
Abstract
The Tanpopo mission will address fundamental questions on the origin of terrestrial life. The main goal is to test the panspermia hypothesis. Panspermia is a long-standing hypothesis suggesting the interplanetary transport of microbes. Another goal is to test the possible origin of organic compounds carried from space by micrometeorites before the terrestrial origin of life. To investigate the panspermia hypothesis and the possible space origin of organic compounds, we performed space experiments at the Exposed Facility (EF) of the Japanese Experiment Module (JEM) of the International Space Station (ISS). The mission was named Tanpopo, which in Japanese means dandelion. We capture any orbiting microparticles, such as micrometeorites, space debris, and terrestrial particles carrying microbes as bioaerosols, by using blocks of silica aerogel. We also test the survival of microbial species and organic compounds in the space environment for up to 3 years. The goal of this review is to introduce an overview of the Tanpopo mission with particular emphasis on the investigation of the interplanetary transfer of microbes. The Exposed Experiment Handrail Attachment Mechanism with aluminum Capture Panels (CPs) and Exposure Panels (EPs) was exposed on the EF-JEM on May 26, 2015. The first CPs and EPs will be returned to the ground in mid-2016. Possible escape of terrestrial microbes from Earth to space will be evaluated by investigating the upper limit of terrestrial microbes by the capture experiment. Possible mechanisms for transfer of microbes over the stratosphere and an investigation of the effect of the microbial cell-aggregate size on survivability in space will also be discussed. Key Words: Panspermia—Astrobiology—Low-Earth orbit. Astrobiology 16, 363–376.
1. Introduction
1.1. Investigation of the panspermia hypothesis and chemical evolution in space
T
Another line of investigation has been carried out to test the presence of terrestrial microbes at high altitude using aircrafts, balloons, and a meteorological rocket (reviewed in Yang et al., 2009a). Terrestrial microbes were isolated from air samples collected in the troposphere and stratosphere. If microbes exist at upper altitudes, it would suggest that microbes might be transferred from Earth to other planets.
Before the origin of life on early Earth, organic compounds were formed in a process called chemical evolution and accumulated on primitive Earth. One of the major sources of organic compounds is that produced in space. Production of organic compounds from such molecules as carbon monoxide, methanol, and ammonia in ice mantles on interstellar dust in molecular clouds by cosmic rays or UV irradiation was examined, and the formation of amino acid precursors has been reported (Kasamatsu et al., 1997; Bernstein et al., 2002; Caro et al., 2002). Complex organic compounds including amino acid precursors were found in carbonaceous chondrites and meteorites. These organics carried by cosmic dust could have been transferred to early Earth (Kvenvolden et al., 1970; Chyba and Sagan, 1992; Cronin and Pizzarello, 1997; Elsila et al., 2009).
1.2. Astrobiology Tanpopo mission
To investigate the panspermia hypothesis and possible space origin of organic compounds, we conducted exposure and capture experiments at the Exposed Facility (EF) of the Japanese Experiment Module (JEM) KIBO of the International Space Station (ISS), which is orbiting at an altitude of about 400 km (Fig. 1a and Yamagishi et al., 2009). The mission is called Tanpopo, which means dandelion in Japanese. Seeds with fluff of the dandelion are spread by winds. We named our project after this process, which resembles panspermia. In the capture experiments, we capture microparticles with ultralow-density silica aerogels (Tabata et al., 2011, 2014, 2015). Blocks of silica aerogel are placed in aluminum Capture Panels (CPs) (Fig. 2a) (Tabata et al., 2015). They are then attached to the surface of the Exposed Experiment Handrail Attachment Mechanism (ExHAM) shown in Fig. 1b. In the exposure experiments, we exposed dried microbes and organic compounds in Exposure Panels (EPs) (Fig. 2b). EPs are also attached to the surface of the ExHAM.
