
Abstract
Within the first billion years of Earth's history, the planet transformed from a hot, barren, and inhospitable landscape to an environment conducive to the emergence and persistence of life. This chapter will review the state of knowledge concerning early Earth's (Hadean/Eoarchean) geochemical environment, including the origin and composition of the planet's moon, crust, oceans, atmosphere, and organic content. It will also discuss abiotic geochemical cycling of the CHONPS elements and how these species could have been converted to biologically relevant building blocks, polymers, and chemical networks. Proposed environments for abiogenesis events are also described and evaluated. An understanding of the geochemical processes under which life may have emerged can better inform our assessment of the habitability of other worlds, the potential complexity that abiotic chemistry can achieve (which has implications for putative biosignatures), and the possibility for biochemistries that are vastly different from those on Earth.
Events following the formation of Earth around a young Sun (see Chapter 3) determined the chemical environments in which life may have originated. This chapter describes the current understanding of the geochemical environment and mechanisms behind the transition of nonliving materials to living matter (i.e., the origins of life or abiogenesis). The focus of this chapter is the period during which life likely originated: from the Hadean Eon (4.56–4.0 billion years ago [Ga]) up to the first era of the Archean Eon (4.0–2.5 Ga) known as the Eoarchean (∼4.0 to 3.6 Ga). The subsequent chapter in this series (Chapter 5) will review our understanding of the evolution of life throughout Earth's history. It is important to note that this chapter is focused on understanding mechanisms for how Earth-like life could have evolved on Hadean Earth and does not discuss in detail abiotic chemistry that could be relevant for the formation of alternative life-forms distinct from our own (i.e., “weird life” or “exotic life”); for a review on this topic the authors kindly direct readers to Chapter 9: Life as We Don't Know It.
Chapter 4.1 will review what we know regarding the geochemical composition of early Earth's crust (lithosphere), oceans (hydrosphere), and atmosphere. Next, Chapter 4.2 describes how the chemical interactions at the interface of each of these geospheres can generate energy, organic compounds, molecular polymers, enantiomeric excess, and chemical networks. In Chapter 4.3 we will discuss geochemical scenarios conducive to processes predicted to have been important for the emergence of living systems. Finally, the chapter will finish by describing the potential chemistry of the simplest system that we could feasibly describe as living (see Chapter 2 for a discussion on defining “life”) and how abiotic chemistry could transition to a functioning protocell (Chapter 4.4).
The Hadean Environment
Earth began forming from the dust and gas of the solar disc around the developing Sun 4.567 ± 0.001 billion years ago (Ga) and continued to accrete over a period of ∼107–108 years (see review by Connelly et al., 2017). Following the main phase of accretion, molten Earth began to differentiate, forming the core, mantle, and crustal layers by around 4.53 Ga (see Chapter 3.4.3.1). This is generally taken to mark the beginning of the geological eon known as the Hadean.
Formation of the Moon and the Late Heavy Bombardment
Oxygen and zinc isotopic values of Earth and lunar samples suggest that the Moon was formed within ∼60 million years of Earth's formation (i.e., at ∼4.5 Ga; Halliday, 2000; Bottke et al., 2015) due to the impact between a Mars-sized planetesimal, named Theia, and proto-Earth (Borg et al., 2022). The collision of Theia and other large impactors likely had immense consequences for the origins and/or pervasiveness of life, as they could have generated more habitable conditions at the impact site (Osinki et al., 2020) by delivering volatiles and organic compounds (see Chapter 3.3.4 and Chapter 4.1.3) and providing the energy needed to generate additional organic materials (Ferus et al., 2019; Takeuchi et al., 2020; Zellner et al., 2020). Alternatively, impactors could have destroyed any extant life (Maher and Stevenson, 1988) not present in the deep subsurface (Grimm and Marchi, 2018). Theia's impact could have generated sufficient energy to vaporize a significant portion of proto-Earth's crust and mantle, resulting in a magma ocean on both planetary bodies (Kasting and Catling, 2003; Sleep, 2010; Carter et al., 2020) and the boiling off of any liquid oceans, essentially sterilizing the planet if life was present (Zahnle and Sleep, 1997). Bombardment of early Earth remained intense 4.25–3.87 Ga (i.e., the Late Heavy Bombardment [LHB]) as evidenced by craters on the Moon, Mars, and Mercury (see Chapter 3.3.4).
