
Abstract
Aims:
In addition to reducing the respiratory function, crystalline silica (SiO2) disturbs the immune response by affecting immune cells during the progression of silicosis. Regulatory T cell (Treg) differentiation may play a key role in the abnormal polarization of T helper cell (Th)1 and Th2 cells in the development of silicosis-induced fibrosis. Alpha-lipoic acid (ALA) has immunomodulatory effects by promoting Tregs differentiation. Thus, ALA may have a therapeutic potential for treating autoimmune disorders in patients with silicosis. However, little is known regarding whether ALA regulates the immune system during silicosis development.
Results:
We found that the expression levels of collagen increased, and the antioxidant capacity was lower in the Lias−/− +SiO2 group than in the Lias +/+ +SiO2 group. The proportion of Tregs decreased in the peripheral blood and spleen tissue in mice exposed to SiO2. The proportion of Tregs in the Lias−/− +SiO2 group was significantly lower than that in the Lias+/+ +SiO2 group. Supplementary exogenous ALA attenuates the accumulation of inflammatory cells and extracellular matrix in lung tissues. ALA promotes the immunological balance between Th17 and Treg responses during the development of silicosis-induced fibrosis.
Innovation and Conclusion:
Our findings confirmed that low expression of lipoic acid synthase aggravates SiO2-induced silicosis, and that supplementary exogenous ALA has therapeutic potential by improving Tregs in silicosis fibrosis.
Introduction
Silicosis is a systemic disease characterized by pulmonary interstitial fibrosis due to the inhalation of crystalline silica (SiO2). Silicosis can decrease quality of life or even cause death, resulting in a heavy financial burden for the patient (Riley and Urbine, 2019). However, the pathogenesis of silicosis remains unclear and treatment remains difficult (Budden et al., 2019; Lederer and Martinez, 2018). Although Pirfenidone and Nidanib can improve respiratory function in patients with idiopathic pulmonary fibrosis, it is uncertain whether they have therapeutic effects (Olenski et al., 2020).
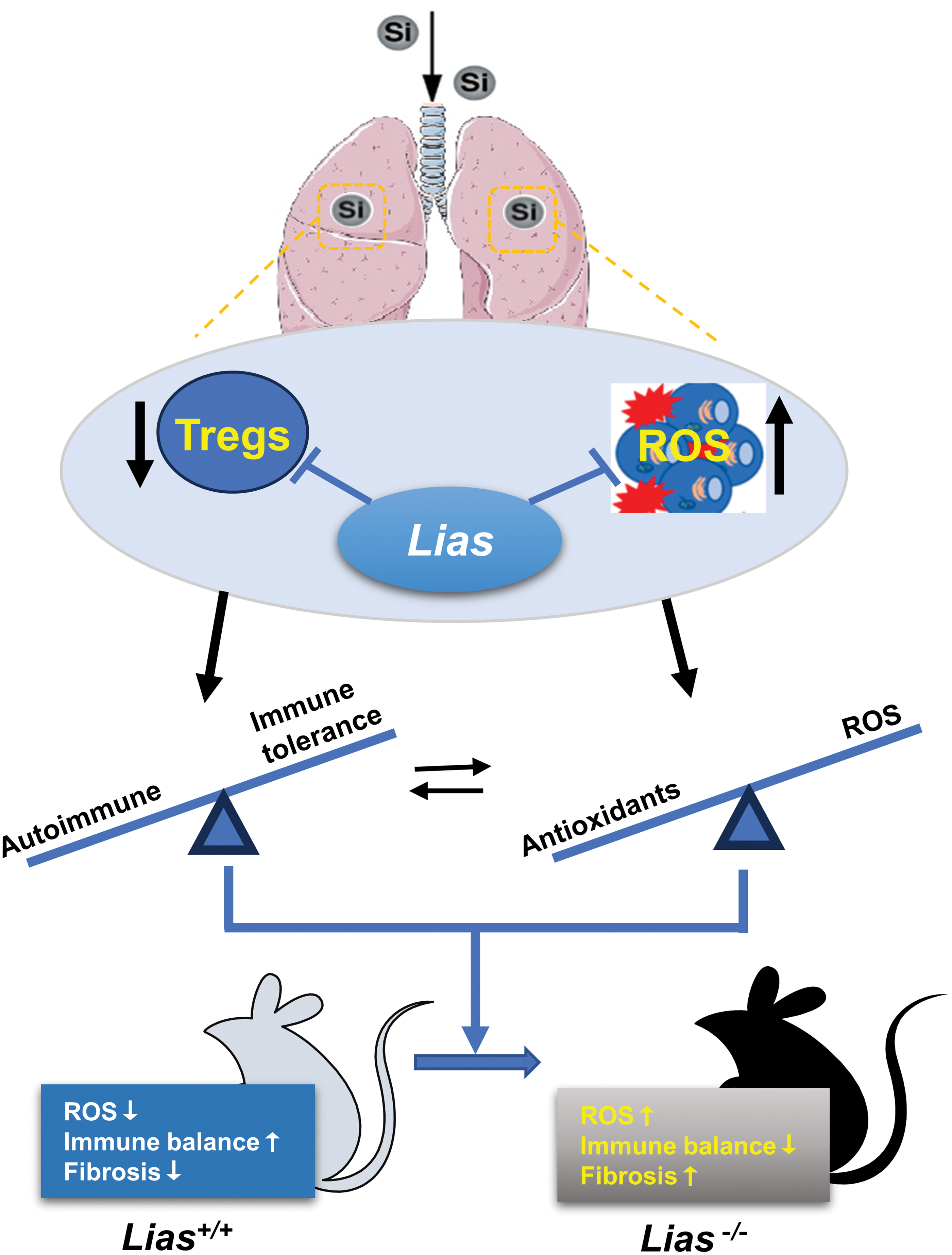
In addition to reducing respiratory function, crystalline silica disturbs immune responses by affecting immune cells, including macrophages, B cells, T cells, and NK cells, during the progression of silicosis-associated fibrosis (Marrocco et al., 2021). Crystalline silica exposure can decrease the proportion of regulatory T cells (Tregs), activate T cell status, and break immunological tolerance against autoantigens (Brilland et al., 2019).
Epidemiological studies have shown that levels of rheumatic factors, antinuclear antibodies, and other autoantibodies are increased in patients with silicosis (Lee et al., 2017; Mayeux et al., 2018). Autoimmune diseases, such as rheumatoid arthritis, systemic sclerosis, and systemic lupus erythematosus, have higher incidence rates in occupationally exposed workers (Ballerie et al., 2020; Ferri et al., 2018).
Innovation
We first used the Lias −/− mice to investigate the role of LIAS in the immune response of silicosis fibrosis. And we found that low expression of LIAS aggravates silica-induced pulmonary fibrosis by inhibiting the differentiation of Tregs in mice.
Silica leads to a disorder in immunomodulatory functions during silicosis fibrosis (Wei et al., 2021). Abnormal polarization of T helper cell (Th)1 and Th2 cells is a characteristic feature of inflammation and fibrosis during silicosis development (Bao et al., 2018). When silica particles are inhaled in the alveolus, they are first engulfed by macrophages to generate excess reactive oxygen species (ROS), and the immune responses are activated through the cytokines such as interleukin (IL)-1β secreted by macrophages (Rivas-Arancibia et al., 2022; Tandon et al., 2020). Silica particles also stimulate macrophages and dendritic cells to initiate antigen processing and presentation pathways, thus activating T lymphocytes (Suzuki et al., 1996).
The transformation of initial CD4+ T cells into Th1 cells is promoted by the stimulation of macrophages by silica particles; subsequently, pulmonary inflammation is initiated by IL-12 and interferon-γ (IFN-γ) secreted by Th1 cells (Wu et al., 2018). As inflammation progresses, Th2 cells from the initial CD4+ T cells are activated to inhibit Th1 cell responses, and promote pulmonary fibrosis by secreting IL-4 and IL-13 (Unutmaz et al., 2010). Silicosis fibrosis can be alleviated by maintaining a balance between Th1/Th2 cell responses in the lungs (Dai et al., 2016).
Tregs are a subpopulation of cells differentiated from the initial CD4+T cells, which can promote immune tolerance and suppress immune responses to maintain cellular immune homeostasis (Dembic, 2019). Tregs inhibit Th1 cell responses by secreting transforming growth factor-β (TGF-β) and IL-10, which can reduce lung inflammation and maintain the balance of Th1/Th2 cells during the initial stage of silicosis fibrosis (Dai et al., 2016). However, the number of Tregs decreases in the bronchoalveolar lavage fluid and peripheral blood of patients with silicosis (Hayashi et al., 2010). Georgi, a Chinese medicinal herb, reduces silicosis-induced fibrosis by increasing the population of Tregs (Liu et al., 2015).