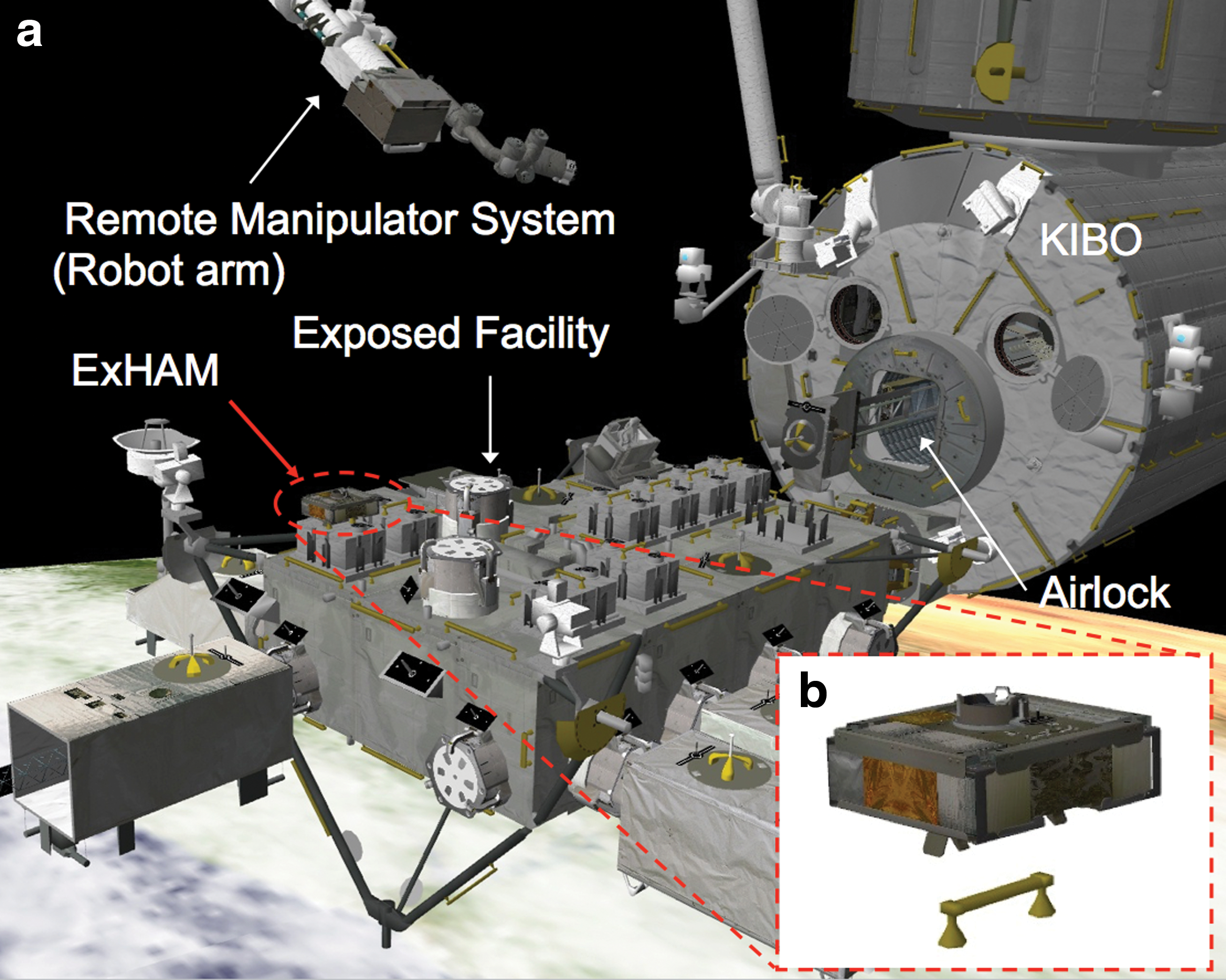
(

(
Capture Panels and Exposure Panels were carried to the ISS by the space carrier Space-X CRS-6 in April 2015. Scott Kelly, a NASA astronaut, attached CPs and EPs to the ExHAM on May 26, 2015. One set of CPs, consisting of about 12 blocks of aerogel, and three EPs were attached on the ExHAM for the first year of operation. The ExHAM was exposed in the depressurized air lock for about 2 weeks. The ExHAM was then moved to the EF-JEM and attached to a handhold on the EF of the JEM-ISS by robotic arms as shown in Fig. 1. After about 1 year of exposure, the ExHAM will be transferred to the pressurized area through the air lock. A set of CPs will be removed from the ExHAM and stored in the pressurized area, and a new set of CPs will be attached to the ExHAM for the next exposure operation. One of the three EPs used for the first-year exposure experiment will be removed from the ExHAM for return to the ground, while two will be left on the ExHAM for the next two exposure operations. The removed CPs and EP will be transferred to the ground. A set of CPs will be replaced with a new set of CPs, and one of the two remaining EPs will be removed from the ExHAM at the end of Year 2. The last EP and the set of CPs attached at the end of Year 2 will be removed at the end of Year 3. The removed CPs and EP will be transferred to the ground each year. The first CPs and EPs will be returned to the ground in mid-2016.
1.3. Six subthemes
The Tanpopo mission consists of six subthemes, as follows: (1) Capture experiment of terrestrial microbes during ISS orbit. (2) Exposure experiment of microbes under the ISS environment. (3) Denaturation experiment of organic compounds in space. (4) Capture experiment of micrometeorites in space. (5) Evaluation of dust capture performance of ultralow-density aerogel. (6) Monitoring of space debris.
We are investigating Subthemes 1, 4, and 6 with CPs and Subthemes 2 and 3 with EPs. Evaluation of aerogel blocks in Subtheme 5 will be done by testing the capability of aerogel blocks in Subthemes 1, 4, and 6.
The panspermia hypothesis will be addressed in Subthemes 1 and 2. In Subtheme 1, we will capture terrestrial microparticles and evaluate whether the microparticles harbor terrestrial microbes. If terrestrial microbes are found, this result will extend the upper limit of the existence of terrestrial microbes. The exposure experiment of microbes is to test the survivability of microbes under space conditions (e.g., temperature changes, high vacuum, cosmic rays, and wide spectrum of UV light).
