
Abstract
Life on Mars, if it exists, may share a common ancestry with life on Earth derived from meteoritic transfer of microbes between the planets. One means to test this hypothesis is to isolate, detect, and sequence nucleic acids in situ on Mars, then search for similarities to known common features of life on Earth. Such an instrument would require biological and chemical components, such as polymerase and fluorescent dye molecules. We show that reagents necessary for detection and sequencing of DNA survive several analogues of the radiation expected during a 2-year mission to Mars, including proton (H-1), heavy ion (Fe-56, O-18), and neutron bombardment. Some reagents have reduced performance or fail at higher doses. Overall, our findings suggest it is feasible to utilize space instruments with biological components, particularly for mission durations of up to several years in environments without large accumulations of charged particles, such as the surface of Mars, and have implications for the meteoritic transfer of microbes between planets. Key Words: Laboratory simulation experiments—Life-detection instruments—Nucleic acids—Mars—Panspermia. Astrobiology 13, 68–78.
1. Introduction
N
We are building an instrument and developing protocols to test this hypothesis in situ on Mars by isolating, detecting, and sequencing nucleic acids (Ruvkun et al., 2002; Isenbarger et al., 2008a, 2008b). Our nominal approach relies on polymerase chain reaction (PCR) amplification as part of the sequencing process, because current single-molecule sequencing approaches are not yet suitable for space applications. The required reagents must function after months to years of space radiation exposure.
Many of these reagents, including enzymes, proteins, nucleotides, and dyes, have potential applications in other astrobiology investigations. For example, immunoassay (Parro et al., 2011; Sims et al., 2012) or oligonucleotide microarray approaches to life detection also utilize biological components (antibodies or nucleic acids) and fluorescent dyes. While these biochips avoid the complexity of PCR, they also lack its sensitivity. While immunoassays can target a variety of molecules, they are limited to a predefined set of targets and generally require a higher number of target molecules, whereas metagenomic sequencing can target any nucleic acid molecule. Oligonucleotide tiling microarrays also rely upon known or predicted sequences and suffer from high background noise. Tiles covering all possible sequences must be short and thus yield little new sequence information. Thus, immunoassay and metagenomic sequencing approaches are complementary; while the former offers a broader molecular diversity of predefined targets, the latter offers the potential for definitive and information-rich interrogation of any nucleic acid–based extant life.
To validate these reagents for use in situ on Mars and more generally in space, we exposed them to a variety of radiation sources selected to approximate the space radiation environment and compared their non-irradiated versus irradiated performance. We focused on radiation to the exclusion of other factors such as vibration or temperature because the former cannot be avoided and is essentially unique to the space environment. First, we briefly describe the main sources of space radiation and justify the target fluences or doses chosen for this study.
Space radiation is principally composed of galactic cosmic radiation (GCR) and solar radiation (Curtis and Letaw, 1989; Task Group on the Biological Effects of Space Radiation, 1996; Committee on Solar and Space Physics and Committee on Solar-Terrestrial Research, 2000; Committee on the Evaluation of Radiation Shielding for Space Exploration, 2008). By particle flux at peak during solar minimum, GCR consists of 90% protons (total flux ∼4 cm−2 s−1), 9% helium ions (alpha particles), and 1% heavy ions (HZE) with a broad fluence peak near or below 1 GeV. Solar radiation is mainly composed of low-energy protons (<1 MeV, 1012 cm−2 s−1), solar storm protons (∼1 MeV, 105 cm−2 s−1), and solar flare protons (10 to 103 MeV at 105 to 10 cm−2 s−1); the latter two are associated with a solar particle event (SPE). We neglect UV radiation because it can be blocked with minimal shielding. Energized electrons are principally a problem near planets with magnetic fields, such as in Earth's outer Van Allen belt, where high-energy solar wind electrons (0.1–10 MeV) can be trapped by interactions with Earth's magnetic field. The Van Allen belts also contain high-energy protons, up to >108 cm−2 s−1 at >0.1 MeV, or <103 cm−2 s−1 at >400 MeV, in the most intense regions. These high-energy protons have a flux versus energy profile similar to SPEs. Trips to Mars based on standard chemical propulsion will have transit times through the Van Allen Belts that limit their contribution to total dose.
The contribution of neutrons to the total radiation dose (10–60%) depends upon spacecraft shielding, as it is derived in large part from HZE secondary radiation (Bartlett et al., 2006). Neutrons also contribute significantly to total dose on the surface of Mars; for example, the Mars Odyssey high-energy neutron detector mapped near-surface (<1 m depth) water on Mars by taking advantage of hydrogen's ability to moderate these secondary neutrons (Boynton et al., 2002). Neutron fluxes at 1.5 m above the surface of Mars have been estimated as 80 cm−2 s−1 (typical) or up to 145 cm−2 s−1 during a SPE (Le Postollec et al., 2009a, 2009b). The majority of martian neutrons' energies range from epithermal (10–1000 of keV) to fast (between 1 and 104 MeV), with the bulk below 10 MeV and peaks at 0.6 and 6 MeV (Le Postollec et al., 2009a).
Prior irradiation studies have tended to focus on the cellular through organism level or on nonbiological spacecraft components such as electronics and spacecraft materials. More recently, instruments have been proposed that involve chemical and biological components. For example, the Signs of Life Detector instrument (Parro et al., 2005, 2011) and the Life Marker Chip (Sims et al., 2012), payload concepts originally planned for the European Space Agency 2018 ExoMars mission, use antibodies to recognize protein domains and other biochemical structures. A related study (Le Postollec et al., 2009a) demonstrated that antibodies and fluorescein dye were able to retain their function after 0.5 or 5 MeV neutron bombardment at a fluence equivalent to 1000 months on the surface of Mars. Also, exposure of the fluorescent dyes fluorescein and Alexa Fluor 633 to low-energy proton and alpha radiation, simulating unprotected exposure during a cruise to Mars, found that both dyes retained their fluorescent properties after irradiation (Thompson et al., 2006). More recently, it was shown that antibodies and fluorophores survive Mars-relevant doses of gamma irradiation (de Diego-Castilla et al., 2011), and DNA aptamers retain their binding affinity after 2 MeV proton exposure at fluences of 109 to 1011 cm−2 (Baqué et al., 2011).