Autoimmune responses are induced by damage to autoreactive lymphocytes (Alissafi et al., 2020). The immunoregulatory efficacy of Tregs is reduced by the oxidative stress induced by chronic exposure to environmental toxicants (Chavez and Tse, 2021). Antioxidants may play a critical role in maintaining the compensatory balance of the inflammatory responses between Tregs and Th17 cells (Chavez and Tse, 2021). Alpha-lipoic acid (ALA) is an efficient antioxidant for small molecules generated in mitochondria that are both lipid- and water soluble (Solmonson and DeBerardinis, 2018).
ALA regulates the balance of Th17/Tregs and enhances the Nrf2/HO-1 signaling pathway in allergic disease (Van Nguyen et al., 2020). ALA inhibits autoimmune responses by decreasing the population of Th1 cells, and inhibiting the differentiation of Tregs in spontaneous diabetic and autoimmune recurrent mice (Huang et al., 2022; Solmonson and DeBerardinis, 2018). Overexpression of lipoic acid synthase (LIAS) increased the population of Tregs and inhibited inflammatory responses in APOE −/− mice (Tian et al., 2020).
ALA has been reported to have an immunomodulatory effect, and may therefore have therapeutic potential in autoimmune disorders in patients with silicosis. We previously found that ALA can reduce silicosis fibrosis in mice (Cui et al., 2009; Li et al., 2018). However, little is known about whether ALA, which enhances the antioxidant capacity in mice, can regulate immune responses during the development of silicosis.
In particular, the mechanism by which ALA affects the function of Th lineages during silicosis progression remains unclear. We hypothesized that ALA promotes Treg production and plasticizes Th cell subsets by influencing the redox microenvironment during the progression of silicosis-induced fibrosis. In this study, we found that ALA significantly increased Tregs in the circulation and significantly decreased lymphocyte infiltration during the development of silicosis. These results suggest that ALA alleviates silicosis by modulating immune responses (Fig. 1).
Results
The degree of SiO2-induced pulmonary fibrosis was aggravated in Lias−/− mice
We identified the mouse genotypes using polymerase chain reaction (PCR). A single band of 500 bp appeared in Lias+/+ mice, a single band of 300 bp appeared in Lias−/− mice, and the DNA ladders in Lias+/− mice comprised of two bands of 300 and 500 bp (Fig. 2C). Pulmonary LIAS protein levels were measured using Western blotting. Semiquantitative Western blotting showed that pulmonary LIAS levels in the Lias−/− +saline group were lower than those in the Lias+/+ +saline group. LIAS levels were significantly lower in the Lias−/− +SiO2 group than in the Lias+/+ +SiO2 group (Fig. 2D, E).
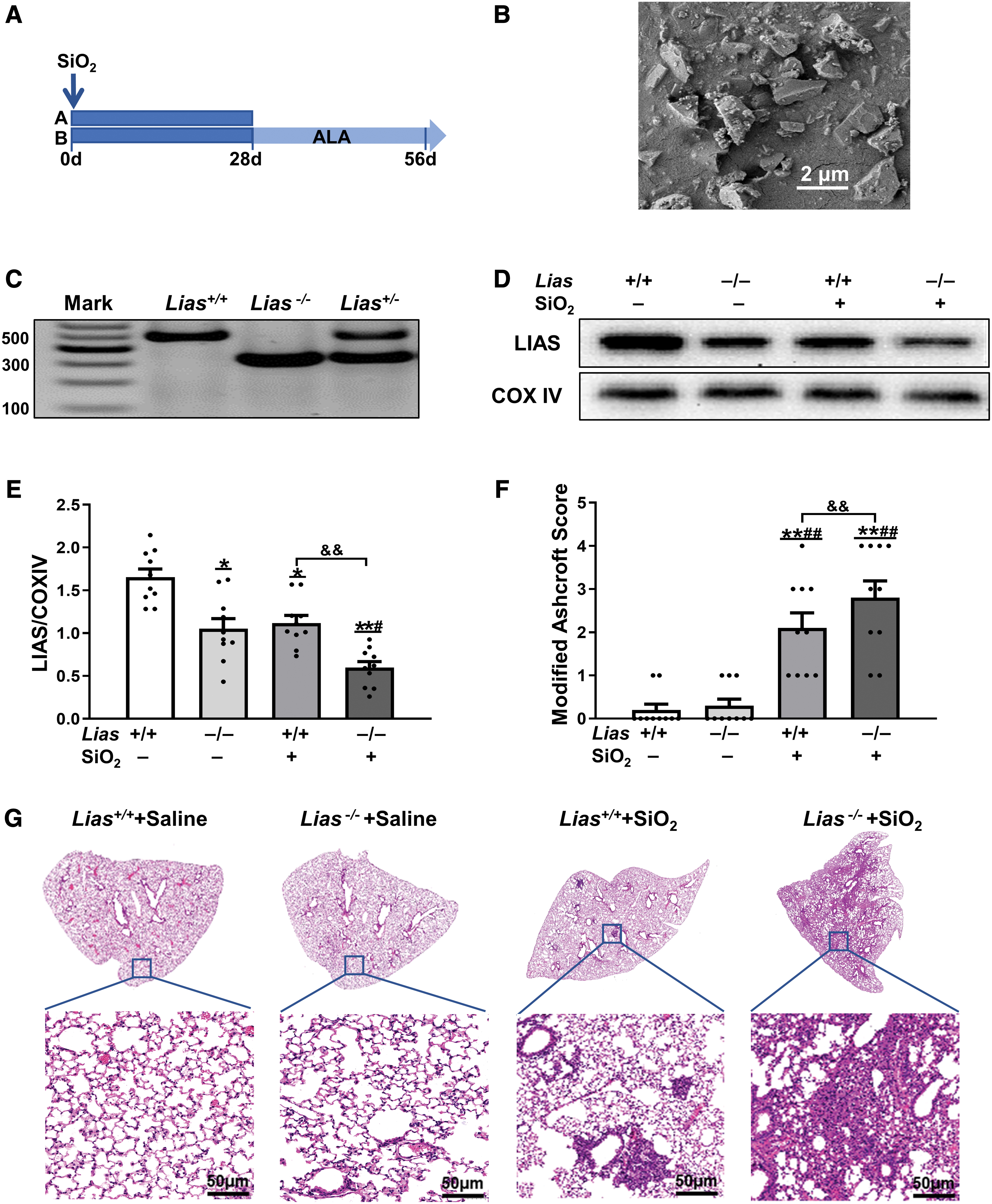
Obvious lung inflammation and cellular nodules were observed in mice exposed to silica in hematoxylin and eosin (H&E)-stained sections. The areas of inflammation and fibrosis in the lung tissue of Lias−/− +SiO2 group were greater than those in Lias−/− +SiO2 group. The degree of fibrosis in mice was determined by measuring the Ashcroft score. The result showed that The Ashcroft score of the Lias−/− +SiO2 group was significantly higher than that of Lias+/+ +SiO2 group (Fig. 2F, G).
Low expression of LIAS increased collagen fiber production and promotes epithelial–mesenchymal transformation induced by SiO2 in Lias−/− mice
We examined the production of collagen fibers in the lungs of mice using Masson staining. In two groups exposed to SiO2, collagen deposition and cellular nodules were observed. Compared with the Lias+/+ SiO2 group, the Lias−/− +SiO2 group showed more areas of collagen expression (Fig. 3A, B).