The possible space origin of organic compounds before the origin of terrestrial life will be addressed in Subthemes 3 and 4. Micrometeoroids will be captured in forms that are as intact as possible and will be analyzed. We will analyze the organic compounds as well as the mineralogical components of the captured particles. The denaturation process of organic compounds will be tested by exposing organic compounds, including glycine, isovaline, hydantoin, and 5-ethyl-5-methylhydantoin, in EPs. Complex amino acid precursors synthesized from a mixture of CO, NH3, and H2O by proton irradiation (Takano et al., 2004) will also be exposed in EPs. To test the denaturation process, these organic compounds will be exposed in the space environment on the EF-ISS. After returning to the ground, the organic compounds in EPs will be analyzed.
Ultralow-density silica aerogel specially designed for the Tanpopo mission will be evaluated in Subtheme 5 (Tabata et al., 2011, 2014). A silica aerogel is composed of dried amorphous porous SiO2. Projectiles orbiting the ISS with hypervelocity (on the order of km/s) are severely damaged upon impact (reviewed in Burchell et al., 2006). Low-density aerogel can reduce the damage caused by impact. Aerogels have been used in space experiments to capture artificial debris and interplanetary dust particles (Yano and McDonnell, 1994; Hörz et al., 2000; Kitazawa et al., 2000; Brownlee et al., 2006). Our silica aerogel block has two layers, one 0.01 g/cc is the lowest-density aerogel ever used in space. Another layer with 0.03 g/cc supports the inside lower-density aerogel from the outside. We have tested possible capture using particles containing microbial cells that were accelerated to 4.2 km/s by a two-stage light-gas gun. We have shown that microbial DNA can be detected in the aerogel after impact (Kawaguchi et al., 2014). The developed aerogel can be used for next-generation sample return missions in the Solar System (e.g., Enceladus). The function of the aerogel will be analyzed by evaluating the performance of aerogel in Subthemes 1, 4, and 6.
The amount of space debris smaller than 1 mm in diameter in low-Earth orbit (LEO) has increased (Higashide et al., 2012). To obtain the latest data on the distribution of space debris at LEO, we will capture and count the number of metal particles in the aerogel in Subtheme 6. We will also count the impact craters on the aluminum surface of CPs.
2. Investigation of the Upper Limit of Terrestrial Microbes
2.1. Capture experiment of microbes in the upper atmosphere
The existence of microbes in Earth's biosphere has been explored. Capture experiments of microbes have been performed at altitudes up to 77 km above Earth's surface by airplanes, observation balloons, and a meteorological rocket (Rogers and Meier, 1936; Greene et al., 1965; Soffen, 1965; Fulton, 1966; Imshenetsky et al., 1978; Narlikar et al., 2003; Wainwright et al., 2003; Griffin, 2004, 2008; Yang et al., 2008a, 2008b; Shivaji et al., 2009; Smith et al., 2010). Imshenetsky et al. (1978) reported that eight viable, culturable microbial cells (CFU) were isolated from air samples collected between 48 and 77 km in altitude. Accordingly, the highest recorded microbial sampling studies were carried out at altitudes between 48 and 77 km. The microorganisms isolated from high-altitude air samples (Table 1) include the spore-forming bacterium Bacillus spp. and close relatives such as Paenibacillus spp., spore-forming fungal Penicillium sp., and Aspergillus sp., and the non-spore-forming bacterium Deinococcus spp. (reported as Micrococci in the literature; however, because Deinococcus radiodurans was initially named Micrococcus radiodurans, the identification of this bacterium as a member of the Micrococci may have been a misidentification) (Table 1). Bacillus spp., including the new strains, were typically isolated at different locations with various environmental conditions on Earth (Table 1). The high-altitude isolated microbes showed resistance against γ rays and UV (Yang et al., 2008a; Smith et al., 2011). The isolated bacterial cells tended to form aggregates, which is advantageous for shielding the cells within the aggregates against intense UV (Yang et al., 2008a; Smith et al., 2011). The airborne microbes are viable, although they may be in a dormant state at high altitude.
These capture experiments of microbes at high altitude have been performed over land, sea, and costal regions (Table 1). The locations from which these high-altitude bacteria originated are difficult to identify. However, some species that are typically found in specific places on the ground or in the ocean were also found at high altitude. A strain of the bacterium Streptomyces maritimus isolated from the upper altitude over Hyderbad, India, is an organism normally found in marine sediments (Shivaji et al., 2009; Bernan et al., unpublished, 2000). Some bacteria and fungi (Macrosporium sp., Papulaspora sp., Acinetobacter radioresistens, and Bacillus endophyticus) isolated from the upper altitude over land areas are known to be plant-associated microbes (Rogers and Meier, 1936; Imshenetsky et al., 1978; Shivaji et al., 2009). Many other bacteria and fungi have commonly been isolated from soil or other common terrestrial environments other than the ocean. Based on these results, the majority of high-altitude bacteria and fungi appear to have originated from land areas. These airborne microbes are transported over land and sea areas, because similar species were frequently isolated at high altitude over both the sea and land.
