. Subsequently, the sections were treated with a Fast Green dye solution for 2 min.
Immunohistochemistry (IHC) is widely used for the visualization of specific proteins or antigens within tissue sections, enabling the examination of their localization and distribution. Antigen retrieval was conducted by treating the sections with pepsin for 30 min at 37°C. Initially, a 3% hydrogen peroxide solution was employed to quench the reaction, eliminating any inherent peroxidase activity that could potentially interfere with subsequent staining procedures. Subsequently, the slides were blocked with goat serum for 30 min to prevent the nonspecific binding of antibodies and minimize background staining. Immunostaining was performed using specific antibodies targeting the following proteins of interest: GPX4 (1:200, ab125066, Abcam), NLRP3 (1:200, A12694, ABclonal), p20 (1:200, AF4005, Affinity), GSDMD (1:200, ER1901-37, HUABIO), COL2A (1:200, A19308, ABclonal), and SLC7A11 (1:200, 26864-1-AP, Proteintech). The antibodies were incubated with the slides overnight to facilitate specific binding to their respective targets. Subsequently, the tissue sections were exposed to biotin-labeled secondary antibodies for 30 min at room temperature to detect the primary antibodies. Finally, the slides were incubated with horseradish peroxidase (HRP)-conjugated streptavidin for 20 min, enabling the visualization of immunostained regions via the activity of the horseradish peroxidase enzyme. The sections were then treated with a 3,3′-diaminobenzidine (DAB) solution (Zhongshan Jinqiao Biotechnical, Beijing, China) to initiate color development. After being stained with 1% hematoxylin, the sections were mounted and observed under a microscope, which was also used to capture images.
Rat NP cell isolation and culture
Male Sprague–Dawley rats weighing between 200 and 250 g were humanely euthanized to obtain NP tissues from the caudal IVDs. The collected tissue was digested for two hours in 0.25% type II collagenase (Solarbio, Beijing, China). Subsequently, the cells were thoroughly washed and cultured in DMEM supplemented with 10% fetal bovine serum and 1% penicillin–streptomycin. After proliferating and reaching 90% confluence, the cells were passaged by transfer to new culture plates, where they attached and continued to grow.
Cell viability assay
Reagents used to treat cells were aspirated from the culture and replaced with 100 μL of a working solution consisting of a 1:9 ratio of Cell Counting Kit-8 (CCK-8) solution to DMEM. The plate was placed in a 37°C incubator, shielded from the light, and incubated for one hour. Subsequently, the absorbance of the sample was determined at a wavelength of 450 nm with a microplate reader.
Western blot analysis
NP cells were lysed on ice for 30 min to prepare them for analysis. The lysis mixture consisted of 100 μL of RIPA buffer supplemented with 1 μL of a protease inhibitor and a phosphatase inhibitor (CWBIO, Beijing, China). The lysate was centrifuged at 14,000×g for 30 min to separate the cellular components. The protein concentration in the lysates was determined using the BCA Total Protein Assay Kit (CWBIO, Beijing, China). Subsequently, the protein samples were loaded onto SDS–PAGE gels (Epizyme, Shanghai, China) and transferred to PVDF membranes for further analysis. Primary antibodies against β-actin (1:1000, AC006, ABclonal), COL2A (1:500, ab34712, Abcam), Aggrecan (1:500, ab36861, Abcam), GPX4 (1:1000, ab125066, Abcam), NLRP3 (1:1000, A12694, ABclonal), ADAMTS5 (1:1000, A2836, ABclonal), MMP3 (1:1000, A1202, ABclonal), Nrf2 (1:1000, 16396-1-AP, Proteintech), SLC7A11 (1:1000, 26864-1-AP, Proteintech), GSDMD (1:500, 39754, CST), p20 (1:500, AF4005, Affinity), and Ubiquitin (1:1000, YM3636, ImmunoWay), were used to probe the blots. Following membrane washing, HRP-conjugated secondary antibodies (CWBIO, Beijing, China) were applied to the membrane, and an enhanced chemiluminescence (ECL) kit (Vazyme, Nanjing, China) was used to develop the bands. The resulting images were captured using an ECL imager (Syngene G:BOX ChemiXT4, United Kingdom). All original bands have been shown in the Supplementary material (Fig. S1–2).
Coimmunoprecipitation
The experimental procedure commenced by lysing cells using Western blot and IP lysis buffer (Beyotime, Shanghai, China). To ensure the preservation of protein integrity, a cocktail of protease (CWBIO, Beijing, China) inhibitors was added to the lysis buffer. The cells were lysed for 20 min to enable the disruption of cellular membranes and the subsequent release of protein complexes. Subsequently, the cell supernatants were collected after lysate centrifugation at 12000 rpm and 4°C for 10 min to eliminate cellular debris. Specific antibodies targeting the proteins of interest were added to the supernatant. To facilitate the formation of antibody–protein complexes, the mixture was gently rotated overnight at 4°C. The antibody–antigen complexes in the solution were mixed with magnetic beads (Beyotime, Shanghai, China) for 120 min. After being washed with immunoprecipitation (IP) lysis buffer, the immunoprecipitants were eluted after boiling in 1 × SDS-loading buffer for 10 min, and they were used for immunoblotting.
Immunofluorescence
NP cells were fixed with 4% paraformaldehyde for 30 min at room temperature to ensure the preservation of their structural integrity. To facilitate cell membrane permeabilization, a solution containing 0.1% Triton X-100 was used for 10 min. To mitigate nonspecific binding events, goat serum was incubated for 30 min. The specific targets of interest were visualized by employing specific antibodies, which were introduced and incubated overnight at a temperature of 4°C. For enhanced detection, fluorescein-conjugated secondary antibodies (Zhongshan Jingqiao Biotechnical, Beijing, China) and DAPI dye (Solarbio, Beijing, China) were utilized. Ultimately, the cellular structures and fluorescence signals were captured using an Olympus IX73 microscope (Olympus, Tokyo, Japan).
Transmission electron microscopy (TEM)
The NP cells were carefully collected and then subjected to a fixation procedure. Specifically, the cells were incubated in a solution containing 2.5% glutaraldehyde at a temperature of 4°C overnight. Subsequently, the samples were dehydrated with an ethanol gradient and then embedded in Epon 812. The cells were then sliced into 100 nm sections with an ultramicrotome. The cell morphology was captured with a Tecnai G2 transmission electron microscope.