The Moon has had a significant influence on the habitability of Earth. Without the Moon, Earth's axial tilt could oscillate and reach up to 85 degrees (Laskar et al., 1993), instead of the current 23.5 degrees. Thus, the Moon prevents extreme climate swings that would make it difficult for complex life to evolve (i.e., the Rare Earth hypothesis; Ward and Brownlee, 2000). Moreover, the Moon is also responsible for Earth's oceanic tides, and it effectively slows Earth's rotation through “tidal braking,” such that the length of one Earth day increases over time. During the Hadean it is estimated that an Earth day was only ∼12–18 hours long (Williams, 2000; Lathe, 2004). The shorter day-night cycles and cycling of oceanic tides (<8 h periodicity) would have promoted wet-dry cycling on any existing landmass, potentially facilitating the formation of biologically relevant molecules (see Chapter 4.2.4.3). As Earth's rotation slows, the Moon drifts farther away to maintain angular momentum; it is estimated that during the Hadean the Moon was much closer to Earth (ca. 191,130–320,500 km) than it is today (∼385,000 km) (Williams, 2000).
Hadean geology
After the Moon-forming impact, Hadean Earth was covered in a magma ocean (∼1000 km depth; Righter and Drake, 1999). Within around ∼10 million years (i.e., by ∼4.57 Ga), olivine and pyroxene minerals crystallized as the magma ocean cooled, ultimately forming ultramafic igneous rock, known as peridotite (Fig. 4.1; Elkins-Tanton, 2008). If this ultramafic crust was exposed to liquid water <400°C the crust would have oxidized and become hydrated via a process termed serpentinization which produces serpentinite rocks (i.e., hydrous (Mg, Fe)-phyllosilicates) and hydrogen gas from water (see Chapter 4.2.3.1). Otherwise, the dense peridotite crust would have eventually sunk, leaving the upper mantle relatively depleted in magnesium (Mg) and enriched in calcium (Ca) and aluminum (Al), which favor the crystallization of plagioclase minerals (Bourdon and Caro, 2007). Accumulation of plagioclase and pyroxene minerals is thought to have facilitated formation of basaltic rock (see Chapter 3.4.3) that could have formed a long-lasting mafic oceanic crust (Fig. 4.1).
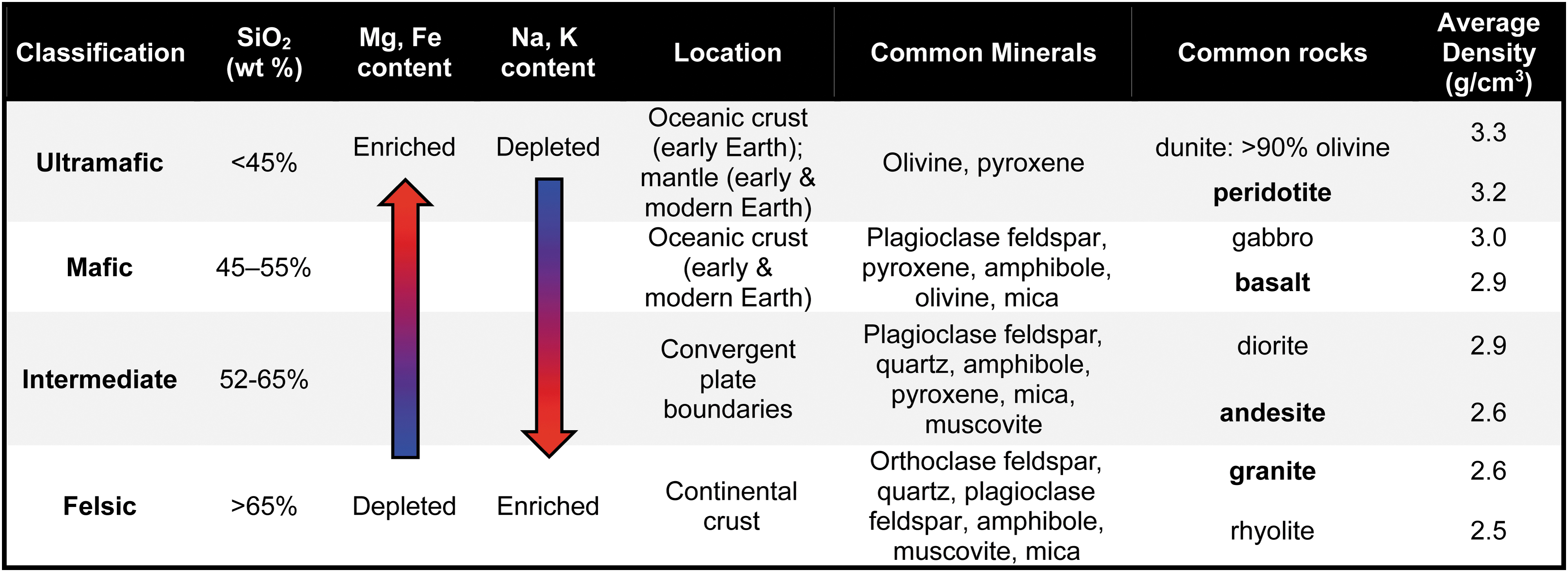
Classification and mineralogy of igneous rocks on Earth. Classes are determined by silica (SiO2) content. Rocks in bold text are those that are the dominant rock type of each class on modern Earth.
Whether continental crust (Fig. 4.1), was also present in the Hadean remains an open question (see Hawkesworth et al., 2020 and references therein). One of the main arguments against the existence of continental crust in the early Hadean is that it requires large volumes of water to facilitate the dissolution and melting of the crust (Collins et al., 2020). Water stabilizes the basaltic melt and delays the onset of recrystallization, thus enabling sufficient time for the lighter elements in the melt to partition, decreasing the upper melt's density, and facilitating its rise and physical