
Abstract
Significance:
Musculoskeletal diseases seriously affect global health, but their importance is greatly underestimated. These diseases often afflict the elderly, leading to disability, paralysis, and other complications. Hydrogen sulfide (H2S) plays an important role in the occurrence and development of musculoskeletal diseases, which may have potential therapeutic significance for these diseases.
Recent Advances:
Recently, it has been found that many musculoskeletal diseases, such as osteoporosis, periodontitis, muscle atrophy, muscle ischemia–reperfusion injury, muscle contraction under high fever, arthritis, and disc herniation, can be alleviated by treatment with H2S. H2S may be conducive to the development of multiple myeloma. The mechanism of action of H2S in the musculoskeletal system has been partly elucidated. A variety of H2S donors and nano-delivery systems provide promising prospects for H2S-based therapies.
Critical Issues:
Related research remains at the level of cell or animal experiments, but clinical research is lacking. The roles of H2S in more musculoskeletal disorders remain largely unknown. The serious consequences of musculoskeletal diseases have not been widely concerned. Targeted delivery of H2S remains a challenging task in musculoskeletal diseases.
Future Directions:
Develop therapeutic drugs for musculoskeletal diseases based on H2S and test their safety, efficacy, and tolerance. Explore the combination of current drugs for musculoskeletal diseases with H2S-releasing components to improve the therapeutic efficacy and avoid side effects. Carry out relevant clinical trials to verify the possibility of its widespread use. Antioxid. Redox Signal. 42, 321–340.
Introduction
Musculoskeletal diseases are an umbrella term for a range of diseases that occur in bone, skeletal muscle, joints, and discs, including osteoporosis, periodontitis, osteosarcoma (OS), and multiple myeloma (MM) that occurs in bones; osteoarthritis (OA) occurring in joints; muscle ischemia, ischemia–reperfusion injury (IRI), and atrophy occurring in skeletal muscles; as well as disc herniation occurring in intervertebral discs. As the global population ages, musculoskeletal diseases are becoming more and more common, yet their importance has been underestimated. Estimates of health care expenditures in the United States from 1996 to 2016 indicate that the cost spent on medical care for lower back and neck pain and other musculoskeletal disorders accounted for the highest proportion compared with any other disease (Dieleman et al., 2020). In 2017, there were ∼1.3 billion cases of musculoskeletal disorders globally, resulting in 121,300 deaths (Safiri et al., 2021). Musculoskeletal diseases often afflict the elderly, leading to disabilities, paralysis, and other complications. Therefore, it is urgent to develop effective therapeutic methods to alleviate or eliminate musculoskeletal diseases.
Hydrogen sulfide (H2S), a colorless toxic gas with a characteristic smell of rotten eggs, is the third endogenous gas signaling molecule discovered after nitric oxide (NO) and carbon monoxide (Calabrese et al., 2007; Wang, 2012). H2S is widely distributed in various tissues, including the brain, heart, kidney, gastrointestinal tract, skeletal muscle, and bone (Li et al., 2011; Wang, 2003). Since the discovery of its physiological role in the brain (Abe and Kimura, 1996), H2S has become the cutting edge of research in recent decades, and H2S donors and inhibitors have become useful tools for investigating the effects of H2S (Powell et al., 2018; Szabó, 2007). Sodium sulfide (Na2S), sodium hydrosulfide (NaHS), and morpholin-4-ium 4-methoxyphenyl(morpholino) phosphinodithioate (GYY4137) (Li et al., 2009; Zhao et al., 2001) are most frequently used H2S donors for in vivo and in vitro experiments, but they all have strong side effects. Research on better H2S donors is entering an exponential exploration era, and many fruitful achievements have emerged. For example, ATB-346, a novel H2S donor, is being developed for treating OA and has completed Phase I clinical trials in healthy volunteers. NO and H2S donors into aspirin, a novel NO and H2S-releasing hybrid, has been proven effective in the treatment and chemoprevention of cancer. SG1002 is the only H2S donor that converts 100% into H2S, making it the most efficient H2S-generating agent in clinical applications (Wallace and Wang, 2015). Inhibitors of H2S typically target the three H2S catalyzing enzymes, including cystathionine β-synthase (CBS), cystathionine γ-lyase (CSE), and 3-mercaptopyruvate sulfurtransferase (3-MST). Aminooxyacetic acid (d’Emmanuele di Villa Bianca et al., 2009), hydroxylamine (Velasco-Xolalpa et al., 2013), and O-(carboxymethyl)hydroxylamine hemihydrochloride (Jiang et al., 2015) are the most common inhibitors of CBS, while DL-propargylglycine and β-cyanoalanine (Madden et al., 2012) are widely used as CSE inhibitors. Pharmacological inhibitors of 3-MST include 2-[(4-hydroxy-6-methylpyrimidin-2-yl)sulfanyl]-1-(naphthalen-1-yl)ethan-1-one and 2-ketobutyric acid, while aspartic acid (Huang and Moore, 2015) is also discovered to inhibit the H2S production catalyzed by 3-MST indirectly by inhibiting cysteine aminotransferase (Jin et al., 2020; Xiao et al., 2015).
A growing number of studies have shown that H2S is a key mediator regulating various physiological functions in biological systems. Due to the high volatility and rapid metabolism of H2S, accurately controlling its release rate is extremely important. Using nanosystems for drug delivery to control H2S may contribute to the clinical treatment of various diseases (Li et al., 2020). However, despite ample evidence indicating that H2S can maintain normal metabolism and homeostasis of bones and skeletal muscles, related reviews are extremely few. Considering the crucial role of H2S in musculoskeletal diseases and its potential therapeutic implications for them, this review summarizes recent research progress on various musculoskeletal diseases. In addition, we delve deeper into relevant mechanisms and signaling pathways, discuss the potential of H2S in treating these diseases, and provide theoretical support for upcoming clinical trials.
Synthesis of H2S in the Musculoskeletal System
In mammals, H2S is mainly produced in the transsulfuration pathway of homocysteine (Hcy) metabolism by two pyridoxal-5′-phosphate (PLP)-dependent enzymes, CBS and CSE, and also by 3-MST, a non-PLP-dependent enzyme (Kamoun, 2004; Sbodio et al., 2019). CBS, CSE, and 3-MST have different tissue distributions and subcellular localization. CBS is mainly located in the central nervous system, kidney, ileum, pancreatic islets, and liver. CSE dominates the cardiovascular system (Kamoun, 2004; Li et al., 2023b; Polhemus and Lefer, 2014). 3-MST is expressed in the brain (Shibuya et al., 2009). CBS and CSE are located in the cytoplasmic matrix, while 3-MST is mainly located in mitochondria (Li et al., 2022; Shibuya et al., 2009).
The three H2S-producing enzymes are expressed differently in osteoprogenitor cells, osteoblasts (OB), osteoclasts (OC), and osteocytes (Fig. 1). Bone marrow mesenchymal stem cells (BMMSCs) express CBS, CSE, 3-MST, and cysteine aminotransferase. According to a bone biopsy, CSE is mainly expressed around the bone marrow microvasculature and less expressed in endosteal cells, while CBS is widely expressed in endosteal cells (Grassi et al., 2016). BMMSCs have the potential to differentiate into OB (Xu et al., 2023), which can be promoted by CBS and CSE (Zheng et al., 2017). The expression of CBS in BMMSCs is higher than that of CSE, and the lack of CBS will reduce the osteogenic capacity of BMMSCs (Liu et al., 2014b). In OB, CSE is the main source of H2S, with only 22% of H2S coming from CBS. 3-MST is not expressed in OB (Zheng et al., 2017). CSE is also highly expressed in OC and bone marrow stromal cells and weakly expressed in osteocytes (Zheng et al., 2017).