The altitude dependence of microbial density based on the capture experiments of microbes (in colony-forming units; CFU) is shown in Fig. 3, which is modified from the figure of Yang et al. (2009a). Because some microbial-sampling experiments have been performed within certain height ranges, the horizontal lines are used to show the respective height ranges in Fig. 3 (Yang et al., 2009a). Many experimental factors differ among the different experiments, such as differences in the sampling methods and culture media used (Yang et al., 2009a). Therefore, the altitude dependence of CFU in Fig. 3 is a rough estimate, because the CFU data at different altitudes from independent investigations cannot be directly compared. The regression line suggests a correlation between microbial density and altitude (km), with the microbial density decreasing from the ground to higher altitudes. If we extrapolate the regression line to the highest altitude where any microbial density data is available, the microbial density at the ISS (ca. 400 km) can be estimated to be about 10−5 to 10−6 CFU/m3. Based on the size of exposed block of aerogel (56 cm2) and the orbiting velocity of the ISS of 8 km/s, the maximum amount of captured terrestrial microbes is estimated to be 1.4 × 103 to 1.4 × 104 CFU for one exposed aerogel block after a 1-year exposure. One of the main subjects in the Tanpopo mission is to test whether the regression line can be linearly extended to the altitude of the ISS.
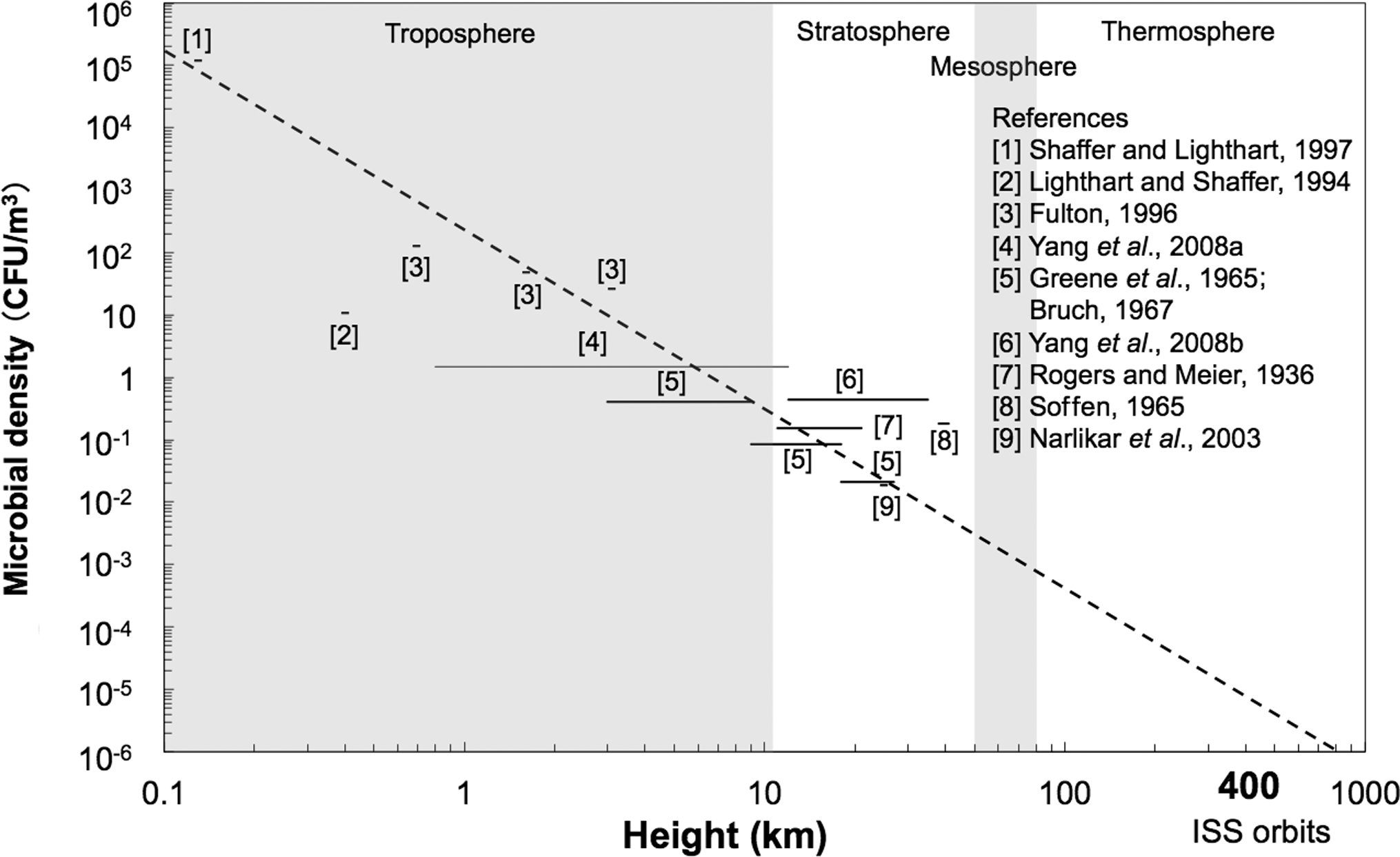
Microbial density depending on the altitude based on previous air-sampling experiments (modified from Yang et al., 2009a). Horizontal lines or filled circles were drawn based on the data at ambient temperature and pressure from each reference. Because some microbial sampling studies were carried out in certain altitude ranges, the horizontal lines show the respective sampling altitude ranges. The inserted dashed line indicates the estimated microbial density versus height dependence.