2. Materials and Methods
2.1. Target fluence or dose
We selected target fluences or water-equivalent doses at levels conservatively above what would be expected for a 2-year mission duration (Table 1). We carried out a neutron exposure at the MIT Nuclear Radiation Laboratory (NRL), a proton exposure at the Massachusetts General Hospital (MGH) Burr Center for Proton Therapy, and a proton and heavy ion exposure at the Brookaven NASA Space Radiation Laboratory (NSRL) (Table 2).
Energy is target and [achieved] in MeV/nucleon; SPE indicates solar particle event energy distribution. ND, not determined; NA, not applicable. For non-NSRL exposures we controlled the fluence, while for NSRL exposures we controlled the water-equivalent dose, and the fluence and flux are calculated from linear energy transfer tables.
For the MIT NRL neutron exposure, we selected a fluence above the worst-case scenario (145 neutrons cm−2 s−1 during a SPE) for 2 years on the surface of Mars (Le Postollec et al., 2009a, 2009b).
At MGH, we targeted the indicated fluences for two monoenergetic proton beams (25 MeV, 40 MeV), a high fluence polyenergetic SPE simulation (equivalent in dose to one day near Jupiter's moon Europa), and an extremely high fluence of 25 MeV protons (equivalent in dose to 2 or more years near Europa). For these proton exposures we controlled fluence. Proton fluence due to SPEs was based on the 370 known events over 2/1/1954 to 12/31/2007; weighting the event size by the predicted frequency [from Table 3 in Kim et al. (2009)] gives an approximate total fluence of 3×109 protons/cm2 with energy ≥30 MeV for a 2-year mission. As a target, we chose a target exposure around 30-fold higher (1011) to provide a margin of safety.
Energy is target value. ND, not determined (because components worked together).
Approximated by Maxwell distribution with beam energy 2.1 MeV, most probable energy 0.7 MeV.
Master mix includes Taq polymerase, PCR buffer, dNTPs, primers (excludes template and SYBR Green I).
Ribosomal primer 357F failed.
Some nonspecific amplification occurred but no different than amplification observed with 357F primer alone, indicating off-target effect such as primer-dimer.
At NSRL, we targeted 1 and 5 Gy doses of 1 GeV oxygen and iron ions, and a proton SPE simulation (Supplementary Table S2; Supplementary Data are available online at
2.2. Neutron exposure
Samples were irradiated at the MIT Nuclear Radiation Laboratory by using a californium (252Cf) source with reported emission 2.4×108 s−1 and reported dose rates at 1 m in air of 2.31 mSv/h (neutron) and 0.168 mSv/h (gamma rays) at its original mass of 104.3 μg. Assuming quality factors of 10 and 1, respectively, these dose rates correspond to 0.23 and 0.17 mGy/h. The calculated emission rate was 1.64×108 s−1 after adjustment for the exposure time (t 1/2=2.64 years, t exposure=1.44 years after calibration). The desired neutron fluence was set at 1010 cm−2, just above the worst-case estimate for 2 years on the surface of Mars, or around twice the typical fluence expected during this period.
We treated the source as a point mass located at the active end of the source and controlled the radial distance R of reagents from the source. Since californium also emits alpha particles, R should be greater than the distance an alpha particle will travel in air (∼4 cm); in practice, alpha particles emitted within the source may be captured there as well. Second, R must be significantly smaller than the mean free path of a fast neutron in air (approximated as 78% nitrogen, 22% oxygen), which is ∼20 m. Finally, to keep manufacturing costs low and limit sample exposure time we chose a small radius, R=10 cm, giving an exposure time of 21.2 h.
We printed (Elite 3D, Dimension Printing) an ABS plastic tube rack with a spherical surface (Supplementary Fig. S1) to hold standard 1.5–2 mL tubes at this distance from the source. In practice, the ends of the tubes ranged from around 6.5 to 10 cm away from the source, so we can expect a 2.4× higher fluence at the base of a long tube relative to the estimate. The mean free path of a neutron also contributed to total variation in fluence: 252Cf has a most probable neutron energy of 0.7 MeV and average neutron energy of 2.1 MeV (Hjalmar, 1955; Singleterry and Thibeault, 2000). In water, the neutron mean free paths at these energies are 2.3 and 4.2 cm. Based on atomic cross sections, after 2 cm of liquid depth, the fluence may drop by up to a factor of 2.5× or 1.7×, respectively, but the fluence may decline by less than this estimate due to elastic scattering of neutrons in water. Thus, we can expect the portion of a sample closest to the source to be irradiated above our desired fluence and the portion near 10 cm to be irradiated below our target fluence in the worst case. Both radial position and neutron mean free path contribute to dose variability, but in terms of average dose these two effects oppose each other. Finally, samples experienced additional doses associated with gamma irradiation from the 252Cf source and secondary production of gamma rays during elastic scattering of neutrons in water.
A 2.5 cm thick polystyrene sheet with a predrilled hole for the source was used to support the tube rack. After loading of samples, the rack was aligned to the source by using a predrawn circle on the sheet, and the rack and sheet were placed inside a cooler (Coleman Ultimate Extreme) on top of dry ice. Sample temperatures were measured before, during, and after the exposure (U12-015 HOBO temperature data logger, Onset Computer). Prior tests suggested that, due to poor air circulation, the air temperature could take longer than 30 min to transition from room temperature to below freezing, although once cold, temperatures would stay below freezing for several days. We dumped about 2 L room-temperature deionized water onto the dry ice before closing the lid, which caused CO2 sublimation and indicated freezing temperatures within about 5–10 min (approximately the response time of the data logger in still air). During exposure, the temperature sensor was placed on the polystyrene sheet but away from the source. After the exposure, the samples were repackaged into a freezer box and stored on dry ice until they were returned to −20°C in the lab.