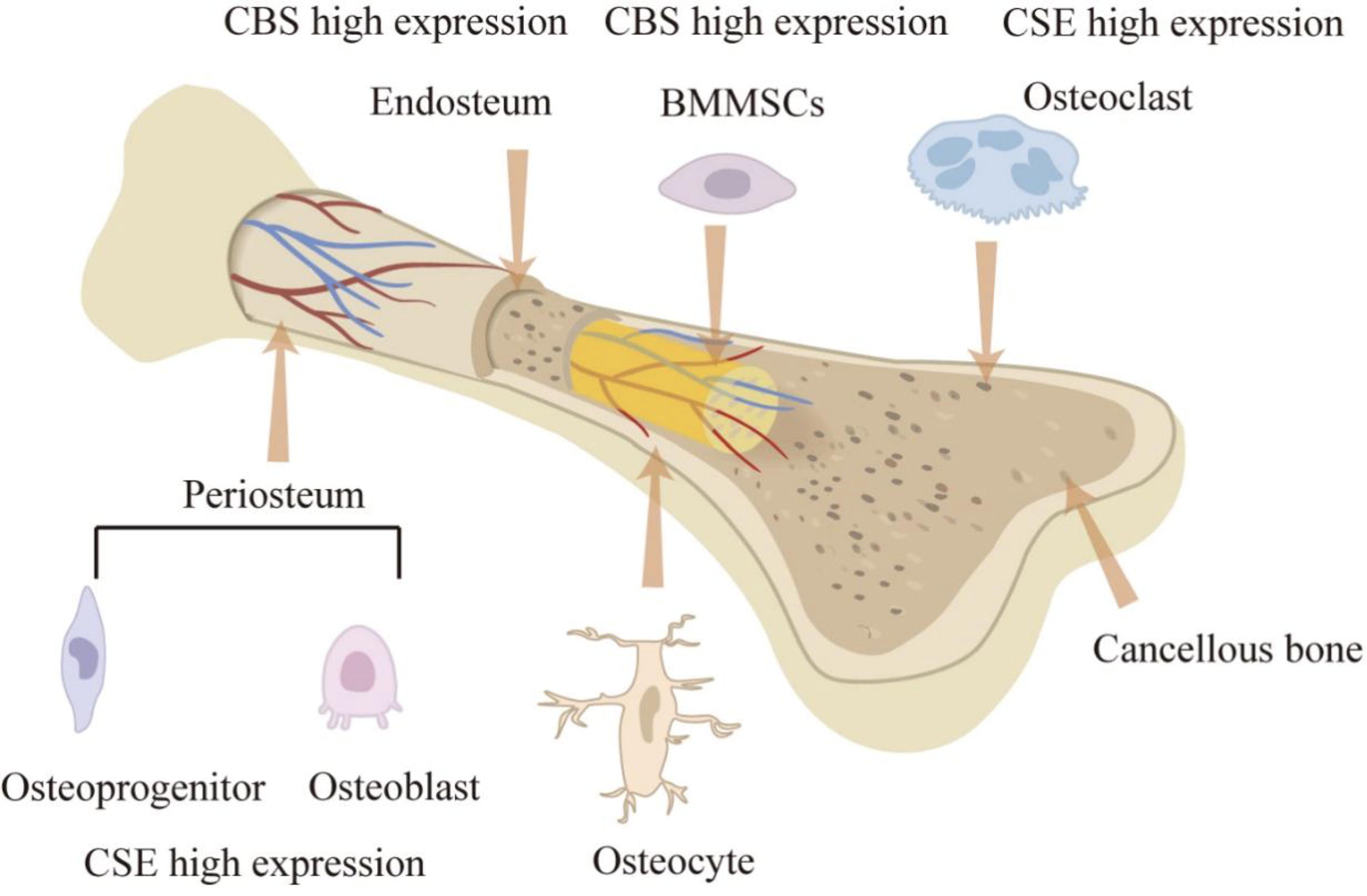
The diagram illustrates the expression levels of H2S-related synthases in normal bone tissue. BMMSCs, bone marrow mesenchymal stem cells; CBS, cystathionine β-synthase; CSE, cystathionine γ-lyase; H2S, hydrogen sulfide. Color images are available online.
In skeletal muscle, there are species differences in the expression of H2S-producing enzymes. CBS and CSE are both highly expressed in human skeletal muscle but rarely expressed in mice (Chen et al., 2010; Veeranki and Tyagi, 2015a; Veeranki and Tyagi, 2015b). All H2S-producing enzymes (CBS, CSE, and 3-MST) are present in rat skeletal muscle but produce less H2S than that in the liver and kidney (Du et al., 2013). In sheep and cattle, the expression of 3-MST in skeletal muscle is lower than that in other tissues (Aminlari et al., 1989). A protein localization analysis indicates that CBS and 3-MST are mainly expressed in and around the skeletal muscle endomysium, while CSE is distributed in the cytoplasm of myofibroblasts (Du et al., 2013). However, the species differences of H2S production in skeletal muscles are still unclear, and further studies are needed to elucidate the molecular mechanisms.
The Role of H2S in Bone-Related Diseases
Bone is an endocrine organ (Fukumoto and Martin, 2009; Guntur and Rosen, 2012). In the normal metabolism of bone, the increased bone mass due to the reconstruction of OB is equal to the decreased bone mass due to the absorption of OC. Therefore, the bone homeostasis can be achieved (Hadjidakis and Androulakis, 2006; Martin and Seeman, 2008; Rodan and Martin, 2000; Sommerfeldt and Rubin, 2001). On the contrary, the disorder of bone metabolism can cause bone diseases.
Osteoporosis
Osteoporosis, caused by the imbalance of bone formation and resorption, is a systemic metabolic bone disease characterized by osteopenia and the microstructural degradation of bone tissue, including primary and secondary osteoporosis. There are three types of primary osteoporosis: postmenopausal osteoporosis (PMO) (type I), senile osteoporosis (type II), and idiopathic osteoporosis. It has been reported that serum H2S levels, CBS, and CSE may be associated with the pathogenesis of osteoporosis (Hao et al., 2021).
H2S resists estrogen deficiency-induced osteoporosis
PMO, an aging-related bone disease caused by the cessation of ovarian function and the lack of estrogen, mainly occurs in postmenopausal women (Grassi et al., 2016; Ji and Yu, 2015; Riggs et al., 1998; Riggs and Melton, 1983; Zhai et al., 2017). The primary function of estrogen is to regulate the lifespan of mature OC through the Fas/Fas-ligand system (Nakamura et al., 2007). Estrogen deficiency can promote bone resorption by increasing mature OC, eventually leading to bone loss (Weitzmann and Pacifici, 2006).
H2S can regulate the Wnt/β-catenin pathway, which is involved in bone development and metabolism, as well as the differentiation and proliferation of chondrocytes, mesenchymal stem cells, OC, and OB (Vuong and Mlodzik, 2022). In the Wnt/β-catenin pathway, β-catenin is a key component. Most of the β-catenin in the cytoplasm binds to E-cadherin in the cell membrane and mediates intercellular adhesion. A small amount of β-catenin forms a destructive complex with Axin, adenomatous polyposis coli (APC), casein kinase I (CKI), and glycogen synthase kinase-3β (GSK-3β) and is degraded by the ubiquitin–proteasome pathway, maintaining the cytoplasmic β-catenin at a low level. The Wnt/β-catenin signaling pathway is activated in the presence of Wnt molecules. The Wnt ligand binds to the frizzled protein and interacts with the co-receptor lipoprotein receptor-related proteins 5 and 6 to activate the CKI in the cell and recruits the dishevelled (DVL) protein to the cell membrane for phosphorylation. On the one hand, phosphorylated DVL inhibits the degradation of β-catenin phosphorylation. On the other hand, it recruits Axin to destroy the Axin-APC-β-catenin complex so that β-catenin is released from the complex and transported to the nucleus to initiate the transcription and expression of downstream target genes of the Wnt pathway (Gao et al., 2002). Through paracrine and autocrine actions, Wnt signaling plays a crucial role in bone development (Kadowaki et al., 1996; Nusse and Clevers, 2017). Treatment with intraperitoneal injections of H2S donor GYY4137/vehicle every other day at 1 mg/mouse for 4 weeks can normalize serum H2S, increase bone formation, and completely prevent trabecular bone loss induced by ovx in ovariectomized mice. Mechanism studies have shown that GYY4137 activates Wnt signaling by increasing the production of Wnt ligands, thereby increasing the formation of OB in mice. H2S can also alleviate bone loss and osteoporosis caused by spinal cord injury by activating Wnt/β-catenin signaling (Grassi et al., 2016) (Fig. 2).