separation from the lower (denser) melt. This less dense, felsic magma forms granitic rock that accumulates to form continents that can rise above sea level (Grove et al., 2012; Lee et al., 2015). Continental crust could not have been Earth's primary crust and likely occurred only after the existence of an ocean and the onset of plate tectonics (Tang et al., 2016; Greber et al., 2017). Notably, melting of mafic crust can occur in the absence of subduction (via melting of the underlayer of oceanic crust; Bédard, 2006), but this mechanism would not generate sufficient felsic melts given that rocks at these depths are generally depleted in water (Tang et al., 2016). Although it is uncertain when plate tectonics began, there is some evidence it was ∼3 Ga, and proposed dates range from 4.4 Ga to 700 million years ago (Ma; see reviews by Condie, 2018; Hawkesworth et al., 2020).
The hot early stages of Earth's formation and its accretion in a relatively water-depleted region of the Solar System suggest water and other volatiles were derived from exogenous sources, namely comets and asteroids, in a later stage of its formation known as the “late veneer” (see Chapter 3.3.4). Hydrogen isotopes in Solar System materials suggest that carbon-rich asteroids delivered the bulk of Earth's water (Albarède, 2009) as early as 4.2–4.4 Ga (according to oxygen isotopes in zircons; Borisova et al., 2022). For a brief overview of how zircons inform prebiotic studies, see Rodriguez (2023). As the planet cooled, water vapor in the atmosphere condensed and rained out, lowering temperatures that triggered further condensation and a growing ocean (Abe, 1993). By the Hadean–Archean transition, global oceans are predicted to have been more voluminous than their modern counterparts (Foley et al., 2003; Sleep, 2010).
There is still little that can be said with certainty regarding the geochemical conditions of the Hadean ocean other than that it was anoxic, enriched in ferrous iron and sulfides, and depleted in sulfate (see Table 4.1, Chapter 4.2.2.5). Hadean oceanic waters were likely temperate (0–50°C; Hren et al., 2009; Krissansen-Totton et al., 2018) and, due to the relatively high partial pressure of atmospheric CO2, mildly acidic (i.e., Sleep et al., 2001). The pH of the oceans, having been buffered by the equilibrium between carbonates and dissolved CO2, is predicted to have progressed from slightly acidic (pH ∼5.8) to near neutral (pH ∼6.8) by the end of the Hadean (Sleep et al., 2001; Krissansen-Totton et al., 2018). As crustal and mantle temperatures in the Hadean were higher than today, it is thought that dissolution of rocks would have been faster, resulting in ocean salinity potentially double or even 10 times modern levels (Knauth, 2005). However, the Hadean/Archean ocean salinity remains uncertain, and fluid inclusions from 3.5–3.0 Ga rocks suggest a salinity similar to modern oceans, except for K for which the Hadean oceans may have been relatively depleted (today, marine K is sourced from continental crust; Marty et al., 2018).