2.3. Proton irradiation at MGH
Samples were exposed to high-energy protons at the Francis H. Burr Proton Therapy Center at Massachusetts General Hospital by using a cyclotron that can accelerate protons up to 235 MeV. We generated near-monoenergetic beams (Supplementary Table S1) with target energies of 25 and 40 MeV and target fluence of 109 cm−2 (estimated dose 3.6 Gy). Briefly, the source beam (229 MeV) passes through a lexan degrader, a Lucite contoured scatterer (which helps flatten the beam profile), and an ion chamber, with air losses bringing the beam to near 39 MeV. An additional Lucite sheet (degrader) was used for the 25 MeV target condition to bring the beam to 27 MeV. Samples were placed immediately behind the degrader for the 25 [27] MeV beam and at the same position for the 40 [39] MeV beam. We used a diode cross to align the beam to achieve a flat region about 8 cm across (Supplementary Fig. S2). Samples in thin-walled (∼0.25 mm) PCR tubes (polypropylene) were preloaded into the central 8×8 cm of thin-walled (∼0.25 mm) polyethylene 96-well PCR plates. These plates were kept on ice except during exposure, when they were exposed to room temperature and proton-mediated heating. The four samples at the extreme corner positions may have experienced beam intensities 25–50% lower due to beam non-uniformity. For the SPE exposure, we used a rotating energy shifter as previously described (Cascio and Sarkar, 2008) to approximate the energy spectrum of the September 1989 SPE. When using the Burr Center accelerator, this energy shifter closely approximates the relative proton fluence of that event for energies from 35 to 205 MeV. We also performed an extreme exposure, using the 25 MeV target setup with a target fluence of 1014 cm−2.
2.4. Proton and heavy ion irradiation at NSRL
Samples were irradiated at the NASA Space Radiation Laboratory at Brookhaven National Laboratory. Sample tubes were placed within a freezer box, and the outer row was loaded with water-filled tubes (Supplementary Fig. S3A). We used 1 GeV oxygen (O-16) and iron (Fe-56) ions generated by the Electron Beam Ion Source, and a proton SPE exposure was simulated by using the 200 MeV linear accelerator. Typical beam uniformity was <2% over a 13×13 cm area (Supplementary Fig. 3B–3D). The SPE exposure simulated the energy spectrum of the August 1972 SPE with the use of 11 monoenergetic beams from 50 to 150 MeV in 10 MeV increments; the full width at half maximum energy spread varied with the target energy level and ranged from 10–12 MeV at 50 MeV to 5 MeV at 150 MeV. Samples were kept in −20°C or on ice (cold packs) except during exposures and during activation checks. Additional beam characterization data is available online (
2.5. Reagents
In all irradiations, we included standard components of a PCR, including Taq polymerase and its associated PCR buffer (11647687001, Roche Diagnostics, Indianapolis, IN), deoxyribonucleotide triphosphate monomers (dNTPs, CB4420-2, Denville Scientific, South Plainfield, NJ), single-stranded DNA (ssDNA) primers targeting the ribosomal 16S gene (desalted custom oligos, Integrated DNA Technologies, Coralville, IA; for sequences see methods; volume varied depending upon synthesis yield, concentration 1 nmol/μL), and Escherichia coli genomic DNA (gDNA; 14380 10 MG, USB/Affymetrix, Inc., Santa Clara, CA). We also irradiated 100× SYBR Green I (diluted from S7585, Invitrogen, Carlsbad, CA), a dye that increases its fluorescence greatly upon binding double-stranded DNA (dsDNA), which is used to monitor amplification during quantitative (real-time) PCR (qPCR) (Zipper et al., 2004), as well as a qPCR mastermix (iQ SYBR Green Supermix, 170-8880, Bio-Rad, Hercules, CA).
In the NSRL exposures, we also included lyophilized buffer, dNTPs, primers, gDNA, and a lyophilized master mix in an 8-PCR-tube strip format (High Yield PCR EcoDry Premix, ClonTech, Mountain View, CA). To study the effect of lyophilization during irradiation despite possible effects of lyophilization itself, we first lyophilized (Virtis FreezeMobile 25EL, SP Scientific, Gardiner, NY) matching reagent tubes and then rehydrated half of the tubes; because the EcoDry Premix was already lyophilized, we simply rehydrated some of the tubes before irradiation.
All reagents were expected to remain frozen during the brief exposures, with the exception of the longest-duration proton exposure at MGH. Reagents were used at commercially provided concentrations and volumes except as indicated above and in Supplementary Table S4.
2.6. Reagent performance assessment
We used qPCR as our primary method to assess reagent performance. After qPCR analysis, melt curves were used to visualize melting domains, and gel electrophoresis was used to visualize the expected band. For each exposure, we carried out a separate experiment, including both irradiated reagents and non-irradiated controls. We first ran qPCRs with all irradiated components and then tested individual components if a PCR failed or showed reductions in performance. As an added check, selected PCR products were sequence verified.
For historical reasons, PCR assays used all-bacterial ribosomal 16S primers 357F-19 (5′ CTC CTA CGG GAG GCA GCA G 3′) and 1492R-22-Y6 (5′ TAC GGY TAC CTT GTT ACG ACT T 3′) and E. coli purified DNA. This simulates targeted PCR for in situ life detection, although our nominal approach is metagenomic sequencing and, hence, is not limited by knowledge of specific sequences. Beginning experiments were performed on MBS satellite 0.2G Thermal Cycler (Thermo Electron Corp, Marietta, OH) with parameters consisting of 1 min at 95°C, 30 cycles of 15 s at 94°C, 30 s at 55°C, 1 min at 72°C and held for 7 min at 72°C. PCR products were run on 1% agarose gels in 1× TAE (40 mM Tris acetate, 1 mM EDTA) and visualized with ethidium bromide. Real-time PCR was performed with 0.5× SYBR Green I. Thermocycling (Bio-Rad CFX96 Real Time System, C1000 Thermocycler) parameters included 5 min at 95°C, 30 cycles of 10 s at 94°C, 30 s at 60°C, 2 min at 72°C; followed by 7 min at 72°C. A melt curve was performed at 65–95°C with 0.5°C×5 s increments. Products were sequence verified through Sanger sequencing of amplicons with 16S reverse primer 907R-20-R15 (5′ CCG TCA ATT CCT TTR AGT TT 3′).
To assay for neutron-induced changes in SYBR Green I structure, we combined 25 or 40 μL 100× SYBR Green I with an equal volume of MeOH and injected into a Bruker Esquire 3000 Plus mass spectrometer in positive ion mode. We monitored a mass to charge (m/z) range of 50–1000 for 5 min.