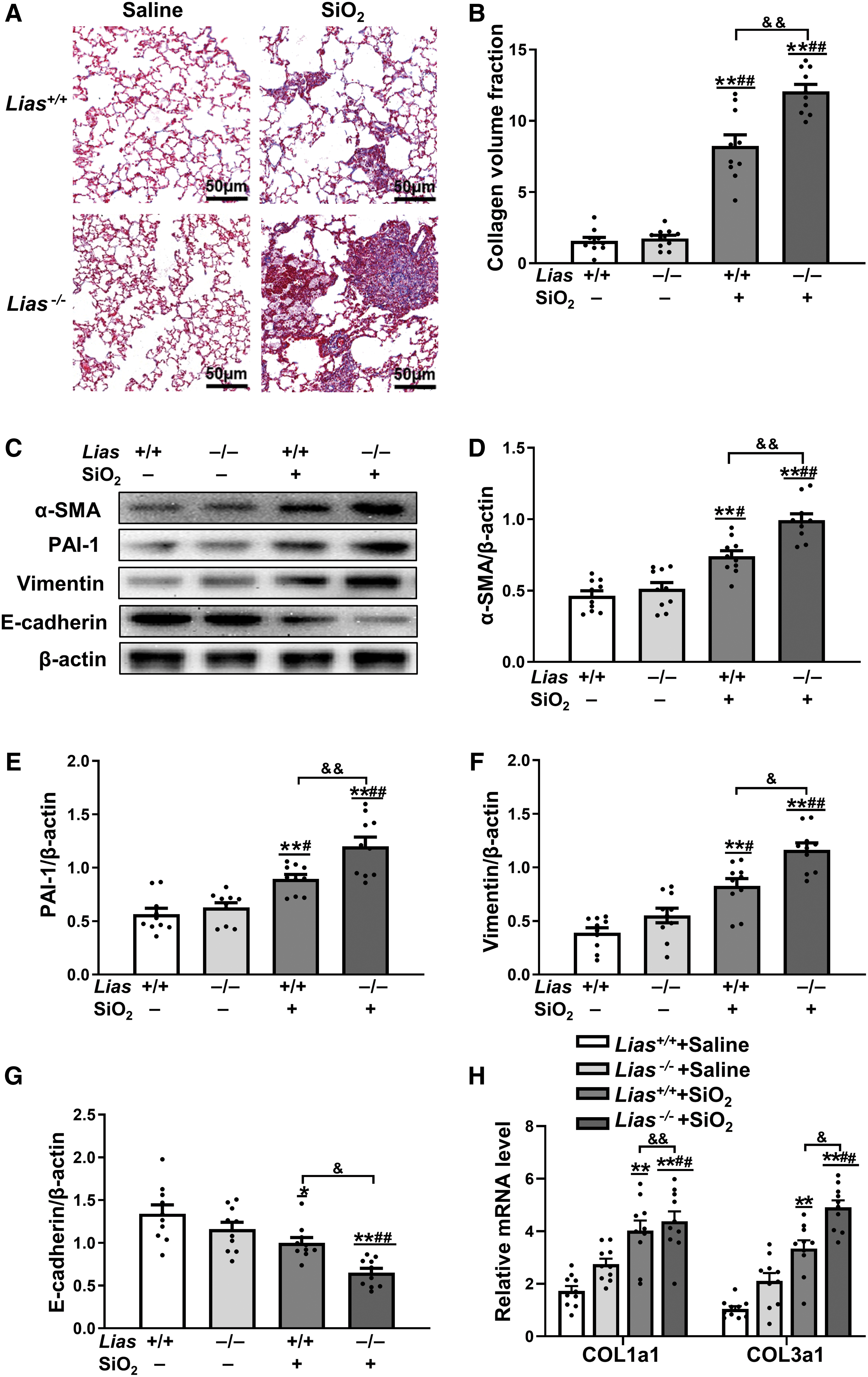
The expression of α-smooth muscle actin (α-SMA), vimentin, and E-cadherin in the lungs of the mice was measured using Western blotting to evaluate the degree of epithelial–mesenchymal transition (EMT). The results showed that the expression of α-SMA and vimentin in mice exposed to SiO2 was higher than that in matched control mice. The expression of E-cadherin, an epithelial cell marker, in mice exposed to SiO2 was lower than that in matched control mice. The expression levels of vimentin and α-SMA in the Lias−/− +SiO2 group were significantly higher than those in the Lias+/+ +SiO2 group.
Furthermore, E-cadherin expression was significantly lower in the Lias−/− +SiO2 group than in Lias+/+ +SiO2 group. In addition, plasminogen activator inhibitor 1 (PAI-1), the main inhibitor of tissue-type plasminogen activator, was expressed at higher levels in SiO2-exposed mice. PAI-1 expression in the Lias−/− +SiO2 group was significantly lower than that in Lias+/+ +SiO2 group (Fig. 3C–G).
According to the quantitative polymerase chain reaction (qPCR) results, the mRNA expression levels of type I and type III collagen in mice exposed to SiO2 were higher than those in matched control mice. The expression levels of type I and type III collagen in the Lias−/− +SiO2 group were significantly lower than those in Lias+/+ +SiO2 group (Fig. 3H).
Low expression of LIAS enhances oxidative stress and promotes the damage to mitochondrial function during SiO2-induced fibrogenesis
The total antioxidant capacity (TAC) of serum was determined (Fig. 4A). Total antioxidant levels were lower in SiO2-exposed mice than in mice that received saline, and a significant reduction in total antioxidants was observed in the Lias−/− +SiO2 group. Levels of catalase (CAT), superoxide dismutase (SOD), and malondialdehyde (MDA) were measured in the lung tissue. Similarly, the levels of serum SOD, CAT, and glutathione (GSH) in SiO2-exposed mice were decreased, and the serum MDA content was significantly increased compared with the matched control group mice.
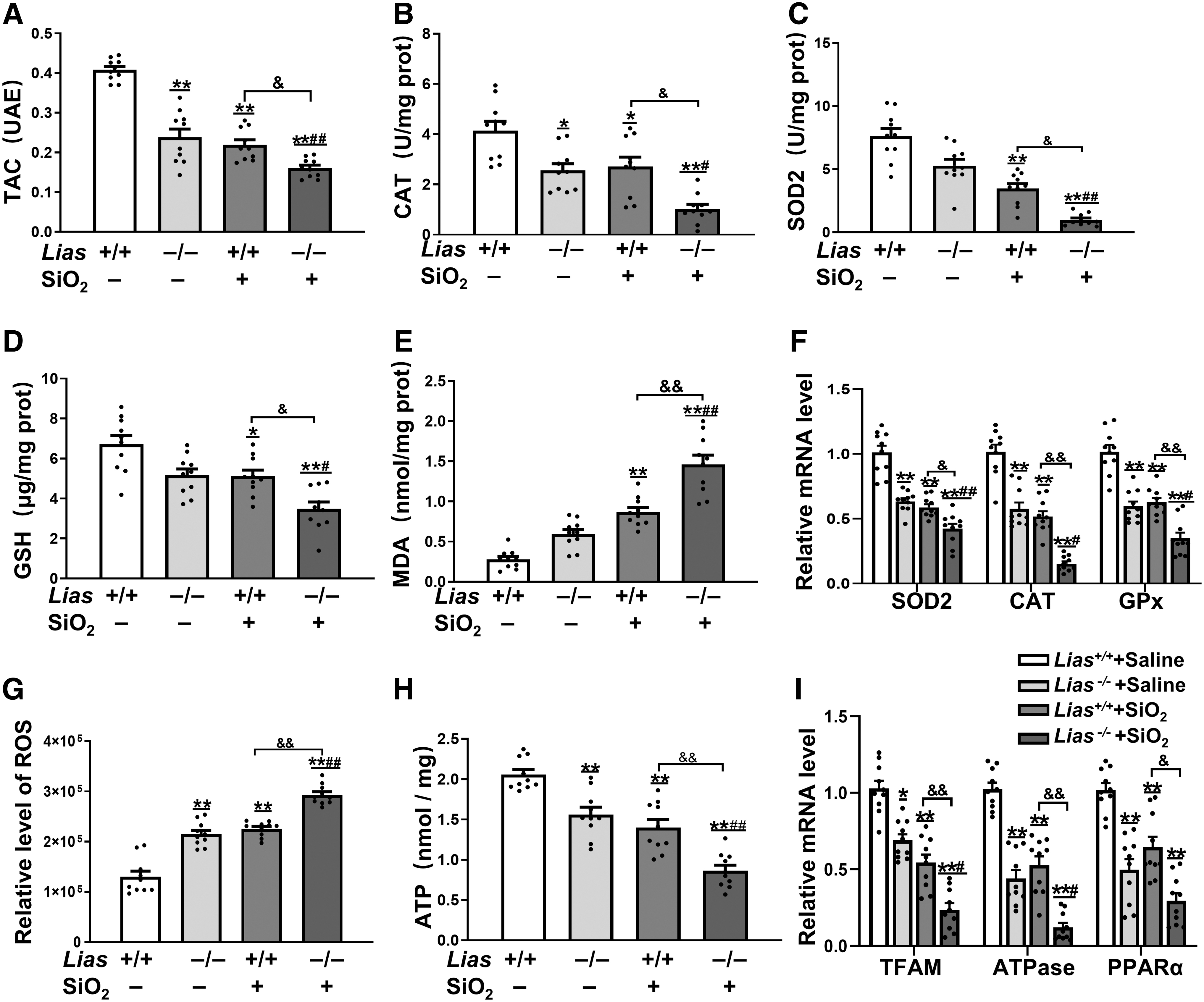
There was a significant reduction in SOD, CAT, and GSH levels and a significant increase in MDA in the Lias−/− +SiO2 group compared with those in the Lias+/+ +SiO2 group (Fig. 4B–E). qPCR results showed that the levels of CAT, SOD2, and glutathioneperoxidase (GPx) mRNA were lower in SiO2-exposed mice than in mice that received saline. A significant reduction in SOD2, CAT, and GPx mRNA levels was observed in the Lias−/− +SiO2 group compared with the Lias+/+ +SiO2 group (Fig. 4F).
ROS levels in the lung tissues were evaluated using an immunofluorescence assay. The results showed that the pulmonary ROS levels in mice exposed to SiO2 were higher than those in matched control mice (Fig. 4G). There was a significant increase in pulmonary ROS levels in the Lias−/− +SiO2 group compared with those in the Lias+/+ +SiO2 group. We also measured adenosine triphosphate (ATP) levels in lung tissue using a chemiluminescence kit.