Predicted Geochemistry for Early Earth versus That of Modern-Day Earth
Predicted Geochemistry for Early Earth versus That of Modern-Day Earth
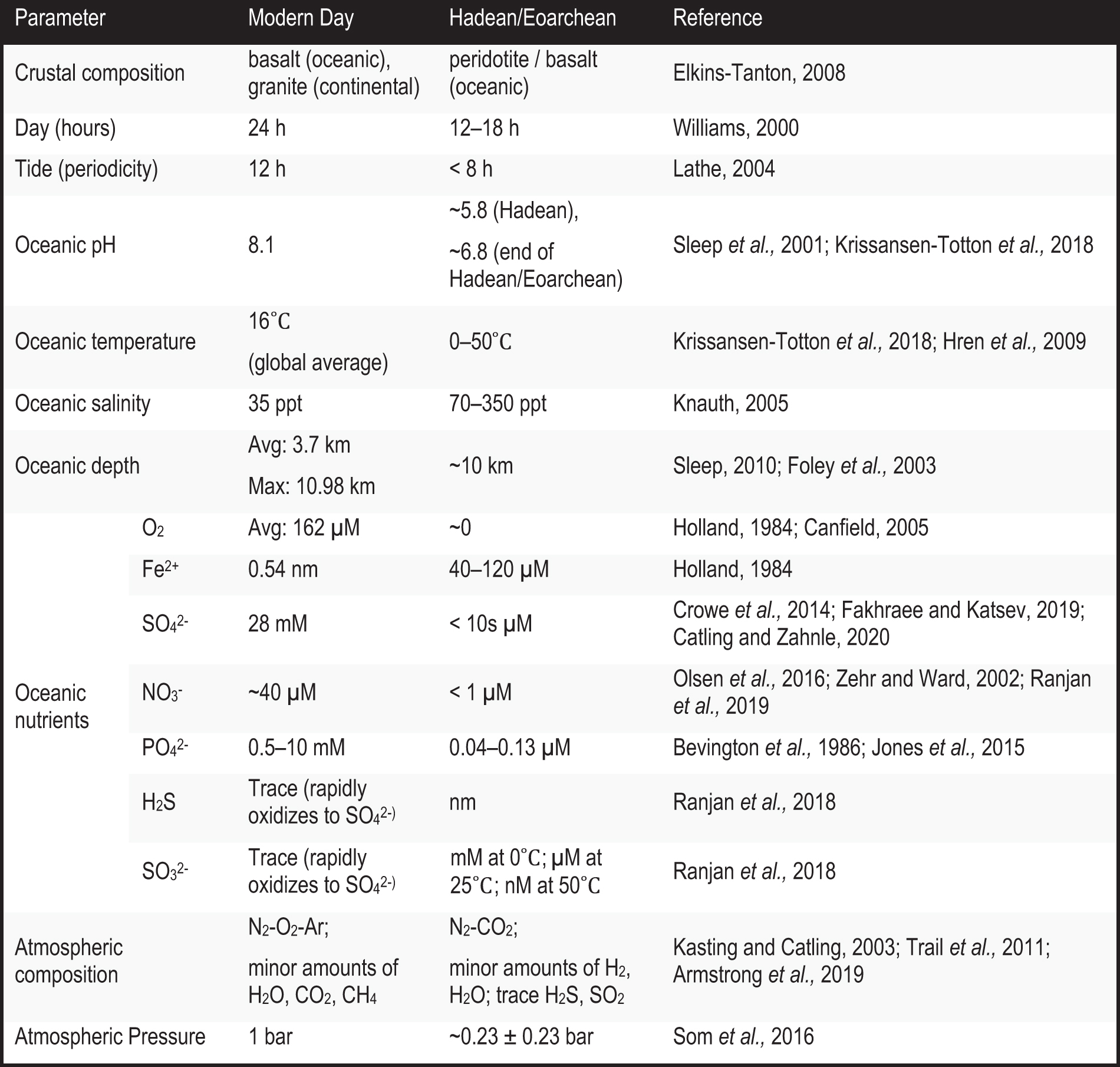
See Chapter 4.1 for details regarding the composition of Hadean Earth's crust, ocean, and atmosphere; see Chapter 4.2.2 for additional details regarding the abiotic cycling and chemical availability of CHONPS elements in the atmosphere and ocean.