Lactate dehydrogenase assay
The amount of lactate dehydrogenase (LDH) released from NP cells was measured in this study by CytoTox96 nonradioactive cytotoxicity assay (Promega, US). After treatment, a 50-μL volume of supernatant was collected and thoroughly mixed with 50 μL of LDH reagent to ensure proper combination. The resulting mixture was then incubated for 30 min. Subsequently, 50 μL of stop solution was added to the mixture to terminate the reaction. The absorbance of the solution was measured at a wavelength of 492 nm using a microplate reader.
ROS levels
Cells were incubated with 10 μM dichlorodihydrofluorescein diacetate (DCFH-DA; Beyotime, Shanghai, China) at 37°C for 30 min to facilitate penetration of the dye. Subsequently, the cells were collected and rinsed with PBS to eliminate any residual DCFH-DA. Specifically, a BD FACSVerse instrument was utilized for flow cytometry analysis.
Lipid ROS levels
Lipid ROS levels were evaluated by treating cells with a 10 μM C11-BODIPY 581/591 probe (Thermo Fisher Scientific, US). Subsequently, the cells were incubated at 37°C for 30 min to allow the necessary reactions. Following the incubation period, the cells were thoroughly examined using an Olympus IX73 microscope (Olympus, Tokyo, Japan).
Determination of the superoxide anion level in mitochondria
Cells were incubated with two specific stains, MitoTracker Green FM (Yeasen, Shanghai, China) and MitoSOitX Red (Yeasen, Shanghai, China) at 37°C for 30 min, to ensure effective interaction between the stains and the targeted cellular components. Subsequently, the cells were gently washed with PBS to eliminate any stain that had not bound or had bound nonspecifically to untargeted substrates. To visualize and analyze the stained cells, we employed an Olympus IX73 microscope (Olympus, Tokyo, Japan).
RNA interference
Small interfering RNA (siRNA) was employed to interfere with the transcription of specific target genes in NP cells. The siRNAs targeting the genes were synthesized by IGE Biotechnology (Guangzhou, China). The transfection procedure was carried out following the manufacturer’s instructions. The following siRNA sense sequences are used: Nrf2-siRNA1, 5‘-CGAGAAGUGUUUGACUUUA-3’; Nrf2-siRNA2, 5‘-GGAUGAAGAGACCGGAGAA-3’; Nrf2-siRNA3, 5‘-GAGUUACAGUGUCUUAAUA-3’; GPX4-siRNA1, 5‘-GCGUGUGCAUCHUCACCAA-3’; GPX4-siRNA2, 5‘-GGUUUGACAUGUACAGCAA-3’; GPX4-siRNA3, 5‘-CCAAGUUUCUCAUUGAUAA-3’.
Gene expression data
In this study, we procured two lists of distinct genes, one for pyroptosis and one for ferroptosis genes, from reputable sources. The pyroptosis list included 365 genes and was obtained from GeneCards (https://www.genecards.org). Similarly, the ferroptosis list, comprising 634 genes, was acquired from FerrDb (http://www.zhounan.org/ferrdb/current). A total of 24 NP samples with varying degrees of degeneration were collected for analysis; 16 samples were obtained from the IDD group (Thompson grade III/IV/V) and 8 samples were obtained from the CTR group (Thompson grade I/II). Data were obtained from the GSE70362 dataset.