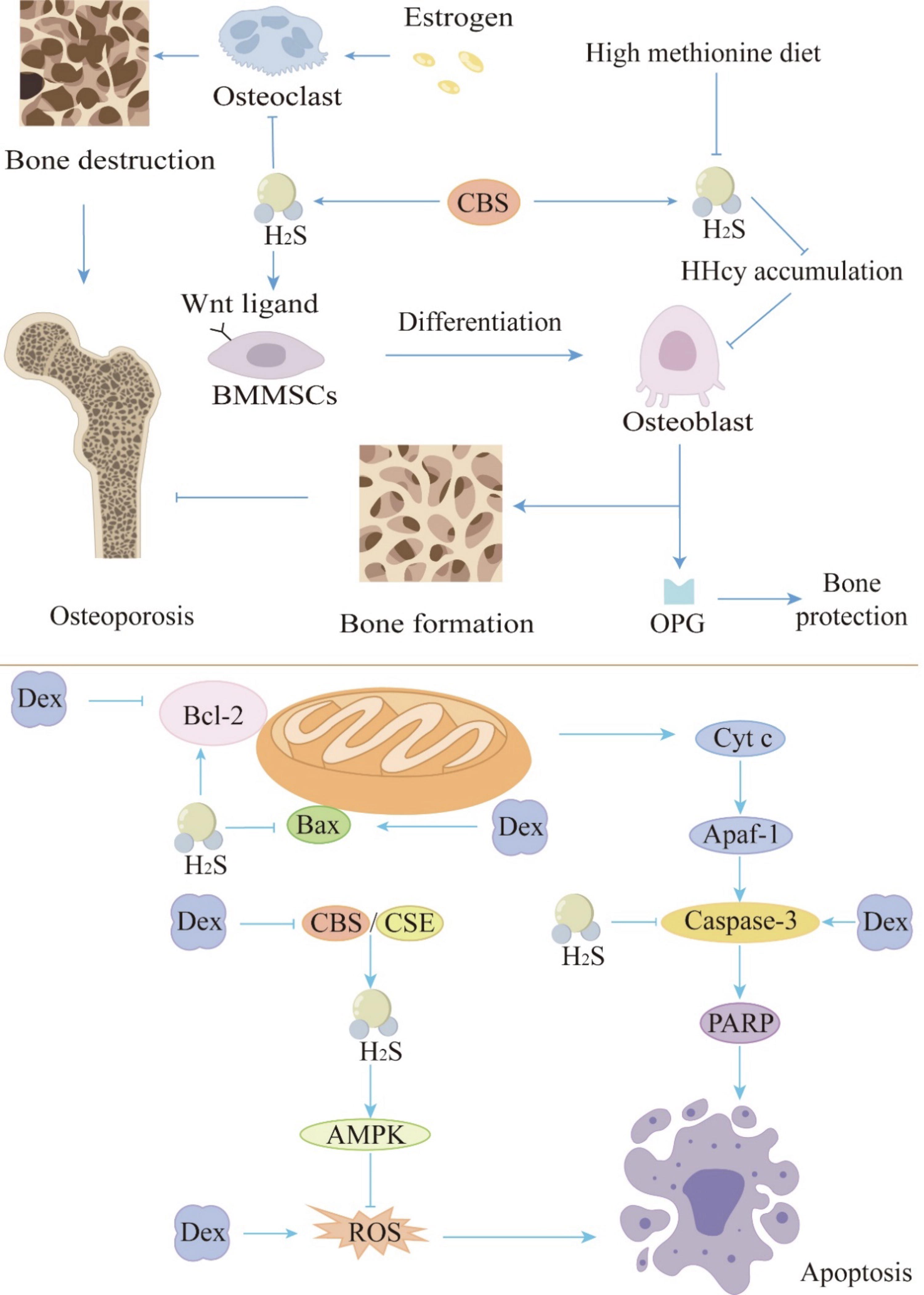
Potential mechanisms of the inhibitory role of H2S in osteoporosis. Osteoporosis is a systemic metabolic bone disease characterized by reduced bone mass and degradation of bone tissue microstructure, caused by an imbalance between bone formation and resorption. It can be triggered by various factors such as estrogen deficiency, HHcy, and long-term or excessive use of glucocorticoids. H2S can inhibit osteoporosis by acting on relevant targets either directly or indirectly. In estrogen-related osteoporosis, H2S can regulate the Wnt/β-catenin pathway by increasing the production of Wnt ligands, thus activating Wnt signaling. This activation promotes the formation of osteoblasts while inhibiting osteoclast formation, thereby alleviating osteoporosis. Supplementation with exogenous H2S might be an effective method for treating HHcy-induced osteoporosis. H2S helps resist HHcy-induced osteoporosis by lowering the expression of RANKL in osteoclasts, thereby inhibiting HHcy accumulation. Long-term and/or excessive use of glucocorticoids such as Dex can cause osteoblast damage and significantly reduce the levels of CBS and CSE in cells, leading to osteoporosis. H2S regulates nicotinamide adenine dinucleotide phosphate homeostasis by activating AMPK signaling and inhibiting ROS production. Additionally, H2S promotes the expression of Bcl-2 while suppressing Bax and caspase-3 expression, thus resisting glucocorticoid-induced osteoporosis. The tip arrows indicate the effect of promoting generation or activating expression; flat-end arrows indicate inhibition of generation or inhibition of expression. AMPK, adenosine monophosphate-activated protein kinase; Apaf-1, apoptotic protease activating factor 1; Bcl-2, B-cell chronic lymphocytic leukemia/lymphoma-2; Bax, Bcl-2-associated X protein; Dex, dexamethasone; Cyt C, cytochrome c; HHcy, hyperhomocysteinemia; OPG, osteoprotegerin; PARP, poly (adenosine diphosphate-ribose) polymerase; RANKL, receptor activator of nuclear factor-κB ligand; ROS, reactive oxygen species. Color images are available online.
H2S resists hyperhomocysteinemia-induced osteoporosis
Methionine metabolism disorder caused by a large amount of methionine intake in the Western diet may increase serum Hcy. Hyperhomosysteinemia (HHcy), characterized by elevated Hcy, is a significant independent risk factor for osteoporosis (van Meurs et al., 2004). HHcy can lead to osteoporosis by reducing the activity of OB (Vijayan et al., 2013). HHcy-induced phosphorylation of protein phosphatase 2A could result in downregulation of insulin-sensitive forkhead box transcription factor 1 (FOXO1) and p38 mitogen-activated protein kinase (MAPK) signaling mechanisms—thus, preventing the synthesis of osteoprotegerin (OPG) in OB (Vijayan et al., 2013). On the contrary, HHcy increased the synthesis of receptor activator of nuclear factor-κB (NF-κB) ligand (RANKL) in OB through the c-Jun/c-Jun N-terminal kinase (JNK) MAPK signal transduction mechanism. As a result, bone loss was aggravated due to the decreased OB activity and increased OC activity. Early studies have found that CBS deficiency is the most common cause of HHcy (Beard and Bearden, 2011; Nakladal et al., 2022; Uren et al., 1978; Watanabe et al., 1995), suggesting that CBS deficiency may lead to osteoporosis. Mice fed a high methionine diet show downregulation of CBS and CSE expression, leading to a decrease in endogenous H2S levels, eventually resulting in HHcy accumulation and osteoporosis (Behera et al., 2018a). BMMSCs have the biological characteristics of differentiating into OB, which can promote bone growth and repair. The decrease of H2S caused by CBS deficiency can downregulate protein kinase C/extracellular signal-regulated kinase (ERK)-mediated Wnt/β-catenin signaling by inhibiting the sulfidation of cysteine residues on multiple calcium (Ca2+) transient receptor potential channels, thereby inhibiting the self-renewal and osteogenic differentiation of BMMSCs (Liu et al., 2014b; Mustafa et al., 2009). In addition, HHcy induced by CBS deficiency can cause oxidative damage and dysfunction of BMMSCs (Behera et al., 2018b).
Supplementation with exogenous H2S may be an effective way to treat HHcy-induced osteoporosis. Studies have shown that after daily intraperitoneal injection of 10 mg/kg of NaHS for 6 weeks, the level of OPG in mice is restored and the expression of RANKL in OC is reduced by inhibiting the c-Jun/JNK-p pathway, thus achieving bone protection and bone resorption prevention (Behera et al., 2018a). Additionally, after treating mouse embryo osteoblast precursor cells MC3T3-E1 with NaHS at a concentration of 30 μM as a H2S donor for 24 h in vitro, H2S can alleviate HHcy-induced OB dysfunction by restoring mitochondrial function, reducing oxidative stress and OB apoptosis (Zhai et al., 2019). Mitochondria are the primary source of adenosine triphosphate (ATP). During initial bone formation and subsequent bone remodeling, OB can exhibit robust mitochondrial biogenesis, increased ATP production, and reduced mitochondrial stress. Oxidative stress in mitochondria can impair mitochondria function by opening the mitochondrial permeability transition pore (Calabrese et al., 2010; Shum et al., 2016). Research by Gao et al. indicates that inhibition of mitochondrial activity or increased production of mitochondrial superoxide significantly inhibits OB differentiation (Gao et al., 2018). Additionally, apoptosis of OB caused by mitochondrial dysfunction is also reversed. This indicates that H2S may relieve osteoporosis by restoring mitochondrial function (Fig. 2).
In summary, H2S can resist HHcy-induced osteoporosis by promoting osteogenesis and attenuating OC activation.