Bacteria, fungi, and viruses from seawater, soil, and vegetation are transported to the troposphere up to about 10 km by storms and strong upward winds worldwide (e.g., Smith et al., 2012). These particles are called bioaerosols. Bioaerosols are lifted by strong upward winds, monsoons (Randel et al., 2010), and hurricanes (Deleon-Rodriguez et al., 2013) and are transported by the prevailing westerly winds and trade winds (Hamilton and Lenton, 1998; Kellogg and Griffin, 2006; Griffin, 2007; Maki et al., 2008; Smith et al., 2012).
Recent studies have detected microbes as well as mineral particles in cloud precipitates. (Alfreider et al., 1996; Bauer et al., 2002; Amato et al., 2005, 2007; Segawa et al., 2005; Ahern et al., 2007; Bowers et al., 2009; Delort et al., 2010; Després et al., 2012). These bioaerosols in clouds contribute to precipitation as cloud condensation nuclei that form liquid cloud droplets or ice condensation nuclei that form ice particles (Schnell and Vali, 1976; Maki and Willoughby, 1978; Sun and Ariya, 2006; Deguillaume et al., 2008; Pratt et al., 2009; Delort et al., 2010; Smith et al., 2010; Després et al., 2012; Murray et al., 2012). Cloud condensation nuclei are the subset of particles that serve as seeds for cloud formation. Ice condensation nuclei are the particles that serve as seeds for ice crystals at the upper troposphere below −33°C. Therefore, bioaerosols have important roles in the climate of Earth.
The next question is, how are microbes or bioaerosols transported above the troposphere? Some hypotheses for the transportation mechanisms of microbes above the stratosphere are shown in Fig. 4. Human activities transport terrestrial microbes to the upper atmosphere via airplanes, observation balloons, meteorological rockets, rockets used for space exploration, and space shuttles (Bucker and Horneck, 1968; Griffin, 2004; Yang et al., 2009a; Smith, 2013). Natural phenomena can also push terrestrial microbes over the troposphere, as shown in Fig. 4. The triangle bar in Fig. 4 shows the frequency of natural phenomena. It has been suggested that bioaerosols are transported over the troposphere by thunderstorms (Dehel et al., 2008), volcanic eruptions (Griffin, 2004; Van Eaton et al., 2013), and meteorite impacts (Kring, 2000; Mileikowsky et al., 2000; Gladman et al., 2005; Worth et al., 2013). The upward wind can provide vertical lifting against gravity for sufficiently large (up to 3–5 μm) aerosol particles with densities up to 1.0–1.5 g/cm3 at stratospheric and mesospheric altitudes (Gryazin and Beresnev, 2011). Microparticles carrying microbes originating from volcanic eruptions might be transported to near-mesosphere altitude. Indeed, B. luciferensis, which was originally isolated from volcanic soils, was isolated from an altitude of 20 km (Griffin, 2008). In addition to these mechanisms, we hypothesize that cloud condensation nuclei and ice condensation nuclei of bioaerosols in clouds are accelerated by the electric forces of transient luminous events in the high atmosphere, and as a result, bioaerosols are transported over the stratosphere or mesosphere. Transient luminous events such as lightning-induced sprites (Franz et al., 1990; Boeck et al., 1995; Sentman et al., 1995; Lyons, 1996; Hardman et al., 2000; Pasko et al., 2002; Su et al., 2002; Takahashi et al., 2003), gigantic jets (Su et al., 2003; Cummer et al., 2009), and upwardly discharged blue jets (Wescott et al., 1995, 1996, 1998, 2001; Pasko et al., 2002; Mishin and Milikh, 2008) have been discovered in the region between thunderclouds and the ionosphere. However, whether microorganisms could survive such high electric charges is an open question. If the capture experiment at ISS orbit finds the same microbial species that are found at high altitude, this would suggest that terrestrial microbes are uplifted by the hypothesized mechanisms (Fig. 4). If we are able to identify the strains of captured microorganisms and their origins, we may be able to discuss which transporting mechanism is plausible.