3. Results
With the exception of the two highest-dose proton exposures, all conditions resulted in successful PCR amplification defined as the presence of the expected 1157 bp band during gel electrophoresis of each PCR product (Table 3). However, we observed some degradation in performance as described below.
3.1. Neutron irradiation
We detected no nuclear activation of the samples post-exposure. qPCR analysis revealed successful DNA amplification but some apparent degradation when all reagents were irradiated (Fig. 1A–1C, left). Samples with template (dark lines) generally amplified ∼10 cycles before samples without template (light colors) (Fig. 1A–1B, 1E), indicating only minor contamination in the negative controls. Melt curve analysis revealed a single (expected) product in samples with template, and probable primer-dimer or low-level contamination in samples without template (Fig. 1C). When only a single irradiated reagent was included, qPCR curves were similar to the non-irradiated case except for SYBR Green I (Fig. 1A, center, orange curve), which was similar to the all-irradiated case. The converse was also true: when PCR reactions included all irradiated reagents except the one indicated reagent (Fig. 1A, right), only lack of irradiated SYBR Green I mimicked the non-irradiated case. Substitution of irradiated SYBR Green I caused a reduction in end-point fluorescence (Fig. 1D, orange to red curves), while substitution of all other irradiated reagents caused essentially no change in the qPCR curves (Fig. 1D, red to brown curves). This reduction in end-point fluorescence could be associated with concentration differences that could be driven in part by irradiation-induced destruction or photo bleaching. To search for evidence of SYBR Green I destruction, we analyzed irradiated and non-irradiated SYBR Green via mass spectroscopy and identified four main mass/charge peaks (Fig. 1F). The main peak at 509.3 matches the predicted exact mass of SYBR Green I (C32H37N4S+, exact mass 509.27 amu); additional peaks at 255.1, 168.0, and 101.1 correspond closely to potential m/z peaks for charges z=+2 to +4. Thus, we found little evidence for neutron-induced damage to SYBR Green I at this fluence.
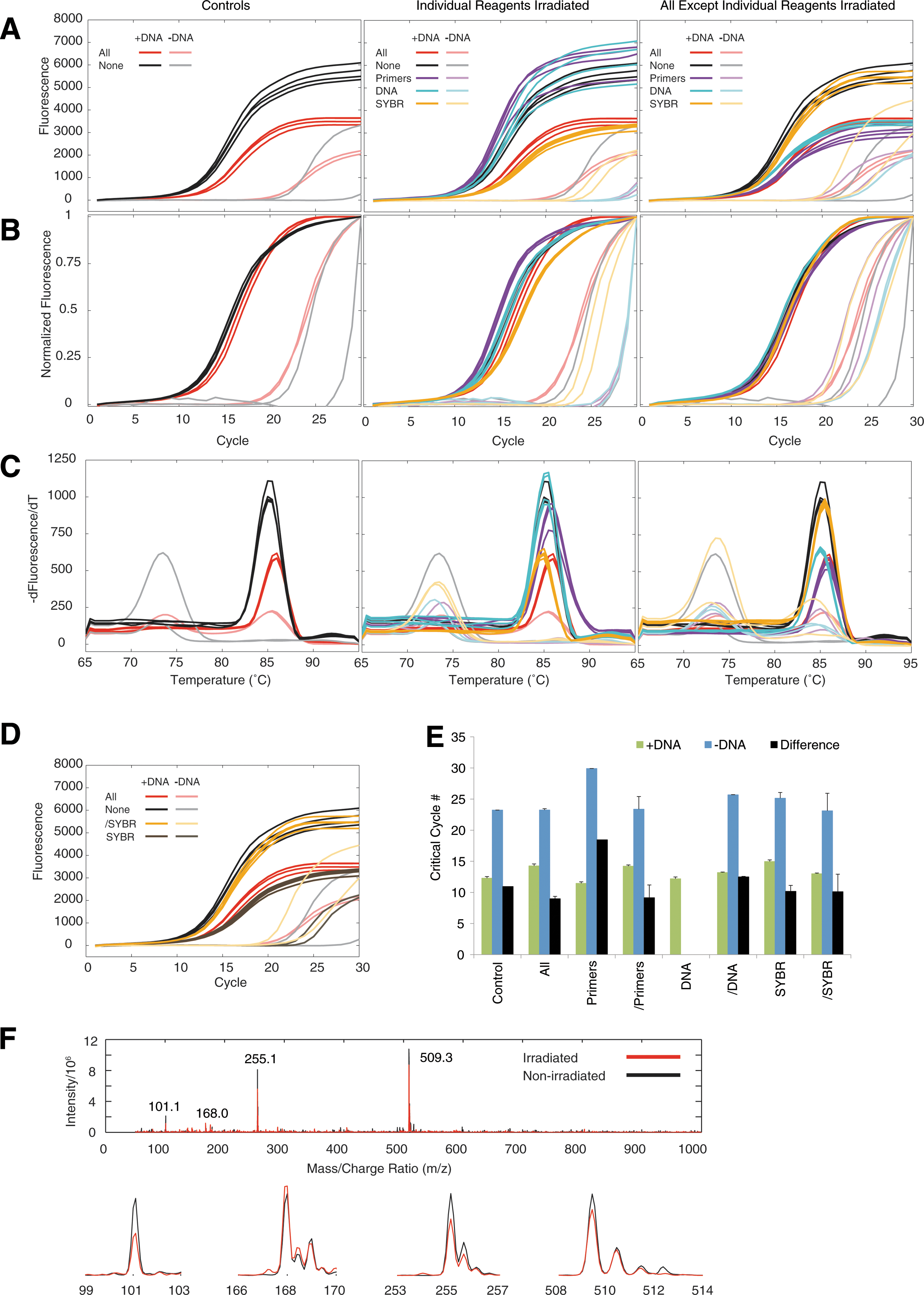
Impacts of neutron irradiation. (
3.2. Proton irradiation at MGH
We found successful amplification after low-dose monoenergetic proton irradiation as demonstrated by real-time PCR, melt-curve analysis, and gel electrophoresis (Fig. 2). Tubes with DNA (dark) amplified earlier (Fig. 2A, 2C) than no-template controls (light) whether the master mix used was irradiated (red) or not (black). However, the 15.6±4.9 (mean±SD) cycle difference between DNA and no-template controls dropped to 7.1±0.8 cycles after irradiation, due in part to critical cycle delay of 4.0±0.4 cycles for the irradiated templates (Fig. 2E). Melt-curve analysis (Fig. 2B, 2D) revealed three melting domains, indicating primer-dimer, background contamination, and the expected ribosomal amplification products. In a separate experiment where only the DNA template was irradiated, we also found successful amplification (Fig. 2F). Monoenergetic proton exposure at 40 MeV gave similar results (not shown). In contrast, at 180 Gy with a SPE spectrum, proton irradiation resulted in an inability to amplify irradiated DNA template and an inconsistent ability to amplify non-irradiated DNA template when using irradiated primers (Table 3). The highest-proton-dose sample tubes were visibly yellowed, and due to activation, the sample was maintained at room temperature over 24 h. Tests in which these reagents were used uniformly failed to PCR. To confirm that this outcome was not simply due to storage at room temperature, we stored equivalent reagents at room temperature for 48 h before qPCR and found that all samples with template amplified successfully and all no-template controls failed to amplify (Supplementary Fig. S4).