H2S resists glucocorticoid-induced osteoporosis
Glucocorticoids (GCs) are reported to increase the incidence of secondary osteoporosis. As a novel regulator, H2S is associated with GC-inhibited bone formation (Ma et al., 2019b). Long-term and/or excessive use of GCs such as dexamethasone (Dex), an anti-inflammatory and immunosuppressive drug, will induce OB damage. The levels of CBS and CSE in MC3T3-E1 OB cells treated with Dex are significantly reduced. Treatment of MC3T3-E1 cells with H2S donor NaHS (250 μM) in vitro can inhibit Dex-induced OB injury, which is achieved by regulating nicotinamide adenine dinucleotide phosphate homeostasis via activating adenosine monophosphate-activated protein kinase signaling and inhibiting reactive oxygen species (ROS) production, thereby protecting cells from oxidative stress (Yang et al., 2014). NaHS can also alleviate Dex-induced MC3T3-E1 OB cell dysfunction by increasing silent information regulator 1 and peroxisome proliferator-activated receptor gamma co-activator-1 α protein expression to improve mitochondrial function in vitro (Ma et al., 2019a). Another H2S donor, GYY4137/vehicle at 1 mg/rat, can accelerate the formation of OB by activating Wnt signaling (Ma et al., 2019b). Therefore, H2S can resist GC-induced osteoporosis by promoting osteogenesis (Fig. 2).
H2S reduces osteoporosis by resisting oxidative stress
Oxidative stress is not negligible in the development of osteoporosis (Basu et al., 2001; Isomura et al., 2004; Manolagas, 2010). Oxidative stress can induce OB apoptosis and inhibit its differentiation (Calabrese et al., 1996; Li et al., 2023a; Nandy et al., 2024), which is mediated by MAPK pathway (Park et al., 2005). Common MAPK enzymes include ERK1 and 2, p38-MAPK, JNK, and big MAPK 1 (also known as ERK5), which represent different MAPK pathways. Activation of p38 and JNK pathways leads to inflammation and apoptosis. ERK/MAPK signaling pathway is closely related to cell proliferation and differentiation (Guo et al., 2020). Endogenous H2S and exogenous H2S provided by 100 μM NaHS can protect OB cells from hydrogen peroxide (H2O2)-induced oxidative damage by inhibiting MAPK (p38 and ERK1/2), thereby inhibiting subsequent apoptosis (Xu et al., 2011). SDSS, a novel H2S-releasing compound derived from Danshensu, can significantly reduce the phosphorylation of MAPK pathway proteins at 25 μM in MC3T3-E1 OB cells, including p38, ERK1/2, and JNK (Yan et al., 2017), thereby reducing apoptosis.
Nuclear factor erythroid 2-related factor 2 (Nrf2) is a transcription factor that regulates the redox state of cells (Che et al., 2023). Increasing evidences have highlighted the key role of redox imbalance in the development of bone-related diseases. Nrf2 binds to the negative regulator Kelch-like epichlorohydrin-associated protein 1 (Keap1) dimer and exists in the cytoplasm in an inactive state. When the cells are stimulated by ROS or other nucleophiles, H2S interacts with Keap1 through S-sulfhydration to uncouple Keap1/Nrf2, and the activated Nrf2 is transported to the nucleus and binds to the antioxidant response element or electrophile response element, resulting in the expression of downstream genes regulated by Nrf2 and exerting antioxidant damage. H2S donor SDSS can inhibit oxidative stress in MC3T3-E1 OB cells by activating the Nrf2 signaling pathway, thereby protecting cells from H2O2-induced cell death (Yan et al., 2017).
In addition, H2S can also alleviate spinal cord injury-induced bone loss and osteoporosis by inhibiting oxidative stress and the activity of matrix metalloproteinases (MMPs) and activating Wnt/β-catenin signaling (Yang et al., 2017). On the whole, H2S can prevent osteoporosis by resisting oxidative stress.
Periodontitis
Periodontitis is a chronic inflammatory disease that occurs in periodontal supporting tissues. The imbalance of the oral microbial community is the initial factor of periodontitis and leads to excessive infiltration of immune cells in periodontal tissue. Macrophages constitute the first line of defense against periodontal pathogens. On one hand, they perform a range of defensive functions, including direct recognition and phagocytosis of pathogens, releasing chemotactic factors to recruit immune cells, production of specific pro-inflammatory resolution mediators, and differentiation into the M2 phenotype to exert anti-inflammatory effects (Wang et al., 2021b). On the other hand, various pro-inflammatory factors secreted by macrophages lead to the destruction of periodontal tissues. Neutrophils are the first immune cells recruited to the site of infection, possessing acute anti-infective and pro-inflammatory effects (Greenlee-Wacker, 2016). Excessive activation of neutrophils causes the destruction of periodontal tissues through multiple pathways. For example, neutrophils secrete proteases that degrade the extracellular matrix (ECM) of periodontal cells and activate OC via RANKL, leading to alveolar bone resorption (Greenlee-Wacker, 2016). Apoptotic neutrophils release toxic contents that further damage periodontal tissues. In summary, excessive activation and inadequate immune response are important pathogenic mechanisms of periodontitis.
Most researchers previously believe that H2S could promote the onset of periodontitis and cause significant damage to periodontal tissues. Incubation with 100 ng/mL of H2S leads to necrosis and apoptosis of gingival fibroblasts, accompanied by strong inhibition of superoxide dismutase (SOD) activity and increased production of ROS (Yaegaki et al., 2008). Incubation with 100 ng/mL H2S can inhibit the proliferation of human OB via the MAPK pathway (Imai et al., 2009). However, recent evidence suggests that H2S may be beneficial for cellular protection. Human periodontal ligament stem cells (PDLSCs) co-express CBS and CSE enzymes and further produce H2S. Inhibition of endogenous H2S production with a CBS inhibitor, hydroxylamine, significantly attenuates the proliferation of PDLSCs and reduces their osteogenic and adipogenic differentiation capabilities (Su et al., 2015). These controversial results may be due to different concentrations of H2S. Physiological concentrations of H2S promote the proliferation of PDLSCs, while high concentrations exhibit cytotoxic effects. Interestingly, Herrera et al. (Herrera et al., 2015) show that oral administration of 16 mg/kg ATB-346 (an H2S-releasing naproxen derivative) daily inhibits alveolar bone loss and inflammation in ligature-induced periodontitis rats. Similarly, oral administration of the H2S donor Na2S, with a single oral dose of ∼10–12 mg/kg/day, alone suppresses strong infiltration of inflammatory cells and loss of connective tissue in gingival papillae secondary to periodontitis, suggesting the therapeutic potential of H2S in periodontitis. This contrasts with earlier research by Yaegaki K et al., possibly due to different experimental models. Yaegaki K et al.’s conclusions are based on cellular-level studies, potentially overlooking the holistic effects of H2S. In contrast, Bruno Schneider Herrera et al.’s findings may be more compelling. In summary, the role of H2S in periodontitis is concentration dependent and manifests different outcomes across different models.
Bone-related cancer
MM is a malignant tumor of plasma cells characterized by the proliferation of cancer cells in the bone marrow, leading to bone damage. In the bone marrow microenvironment of MM, malignant plasma cells, OC, and bone marrow stromal cells interact to produce a large number of cytokines. These cytokines disrupt bone remodeling balance by promoting OC differentiation and inhibiting new bone formation, leading to lytic bone lesions. As the disease progresses, 80% to 90% of patients experience lytic lesions and severe bone pain at initial diagnosis. More than 50% of patients experience compressive fractures, significantly reducing the quality of life of patients with MM (Kyle et al., 2003). The concentration of H2S in the serum of patients with MM is significantly higher than that in the healthy control group. A recent study has shown that at 200 μM NaHS accelerates the cell cycle of MM cells by reducing the distribution in the G0/G1 phase and increasing it in the G2/M phase, indicating a promotional effect of H2S on MM proliferation (Zhang et al., 2020a). Analysis of bone marrow samples from newly diagnosed patients with MM without any treatment (Zhang et al., 2020a) reveals that the messenger ribonucleic acid (mRNA) expression of CBS, rather than CSE, is elevated in MM cells. Inhibition of CBS, but not CSE, significantly suppresses H2S production in MM cells, indicating that the increased H2S in MM cells is synthesized via CBS. In addition, CBS inhibitors can suppress the proliferative ability of MM cells in a concentration-dependent or time-dependent manner, suggesting that reducing H2S levels or inhibiting CBS rather than CSE activity may be effective strategies for treating MM.