Statistical analyses
Data analyzed in this study were derived from a minimum of three independent experiments, and the results are presented as the mean ± standard deviation (SD). Statistical analyses were performed using GraphPad Prism 8 software. To compare values between the two groups, an unpaired two-tailed Student’s t-test was performed. In addition, to examine differences among multiple groups, a one-way analysis of variance (ANOVA) was performed. In both analyses, a p value <0.05 was considered statistically significant. Electronic laboratory notebook was not used.
Results
A synergistic interplay between ferroptosis and pyroptosis in human NP tissue
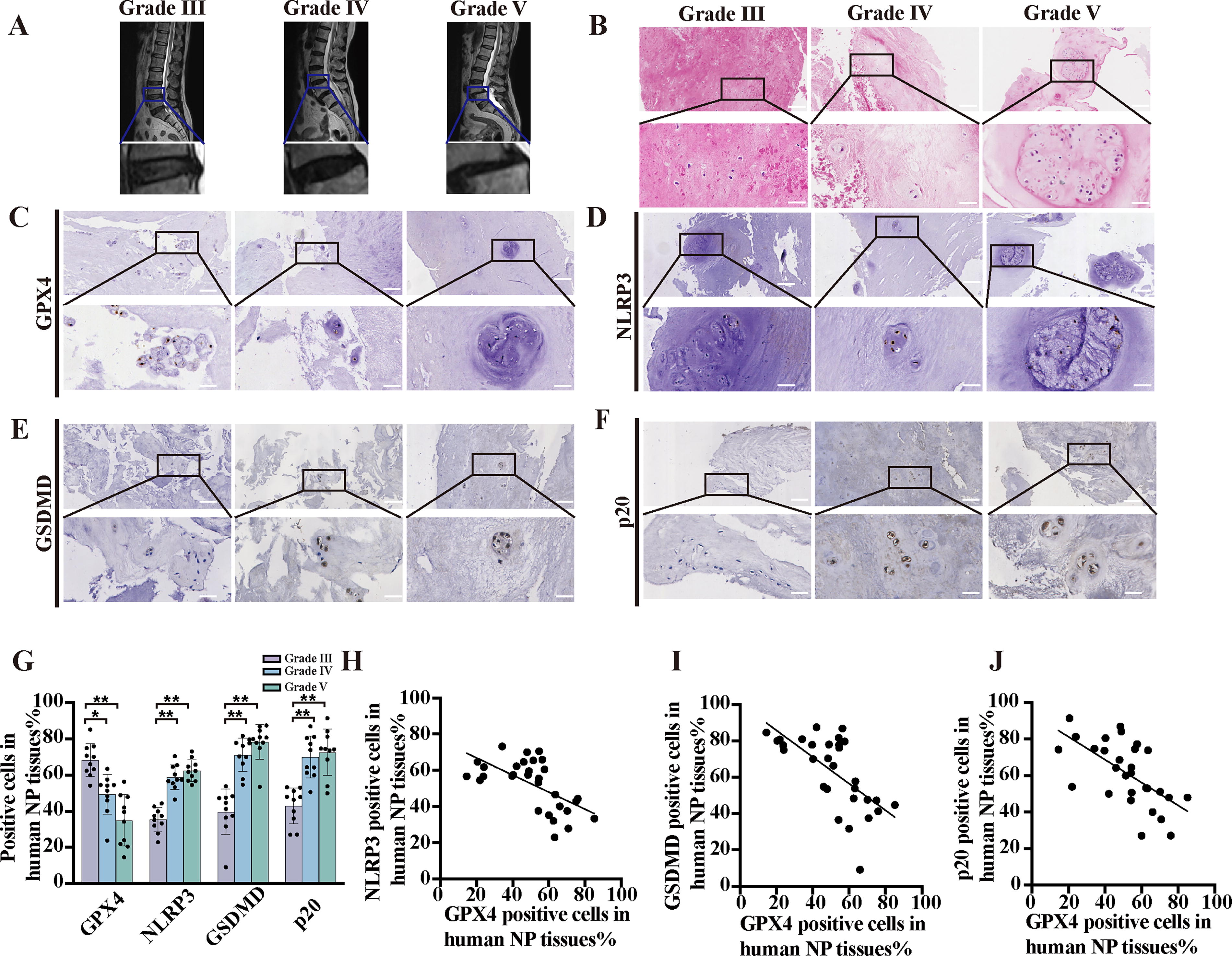
To evaluate the involvement of ferroptosis and pyroptosis in promoting IDD, a total of 30 human NP tissues exhibiting various levels of degeneration according to the Pfirrmann grading system were collected and subjected to magnetic resonance imaging (MRI) (Fig. 2A). In this investigation, we employed H&E staining to evaluate the cell density and presence of atypical cell clusters in NP tissue specimens with various Pfirrmann grades (Fig. 2B). In addition, we performed IHC analyses to measure the expression of GPX4, p20, GSDMD, and NLRP3 in degenerative NP tissues (Fig. 2C–G). Our findings revealed a significant reduction in GPX4 expression and a concurrent increase in p20, GSDMD, and NLRP3 expression compared with those in the normal group. To further explore the potential relationship between GPX4 and p20, GSDMD, and NLRP3 expression in NP tissues with similar Pfirrmann grades, we performed a single-factor linear regression analysis (Fig. 2H–J). Importantly, our results demonstrated a negative correlation between GPX4 and p20 expression (Y = −0.6529*X + 93.80, r = 0.4028, p = 0.0002), GPX4 and GSDMD expression (Y = −0.7333*X + 100.5, r = 0.43, p = 0.0001), and GPX4 and NLRP3 expression (Y = −0.5123*X + 80.27, r = 0.4776, p = 0.0043), suggesting a potentially significant role for GPX4 in modulating pyroptosis.
Intradiscal injection of glutamine alleviates puncture-induced IDD ex vivo and in vivo
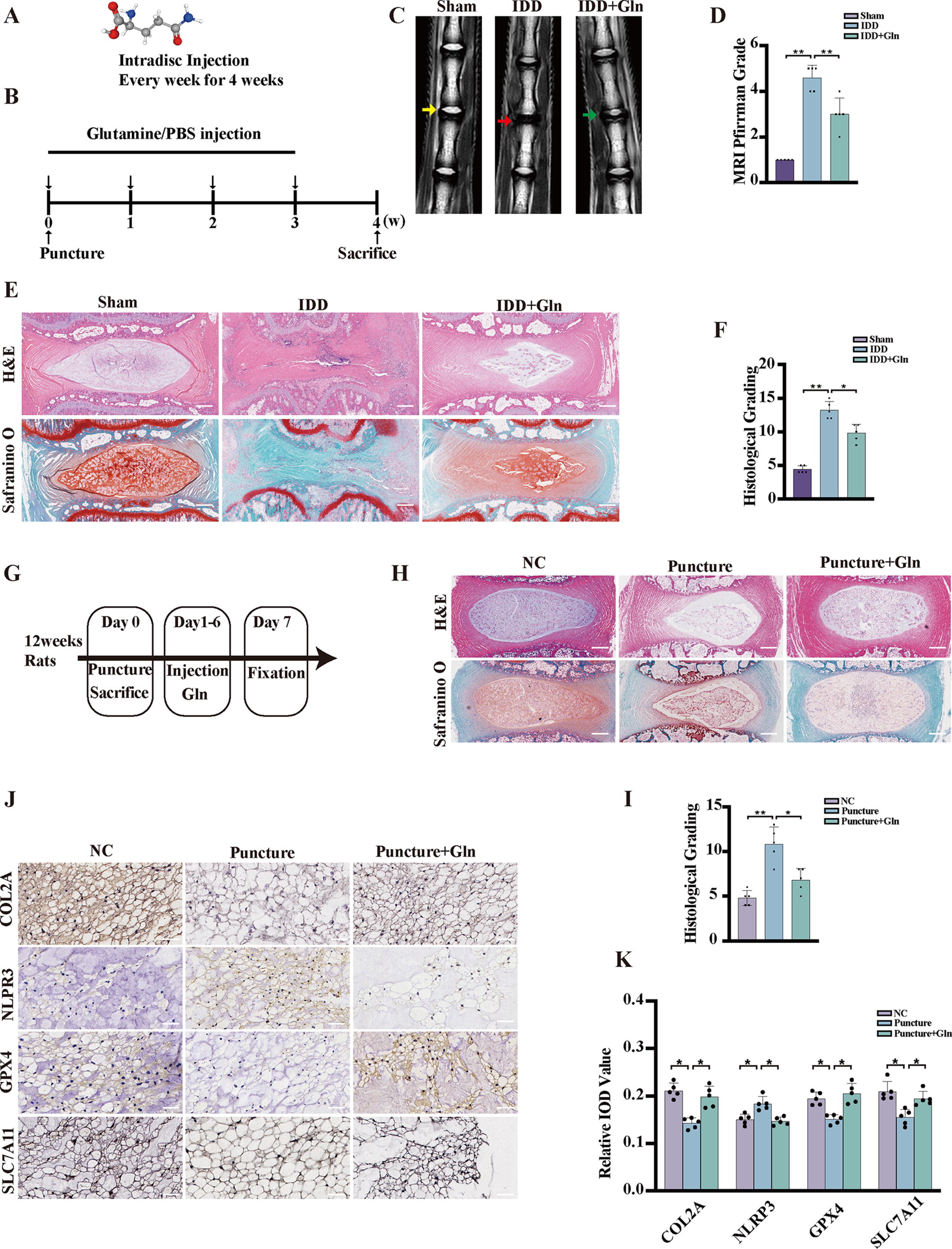