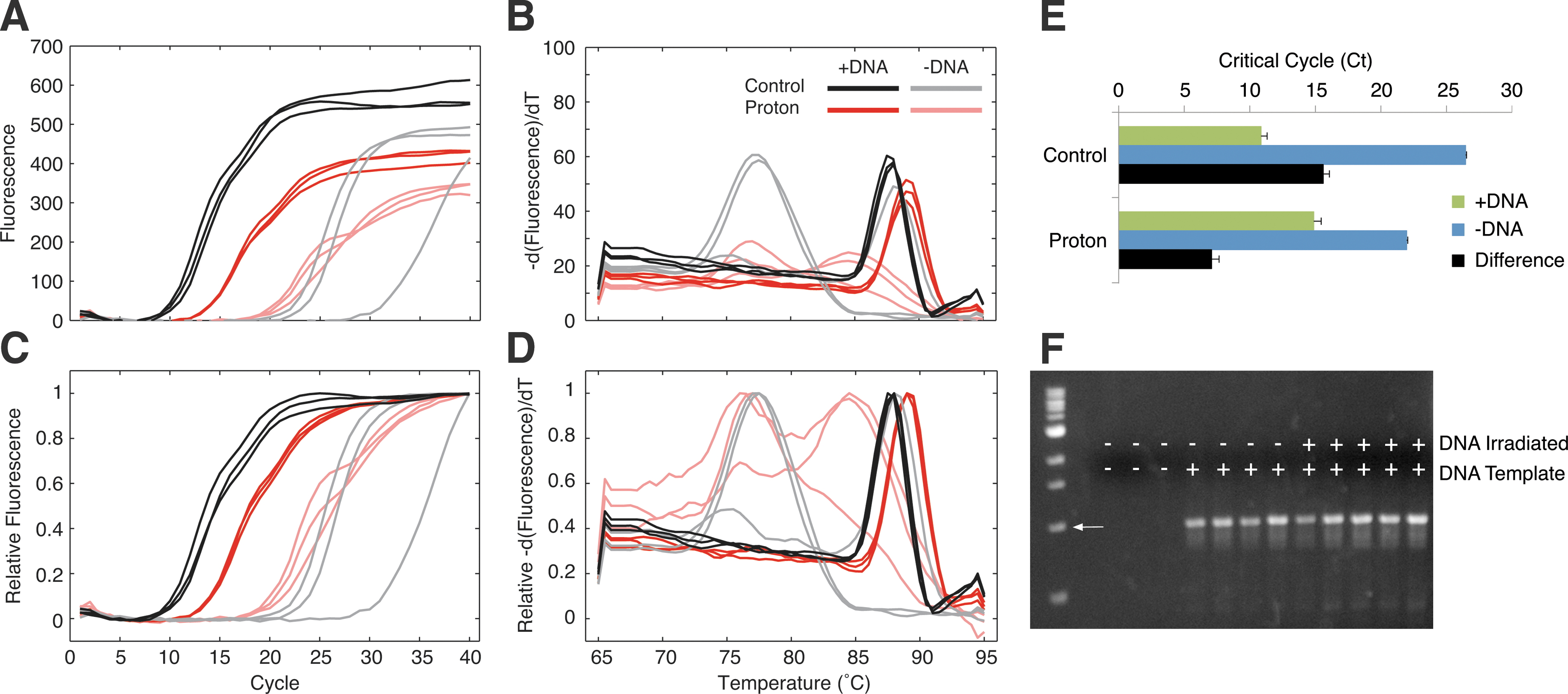
Successful PCRs after monoenergetic proton irradiation. (
3.3. Proton and heavy ion irradiation at NSRL
There was little impact of heavy ion irradiation on PCR (Fig. 3). Although there was variability in the qPCR end-point fluorescence (Fig. 3A, top row), after normalizing by end-point fluorescence, the qPCR curves were similar across all ions and doses (Fig. 3A, bottom row). The melt curves revealed a single melting domain in samples with template, and primer-dimer in no-template controls (Fig. 3B). In contrast, for the iQ SYBR Premix we found lower variation in end-point fluorescence (Fig. 3C), suggesting that end-point fluorescence differences result from variation in concentrations associated with pipetting rather than any impact of irradiation.