The results showed decreased levels of ATP in the lung tissues of the two groups of mice exposed to SiO2, and a significant reduction in ATP levels in the Lias−/− +SiO2 group compared with the Lias+/+ +SiO2 group (Fig. 4H). In addition, the qPCR results showed that the expressions of mitochondrial transcription factor A (TFAM), ATPase, peroxisome proliferators-activated receptor alpha (PPARα), and COXIV mRNA in SiO2-exposed mice were lower than those in matched control mice, and there was a significant reduction of the expressions in the Lias−/− +SiO2 group compared with the Lias+/+ +SiO2 group (Fig. 4I). Low expression of LIAS enhances ROS production and promotes mitochondrial damage induced by silica in the lung tissue.
Low expression of LIAS inhibits the differentiation of Tregs and exacerbates the imbalance of Th1/Th2 during SiO2-induced fibrogenesis
To further investigate the effects of low LIAS expression on immune responses in SiO2-exposed mice, we examined the population of CD4+CD25+FOXP3+ T cells in the peripheral blood using flow cytometry. The percentage of CD4+CD25+FOXP3+ T cells in SiO2-exposed mice was lower than that in matched controls. The percentage of CD4+CD25+FOXP3+ T cells in Lias−/− +SiO2 group was significantly lower than that in the Lias+/+ +SiO2 group (Fig. 5A–D).
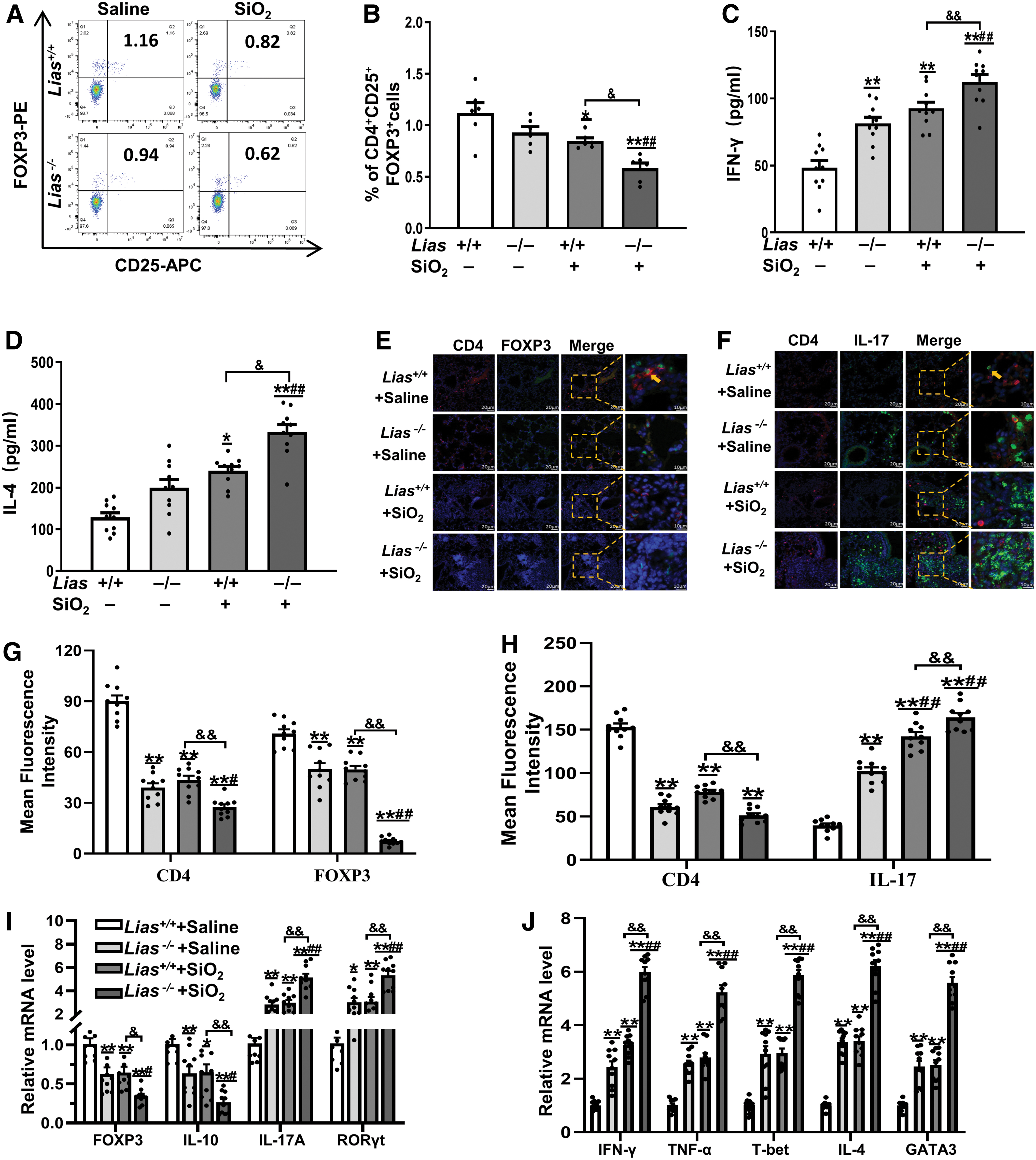
The IFN-γ, tumor necrosis factor-α (TNF-α), and T cell-specific T-box transcription factor (T-bet) linked with Th1 responses, and IL-4 and GATA binding protein-3 (GATA-3) linked with Th2 responses were examined in lung tissue by enzyme-linked immunosorbent assay (ELISA) and qPCR. The levels of IFN-γ and IL-4 in SiO2-exposed mice were higher than those in the matched controls.
The levels of IFN-γ and IL-4 in Lias−/− +SiO2 group were significantly lower than those in Lias+/+ +SiO2 group (Fig. 5C, D). Further analysis by qPCR showed that the levels of IFN-γ, TNF-α, T-bet, IL-4, and GATA-3 expression in SiO2-exposed groups mice were higher than those in matched controls and their expression in the Lias−/− +SiO2 group was significantly increased compared with Lias+/+ +SiO2 group (Fig. 5H).
CD4+FOXP3+ and CD4+IL-17+ cells in the lung tissue were measured by immunofluorescence. Compared with the matched control group, the number of CD4+FOXP3+ cells in the lung tissue of the SiO2-exposed group was significantly lower. However, the expression levels of CD4+IL-17+ cells in the lung tissue were significantly higher than those in the matched control group (Fig. 5E, F). Subsequently, we verified the expression of Tregs and Th17 cells in the lung tissue using qPCR. These results are consistent with those of the immunofluorescence analysis (Fig. 5G). These results show that low LIAS expression can activate Th17 cells by inhibiting Tregs in lung tissue.
Low expression of LIAS inhibits the differentiation of Tregs in Lias+/+ and Lias−/− mice spleen during SiO2-induced fibrogenesis
The population of CD4+CD25+FOXP3+ T cells in the spleen was measured using flow cytometry. The results showed that the population of CD4+CD25+FOXP3+ T cells in SiO2-exposed mice was lower than that in matched controls. The population of CD4+CD25+FOXP3+ T cells in the Lias−/− +SiO2 group was significantly lower than that in the Lias+/+ +SiO2 group (Fig. 6A, B). We also measured the expression levels of FOXP3+ T cells using immunohistochemistry. The results showed that the number of FOXP3+ T cells in the spleen decreased in SiO2-exposed mice. The number of FOXP3+ T cells in the Lias−/− +SiO2 group was lower than that in the Lias+/+ +SiO2 group (Fig. 6E, F).