Notably, glutamine, the most abundant free amino acid in both plasma and intracellular amino acid pools (Fig. 3A), exhibits a critical function in ROS clearance by functioning as a precursor or facilitating GSH synthesis (Chen et al., 2016, DeBerardinis and Cheng, 2010). To elucidate the involvement of glutamine in the progression of IDD, a rat model of IDD was induced through needle puncture (Fig. 3B). MRI assessments revealed that the T2-weighted signals of IVDs in rats subjected to PBS treatment were notably diminished compared with those in rats administered with glutamine injections (Fig. 3C, D). Histological evaluations employing H&E staining and SO&FG staining demonstrated substantial mitigation of IVD histological degeneration with glutamine treatment (Fig. 3E, F). These findings collectively suggest that the progression of IDD may be subject to regulation by glutamine. An ex vivo experiment was conducted to further investigate the effect of glutamine on IDD puncture models (Fig. 3G). H&E staining and Safranin O staining were performed to examine the IVD structure in the puncture group. A notable feature observed in these degenerative discs was the loss of distinct demarcation between NP and AF tissues. In degenerating discs, this boundary was less discernible, indicating structural changes and compromised disc integrity. However, the administration of glutamine treatment conferred protective effects on IVDs (Fig. 3H, 2). Furthermore, the results obtained from an IHC analysis of the glutamine group indicated an increase in the levels of the ECM component COL2A compared with that in the puncture group. Lower levels of NLRP3 were observed in the glutamine group than in the puncture group. Notably, the levels of ferroptosis biomarkers, including SLC7A11 and GPX4, were significantly decreased in the puncture group and increased in the glutamine group (Fig. 3J, K). Our findings suggest a potential regulatory role for glutamine in protecting ferroptosis and pyroptosis pathways in IDD.
Glutamine protects NP cells from TBHP-induced ECM degradation and pyroptosis in vitro
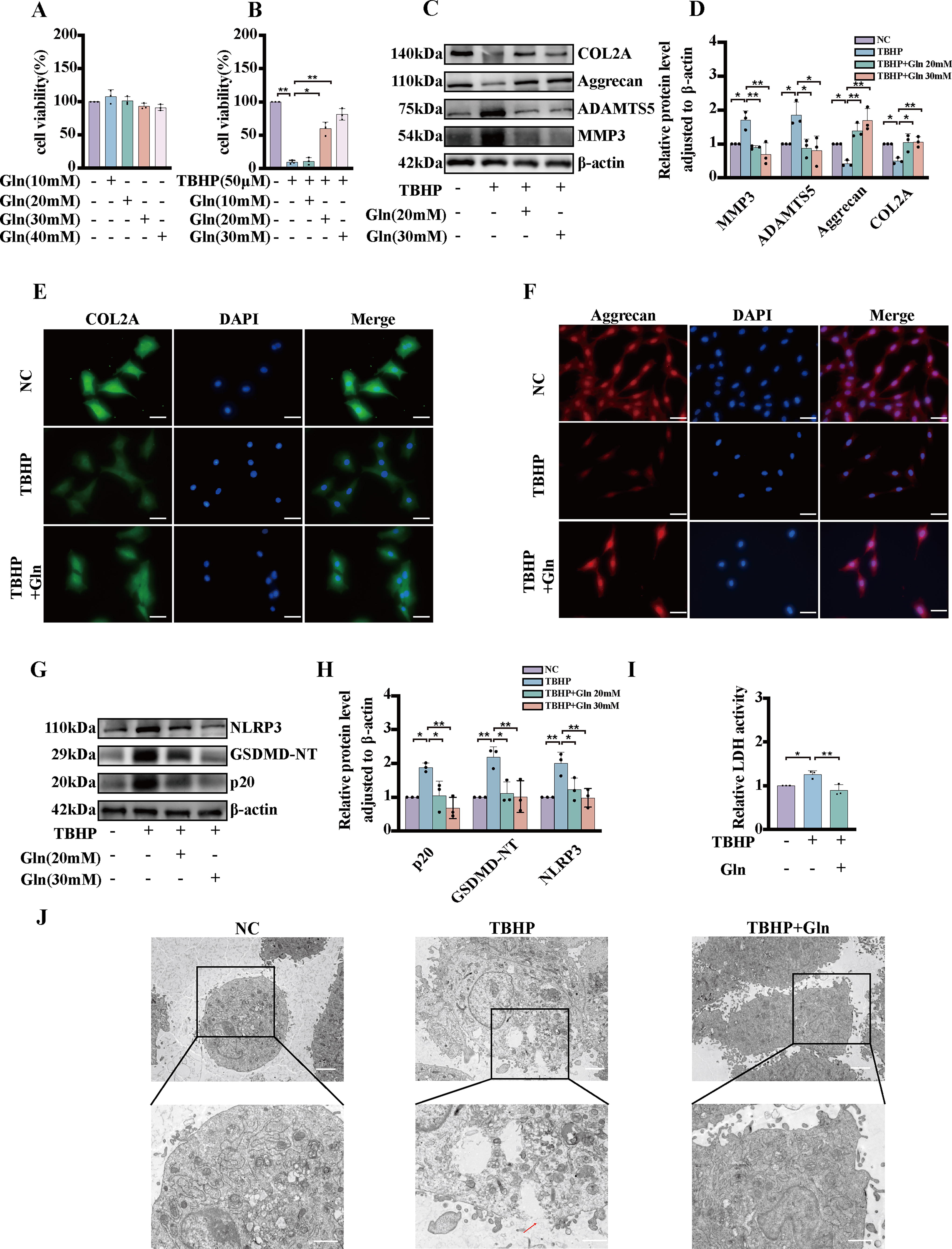
Our findings revealed that glutamine treatment did not exert a discernible effect on cell viability when administered at a concentration of <40 mM for 24 h (Fig. 4A). Furthermore, our results indicated that treatment with 20 mM glutamine conferred protection against TBHP (Fig. 4B). The degeneration of NP cells is positively associated with ECM metabolism. In the present study, TBHP treatment significantly reduced the synthesis of COL2A and Aggrecan. Additionally, TBHP treatment led to an increase in the expression levels of MMP3 and ADAMTS5. However, these deleterious alterations induced by TBHP were largely reversed by pretreatment with glutamine (Fig. 4C, D). In addition, immunofluorescence assay results revealed that glutamine significantly attenuated the TBHP-induced degeneration of COL2A and Aggrecan (Fig. 4E, F). Supplementation with exogenous glutamine was a crucial factor in the restoration of the metabolic balance after samples were subjected to TBHP-mediated