Hypothetical mechanisms for transfer of microbes over the stratosphere. The red arrows indicate the possible altitudes to which the terrestrial microbes can reach by each respective mechanism. The image of the spacecraft is from NASA. (Color graphics available at
2.2. Investigation of the existence of terrestrial microbes at LEO in the Tanpopo mission
We are attempting to capture terrestrial microparticles orbiting or reaching ISS altitude. Our targets are microparticles or bioaerosols that consist of clay minerals and microbes. Our scientific goal is to obtain data to determine the microbial density at an altitude of approximately 400 km. In the pressurized area of the ISS, astronauts attached the CPs containing aerogel blocks to the three surfaces of the ExHAM: the surface toward the moving direction of the ISS, the port side of the ISS, and the space side (Fig. 1b). The ExHAM was transferred through the air lock and deployed to a handhold on the EF-JEM using the Japanese robotic arms (Fig. 1). After about 1 year of exposure of the CPs, the ExHAM will be transferred back inside the pressurized area using the robotic arms. Exposed CPs will be removed from the ExHAM and stored in the pressurized area. After CPs are returned to our ground laboratory, we will inspect aerogels for impact tracks and captured microparticles using a stereoscopic microscope. We expect that the terrestrial particles will collide with the CPs-aerogels mainly on the ISS forward and port-side surfaces. We will cut out impact tracks including the microparticles. The bioaerogel will be cut in half along the center lines of the tracks. First, microbes in tracks will be visualized by fluorescence microscopy after staining with a DNA-specific fluorescent dye. We have established a method to distinguish microbial DNA from inorganic compounds after staining the aerogel with the DNA-specific fluorescent dye SYBR Green I. The fluorescence from the dye bound to DNA decreases faster than that of the autofluorescence of inorganic particles (Kawaguchi et al., 2014). If microbial DNA is detected by the fluorescence method, we will amplify the microbial small subunit ribosomal RNA gene by PCR and compare the DNA sequences with the database of terrestrial microbial DNA sequences. If the captured microbial species from the ISS orbit are similar to the species captured at high altitudes over Earth (e.g., spore-formers), the mechanisms for transporting terrestrial microbes as bioaerosols from the ground can be extended to the ISS orbit altitude (Fig. 4).
A crucial aspect of our studies is how to prevent microbial contamination throughout the experimental procedure from preparation to analysis. In general, it is not possible to totally eliminate microbial contamination. However, we have already estimated the probability of microbial contamination. Contamination by the 16S rRNA gene in our handmade aerogels is less than the detection limit of the PCR assay (Tabata et al., 2011). We also confirmed that the aerogel does not inhibit PCR (Tabata et al., 2011). We handled aerogel blocks in the class 1000 experimental clean room on the ground, with its surface exposed to open air. It should be noted that the aerogel might be contaminated by floating microbes and might accidentally be touched by astronauts in the pressurized area of the ISS. Airborne microbial contamination may also occur during the transport from the ISS to the ground. Accordingly, we will cut off the area near the surface of aerogels, including shot traces, where the possibility of contamination is high. After cutting off the aerogel surface, we will use the inside portion of the tracks for fluorescence analysis and PCR. During the repressurization process of the air lock, tracks present on exposed aerogels may be contaminated by airborne microbes. We tested this possibility during the two-stage light-gas gun high-velocity impact experiment. The aerogel block was impacted by clay particles without microbes, in a vacuum, and repressurized. Microbial contamination in the tracks of the aerogel block was below the detection limit when observed by a fluorescence microscope (Kawaguchi et al., 2014).
3. Investigation of the Interplanetary Transfer of Microbes
3.1. The survival of microbes in space
Arrhenius proposed that a single spore could have been transferred to interplanetary space by radiation pressure from the Sun (Arrhenius, 1903). To investigate the possibility of microbial survival during interplanetary transfer in the space environment, exposure experiments with terrestrial microbes have been performed since the 1960s (reviewed by Horneck et al., 2010; Baranov et al., 2009; Olsson-Francis and Cockell 2010; de Vera et al., 2012; Rabbow et al., 2012, 2015). Various bacteria and fungi have been used for space exposure experiments in LEO using space stations (e.g., ISS, Mir) where solar UV is abundant. The results indicated that a monolayer of spores of Bacillus subtilis were killed by solar UV (Horneck, 1993), but multilayered Bacillus spores survived for 6 years in LEO (Horneck et al., 1994). A single spore or spores attached to micron-sized grains, as suggested by the theory of panspermia, will not survive in space (Horneck et al., 2001). Bacillus spores with clay and glucose as “mixed layers” survived for 21 days in LEO (Horneck et al., 2001). The rock-colonizing lichen Xanthoria elegans survived for 1.5 years in the ISS orbit (Onofri et al., 2012). The results suggested that small rocks that are sufficiently large to protect organisms against intense solar UV could serve as carriers, and as a result the lithopanspermia hypothesis was proposed (Horneck et al., 2002).
3.2. Interplanetary transfer of cell aggregates
Some microbes isolated from the upper atmosphere tend to form clumps or cell aggregates (Lighthart, 1997; Harris et al., 2002; Wainwright et al., 2004; Yang et al., 2008a). We have isolated Deinococcus aerius and Deinococcus aetherius, which are new species of Deinococcus, from air dust samples at the upper troposphere and lower stratosphere (Yang et al., 2008a, 2009b, 2010). Deinococcus aetherius forms cell aggregates during culture and exhibited higher resistance to UV than D. radiodurans and D. aerius (Yang et al., 2008a; Kawaguchi et al., 2013). The results suggest that the ability to form cell aggregates could be important for UV resistance at high altitude (Mancinelli and Klovstad, 2000; Horneck et al., 2001; Yang et al., 2008a; Smith et al., 2011). Several species of bacteria and fungi form cell aggregates on the surfaces of rocks, plants, and so on. These natural clumps of cells are known as biofilms. Bacterial biofilms are surface-associated, multicellular, complex microbial communities (O'Toole et al., 2000; Stoodley et al., 2002). Our previous report indicated that in aggregates, deinococcal cells near the surface layer were killed by UV radiation, and the layers of killed cells protected the cells underneath from UV damage (Kawaguchi et al., 2013). Deinococcus radiodurans, D. aerius, and D. aetherius are estimated to survive after 1 year of exposure under vacuum, temperature changes, heavy ions, and γ rays (Kawaguchi et al., 2013). Submillimeter deinococcal aggregates shield internal cells against the intense solar UV, and such cells survive the LEO environment for 1 year. We would like to emphasize the importance of cell aggregates as the ark for interplanetary transfer of microbes. We named this concept massapanspermia (Kawaguchi et al., 2013). Massa means mass in Latin. The European missions in Biofilm Organisms Surfing Space (BOSS) are also investigating the survivability of planktonic biofilms in space. The exposure experiment in space in EXPOSE-R2 has been in progress at the European Space Agency facilities in the ISS since August 2014. EXPOSE-R2 includes BOSS and Biology and Mars Experiment (BIOMEX) (de Vera et al., 2012). In BOSS, the following hypothesis will be tested: Biofilm lifestyle is better suited to support long-term survival under space and martian conditions than planktonic conditions (Baqué et al., 2013).