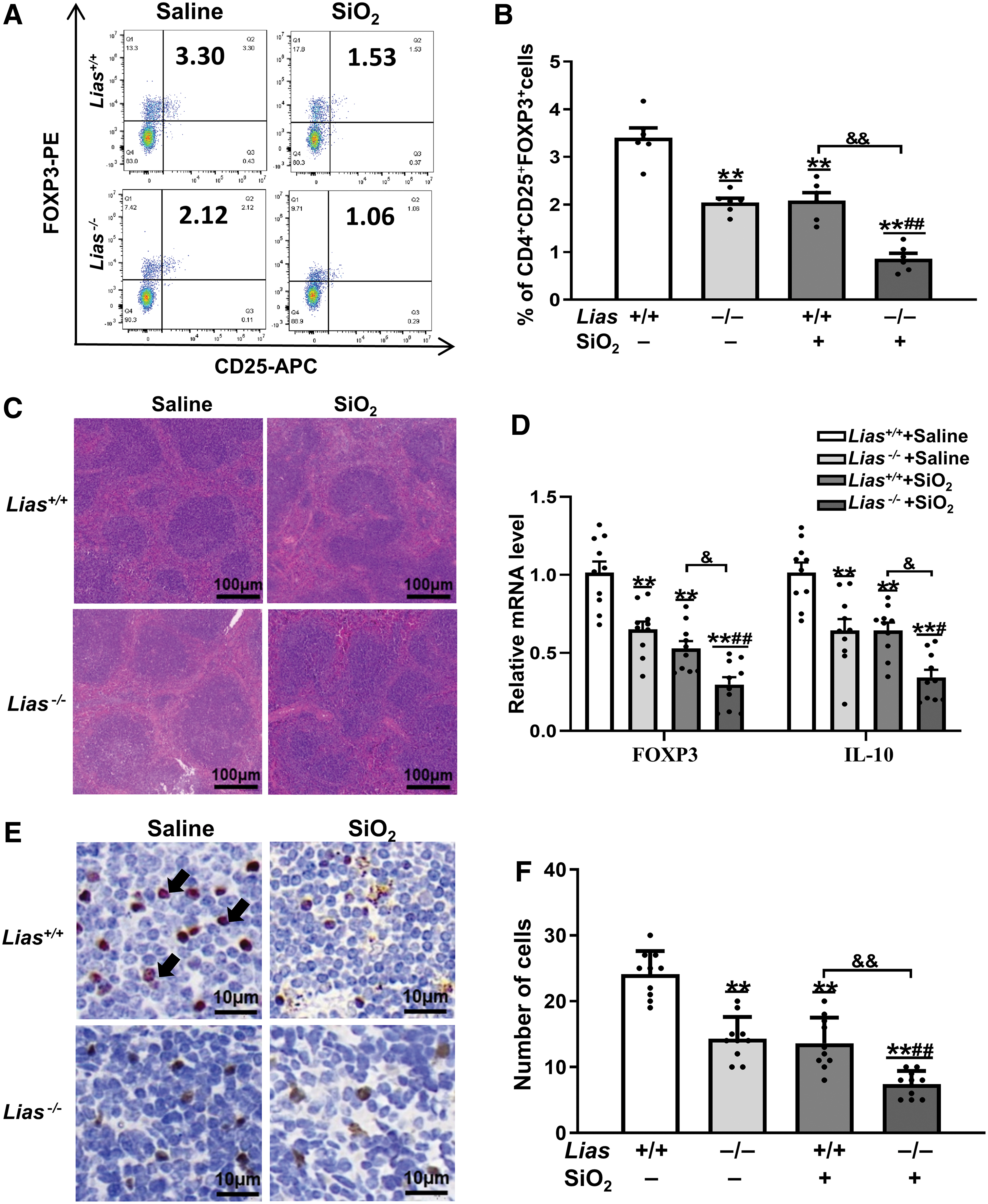
The results of histological analysis showed intact white pulp, and the boundary between the red pulp and white pulp was unclear in the SiO2-exposed mouse spleen. Compared with Lias+/+ +SiO2 group, the Lias−/− +SiO2 group exhibited more severe pathological lesions (Fig. 6C). The mRNA levels of the IL-10 and TGF-β and FOXP3 in the SiO2-exposed mice spleen were decreased compared with the matched controls. The mRNA levels in the Lias−/− +SiO2 group were lower than those in the Lias+/+ +SiO2 group (Fig. 6D).
Exogenous ALA attenuated SiO2-induced pulmonary fibrosis and decreased SiO2-induced collagen deposition in Lias+/+ and Lias−/− mice
The pulmonary tissue slices were stained with H&E. The results showed infiltration of inflammatory macrophages and cellular nodules in SiO2-exposed Lias+/+ and Lias−/− mice. Compared with SiO2-exposed mice, those with exogenous ALA supplementation had reduced modified Ashcroft scores (Fig. 7A, B). Accordingly, the results showed that the mRNA expression of type I and III collagen in SiO2-exposed mice was higher than that in mice that had received saline, and that exogenous ALA supplementation decreased the expression of these mRNAs compared with SiO2-exposed mice (Fig. 7C).
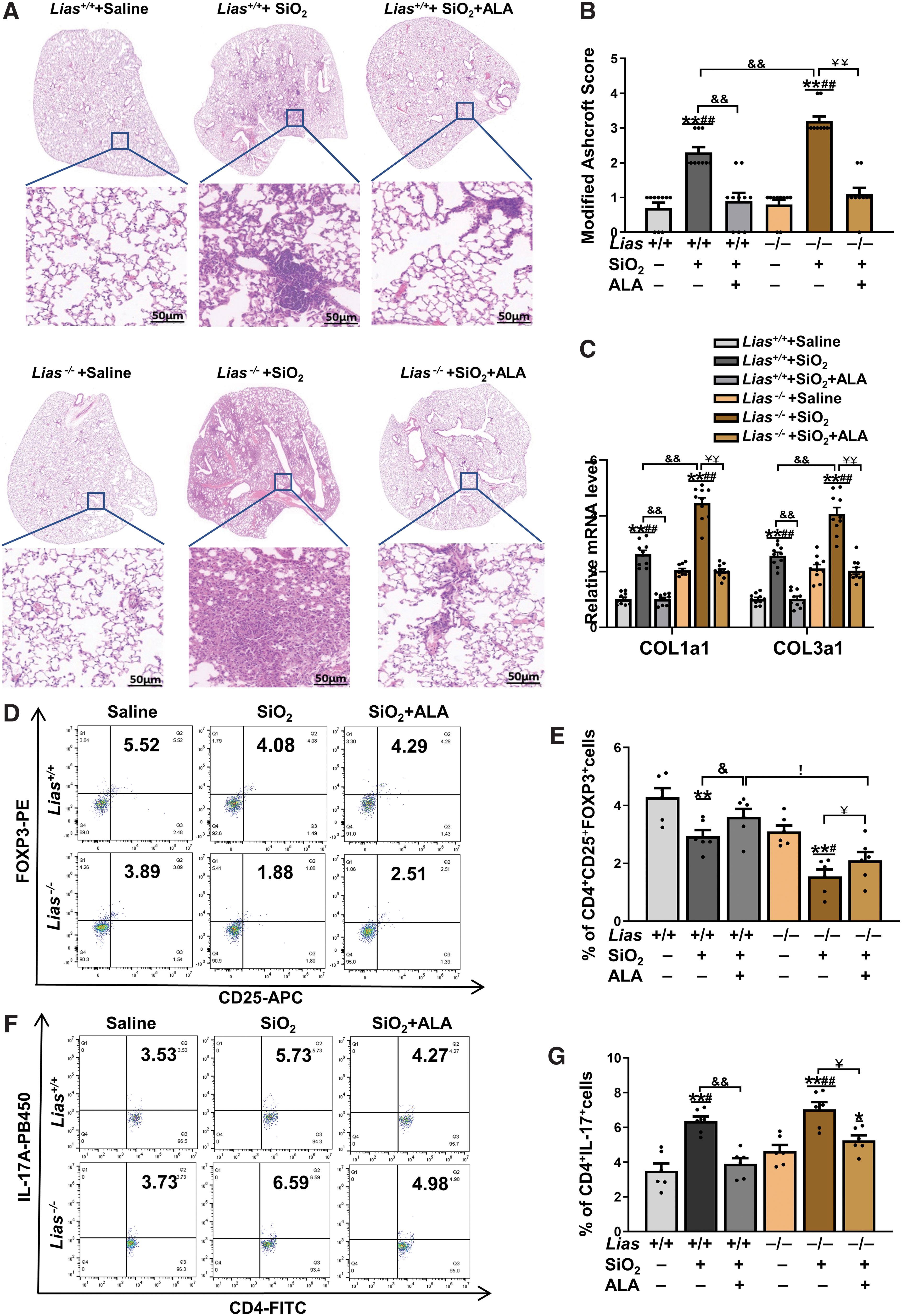
Populations of CD4+CD25+FOXP3+ T cells and CD4+IL-17A+ T cells in the spleen were measured using flow cytometry. The results showed that the population of CD4+CD25+FOXP3+ T cells, was increased and the population of CD4+IL-17A+T cells was decreased in the exogenous ALA-supplemented mice (Fig. 7D–G).
Exogenous ALA inhibited the oxidative stress in the lungs and maintained the balance of Th1/Th2 cells during SiO2-induced fibrogenesis
We detected the activities of SOD, CAT, and MDA in the lung tissue. The activities of SOD and CAT decreased, and MDA levels increased in the SiO2-exposed groups (Fig. 8A–C). Exogenous ALA supplementation increased the activities of SOD and CAT, and reduced MDA levels. We also measured ROS levels in lung tissue using fluorescent probes. The results showed that the levels of pulmonary ROS in SiO2-exposed mice were higher than those in mice that received saline, and exogenous ALA supplementation decreased ROS levels in SiO2-exposed mice (Fig. 8D). Exogenous ALA inhibits SiO2-induced ROS production in Lias+/+ and Lias−/− mice during SiO2-induced fibrogenesis.