oxidative stress. IDD is a degenerative age-related ailment that manifests by oxidative stress and pyroptotic NP cells. Notably, glutamine treatment has shown promise in ameliorating TBHP-induced pyroptosis in NP cells. To substantiate this purported effect, Western blot analysis was performed to assess the impact of glutamine treatment on the TBHP-triggered expression of pyroptosis-related proteins. The results demonstrated that glutamine treatment effectively attenuated the TBHP-induced increase in the expression of NLRP3 cleaved caspase-1 and GSDMD-N, which are associated with pyroptosis (Fig. 4G, H). Additionally, glutamine treatment successfully counteracted the excessive secretion of LDH observed after TBHP exposure, as corroborated by LDH assay (Fig. 31). Furthermore, TEM was employed to evaluate the morphological changes induced by NP cells undergoing pyroptosis. Our results indicated that after TBHP treatment, cells were swollen, organelles, including the nucleus, disappeared, and cell membranes developed pores and ruptured. However, pretreatment with glutamine inhibited these pyroptotic characteristics of NP cells treated with TBHP (Fig. 4J).
Glutamine protects NP cells from TBHP-induced ferroptosis in vitro
To investigate the protective effect of glutamine against ferroptosis, we performed lipid ROS, MitoSOX, and MitoTracker staining with NP cells. Our results indicated that glutamine treatment inhibited the TBHP-induced downregulation of the ferroptosis-associated proteins, GPX4 and SLC7A11 (Fig. 5A, B). To evaluate the ferroptosis rate, NP cells were incubated with the DCFH-DA probe. The results demonstrated that TBHP treatment of NP cells led to an elevation in ROS levels, indicating that the cells underwent ferroptosis. However, this increase in ROS levels was effectively counteracted by glutamine treatment (Fig. 5C, D). Furthermore, our findings revealed that TBHP treatment increased lipid ROS levels, a hallmark of ferroptosis. Again, glutamine successfully attenuated the increase in lipid ROS levels induced by TBHP treatment (Fig. 5E–F). Moreover, TBHP exposure significantly increased mitochondrial ROS levels and mitochondrial membrane damage in NP cells, but glutamine treatment inhibited these effects (Fig. 5G–H). Glutamine pretreatment has been previously demonstrated to upregulate the expression of GPX4 in NP cells (Fig. 41). TEM provided evidence showing internal vesicle formation and crista loss in affected mitochondria. Notably, the deleterious effects of TBHP treatment on NP cells were attenuated by glutamine treatment (Fig. 5J).
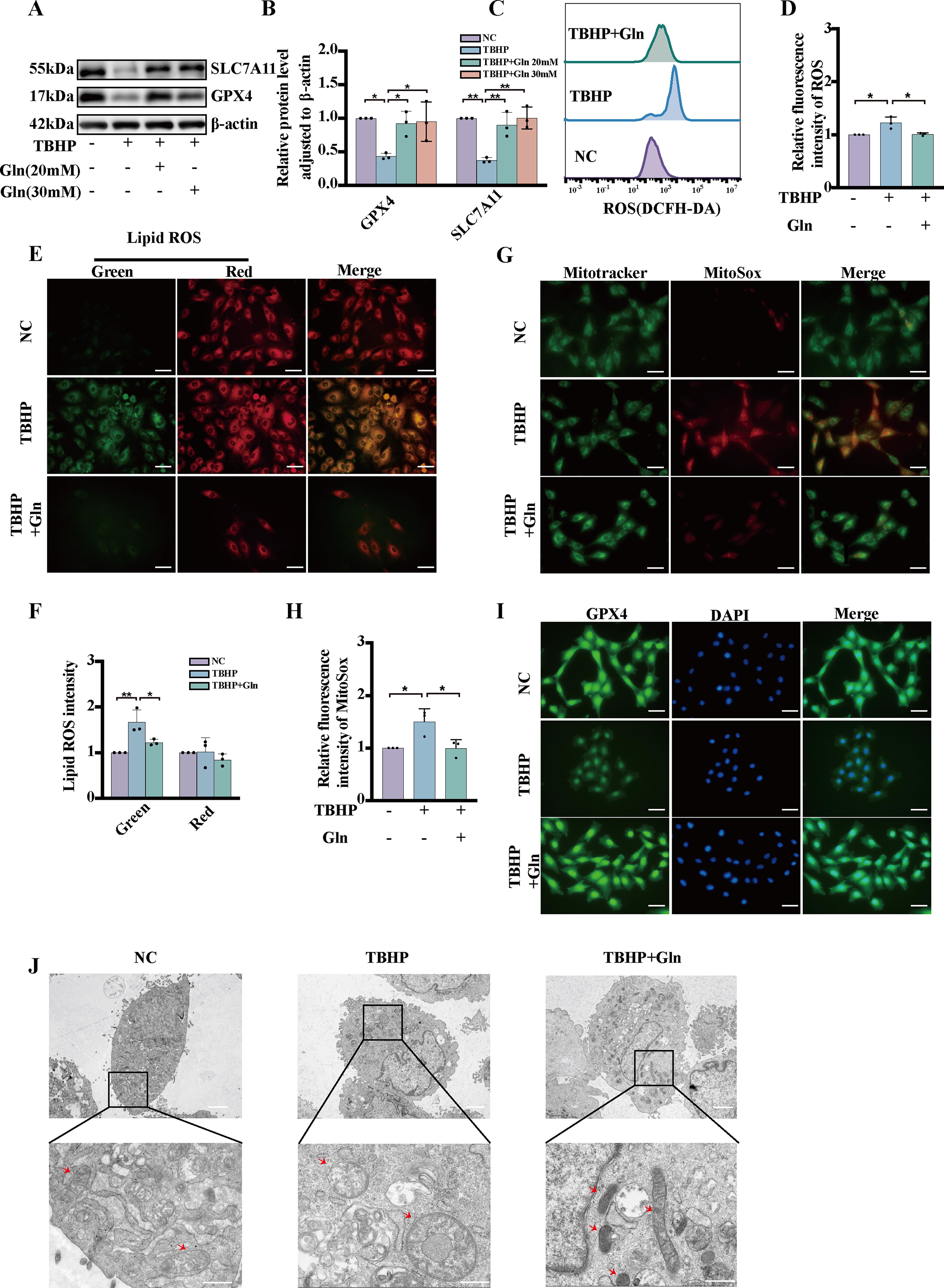
Our findings suggest that glutamine inhibits TBHP-mediated ferroptosis in NP cells by regulating mitochondrial function and subsequent ROS production.