The phosphatidylinositol 3-kinase (PI3K)/protein kinase B (Akt) pathway plays a crucial role in the growth, survival, and drug resistance of MM cells (Younes et al., 2007). In the PI3K/Akt pathway, extracellular signals bind to activated Ras protein receptors, activating PI3K (phosphoinositide 3-kinase), which phosphorylates phosphatidylinositol-4,5-bisphosphate to convert it into phosphatidylinositol 3,4,5-trisphosphate (PIP3). PIP3 binds to Akt, recruiting Akt to the cell membrane, thereby activating Akt (p-Akt). Akt phosphorylates serine 9 of GSK-3β, reduces its phosphorylation level, induces apoptosis, and thereby regulates cell proliferation and differentiation (Davies, 2012). Moreover, Akt regulates cell growth through the mammalian target of rapamycin (mTOR) complex signaling. Akt also modulates NF-κB signaling by phosphorylating inhibitory kappa B kinase α and tumor progression locus 2. Studies have shown that NaHS significantly increases Akt phosphorylation (Zheng et al., 2016). Importantly, LY294002, a PI3K/Akt inhibitor, blocks NaHS-induced PI3K/Akt activation and promotes MM proliferation. Overall, MM cells upregulate CBS, leading to excessive H2S production, which enhances MM proliferation through the PI3K/Akt pathway. This provides three potential therapeutic strategies for MM treatment: H2S clearance, CBS inhibition, and PI3K/Akt inhibition (Fig. 3).
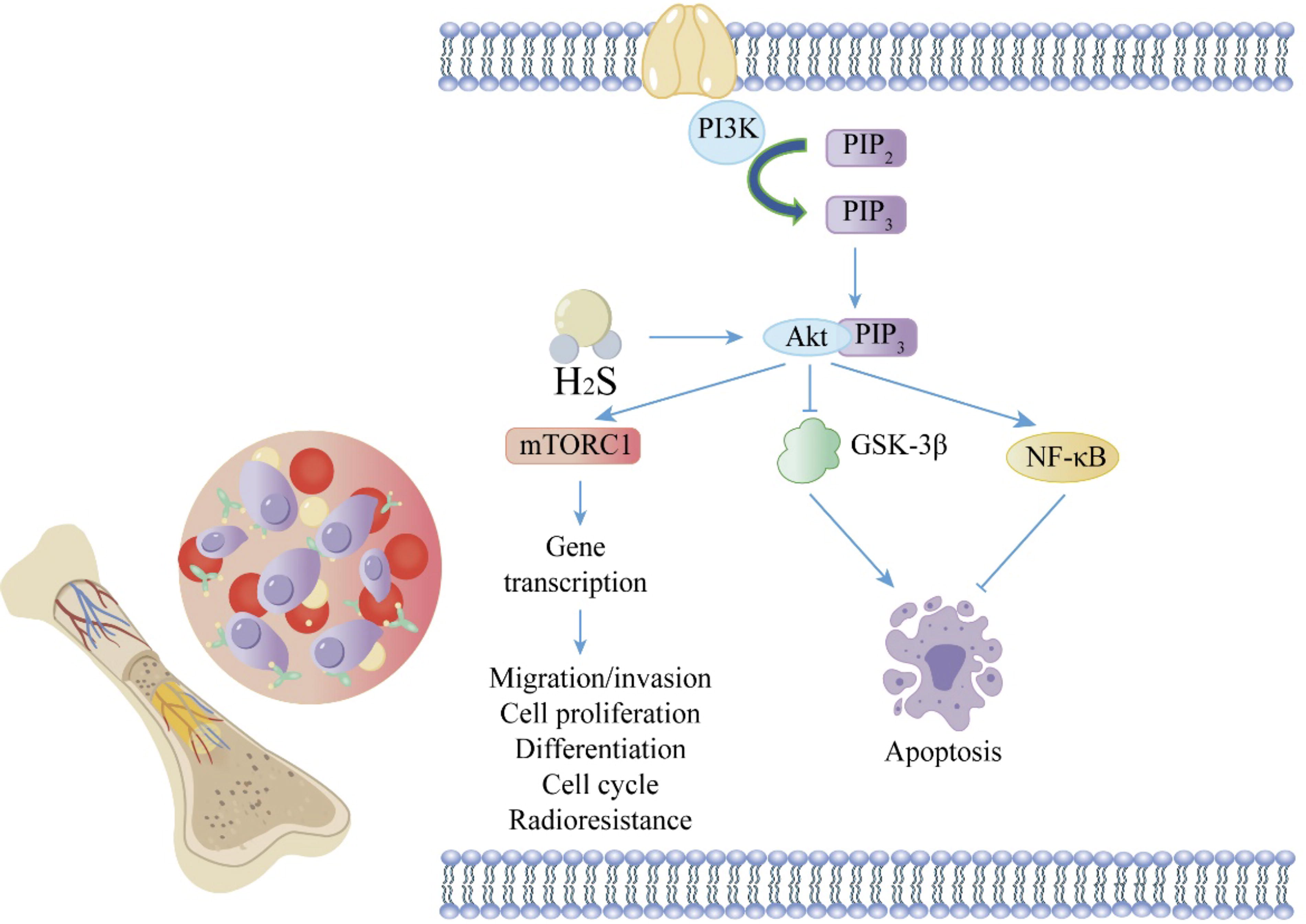
The PI3K/Akt pathway plays a significant role in the occurrence and development of multiple myeloma. In the PI3K/Akt pathway, PI3K phosphorylates PIP2, converting it to PIP3. PIP3 binds to Akt, recruiting Akt to the multiple myeloma cell membrane and thus activating it. Akt then phosphorylates GSK-3β on serine 9, reducing its phosphorylation level and inducing apoptosis. Additionally, Akt regulates cell growth through modulation of NF-κB and mTORC signaling pathways. H2S significantly increases Akt phosphorylation, thereby enhancing the PI3K/Akt pathway and promoting multiple myeloma proliferation. The tip arrows indicate the effect of promoting generation or activating expression; flat-end arrows indicate inhibition of generation or inhibition of expression. Akt, protein kinase B; GSK-3β, glycogen synthase kinase-3β; mTORC, mammalian target of rapamycin complex; NF-κB, nuclear factor-κB; PI3K, phosphatidylinositol 3-kinase; PIP2, phosphatidylinositol 4,5-bisphosphate; PIP3, phosphatidylinositol 3,4,5-trisphosphate. Color images are available online.
OS is the most common primary malignant tumor of bone characterized by high local invasiveness and a tendency to metastasize. In both primary OS and pulmonary metastatic OS, the expression of CBS and especially CSE is decreased (Gong et al., 2022). However, whether supplementation of H2S can inhibit OS remains unknown. Doxorubicin (DOX) is one of the first-line drugs for OS treatment, but its effectiveness is limited by the efflux pump P-glycoprotein (Pgp) and the risk of cardiotoxicity. Gazzano et al. demonstrate that intravenous injection of 200 μL of the sterile physiological solution containing 5 mg/kg of synthetic DOX conjugated with H2S-releasing moieties exhibits reduced cardiotoxicity and enhanced efficacy against Pgp-overexpressing OS cells compared with DOX alone. Hyaluronic acid liposomes containing H2S-releasing DOX effectively induce cell death in multidrug-resistant OS cells, thereby inhibiting OS growth (Gazzano et al., 2019). However, whether H2S alone can inhibit OS also lacks direct evidence. In conclusion, the effect of H2S on OS requires more experiments to demonstrate.
The Role of H2S in Skeletal Muscle
Muscle atrophy and muscle weakness
The study by Majumder et al. shows that HHcy caused by CBS deficiency can lead to oxidative stress and endoplasmic reticulum (ER) stress responses, resulting in skeletal muscle atrophy. On the one hand, the expression levels of Atrogin-1 and MuRF-1 are increased by inducing phosphorylation of JNK, c-Jun, and FOXO1A. On the other hand, pro-inflammatory cytokines, such as interleukin (IL)-6 and tumor necrosis factor (TNF)-α, are upregulated (Majumder et al., 2018; Majumder et al., 2019). In addition, during hind limb ischemia, HHcy caused by CBS deficiency can also aggravate skeletal muscle dysfunction by attenuating angiogenesis via reducing the levels of hypoxia-inducible factor-1α and peroxisome proliferator-activated receptor gamma coactivator-1α in muscle fibers and inhibiting the vascular endothelial growth factor-endothelial NO synthase-NO signaling pathway (Veeranki et al., 2014). HHcy-induced skeletal muscle weakness is involved in mitochondrial dysfunction and epigenetic modifications (Veeranki et al., 2015). On the one hand, HHcy leads to decreased muscle performance by hindering the formation of large muscle fibers, thus affecting the movement of CBS+/− mice. On the other hand, mitochondrial function can be impaired by HHcy via upregulation of miR-494 and reduction of mitochondrial transcription factor A, thereby inhibiting the production of ATP and causing skeletal muscle weakness. In skeletal muscle cells treated with Hcy, NaHS can reverse oxidative stress and alleviate the elevation of ER stress markers, such as glucose-regulating protein 78 (GRP78), activating transcription factor 6, phosphorylation of inositol-requiring enzyme 1α, and pJNK both in vitro and in vivo (Majumder et al., 2018). The skeletal muscle shows the gene and protein expression of three enzymes that produce H2S. Aerobic exercise can increase endogenous H2S by upregulating the expression of H2S-producing enzymes (Du et al., 2019; Kar et al., 2019; Seifi et al., 2019; Shao et al., 2021; Strutynska et al., 2022; Wang et al., 2017; Yang et al., 2021). Studies have shown that exercise can alleviate the effect of HHcy on poor muscle remodeling caused by CBS deficiency (Winchester et al., 2018). However, there is still a lack of direct evidence to determine whether this effect is mediated by an increase in H2S content. In summary, a deficiency in CBS could cause a lack of H2S, resulting in the accumulation of Hcy, mitochondrial damage, oxidative stress, and ER stress, which in turn causes muscle atrophy and weakness. Furthermore, exogenous H2S can reverse this process (Fig. 4).