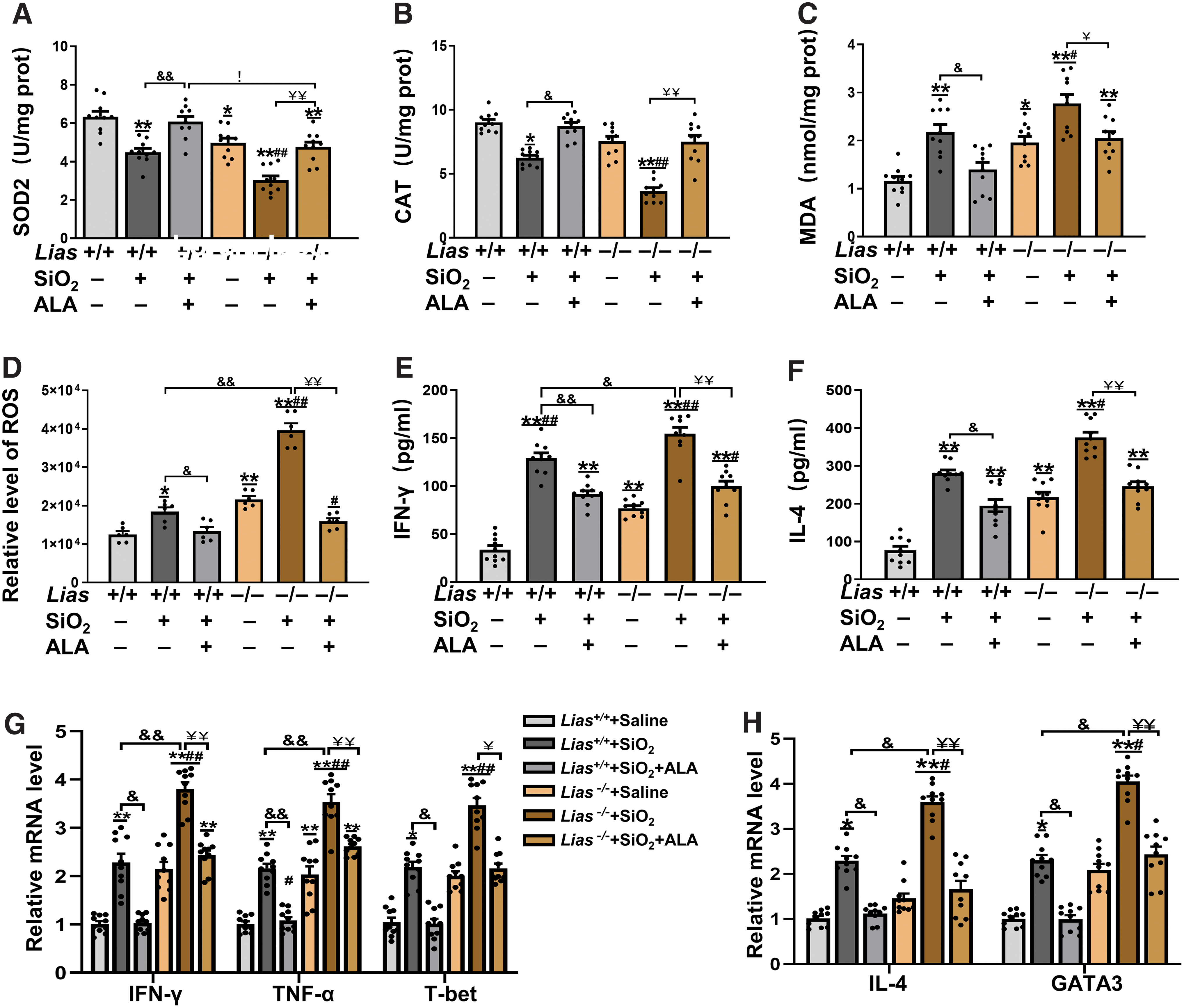
To explore the effects of exogenous ALA on the balance of Th1 and Th2 cells, we measured Th1-related cytokines including IFN-γ, TNF-α, and T-bet, and Th2-related cytokines including IL-4 and GATA3 in lung tissue with ELISA and qPCR (Fig. 8E–H). The results showed that exogenous ALA inhibited the secretion of inflammatory cytokines in Lias+/+ and Lias−/− mice induced by SiO2.
Discussion
Silicosis is a chronic and progressive disease characterized by pulmonary fibrosis, for which there is no effective treatment. A population of multiple cell types, including alveolar macrophages, pulmonary epithelial cells, and fibroblasts, is involved in the pathological process of silicosis-induced fibrosis (Pollard, 2016). Recent studies have shown that there is an immune function disturbance during silicosis fibrosis, which is related to an imbalance in immune cells and immune microenvironment homeostasis in the lung tissue (Pollard, 2016). Lipoic acid, a natural organic compound, is a strong antioxidant. Studies have shown that ALA has a regulatory effect on cellular immune responses (Lee et al., 2012; Tsou et al., 2014).
In this study, Lias−/− mice with low endogenous LIAS expression were established using homologous recombination technology, which can be stably inherited in generations. First, we confirmed the genotype and LIAS protein expression levels. The results showed that the degree of SiO2-induced pulmonary fibrosis was aggravated in Lias−/− mice. This is consistent with our previous findings that the degree of fibrosis was reduced in overexpression of LIAS in LiasH/H mice (Zhao et al., 2020).
EMT is a process in which epithelial cells gradually transform into mesenchymal cells and lose their original functions (Xi et al., 2014). EMT is a key factor in the pathogenesis of silicosis-related fibrosis. Therefore, we detected biomarkers of EMT in lung tissue by Western blotting. The results showed that low LIAS expression increased collagen fiber production and promotes epithelial–mesenchymal transformation induced by SiO2 in Lias−/− mice. This indicates that low LIAS expression affects the process of epithelial–mesenchymal transformation in the lung tissue of Lias−/− mice.
During SiO2-induced fibrogenesis, oxidative stress induced by excessive ROS drives the progression of fibrosis, which is accompanied by an imbalance between the oxidative and antioxidant defense systems (Sayan and Mossman, 2016). To elucidate the relationship between low LIAS expression and oxidative stress during SiO2-induced fibrogenesis, the antioxidant levels were measured in each group. The results showed that, compared with the matched control group mice, the activities of SOD and CAT were decreased, and MDA levels were increased in SiO2-exposed groups mice.
The antioxidant levels in the Lias−/− +SiO2 group were lower than those in the Lias+/+ +SiO2 group. These results indicate that low expression of LIAS leads to damage to antioxidant capacity. Furthermore, ROS levels in the lung tissue were detected. The results showed higher ROS levels in the SiO2-exposed group than in the control group, and higher ROS levels in the Lias−/− +SiO2 group than in the Lias+/+ +SiO2 group.
This indicated that low LIAS expression exacerbated ROS production during SiO2-induced fibrogenesis. ROS are mainly generated in mitochondria of alveolar epithelial cells in response to exogenous stimuli (Ning et al., 2019). Therefore, proteins related to mitochondrial function in the lung tissue were measured. Low LIAS expression leads to mitochondrial damage due to excessive ROS during SiO2-induced fibrogenesis, whereas mitochondrial damage conversely increases ROS production.
Excessive ROS levels can lead to immune suppression by affecting Treg function (Pollard, 2016; Stanek et al., 2011). Therefore, the Treg profile and Treg-associated cytokines in lung tissue were measured to explore the effects of low LIAS expression on immune responses during SiO2-induced fibrogenesis. The expression of Foxp3, a key transcription factor of Tregs, was detected in the spleen using immunohistochemistry. Flow cytometry and immunofluorescence showed that low expression of LIAS inhibited Treg differentiation.
Next, the expression of IFN-γ and IL-4 in the lung tissue was measured by ELISA and qPCR. These results suggest that low LIAS exacerbated the imbalance of Th1/Th2 during SiO2-induced fibrogenesis. One possible reason is that the activation and differentiation of immune cells depend on the microenvironment, including antigens, cytokines, and oxidation signals (Canli et al., 2016). Tregs inhibit Th1 and Th2 responses by affecting the relevant cytokines and transcription factors. These data suggest that low expression of the LIAS gene decreases Tregs, and greatly influences the function of Tregs in the inhibition of Th1 and Th2 responses during silica-induced inflammation and fibrosis (Zhu et al., 2006).
Previous studies have shown that the balance between Th17 and Treg cells plays a crucial role in silicosis fibrosis (Ivanov et al., 2006). The expression of IL-17A and retinoid-related orphan nuclear receptor γt (RORγt) in the lung tissue was measured by qPCR. IL-17A and RORγt, key transcription factors in IL-17 production, were significantly increased in the silica-treated groups compared with the matched control group (Ivanov et al., 2006). Similarly, the IL-17A and RORγt levels in Lias−/− +SiO2 group mice were higher than those for the Lias +/++SiO2 group. This suggests that low LIAS expression exacerbates the differentiation of Th17 cells during SiO2-induced fibrogenesis.