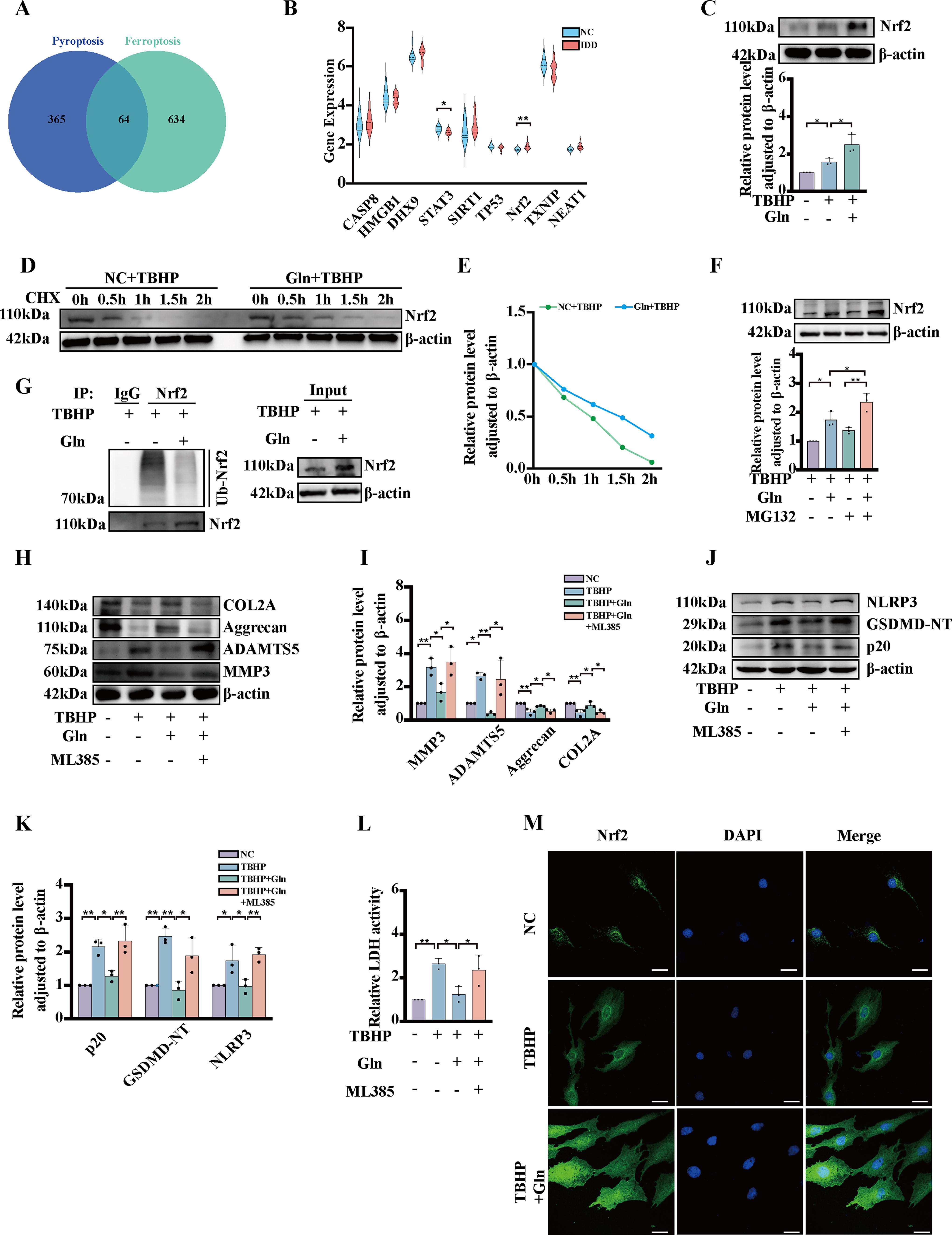
Glutamine promotes Nrf2 accumulation by inhibiting its ubiquitin–proteasome degradation and activating antipyroptosis signaling
For analysis, 365 pyroptosis-related genes were obtained from GeneCards. Furthermore, a separate set of 634 ferroptosis-related genes was obtained from FerrDb. By comparing the two lists, 64 common cell death-related genes were identified and selected for further analysis (Fig. 6A). The ten most differentially expressed genes among the 64 common genes are visualized in a violin plot constructed with data from GSE27494 (Fig. 6B). The results indicated that Nrf2 showed the highest differential expression between the two groups. In vitro, experiments showed that the protein levels of Nrf2 were increased by glutamine treatment (Fig. 6C). Additionally, to investigate the mechanism underlying the glutamine-mediated upregulation of Nrf2, cycloheximide (CHX) chase experiments were conducted. Our results suggest that glutamine extended the half-life of Nrf2 (Fig. 6D, E). Following treatment with MG132, a proteasome inhibitor, the protein level of Nrf2 was increased, particularly in the glutamine-treated group (Fig. 6F). Furthermore, a ubiquitination assay showed that TBHP increased the ubiquitination rate of Nrf2, while glutamine delayed Nrf2 ubiquitination and degradation (Fig. 6G). These findings provide evidence that glutamine inhibits the degradation of Nrf2 by blocking the ubiquitin–proteasome pathway. This inhibitory effect allows Nrf2 to accumulate and thus exert its protective effects. In the context of NP cells, Nrf2 has been found to contribute significantly to the protective effects of glutamine against TBHP-induced pyroptosis as well as ECM degradation. To investigate the specific role of Nrf2 in mediating the protective effects of glutamine, we used the small-molecule inhibitor of Nrf2, ML385. A Western blot analysis revealed that glutamine treatment significantly suppressed the levels of ADAMTS5, MMP3, NLRP3 GSDMD-NT, and p20 while markedly decreasing the levels of COL2A and Aggrecan in the presence of TBHP. In contrast, ML385 treatment reversed the protective effects of glutamine (Fig. 6H–K). In addition, ML385 treatment notably increased the level of LDH with the addition of both TBHP and glutamine (Fig. 6L). Furthermore, glutamine increased the nuclear translocation rate and thus the level of Nrf2 in the nucleus (Fig. 6M). These results indicate that glutamine protects against TBHP-triggered pyroptosis and ECM degradation via Nrf2 signaling pathway activation.