Earth's primordial atmosphere was likely lost upon the impact of Theia to form the Moon, and its subsequent (i.e., secondary) atmosphere was likely formed via a combination of volcanic outgassing and exogenous delivery of volatiles during the Late Veneer (see Chapter 3.4.1.1). In addition, this atmosphere is estimated to have been less dense than today's, perhaps as low as ∼0.23 ± 0.23 bar (Som et al., 2016). The atmosphere was anoxic up until ∼2.1 Ga (Chapter 5.2) and is thought to have been composed mostly of nitrogen (N2) and carbon dioxide (CO2) with some H2, H2O (vapor), hydrogen sulfide (H2S), and sulfur dioxide (SO2) gases (Kasting and Catling, 2003; Armstrong et al., 2019). Reduced gases (e.g., CH4, CO, NH3) were likely only generated transiently from impact events or lightning discharges (Kress and McKay, 2004; Pizzarello et al., 2011; Zahnle et al., 2020).
In the Hadean, the Sun is thought to have only been 70% as luminous as it is now (Rosing et al., 2010). If the Hadean had the same greenhouse effect and albedo as Earth today, the planet would have been too cold for surface liquid to have existed, yet there is evidence of liquid water during this period. This apparent contradiction, called the Faint Young Sun Paradox (FYSP), can be resolved by considering the warming effect of greenhouse gases, such as CO2 (see Chapter 3.4.1.4). Alternative explanations for the FYSP include (i) reduced albedo of a less cloudy, landless early Earth (Rosing et al., 2010); (ii) N2-H2 atmospheric collisions that could have induced each molecule to behave as a greenhouse gas (Kasting et al., 2013; Wordsworth and Pierrehumbert, 2013); and (iii) the greenhouse gas effects of nitrous oxide (N2O), generated via oxidation of N2 during super-flare events from the young Sun (Airapetian et al., 2016). See Charnay et al. (2020) for a detailed review of the FYSP.
Life in the Hadean/Eoarchean