Tregs are crucial for the integration and maintenance of immune homeostasis, and mainly develop in the thymus (Richards et al., 2015). During the maturation process, Tregs migrate from the thymus to secondary lymphoid tissues, such as the lymph nodes and spleen, or peripheral nonlymphoid tissues (Richards et al., 2015). Therefore, the expression of Tregs in the spleen was examined to determine the effects of silica on Treg differentiation. Histological examination of the spleen showed that silica damaged the spleen structure of the mice, which was not intact in the white pulp, and the boundary between the red pulp and white pulp was unclear. These results were consistent with those of Zhao et al. (2022).
The responses of Th17 and Th1 cells may increase to promote lung inflammation due to the suppression of Tregs induced by SiO2. Excessive lung inflammatory responses increase Th2 cell function. During tissue repair, hyperplasia of the lung extracellular matrix and abnormal deposition were observed. Our results showed that the proportion and function of Tregs were inhibited, the levels of ROS in lung tissue were increased, and the EMT in lung tissue was enhanced in the Lias−/− +SiO2 group compared with the Lias +/++SiO2 mice.
It was first demonstrated that ALA has immunoregulatory effects on SiO2-induced pulmonary fibrosis in a Lias−/− mouse model. Furthermore, we explored whether exogenous ALA supplementation could attenuate silicosis-induced fibrosis by regulating the immune response. Fibrotic thickening, collagen deposition, and cellular nodules were observed in the lungs of the silica-exposed mice. Compared with silica-exposed mice, there were fewer areas of fibrotic bands in the lungs of the exogenous ALA supplement group. Our data showed that ALA treatment contributed to the inhibition of inflammation due to promotion of the Tregs response by ALA in the process of silicosis fibrosis.
We investigated the effects of ALA on the activity of antioxidant enzymes and silica-induced inflammatory responses. ALA is a strong antioxidant that acts as a free-radical scavenger. ALA can improve the antioxidant capacity (Shay et al., 2009). The results showed that TAC levels in the peripheral blood and SOD, CAT, and GPx levels in the lung tissue of mice in the exogenous ALA group were significantly higher than those in the matched silica-exposed mice. However, the MDA content in the lung tissues of mice in the exogenous ALA supplementation group was significantly lower than that in the matched exposed mice. Exogenous ALA supplementation improved antioxidant capacity and inhibited oxidative stress induced by silica. ALA may alleviate silicosis-induced pulmonary fibrosis by maintaining an imbalance between the antioxidant defenses and oxidative damage (Jakubczyk et al., 2020).
Inflammatory factors increase during the development of silicosis-associated pulmonary fibrosis, and lung inflammation drives the progression of fibrosis (Sayan and Mossman, 2016). Our results showed that the levels of inflammatory factors in the lung tissue of mice in the exogenous ALA supplementation group were significantly lower than those in the matched silica-exposed mice. Exogenous ALA supplementation inhibits silica-induced inflammatory responses.
Conclusion
Taken together, the evidence obtained from this study shows that LIAS affects the cellular immune response in SiO2-induced pulmonary fibrosis in mice. We found that SiO2-induced pulmonary fibrosis was aggravated in Lias−/− mice because low LIAS expression reduced the population of Tregs and enhanced the immune response. Exogenous ALA attenuated SiO2-induced pulmonary fibrosis by inhibiting oxidative stress in the lungs and maintaining the Th1/Th2 cell balance during SiO2-induced fibrogenesis. Our findings indicate that ALA is a potential prophylactic and/or therapeutic intervention for SiO2-induced pulmonary fibrosis.
Materials and Methods
Crystalline silica
Crystalline silica particles (>99% purity) were purchased from Silica Company (Frederick, MD), and >97% of the particles were 1–5 μm in diameter. After grinding for 2 h in a sterile agate mortar, the particles were dehydrated for 6 h at 180°C in a pyrostat to denature the endotoxins. Scanning electron microscopy (Hitachi, Tokyo, Japan) showed that the particles were still irregular, and the size of most particles was <2 μm (Fig. 2B). The weighed silica particles were suspended in sterile normal saline at a concentration of 50 mg/mL, and 2000 units of penicillin per milliliter were added before silica particle exposure.
Alpha-lipoic acid
ALA (Sigma-Aldrich, St. Louis, MO) was diluted to a concentration of 100 mg/mL in ultrapure water, and its solubility was increased by adjusting the pH from 4 to 5–6 with 1.0 M sodium hydroxide.
Animal experiments
Animal model
Lias-floxed mice with a loxP-modified 3′-untranslated regions of the LIAS gene were gifted by professor Oliver from the University of North Carolina. Lias−/− mice with low expression of the Lias gene were generated by crossing the Lias-floxed mice with E2a-Cre mice (Cat. 003724; Jackson Lab, Bar Harbor, ME) on a C57BL/6J background. The genotypes of the Lias−/− and Lias +/+ mice with wild-type Lias gene were identified using PCR.
The mice were housed under standard conditions at a temperature of 22°C ± 1°C and 12-h light/dark cycle and 40%–60% humidity, water ad libitum, and were fed a standard laboratory rodent diet (Puluteng, Inc., Shanghai, China). The experimental protocol was approved by the Animal Care and Use Committee of Xinxiang Medical University (Approval No. XYLL-2017086).
Experimental design
Lias−/− mice with low expression of the Lias gene were used to assess the effects of ALA on cellular immune responses in silicosis mice. Twenty-eight-week–old Lias+/+ and Lias−/− male mice were randomly divided into two groups (n = 10 per group): Lias+/+ +saline, Lias−/− +saline, Lias+/+ +SiO2 group, and Lias−/− +SiO2. To successfully establish the pulmonary fibrosis model, a 50 μL volume of silica suspension at a concentration of 50 mg/mL was administered by single intratracheal instillation to the Lias+/+ +SiO2 group and Lias−/− +SiO2 group mice according to the previous research (Zhao et al., 2020), while sterile saline with the same volume was administered to the Lias+/+ + saline group and Lias−/− +saline group mice.
After 28 days, peripheral blood was collected through the femoral artery, and the mice were anesthetized with isoflurane. The inferior lobes of the right lung and a part of spleen were fixed with 4% paraformaldehyde. Living pulmonary tissues were immediately harvested for the evaluation of ROS and ATP levels. Suspended cells were obtained from the living spleen and peripheral blood for flow cytometry. The rest of these tissues were stored at −80°C.
The effects of supplementary exogenous ALA on cellular immune responses were evaluated in mice of two genotypes during SiO2-induced fibrogenesis (Fig. 2A). Thirty-eight-week–old Lias+/+ and Lias−/− male mice were randomly divided into three groups (n = 10 per group): saline, SiO2, and SiO2+ALA. A 50 μL volume of SiO2 suspension at a concentration of 50 mg/mL was administered by single intratracheal instillation to the SiO2 and SiO2+ALA groups, while the same volume of saline was administered to the saline groups.
After 28 days, ALA was orally administered at a dose of 100 mg/kg, which falls within a pharmaceutical dose range (Mahmoudinezhad and Farhangi, 2021), to the SiO2+ALA groups once a day for 28 consecutive days. Equal volumes of sterile saline were administered to the saline and SiO2 groups by gavage. After 56 days, all animals were treated following for the process in which it was performed for 28 days. The mice were euthanized with CO2 according to a previous experimental protocol (Boivin et al., 2016).
Pathological examination
For the histological analysis, the inferior lobes of right lung and one-third of the spleen tissue were fixed in 4% paraformaldehyde for 24 h. The tissues were then embedded in paraffin and cut into 4 μm-thick slices. Lung slices were stained with H&E and Masson's trichrome using standard procedures. Spleen slices were stained with H&E. Histopathological features of the lungs and spleens were observed under an optical microscope (Model DFC360 FX; Leica, Frankfurt, Germany). The degree of fibrosis was scored according to the modified Ashcroft histopathology system, based on the blind method.
ROS levels measured by immunofluorescence
A single-cell suspension from the lung tissue was prepared by grinding with frosted glass sheets. After rinsing with phosphate-buffered saline (PBS) containing 0.1% fatal bovine serum, the washing solution was filtered through 200 nylon mesh to remove impurities. Then, the filtrates were centrifuged for 5 min at 4°C, 12,000 g. The sediment was resuspended in PBS containing 0.1% bovine serum albumin (BSA) and cultured in the RPMI1640 medium (Hyclone; Thermo Fisher Scientific, Waltham, MA) at 5% CO2, 37°C for 6 h.