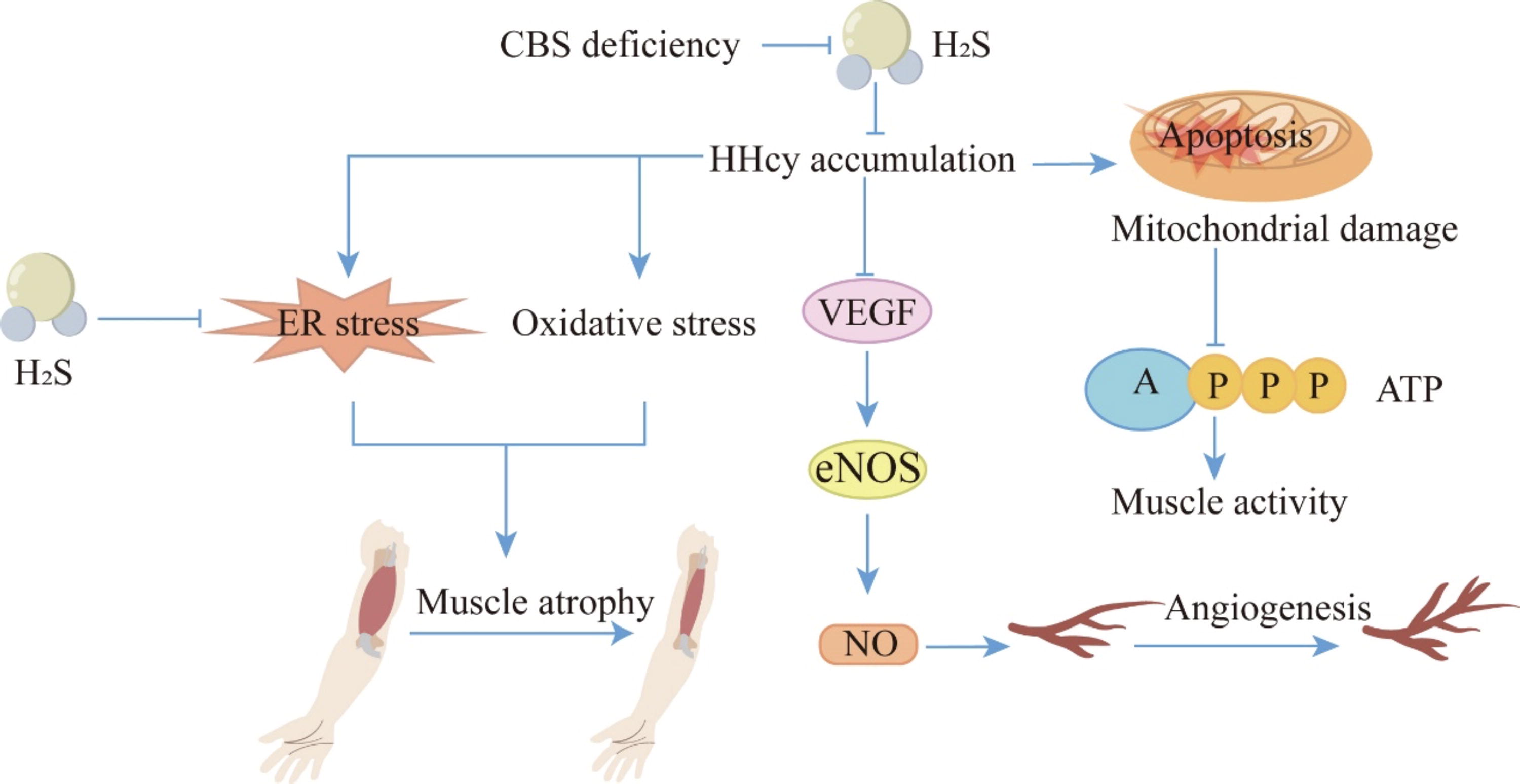
Inhibitory role of H2S in muscle atrophy and muscle weakness. CBS deficiency-induced HHcy can lead to oxidative stress and ER stress responses, causing skeletal muscle atrophy. Additionally, HHcy resulting from CBS deficiency can suppress the VEGF-eNOS-NO signaling pathway, which impairs angiogenesis and exacerbates skeletal muscle dysfunction. On the contrary, HHcy damages mitochondrial function, thus inhibiting ATP production and leading to skeletal muscle weakness. H2S can reverse oxidative stress and alleviate the increase in ER stress markers, thereby mitigating muscle atrophy. The tip arrows indicate the effect of promoting generation or activating expression; flat-end arrows indicate inhibition of generation or inhibition of expression. ATP, adenosine triphosphate; eNOS, endothelial nitric oxide synthase; ER, endoplasmic reticulum; NO, nitric oxide; VEGF, vascular endothelial growth factor. Color images are available online.
Duchenne muscular dystrophy (DMD) is a neuromuscular disorder caused by mutations in the DMD gene. The dystrophin protein is a crucial component of the dystrophin-associated protein complex. It not only links the actin cytoskeleton to the ECM but also regulates NO synthase activity and Ca2+ homeostasis (Calabrese et al., 2007; Łoboda and Dulak, 2024; Mancuso et al., 2008). DMD is characterized primarily by Ca2+ imbalance, chronic inflammation, autophagy changes, and dysregulation of angiogenesis. Heart failure is the leading cause of death in DMD patients (Kaziród et al., 2022). In addition to traditional corticosteroid treatments, researchers are exploring new therapeutic strategies such as H2S donors to find more effective treatments. Due to its anti-inflammation, antioxidant (Shefa et al., 2018), antifibrosis, and cardioprotective properties (Kondo et al., 2013), H2S is considered a potential therapeutic molecule for DMD. Data suggest that H2S has shown effects on muscle wasting in Caenorhabditis elegans and mouse models (Ellwood et al., 2021), but research on cardiac protection is still ongoing. Currently, research on H2S in DMD is in its early stages, with NaHS shown to moderately alleviate muscle weakness and cardiomyopathy in mdx mice models (Kaziród et al., 2022). SG1002 is the sole donor that can be 100% converted to H2S. Clinical trials have confirmed the safety and bioavailability of H2S and NO in heart failure patients. In mouse studies, 20 mg/kg/day SG 1002 was used (Kondo et al., 2013). Even at the highest dose of 800 mg, administered twice daily for 7 days, no serious side effects are observed, demonstrating good safety and tolerability (Polhemus et al., 2015). SG1002 can increase mitochondrial content in the heart and improve cardiac function (Shimizu et al., 2018), showing potential efficacy for heart failure patients. SG1002 has also achieved promising results in animal models of DMD, atherosclerosis (Bibli et al., 2019), diabetic cardiomyopathy (Barr et al., 2015), and ischemic injury (Rushing et al., 2019). AP39, a mitochondria-targeted H2S donor, has been shown to maintain mitochondrial integrity, reduce oxidative stress, and improve neurological function in cardiac arrest and resuscitation-induced neurodamaged models and also exhibits cardiac protective effects (Karwi et al., 2017). H2S, due to its anti-inflammatory, antioxidant, and anti-fibrotic properties, as well as its ability to promote angiogenesis and cardiac protection, needs to be further investigated for its potential in DMD treatment, which includes studies on its effects on muscle cell membrane stability and its role in enhancing muscle cell regeneration.
Long-term use of GCs can lead to skeletal muscle atrophy and capillary rarefaction. Research by Mohamed Adel et al. indicates that Dex-treated mice significantly reduce the mass, length, and cross-sectional area of the soleus muscle, along with decreased contractility. Additionally, markers of oxidative stress, serum potassium, and creatine kinase levels are significantly increased, while myostatin (a substance that inhibits skeletal muscle growth) levels are significantly elevated, and the expression levels of growth factors are notably reduced. Furthermore, capillary density is significantly decreased. In comparison to the GC group, intraperitoneal injection of 5 mg/kg/day NaHS can improve contractility, reduce oxidative stress and muscle atrophy, and increase capillary density (Adel et al., 2022). A natural H2S-releasing molecule, sulforaphane (30 μM), can prevent skeletal muscle damage in sarcopenia by reducing Dex-induced oxidative imbalance (Micheli et al., 2022).