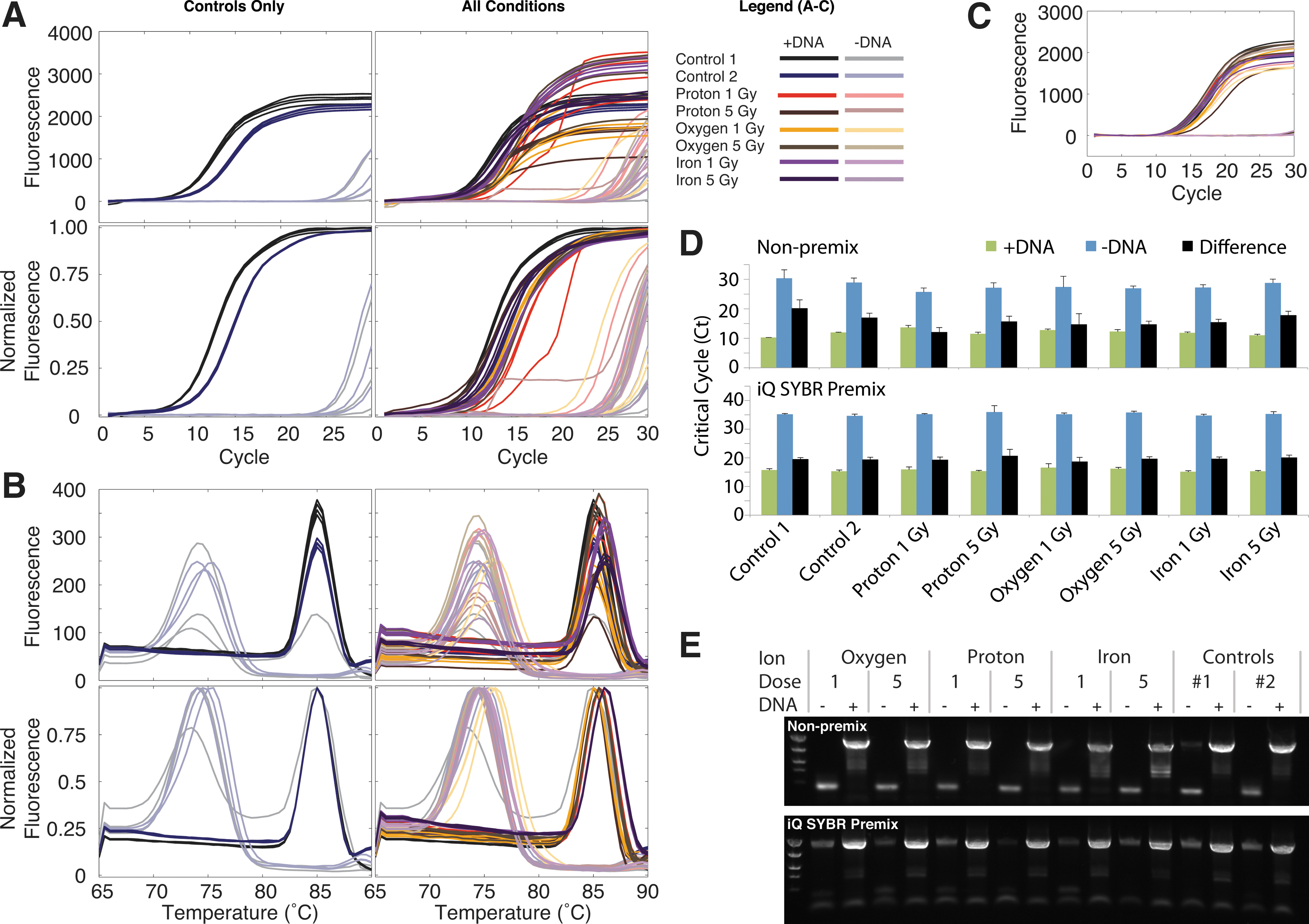
PCR is robust to heavy ion irradiation. (
To identify the effects of irradiation on the exponential phase of PCR, we compared the critical cycle numbers of our non-premix qPCRs. For the non-premix PCRs, samples with template had critical cycle numbers 12–20 cycles lower than no-template controls (Fig. 3D); and a regression of ion, dose, and presence of template on critical cycle number revealed no effect of ion or dose (p>0.5), with only the constant term and presence of template having significance (p<0.001). Limiting the regression to samples with template reveals minor delays in critical cycle associated with irradiation (95% CI: 1.4–3.3, 1.4–3.2, and 0.3–2.1 cycles for proton, oxygen, and iron ions, respectively, with p values of <0.001, 0.012, and <0.001). However, in this regression, higher dose is associated with a lower critical cycle number (95% CI: −0.44 to −0.13, p<0.001), an improbable result that casts doubt on the significance of any ion or dose effect. A regression limited to no-template controls reveals a trend toward increased (2–4 cycles earlier) primer-dimer formation after irradiation (95% CI: −6.6 to −1.0, −5.9 to −0.3, and −5.1 to 0.5 cycles for proton, oxygen, and iron ions, respectively, with p values of <0.01, 0.10, and 0.03). For the iQ SYBR Premix, we found a similar lack of variation across ions and doses; the only significant coefficient other than the constant term or presence of template was found in a regression limited to samples with template, where oxygen ions were associated with a one-cycle delay (95% CI: 0.1–2.1 cycles, p=0.03). The small magnitude of this effect and the lack of any dose effect suggest no impact due to irradiation.
The expected products (in samples with template) and primer-dimer (in no-template controls) were then confirmed via gel electrophoresis (Fig. 3E). In comparison to the non-premix samples, the iQ SYBR Premix samples had a lower tendency to form primer-dimer and by 30 cycles had amplified some background contamination.
To evaluate the effect of lyophilization during proton and heavy-ion irradiation, we performed 30-cycle qPCR with the EcoDry Premix with added SYBR Green I and observed consistent amplification with some variability in end-point fluorescence (Fig. 4). This end-point variability is likely associated with pipetting of SYBR Green I, because this level of variability was not observed for the iQ SYBR Premix, which includes SYBR Green I. Regression analysis revealed no effect of ion or dose (all p>0.2). In contrast, the constant term, presence of template, and lyophilization were significant (all p values<10−8). Hydration before irradiation was associated with a 0.9 cycle delay (95% CI: 0.7–1.1 cycles), indicating a possible, if tiny, benefit of lyophilization at the tested doses. However, this difference was also observed in the control conditions and may thus represent a small systematic bias instead of any benefit of lyophilization during irradiation. We also performed qPCR analysis with lyophilized individual reagents (buffer, primers, dNTPs, DNA, but not SYBR Green I) in a non-premix and identified no observable differences between lyophilized and hydrated reagents with a broad (15-cycle) difference in critical cycle numbers between samples with template and no-template controls (data not shown).