ROS levels in the lung tissue cells were measured using an immunofluorescence assay. The medium was removed, and the cells were rinsed with PBS three times. The cells were diluted at a concentration of 2 × 105 cells per well in 96-well plates and incubated in the medium containing 2′,7′-dichlorofluorescein diacetate at a concentration of 10 μM at 37°C for 20 min. Fluorescence intensity was measured using a microplate reader at excitation and emission wavelengths of 488 and 525 nm, respectively.
ATP levels in lung tissue
Approximately 30 mg of lung tissue was added to 300 μL RIPA lysis buffer (Beyotime Biotechnology, Shanghai, China) containing 3.0 μL PMSF. The tissue was homogenized by grinding with a tissue crusher (KZ-5F-3D; Servicebio, Wuhan, Hubei, China) at 70 Hz for 1 min. After 30 min on ice, the tissue homogenization was centrifuged with 11,000 g at 4°C for 5 min, and the supernatant was taken for testing. Luminescence was measured using a chemiluminescence meter, and ATP levels were calculated according to a standard curve.
Flow cytometry analysis
Cell preparation
A 200 μL sample of peripheral blood was collected through the femoral artery for flow cytometry analysis. The spleen single-cell suspension was prepared in PBS containing 0.1% BSA followed by grinding with frosted glass sheets. The washing solution was then filtered through a 200-mesh sieve. Erythrocyte lysis buffer (3 mL, 00-4333-57; eBioscience, San Diego, CA) was added to the peripheral blood, filtered through the spleen homogenate, and incubated at room temperature in the dark for 5 min, then centrifuged at 4°C, 400 g for 5 min. The sediment was resuspended in PBS containing 0.1% BSA. Cell counts were performed on cell suspensions.
Flow cytometric cell staining
The cells were stimulated with 2 μL/mL cell stimulation cocktail (00-4975-03; eBioscience), including Phorbol 12-myristate 13-acetate, ionomycin, brefeldin A, and monensin for 4 h at 37°C, 5% CO2 in the cell incubator. Then, the cells were incubated with antimouse CD4-FITC (11-0041-82; eBioscience), CD3-PerCP eFlour710 (46-0032-80; eBioscience), and CD25-APC (17-0251-81; eBioscience) at room temperature in the dark for 30 min.
For intracellular FOXP3 and IL-17 staining, cells were fixed and permeabilized (00-5523; eBioscience) according to the manufacturer's instructions. Then, the cells were stained with antimouse FOXP3-PE (12-4771-80; eBioscience), IL-17-eFluor450 (48-7177-80; eBioscience), and incubated at 4°C in the dark for 30 min. All data collection and sorting were performed using the BD FACS Diva software (BD Biosciences, East Rutherford, NJ), and data analyses were performed using FlowJo software (Tree Star, Ashland, OR).
Immunohistochemistry
The lungs and spleen tissue slices were dewaxed in water, sealed with 3% hydrogen peroxide, and repaired by microwave with citric acid buffer. After blocking with 3% BSA for 30 min at room temperature, FOXP3 antibody was added to the spleen sections (diluted 1:200; Servicebio) at 4°C overnight. The membranes were rinsed and incubated with secondary antibodies conjugated to horseradish peroxidase (diluted 1:200; Santa Cruz Biotechnology, Santa Cruz, CA) for 30 min at room temperature. Finally, 3,3′-diaminobenzidine (Santa Cruz Biotechnology) was used as a chromogen solution for 1 min. Representative images were analyzed using Image-Pro Plus 6.0 software to measure the optical density.
Immunofluorescence assay
Paraffin-embedded lung tissue sections were dewaxed in xylene for 10 min and rehydrated in ethanol. Antigen retrieval was performed by heating tissue sections to 100°C for 15 min in 10 mM citrate buffer (pH = 6.0). Sections were then blocked with 3% BSA (Thermo Fisher Scientific) for 30 min at room temperature and incubated with primary antibodies (CD4-FITC, 1:200; FOXP3-PE, 1:200; IL-17-eFluor450, 1:200) in the dark at 4°C overnight. Cy3-conjugated secondary antibodies (diluted 1:200) were then added and incubated at room temperature for 50 min. Nuclei were stained with 4′,6-diamidine-2-phenylindole dihydrochloride. Images were captured using a laser confocal microscope (Leica).
Antioxidative capacity assay
The TAC of the murine serum was measured using a TAC kit (Cell Biolabs, San Diego, CA). Uric acid equivalents were used to determine TAC. The activities of SOD (Solarbio, Beijing, China), CAT (Solarbio), GSH (Solarbio), and MDA (Solarbio) in the lung tissues were measured using colorimetric methods. The assay was performed according to the manufacturer's instructions.
Enzyme-linked immunosorbent assay
The levels of IFN-γ and IL-4 in murine lung tissue were determined using mouse IFN-γ ELISA kits (CSB-E04608m; Cusabio, Beijing, China) and mouse IL-4 ELISA kits (CSB-E04634m; Cusabio). All experiments were performed according to the manufacturer's instructions.
Western blotting
Approximately 20 mg of each lung tissue sample was cut into small pieces, and 200 μL RIPA lysate buffer (Beyotime Biotechnology) containing protease inhibitor was added. Lung tissue was homogenized using an ultrasonic tissue disruptor (Scientz, Ningbo, Zhejiang, China) at 70 Hz for 3–5 min. The homogenate was centrifuged at 4°C and 11,304 rpm for 10–15 min, and total protein in the supernatant was quantified with a commercial kit (BCA Protein Assay Kit; Thermo Fisher Scientific).
Samples containing equal amounts of protein were resolved by electrophoresis on a 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) followed by transfer onto a 0.45 μm polyvinylidene fluoride membrane (Millipore, Bedford, NY). The membrane was blocked with 5% fat-free milk for 2 h and incubated at room temperature for 2 h with the following primary antibodies: anti-LIAS (diluted 1:2000; Proteintech), anti-COXIV (diluted 1:2000; Cell Signaling, Boston, MA), anti-PAI-1 (diluted 1:2000; Cell Signaling), anti-α-SMA (diluted 1:1000; Affinity Biosciences, Cincinnati, OH), anti-Vimentin (diluted 1:1000; Affinity Biosciences), anti-E-cadherin (diluted 1:1000; Affinity Biosciences), and anti-β-actin (diluted 1:2000; Affinity Biosciences).
The membrane was washed three times with Tris-buffered saline (20 mM Tris-Base, 137 mM NaCl, pH 7.6; 5 min, room temperature), and then incubated with a horseradish peroxidase-conjugated secondary antibody (diluted 1:4000; Affinity Biosciences) at room temperature for 1 h. Protein expression was observed using the FluorChem R multifunction imaging analysis system (Protein Simple, San Francisco, CA). The intensity of bands in the Western blot was quantified using ImageJ 6.0 and normalized by the intensity of the β-actin as an internal control.
RNA extraction and real-time reverse transcription-polymerase chain reaction
Total mRNA was isolated from the lung and spleen tissues using TRIzol (Thermo Fisher Scientific), and cDNA was reverse transcribed using the Revert Aid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific). The expression levels of COL1a1, COL3a1, TFAM, COXIV, ATPase, PPARα, SOD, CAT, GPx, FOXP3, IL-10, TGF-β, RORγt, IL-17A, IFN-γ, TNF-α, T-bet, IL-4, and GATA3 were determined using a SYBR qPCR Mix Kit (Novoprotein, Beijing, China). The sequences of the primer pairs used were as follows: Ct values were determined using Design and Analysis 2.5.0 (Applied Biosystems, Waltham, MA) and normalized to actin Ct values using 2−ΔCt.
Statistical analysis
All statistical analyses were performed using the SPSS v20.0 Software (SPSS, Inc., Chicago, IL). The normality of the data distribution was evaluated using the Shapiro–Wilk test. Data are presented as the mean ± standard error of the mean. Multiple group comparisons were performed using one-way ANOVA followed by Tukey's post hoc test. Differences were considered significant at p values <0.05.
Ethics Declaration
The experimental protocol was approved by the Animal Care and Use Committee of Xinxiang Medical University (Approval No. XYLL-2017086), and the experiments were conducted in accordance with the legal requirements of the Basel Declaration.
Data Availability
The datasets generated and/or analyzed during this study are available from the corresponding author on request.
Footnotes
Authors' Contributions
Y.Z. and Se.Y. contributed to conceptualization, writing—original draft preparation, and writing—original draft preparation; Se.Y., M.L., and J.Y. provided methodology; G.X., Sa.Y., and Y.B. provided validation; Se.Y., Y.G., J.Y., and X.Y. performed data curation; G.X. and Sa.Y. provided supervision; Y.Z. assisted with article submission. All authors have read and agreed to the published version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work has been supported by the Talent Support Project of Xinxiang Medical University number XYBSKYZZ202207 (Yingzheng Zhao), the Key Scientific Research Projects of Henan Higher Education Institutions number 23A330001 (Yingzheng Zhao), and the National Natural Science Foundation of China number 81773399 (Yingzheng Zhao).