3.3. Exposure experiment of microbes in the Tanpopo mission
We are going to test the thickness dependence of survivability using dehydrated cells of D. radiodurans, D. aerius, and D. aetherius to evaluate the massapanspermia hypothesis. The dehydrated deinococcal cells will be exposed in EPs. Wells of 2.0 mm diameter on the aluminum sample plates are filled with different amounts of cells. Layers of cells with different thicknesses will be exposed: about 1 μm (corresponding to a single layer), 100, 500, 1000, and 1500 μm. Aluminum sample plates are placed in exposure units as shown in Fig. 5. The upper part of each aluminum plate is exposed to solar UV, and the bottom aluminum plate is placed in the dark. The upper aluminum plates are covered with MgF2 or quartz glass, which shields UV wavelengths less than 150 or 200 nm, respectively. We will compare the fractions of surviving cells to investigate the effects of exposure to different wavelengths of UV light. The exposure units are set in EPs. The EPs are attached to the ExHAM by astronauts in the ISS pressurized area. Three EPs will be exposed for different durations from 1 to 3 years. After the exposure experiment in the Tanpopo mission, EPs will be returned to the ground lab. The survival fraction of exposed cells will be evaluated by colony-formation assays (drop count method). By plotting the survival fractions following different exposure periods, we will extrapolate the survival curve to estimate the survival of microbes during long-term space transfer.
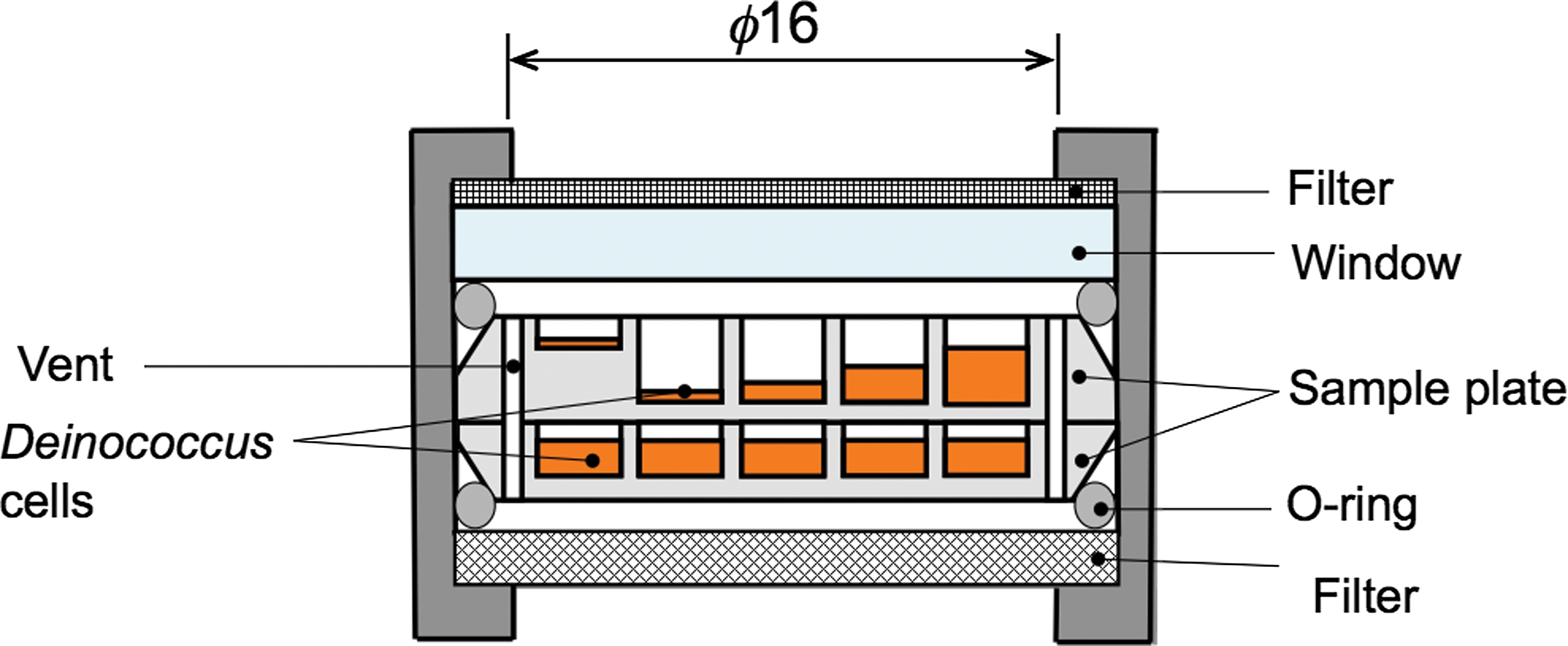
The cross section of an exposure unit. A metal mesh is placed at the top of the window to prevent scattering of accidentally broken windows. Wells of the upper sample plate were filled with deinococcal cells to different depths. The bottom sample plate is for the dark control samples. (Color graphics available at
We will also investigate DNA damage following exposure to the harsh space environment using D. radiodurans DNA repair–deficient mutant strains. Deinococcus radiodurans strain KH311 is defective in nonhomologous end joining (carrying a mutation in the pprA gene: Kitayama et al., 1983; Narumi et al., 2004). Deinococcus radiodurans strain rec30 is defective in homologous recombination (carrying a mutation in the recA gene: Moseley and Copland, 1975). Deinococcus radiodurans strain UVS78 is defective in nucleotide excision repair (carrying a mutation in the uvrA gene) and alternative excision repair (carrying a mutation in the uvdE gene: Moseley and Evans, 1983). After space exposure, the survivability will be investigated by measuring the survival fraction using colony-formation assays. To investigate DNA damage, including double and single strand breaks, pulsed-field gel electrophoresis and quantitative PCR will be conducted. We will also estimate the mutation frequencies of the rpoB gene. These experiments will demonstrate the importance of DNA repair systems in supporting life in the space environment.
In addition to Deinococcus spp., Nostoc sp. HK-01 and Schizosaccharomyces pombe JY3 are being exposed in the Tanpopo mission. Nostoc sp. HK-01 is expected to survive after 1 year of exposure based on ground-simulation experiments (Tomita-Yokotani et al., unpublished data). Although the resistance of S. pombe to solar UV is low, we expect S. pombe cells to survive in space in the dark (Hayashi et al., unpublished data).