By the end of the Hadean, conditions may have been hospitable for life. An overview of major Hadean–Archean events and where both eons fall along the geologic timeline of Earth is given in Fig. 4.2. Table 4.1 summarizes the current understanding of the geochemistry of the Hadean–Eoarchean Earth.
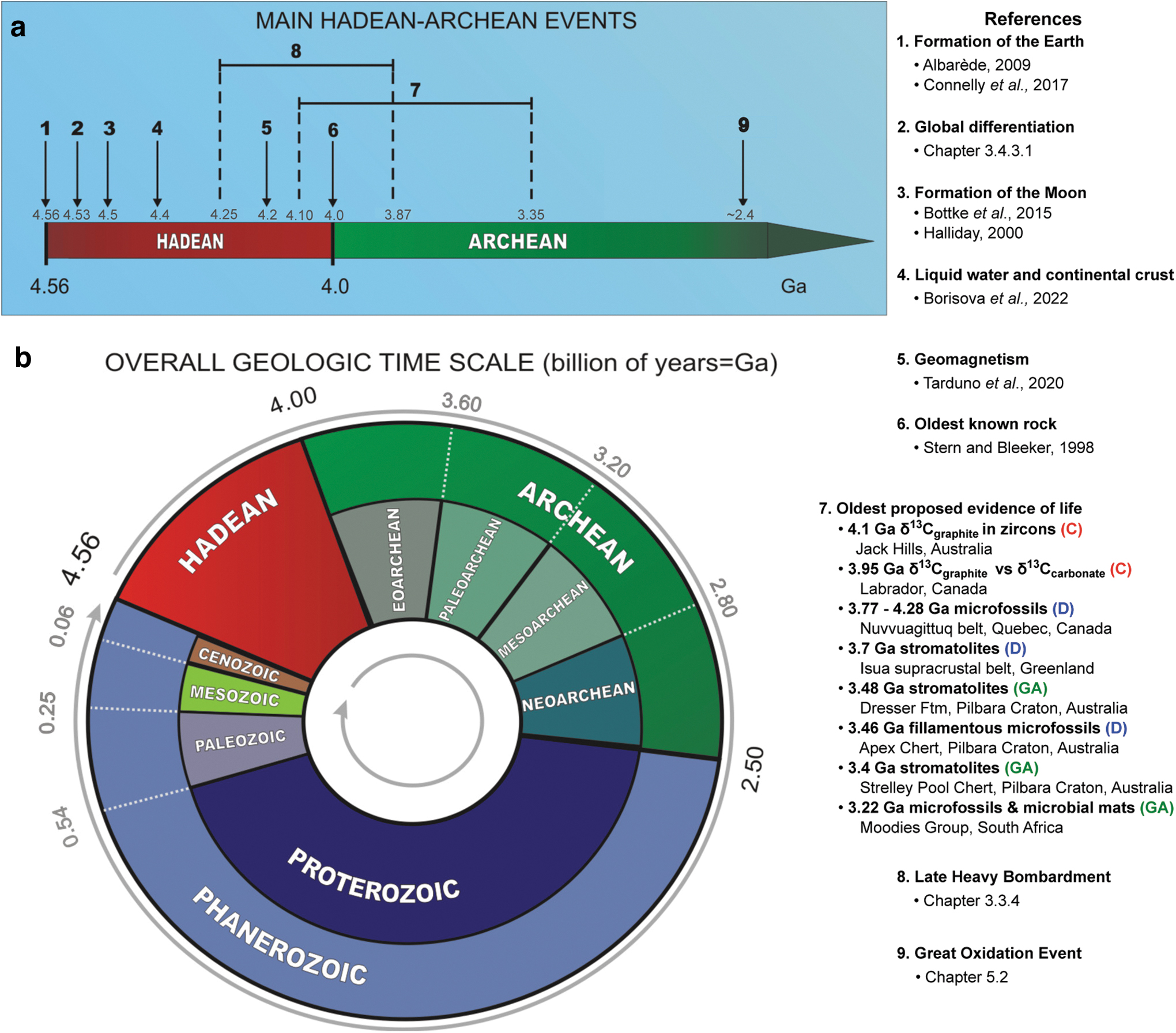
An overview of the geologic history of Earth. (
The oldest evidence for life on Earth stems from a combination of isotopic measurements consistent with metabolic activity and morphological structures, the most convincing being stromatolites. Stromatolites are layered sedimentary structures that formed from the entrapment or precipitation of sediment particles within extracellular polymeric substances secreted from photosynthetic organisms (Fig. 4.3; see Chapter 8.1.2). Although controversial, isotopic ratios and morphological evidence suggestive of biological activity have been reported in geological samples as old as 4.1 Ga (Bell et al., 2015; Dodd et al., 2017; Tashiro et al., 2017; Allwood et al., 2018; Javaux, 2019), and stromatolites from the Isua supracrustal belt in Greenland have been dated to ∼3.7 Ga (Allwood et al., 2018; Zawaski et al., 2020; Nutman et al., 2021). Nonetheless, there is a consensus that life was present by ∼3.5 Ga given the various Archean rocks with strong evidence for life (3.5–3.2 Ga), the most notable being stromatolites identified in the 3.48 Ga Dresser Formation (Australia) and the 3.35 Ga Strelley Pool Formation in Australia (Duda et al., 2016; Djokic et al., 2017; Homann et al., 2018; Baumgartner et al., 2019). The biogenicity of 3.22 Ga sedimentary structures from the Moodies Group Formation in South Africa is also widely accepted (Homann et al., 2018). For a detailed discussion on the oldest evidence for life on Earth, including ongoing debates and uncertainties, see Fig. 4.2 and reviews by Javaux (2019) and Lepot (2020).