Malignant hyperthermia with excessive muscle contractions
Excessive contraction of skeletal muscles is a common complication of malignant hyperthermia (Vellecco et al., 2016). Molecular analysis has shown that the content of H2S in the gluteus maximus muscle of patients with malignant hyperthermia is ∼10 times higher than that in healthy subjects, primarily derived from CBS (Vellecco et al., 2016). Treatment with 0.5, 1, and 3 mM NaHS induced contractions in skeletal muscle cells in patients with malignant hyperthermia, further supporting the above viewpoint. Caffeine or halothane treatment can result in skeletal muscle reactions that are similar to those results in typical malignant hyperthermia. Normal skeletal muscle does not exhibit contractions following caffeine or halothane treatment; however, NaHS treatment enhances the contraction response of normal skeletal muscle to caffeine or halothane, suggesting that elevated H2S concentration may be a crucial step in promoting skeletal muscle contraction under hyperthermic conditions. Blocking the ATP-sensitive potassium channels, a well-recognized role of H2S, can reduce skeletal muscle contraction under hyperthermia. Further research indicates that persulfidation of Kv 7 channels in skeletal muscle from patients with malignant hyperthermia is significantly higher than that in normal biopsies. Moreover, exposure to the Kv 7 channel opener retigabine leads to abnormal contraction in malignant hyperthermia muscle biopsy tissue (Vellecco et al., 2020), suggesting that H2S regulates Kv 7 channels through persulfidation, thereby triggering skeletal muscle contraction under hyperthermia.
Skeletal muscle ischemia and IRI
Skeletal muscle ischemia caused by impaired blood supply is a common clinical issue, which may result in intermittent claudication or even amputation. Compared with healthy individuals, the skeletal muscle of patients with critical limb ischemia (CLI) experience increased oxidative stress, leading to a significant decrease in the mRNA and protein levels of CBS, CSE, and 3-MST, along with a notable decrease in H2S levels. Additionally, activation of Nrf2 and antioxidant proteins such as copper/zinc-SOD, catalase, and glutathione peroxidase are significantly reduced, while malondialdehyde (MDA) and protein carbonyl formation are significantly increased. The results suggest that H2S deficiency-induced cellular oxidative stress may exacerbate muscle damage in patients with CLI (Islam et al., 2015).
Skeletal muscle ischemia causes an initial stress in skeletal muscle, and then the reintroduction of oxygenated blood to the ischemic tissue produces a large amount of ROS, which further exacerbates the stress. In skeletal muscle IRI rats, endogenous H2S levels and production are reduced, accompanied by severe oxidative stress, indicating that H2S imbalance may promote oxidative stress. Importantly, pretreatment with intraperitoneal injection of 56 μmol/kg NaHS in rats reduces MDA levels, decreases H2O2 and superoxide anion levels, but increases SOD activity and protein expression in skeletal muscle, thereby preventing IRI (Du et al., 2013). These results indicate that H2S protects skeletal muscle by combating oxidative stress. Numerous studies have demonstrated that skeletal muscle IRI can be significantly reduced by H2S in vivo and in vitro (Du et al., 2013; Henderson et al., 2010; Henderson et al., 2011; Zhong et al., 2021). By enhancing the activity of SOD and reducing the levels of MDA, H2O2, and superoxide anion, endogenously produced H2S in rat skeletal muscle can play a protective role against oxidative stress and inflammation (Chen et al., 2013). Exogenous supplementation of NaHS (5 μmol/L) via tail vein injection in rats can also reduce skeletal muscle injury after limb IRI by decreasing the content of necrotic material (including myoglobin, lysophosphatidylcholine, and lipid peroxidation) and increasing mitochondrial membrane potential and ATP content (Fu et al., 2019). CSE is the main H2S-producing enzyme in skeletal muscle, and CSE deficiency will aggravate skeletal muscle injury (Zhang et al., 2021b; Zhang et al., 2020b). During skeletal muscle contusion, treatment with 28 μmol/kg NaHS for 15 days can reduce the expression levels of M1 macrophages, profibrogenic cytokines, proinflammatory cytokines, chemokines, MMPs, and oxidative stress factors; inhibit inflammation and oxidative stress; reduce skeletal muscle fibrosis; and partially prevent skeletal muscle injury in mice (Zhao et al., 2020a).
Metabolic disorder of skeletal muscle
A previous study has shown that inhalation of 5 ppm H2S during exercise can significantly reduce oxygen uptake, resulting in increased lactic acid in muscle. Due to the increased level of lactate dehydrogenase, the decreased content of cytochrome C oxidase, and the inhibition of citrate synthase activity, the aerobic metabolism of muscle is inhibited, thus promoting the transformation from aerobic metabolism to anaerobic metabolism. However, heart rate and lung function are not affected (Bhambhani et al., 1996). It is notable that no inhibition of aerobic metabolism caused by H2S is observed in females, suggesting that there are gender differences in the effects of H2S. The reasons for these differences remain unclear. One possible reason could be the beneficial effects of estrogen on skeletal muscle, including alleviating muscle damage, enhancing post-injury repair, preventive effects on the number and function of muscle satellite cells, and potential antioxidant effects following injury, exercise, and/or mitochondrial stress (Liao et al., 2019; Velders and Diel, 2013). However, due to the small sample size in a previous study (only 25 participants, including 12 females), the statistical significances for many indicators are low. Therefore, the existence of such gender differences warrants further investigation.
The Role of H2S in OA
OA, a common degenerative joint disease, is characterized by inflammation and oxidative stress (Creamer and Hochberg, 1997). In the pathogenesis of OA, fibroblast-like synoviocytes (FLS) are key participants, which can produce proinflammatory cytokines and matrix-degrading enzymes (Sieghart et al., 2015). There are a large number of inflammatory mediators in the synovial fluid of patients with OA, such as IL-1β, TNF-α, MMPs, prostaglandin E2 (PGE2), cyclooxygenase (COX)-2, and NO (Amin et al., 2000). IL-1β and TNF-α are two major pro-inflammatory cytokines in the process of OA (Robinson et al., 2016). IL-1β and TNF-α can stimulate articular chondrocytes and synoviocytes to produce catabolic proteases, such as MMPs, which are involved in the degradation of elastin in the ECM (Wojdasiewicz et al., 2014). H2S is widely recognized as an endogenous mediator of anti-inflammation, and the lack of H2S will aggravate OA (Burguera et al., 2014; Nunomura et al., 2022; Rivers et al., 2012; Song et al., 2024; Whiteman and Winyard, 2011). In OA, the decrease in 3-MST levels may partially lead to a decrease in H2S levels, which may be a factor in the aggravation of joint calcification and OA (Burguera et al., 2020; Nasi et al., 2020). A study has found that CSE knockdown could exacerbate pathological calcification (PC), and PC would lead to cartilage degradation in OA (Nasi et al., 2021). H2S exhibits anti-inflammatory and anti-catabolic properties in IL-1β-stimulated OA (Burguera et al., 2014). In FLS, the stimulation from IL-1β can be antagonized by 1 mmol/L NaHS through MAPK and PI3K/AKT pathways (Sieghart et al., 2015). In chondrocytes, H2S can also antagonize IL-1β-induced inflammation and mitochondrial dysfunction-related apoptosis in the same way. Meanwhile, NaHS (0.06, 0.15, 0.3, and 0.6 mM) can reduce the secretion of MMP-13 by inhibiting the expression (or production) of inducible NO synthase/NO and COX-2/PGE2, thereby inhibiting inflammation, protecting cartilage, and ultimately treating OA (Ha et al., 2015; Wang et al., 2021a).
IL-1β and TNF-α can also induce the production of ROS in cartilage. In addition to causing oxidative stress to participate in the pathogenesis of OA (Lepetsos and Papavassiliou, 2016), ROS are also harmful substances involved in cartilage degradation, which can destroy the structure and function of cartilage and lead to damage (Henrotin et al., 2003; Liu et al., 2014a). It is well known that physiological concentrations of H2S scavenge ROS in a variety of ways, including promotion of glutathione synthesis (Lu et al., 2008), improvement of the scavenging of ROS by antioxidant enzymes (Searcy et al., 1995), and inhibition of the activity of ROS synthase (Xu et al., 2011), suggesting its potential to reduce OA by inhibiting ROS. A recent study shows that single intra-articular injection of 200 μM GYY4137 significantly reverses oxidative stress and oxidative damage of deoxyribonucleic acid, protein and lipid membranes caused by excessive ROS in surgery-induced OA rats. GYY4137 can not only reduce oxidative stress by increasing Nrf2 level but also inhibit ER stress and the apoptosis of chondrocytes through GRP78/mTOR pathway in rats (Vaamonde-García et al., 2020).