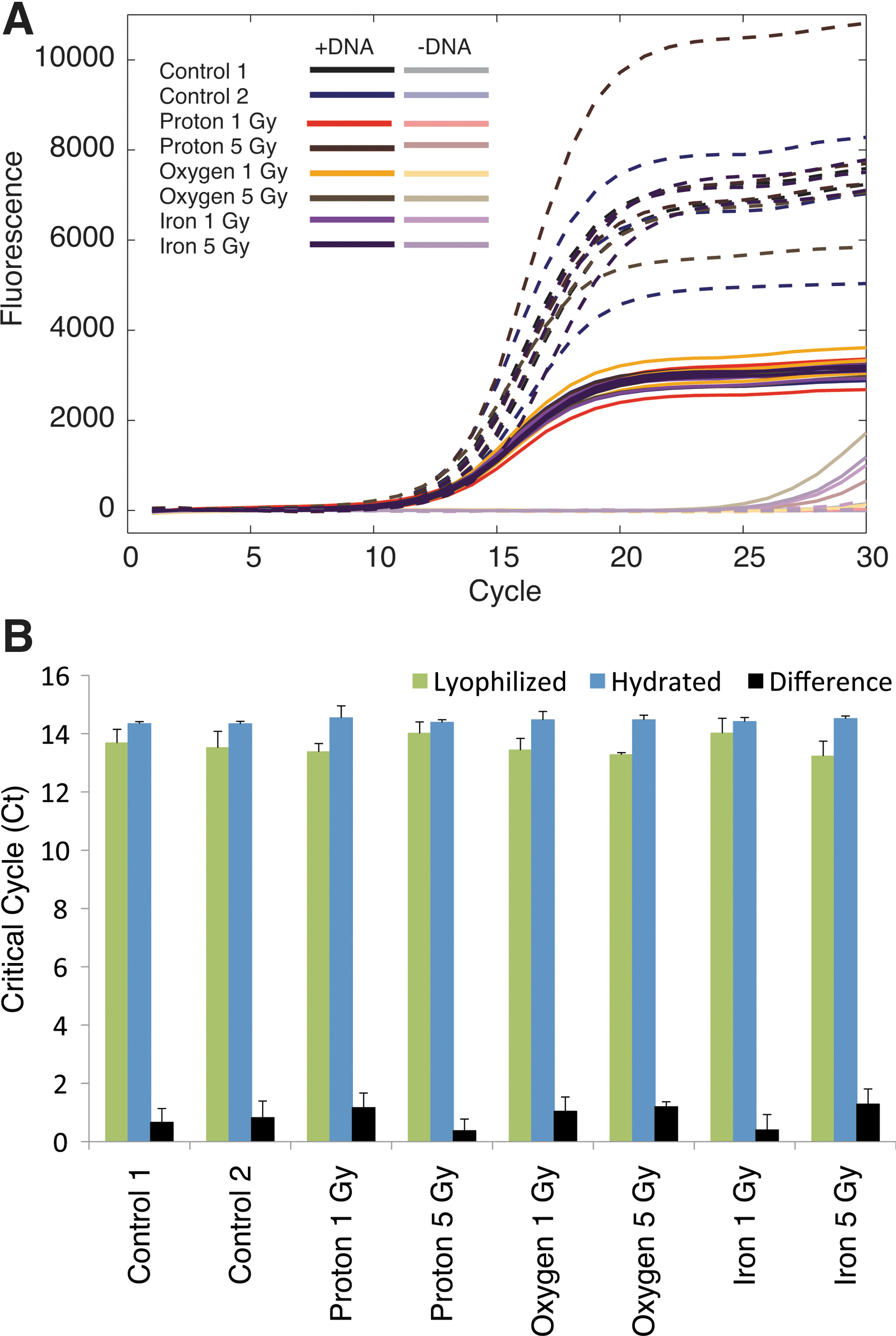
Effects of lyophilization during proton and heavy ion irradiation. (
4. Discussion
4.1. Assessment of irradiation results
We demonstrate that PCR reagents are robust to neutron, proton, and selected heavy ion components of space radiation at a dose commensurate with a 2-year Mars mission.
We first validated californium as a source for future astrobiological studies of neutron bombardment. Of note, the most probable energy of 0.7 MeV closely matches the predicted 0.6 MeV peak on the surface of Mars (Le Postollec et al., 2009a, 2009b). Californium also provides a gamma dose relevant to the radiation environment on Mars (de Diego-Castilla et al., 2011). Our mass spectrometer analysis of irradiated SYBR Green I revealed that variability of end-point fluorescence was not necessarily an indicator of reduced dye performance. In later exposures, we continued to observe variability in end-point fluorescence despite careful control over the SYBR Green I concentration. Because we observed less variability in premix samples, this variability is likely to arise from relative concentration differences in the master mix associated with pipetting, leading to exhaustion of a limiting reagent at different concentrations of the final dsDNA product. Because we saw essentially no impact due to irradiation when using californium, we can conclude that neither the neutron nor the associated gamma dose impaired performance of our reagents.
Overall, we found little impact of the ions and doses tested except for the two highest-dose proton conditions. At the 180 Gy exposure with SPE energy levels, DNA (primers and gDNA) were inactivated by proton irradiation; this inactivation is unlikely to result from dsDNA breaks, as these can be expected to occur with a frequency of one per 10 Gy per 5×109 bases (Cox and Battista, 2005), or 0.02 per E. coli genome (∼5×106 bp). However, single-stranded breaks occur with greater frequency, and the most likely source of inactivation may be nucleobase modification. Baqué et al. (2011) found that 2 MeV proton irradiation did not affect the function of DNA aptamers at fluences up to 1011 cm−2. In contrast, we found inactivation of primers and gDNA at this fluence. This difference may result from the >>2 MeV energy protons used in our SPE simulation but may also result from increased sensitivity of the PCR assay relative to the aptamer assay. While PCR can be inhibited by a defect anywhere along the target region, local damage (such as base modifications or other lesions) does not necessarily disrupt the shape-recognition function of aptamers. Performing the aptamer assay with higher-energy protons or the PCR assay with 2 MeV protons should reveal the source of this difference.