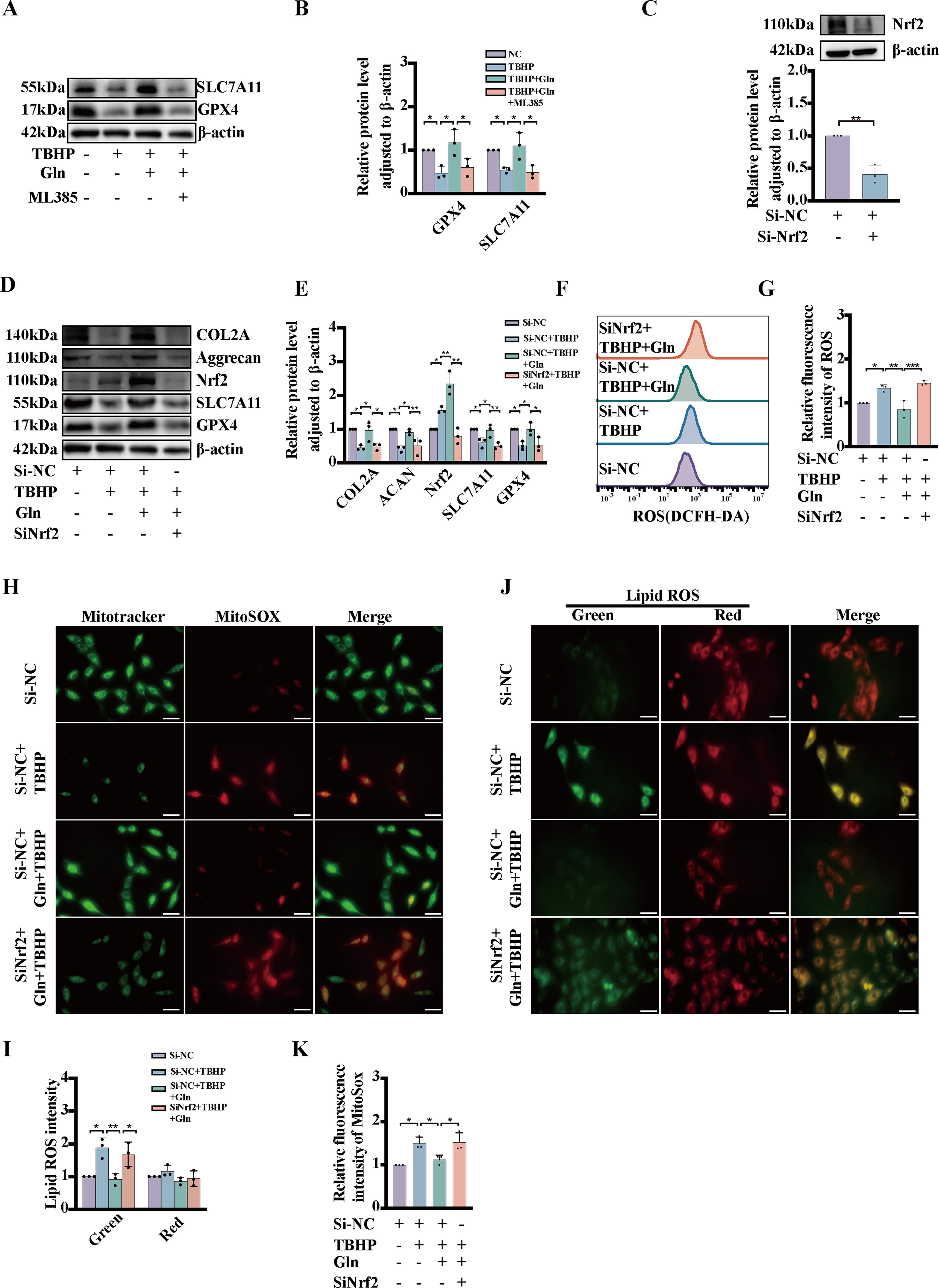
Glutamine protects NP cells from TBHP-induced ferroptosis via Nrf2 signaling activation
Our findings indicate that ML385 caused a decrease in the expression of ferroptosis-associated proteins in NP cells compared with cells treated with both glutamine and TBHP (Fig. 7A, B). To elucidate the functional role of Nrf2 in ferroptosis, we employed siRNA to knock down Nrf2. The efficacy of Nrf2 knockdown was assessed by measuring the protein expression levels of Nrf2 (Fig. 7C). Knockdown of Nrf2 also decreased the protein levels of COL2A, Aggrecan, SLC7A11, and GPX4 after TBHP and glutamine stimulation, as measured by Western blotting (Fig. 7D, E). A flow cytometry analysis revealed that glutamine regulated TBHP-induced changes in cellular ROS levels via Nrf2 activation (Fig. 7F, G). In addition, we found that Nrf2 knockdown led to increased lipid ROS and MitoSOX levels, indicating the exacerbation of ferroptosis damage (Fig. 7H–K). These findings suggest that glutamine protects against TBHP-induced ferroptosis by increasing the level of Nrf2.