4. Conclusion
To investigate the panspermia hypothesis and chemical evolution, the Tanpopo mission will perform experiments to capture terrestrial microparticles, micrometeorites, and space debris using aerogels and will examine effects of exposure of microbes and organic compounds to space conditions. CPs and EPs were carried to the ISS by a space carrier Space-X CRS-6 in April 2015. CPs and EPs were exposed on May 26, 2015. The capture experiment will test the possible presence of terrestrial, microbe-carrying microparticles that are orbiting or reaching altitudes of about 400 km. The capture experiments address the question of whether microbes can escape from the ground into LEO. If terrestrial microbes are found in the ISS orbit, it would suggest an increased possibility of terrestrial microbes reaching higher altitudes, and even other planets and moons (e.g., Earth's moon, Mars). We will test the microbial density-height dependence as shown in Fig. 3. If the captured microbial DNA sequences are found to be similar to those isolated at high altitudes within Earth's atmosphere, this would suggest that the microbes may be uplifted to the mesosphere by some mechanism such as human activities, transient luminous events (e.g., sprites and gigantic jets), and volcanoes (Fig. 4). Additional sampling to detect microbes at different altitudes of Earth's atmosphere and LEO using new methods (e.g., ultralong-duration ballooning and re-use rockets) will reveal details of microbial circulation around Earth. Overall, the control of contamination is most critical for the success of our capture experiments. We have already confirmed low contamination in the aerogels and developed procedures, protocols, and experimental environments with the lowest contamination possible. We will expose layers of dehydrated deinococcal cells with different thicknesses at the ISS for approximately 1, 2, and 3 years to investigate the survivability during interplanetary transfer and analyze the surviving fraction of exposed cells depending on the thickness. From these results, we can judge how cells in large aggregates may survive in the space environment. By plotting the survival fractions following different exposure periods, we will extrapolate the survival curve to estimate the survival of microbes during long-term space transfer. In addition, we will also investigate DNA damage in space. These experiments will demonstrate the importance of DNA repair systems in supporting life in the space environment.
Footnotes
Acknowledgments
We thank all Tanpopo working group members. We especially thank Prof. Kensei Kobayashi (Yokohama National University) and Dr. Yoko Kebukawa (Yokohama National University) for proofing the section describing the organic compounds experiment. We also gratefully thank Scott Kelly (NASA astronaut) and JAXA and NASA operators for exposing the Tanpopo samples on the ISS. We also thank Dr. Hiromi Shibata (The Institute of Scientific and Industrial Research, Osaka University) for his valuable comments on the electrostatic acceleration of transient luminous events. We are grateful for travel grant support from the Society for Promotion of Space Science to attend the international meeting.
Author Disclosure Statement
No competing financial interests exist.