Although we did not find any protective effect of lyophilization under the conditions tested, such stable storage will be beneficial to facilitate tolerance to temperature fluctuations, which will reduce mass, volume, and power requirements associated with reagent storage. Notably, de Diego-Castilla et al. (2011) found lyophilization or vacuum drying allowed fluorescence-conjugated antibodies to tolerate repeated sudden temperature shifts. In this study, we only tested lyophilization under low dose (1 and 5 Gy) proton and heavy ion exposure. It is likely that lyophilization would be protective at higher doses, where measurable decrements in PCR performance occur.
Our fluxes were much higher than would be expected during a Mars mission; however, we expect that any dose-rate effect is likely minimal due to the lack of damage sensing, damage repair, and the lack of heating except in the most extreme exposure. Because we demonstrated successful PCR when using non-irradiated reagents after 48 h of room-temperature storage (Supplementary Fig. 4), the complete inactivation of reagents at the highest proton dose is due to proton irradiation, not temperature. In this case, the complete failure of PCR at this dose places limits on the ability of life as we know it to survive during meteoritic exchange. In particular, under minimal shielding, typical SPE exposure, and no active repair, the high dose condition (and complete PCR failure) would be achieved within 23 thousand years, limiting the timescale of interplanetary transfer within ejecta particles too small to provide radiation shielding.
4.2. Implications for in situ life detection
In situ sequencing provides a sensitivity and specific approach to life detection of nucleic acid–based life. While challenging, extracting nucleic acids from Mars-like soils can likely be addressed (Lui et al., 2011). While we focus here on the sequencing aspect, one promising approach with high contaminant rejection is synchronous coefficient of drag alteration (SCODA), in which rotating electric fields concentrate nucleic acids with extreme contaminant rejection (Broemeling et al., 2008; Pel et al., 2009).
Given this, how can we sequence in situ? Aside from single-molecule sequencing approaches (Eid et al., 2009; Pushkarev et al., 2009), which are impractical for our application due to their complexity, sequencing requires generating thousands to a million identical copies of the same DNA molecule. Of all commercially available sequencing technologies, semiconductor-based sequencing (Rothberg et al., 2011) as commercialized by Life Technologies (Personal Genome Machine by Ion Torrent) enables concurrent sequencing in millions of wells; requires no imaging or optics; and is extremely small, fast, and robust. This approach requires PCR for clonal amplification on beads, followed by sequencing by synthesis with standard dNTPs and polymerase on a ∼3×3 cm semiconductor chip. As dNTPs are flowed over the millions of wells, detection occurs in each well by sensing the hydrogen ions released during nucleotide incorporation. Our results demonstrate that the principal reagents required for sequencing in situ can survive the space radiation expected during a 2-year Mars mission.
Reducing the extent of amplification required during sequencing is desirable for several reasons: First, amplification introduces bias in terms of the relative number of different sequences. Second, amplification can result in spurious results due to contamination or nonspecific amplification. Lesions caused by irradiation may increase the likelihood of nonspecific amplification. Ion Torrent has been scaling their technology toward smaller wells, which will require fewer clonal copies per bead, and this may reduce the required number of PCR cycles. A second possibility is an amplification-free sequencing technology such as nanopore strand sequencing (Oxford Nanopore), in which a dsDNA molecule is unzipped and passes in ssDNA form through a nanopore, changing the ionic current through the pore. Signal processing of current traces can be used to determine the base sequence, although the stated error rate is currently 4%. This technology is not yet commercially available but could permit strand sequencing without amplification, including for non-DNA polymers such as RNA. However, even the nanopore approach currently relies on biological components such as an alpha-hemolysin pore. Our study shows that at least one protein, Taq polymerase, is robust under the conditions studied, suggesting that other proteins like alpha-hemolysin may be adequately robust to be used in space-based life-detection applications. Beyond sequencing in situ, planned life-detection instruments contain biological components (Parro et al., 2011). In addition, NASA has recently begun to explore space applications of synthetic biology; a critical prerequisite is that synthetic biology systems and the biological components of currently envisioned instruments must function despite space radiation.
Our results further support and expand the feasibility of using biological compounds in space, particularly for missions of short duration or in environments without large accumulations of charged particles. This includes our particular target of a 2-year Mars mission as well as other destinations under consideration for human exploration, such as lunar and asteroid missions. Recent discoveries of nucleic acids or their precursors within meteorites (Engel and Macko, 1997; Martins et al., 2008; Schmitt-Kopplin et al., 2010; Callahan et al., 2011; Cooper et al., 2011) and in interstellar space (Hollis et al., 2000) could steer the development of life toward these biomolecules. Thus, it makes sense to search for RNA- or DNA-based life within potential habitable zones even outside the context of meteoritic exchange, such as the probable liquid water oceans beneath Europa (Carr et al., 1998) and Enceladus (McKay et al., 2008; Postberg et al., 2011) and possibly Titan (Baland et al., 2011). These environments may present more of a challenge: Our high-dose proton conditions correspond to 1 day and 2 years, respectively, near Europa, suggesting that any mission there would require more robust reagents and/or significant shielding during any above-surface exposure. Our findings provide further evidence that meteoritic transfer of viable microbes to or from the jovian system is unlikely, given the small number of ejecta and the restriction that high dose rates place upon the residence time of such ejecta.
Footnotes
Acknowledgments
We thank Thomas Bork and William McCarthy for assistance with exposures at the MIT NRL, and Peter Guida, Adam Rusek, Michael Sivertz, and the rest of the NSRL-associated staff for assistance with heavy ion exposures. This work was funded by NASA's ASTID program (NNX08AX15G).
Author Disclosure Statement
No competing financial interests exist.
Abbreviations
dNTPs, deoxyribonucleotide triphosphate monomers; dsDNA, double-stranded DNA; GCR, galactic cosmic radiation; gDNA, genomic DNA; HZE, heavy ions; MGH, Massachusetts General Hospital; m/z, mass to charge; NRL, Nuclear Radiation Laboratory; NSRL, NASA Space Radiation Laboratory; PCR, polymerase chain reaction; qPCR, quantitative (real-time) PCR; SPE, solar particle event; ssDNA, single-stranded DNA.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.