The Role of H2S in Herniated Intervertebral Disc
The intervertebral disc is the largest avascular organ in the human body, consisting of the central nucleus pulposus (NP) and the surrounding annulus fibrosus (Sakai and Grad, 2015). Intervertebral disc degeneration (IDD) is a major cause of neck, shoulder, and lower back pain. The pathological changes associated with IDD primarily involve NP cell aging and excessive apoptosis, degeneration of the ECM, annulus fibrosus fibrosis, and inflammation (Xin et al., 2022). The goals of IDD treatment are the alleviation of inflammation and the protection of cells from apoptosis. Studies have found that the expression levels of CBS and CSE are significantly reduced in human degenerated NP tissues and in NP cells treated with IL-1β. Administration of the H2S donor NaHS has shown protective effects by inhibiting ER stress and mitochondrial dysfunction induced by IL-1β stimulation in vitro, which is associated with the activation of the PI3K/Akt and ERK1/2 signaling pathways. Additionally, in a rat tail disc degeneration model induced by percutaneous needle puncture, NaHS administration partially reverses disc degeneration. The results indicate that H2S plays a protective role in IDD, with its potential mechanisms involving the suppression of ER stress and mitochondrial dysfunction in NP cells induced by IL-1β through the PI3K/Akt and ERK1/2 signaling pathways (Xu et al., 2017). In IDD, pro-inflammatory factors such as TNF-α, IL-1β, and IL-6 are significantly increased, leading to changes in disc structure. ECM-derived matrix proteins induce inflammation in the tissue microenvironment. Among these, the matrix protein N-acetylated proline–glycine–proline (N-Ac-PGP) is cleaved from collagen by MMP8, MMP9, and prolylcarboxypeptidas. The level of N-Ac-PGP is positively correlated with the severity of IDD. Transplantation of NP cells treated with N-Ac-PGP into rat intervertebral discs causes degenerative changes, resulting in disc collapse (Feng et al., 2017). Inflammation signals activated by Toll-like receptors (TLRs) also contribute to IDD development. The expression levels of TLRs 1/2/3/4/6 are dependent on the severity of IDD, with various stimuli-activated inflammation signals accelerating the progression of IDD. Research has found that H2S donor JK1 loaded into a pH- and enzyme-responsive hydrogel prolongs its retention in degenerative discs, reducing NP cell apoptosis and modulating the NF-κB signaling pathway to inhibit ECM degradation and prevent disc degeneration. H2S and its donors can effectively suppress inflammation, oxidative stress, and aging, thereby preventing disc cell degeneration and providing a potential therapeutic option for IDD (Zheng et al., 2019).
Potential Therapeutic Modalities of H2S
H2S has a variety of potential therapeutic effects in the treatment of musculoskeletal diseases. Analyzing papers related to H2S and musculoskeletal diseases in the Web of Science Core Collection database from 2012 to 2024 by means of bibliometric analysis, we find out the research trends and potential frontier hot spots in this field and identify the three diseases that have received the most attention in this field, including bone tumor, periodontitis, and OA. The related studies on these diseases are mainly involved in the fields of molecular biology, pharmacology, and immunology, suggesting that interdisciplinary convergence research represents a current trend in the scientific field. Keyword bumping analysis through CiteSpace reveals that early studies focus on exploring the potential application of H2S in skeletal muscle diseases. In recent years, there has been increasing interest in specific areas such as the development of drug delivery systems, innovative applications of photothermal/photodynamic therapies and fluorescent probe technologies, and the regulation of the tumor microenvironment.
Due to the high volatility and rapid metabolism of H2S, accurately controlling its release rate is crucial. Using nanosystems for drug delivery to control H2S may hold great potential for clinical treatment in various diseases (Nazarnezhada et al., 2020). A silk fibroin porous scaffold loaded with GYY4137 is reported to promote the differentiation of human BMMSCs into OB (Gambari et al., 2019; Raggio et al., 2018). Hydrophobic phase-change materials loaded with NaHS can alleviate diabetic wounds (Lin et al., 2017). Nanofibrous membrane loaded with NaHS could promote wound healing (Lian et al., 2021). Several novel H2S donor-based nanoscale release systems have been designed and have shown promising results in various diseases. These novel H2S donors include 2-acetyloxybenzoic acid 4-(3-thioxo-3H-1,2-dithiol-5-yl)phenyl ester (Lu et al., 2021; Zhang et al., 2021a), JK1 (Wu et al., 2016; 2019; Zhao et al., 2020b; Zheng et al., 2019), androgen deprivation therapy (ADT) (Chen et al., 2020; Hasegawa and van der Vlies, 2014; Liu et al., 2019; Liu et al., 2017; Takatani-Nakase et al., 2017), S-aroylthiooxime (Wang and Matson, 2019), trisulfide (Dao et al., 2021; Dao et al., 2020; Ercole et al., 2017), arylthioamide (Ercole et al., 2016; Lee et al., 2021), 2-aminopyridine-5-thiocarboxamide (Liang et al., 2019), N-(benzoylthio) benzamides (Feng et al., 2015), α-thioetherketone (Xiao et al., 2017), manganese sulfide (He et al., 2020), ferrous sulfide (Xie et al., 2020), polysulfide (2, 2′-dipyridyl tetrasulfide) (Li et al., 2021; Li et al., 2020), phenyl substituent ADT-DOX conjugate (Gazzano et al., 2019), L-lys (Wan et al., 2021), thiosulfate cyanide sulphurtransferase (Mauretti et al., 2016), and S-propargyl-cysteine (Yu et al., 2021) (Table 1).
H2S-Releasing Compounds and Nanosystems
ACS14, 2-acetyloxybenzoic acid 4-(3-thioxo-3H-1,2-dithiol-5-yl)phenyl ester; ADSC, adipose-derived stem cell; ADT, androgen deprivation therapy; APTC, 2-aminopyridine-5-thiocarboxamide; ECs, endothelial cells; Balb/c, Bagg albino c; BSA, bovine serum albumin; Cys, cysteine; FeS, ferrous sulfide; GSH, glutathione; GYY4137, morpholin-4-ium-4-methoxyphenyl (morpholino) phosphinodithioate; H2O2, hydrogen peroxide; H2S, hydrogen sulfide; hCPCs, human cardiac c-Kit+ progenitor cells; hMSCs, human mesenchymal stem cells; HEK293, human embryonic kidney cells; HepG-2, human hepatocellular carcinoma; HPAEC, human pulmonary artery endothelial cell; HUVECs, human umbilical vein endothelial cells; ICR, Institute of Cancer Research; LPM, loperamide hydrochloride; MCF, mouse embryonic fibroblasts; MCF-5, mossy cell fibers; MnS, manganese sulfide; N/A, no content; NaHS, sodium hydrosulfide; NIH 3T3 cells, mouse embryonic fibroblast cells; NSHD, N-mercapto-based H2S donor; PCL, polycaprolactone; PEG, polyethylene glycol; PHDC, peptide-H2S donor conjugate; PTA, PLGH-thiobenzamide; ROS, reactive oxygen species; SATO, S-aroylthiooxime; SD, Sprague–Dawley; SMCs, smooth muscle cells; SPRC, S-propargyl-cysteine; TST, thiosulfate sulfurtransferase; UV, ultraviolet.
Discussion
H2S is crucially involved in musculoskeletal diseases, including osteoporosis, periodontitis, MM, muscle atrophy and muscle weakness, muscle ischemia and IRI, OA, and herniated intervertebral disc, which may become new targets for the diagnosis and treatment of these diseases. Although there has been significant progress in the multiple roles of H2S in musculoskeletal diseases, as well as its production and metabolism in the musculoskeletal system, many researchers are still striving to understand specific mechanisms and signaling pathways in the research fields. There is no doubt that H2S will play an increasingly essential role in the treatment of musculoskeletal diseases, and the exploration of new treatment methods based on H2S will be promising research directions.
Footnotes
Authors’ Contributions
Y.-F.L.: Conceptualization, writing—original draft, and revision of draft. Y.-X.Z.: Writing—original draft. Y.-W.Z.: Revision of draft. A.-Q.T.: Drawing and organizing references. H.-B.L.: Bibliometrics. Y.-L.Y.: Revision of references. Y.-K.Z.: Conceptualization. X.-Y.J.: Project administration. D.-D.W.: Visualization, supervision, and writing—review and editing.
Author Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This work was supported by grants from the