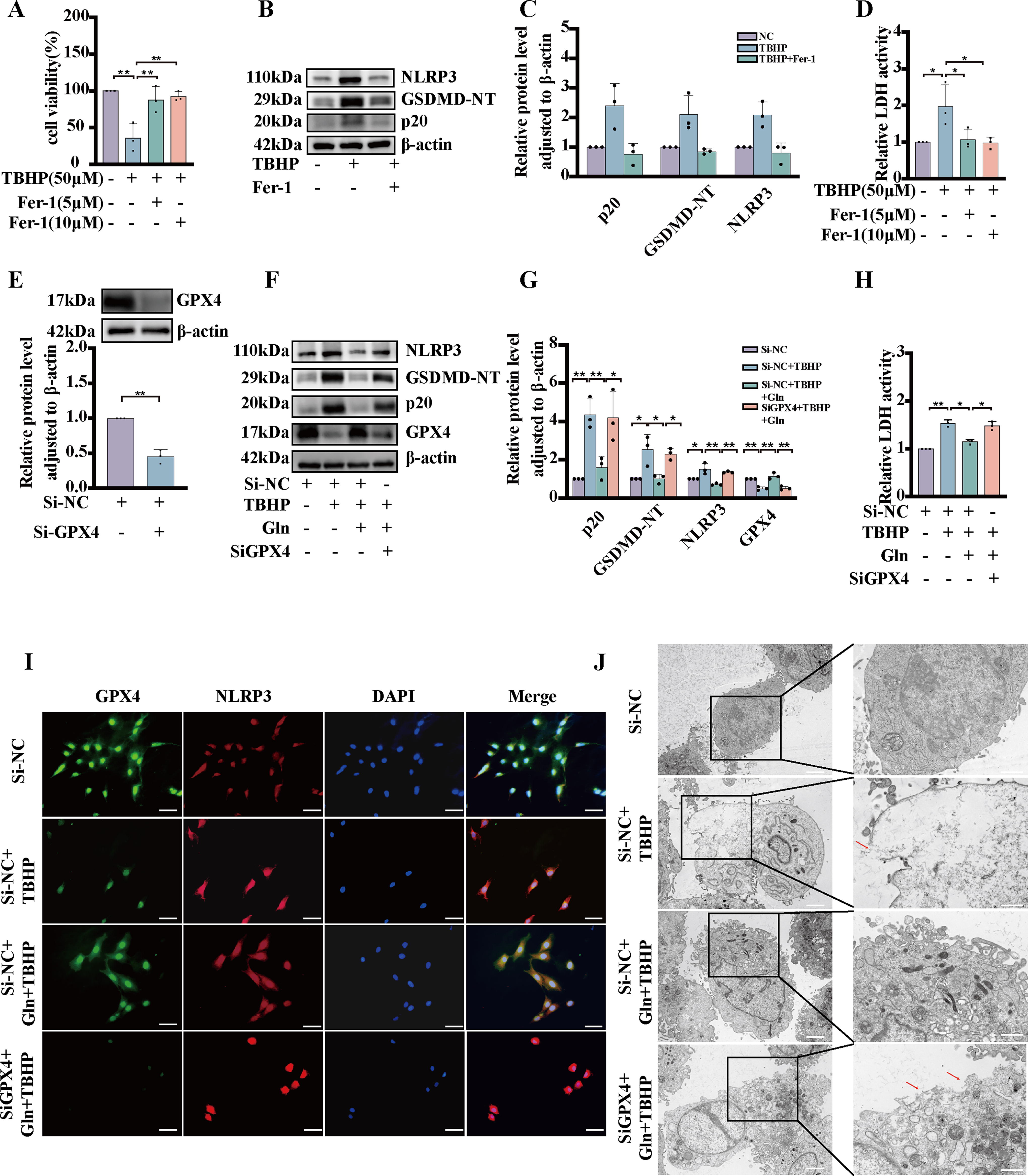
Glutamine attenuates TBHP-induced pyroptosis in NP cells by inhibiting lipid oxidation
Excessive production of ROS has been established as one of the contributing factors to the induction of pyroptosis. (Horng, 2014). In addition, it has been reported that products resulting from lipid oxidation can inhibit the NLRP3 inflammasome (Hsu et al., 2022). Based on previous results (Fig. 2H–J), it is evident that pyroptosis synergistically interacts with ferroptosis in human samples. Consequently, we conducted a series of experiments aimed at elucidating the role of ferrostatin-1, a small-molecule inhibitor of lipid oxidation, in the context of pyroptosis. First, we performed a CCK-8 assay to measure NP cell viability with ferrostatin-1, in the presence of TBHP. Our results indicated that ferrostatin-1 treatment increased the viability of TBHP-induced NP cells (Fig. 8A). Our Western blot data demonstrated that ferrostatin-1 treatment decreased the expression of NLRP3 GSDMD-NT, and p20 in NP cells compared with the control cells (Fig. 8B, C). In addition, the addition of ferrostatin-1 notably decreased the rate of LDH release in the presence of TBHP (Fig. 8D). To investigate the role of the ferroptosis biomarker protein GPX4 in pyroptosis, we used siRNA to knock down GPX4 and measured the efficacy of the knockdown by measuring the protein expression of GPX4 (Fig. 8E). Our findings revealed that the GPX4 knockdown increased the protein levels of NLRP3 GSDMD-NT, and p20 following TBHP and glutamine stimulation, as measured by Western blotting (Fig. 8F, G). Furthermore, GPX4 knockdown significantly increased the rate of LDH release in the presence of TBHP and glutamine (Fig. 8H). Our findings indicate that knocking down GPX4 resulted in elevated NLRP3 levels (Fig. 71). Moreover, our TEM results demonstrated that NP cells with GPX4 knocked down exhibited pyroptosis characteristics following stimulation with TBHP and glutamine (Fig. 8J). Our data suggest that glutamine can attenuate TBHP-induced pyroptosis by inhibiting lipid oxidation.
Footnotes
Author Disclosure Statement
The authors declare that they have no conflicts of interest.
Funding Information
This study was supported by the National Natural Science Foundation of China (No. 82372437 and No. 82202653), the Natural Science Foundation of Guangdong Province (No. 2020A1515011538, No. 2020A1515010060, No. 2021A1515010345, No. 2024A1515012963, and No. 2024A1515010719), the International Science & Technology Cooperation Program of Guangdong Province (No. 2023A0505050136), the Yat-sen Qihang Foundation of Sun Yat-sen Memorial Hospital (No.YXQH202204).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.