A photo and elemental maps of an Archean stromatolite from Bolivia. The stromatolite was procured from Nature's Own mineral and rock shop. Elemental maps were acquired via X-ray fluorescent spectrometry (XRF) using a Bruker M4 Tornado micro-XRF. Each elemental map is on a relative scale (black = depleted; bright color = enriched in the element of interest). As can be seen in the elemental maps, the stromatolite columns are composed of Ca (as calcium carbonates) and P, whereas the interstitial sediments are enriched in Si and Al (from aluminosilicate sediments that infilled the space between columns post deposition). The distribution of Fe and S reflects cyclical fluctuations in the surrounding environment.
Investigations into the origins of life can be either top-down (biological) or bottom-up (chemical). The top-down approach “reverse engineers” life into its most basic constituents to consider the potential materials and reactions utilized by the first life-forms (Weiss et al., 2016; Djokic et al., 2017). Top-down approaches are hindered by the dearth of the Hadean geochemical record. A bottom-up approach considers the fundamental physical and chemical mechanisms of molecular systems in the context of primordial geochemical environments to deduce how simple molecular building blocks develop into more complex biomolecular systems (see Chapter 4.2.4). Experiments simulating early Earth chemistry together with meteorites and returned extraterrestrial samples suggest that early Earth hosted a myriad of organic molecules derived from exogenous (external) and endogenous (internal) sources (see Chapter 3.3 and Chapter 4.2.3).
Energy sources for the origins of life
Energy is a critical ingredient for abiogenesis as it enables the generation and maintenance of biological matter. The higher the amount of energy needed for a reaction to proceed (called the activation energy), the less likely it is to occur spontaneously (see Fig. 4.4). The activation energy can be lowered by a catalyst that in and of itself is not consumed, thereby increasing the rate of the reaction (see Chapter 4.2.6).
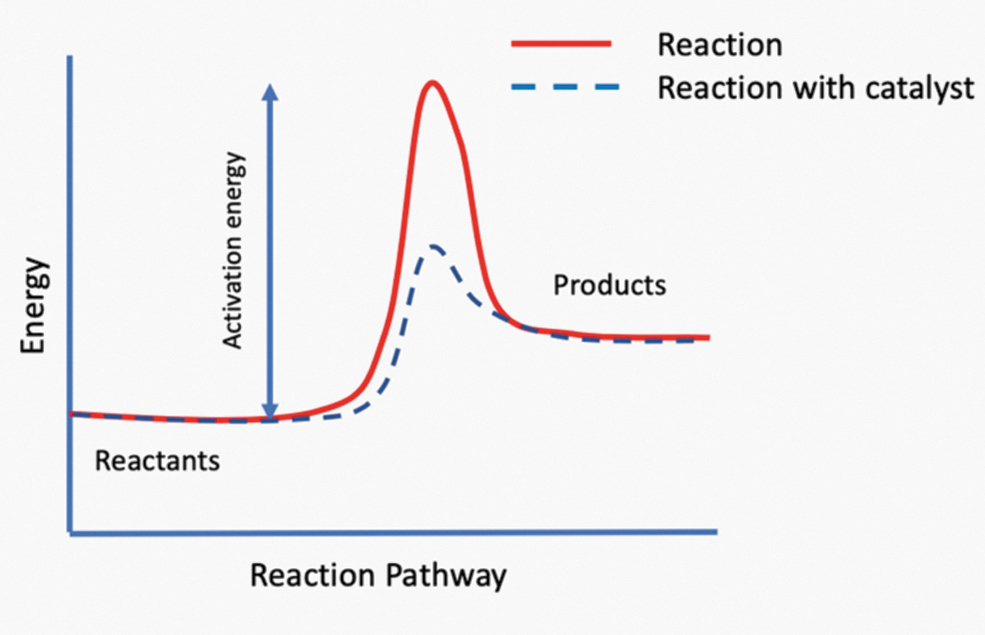
Activation energy of a generic reaction with (blue) and without (red) a catalyst.
Forms of energy are interchangeable (Fig. 4.5). Life takes advantage of this to precisely control when and which chemical reactions take place. The most prevalent example of this is when life harnesses the movement of protons to synthesize adenosine triphosphate (ATP), the primary energy molecule used by all terran life (see Chapter 6.2.1). However, it is unclear whether the ability to harness and store energy was necessary for abiogenesis or whether it emerged during life's evolution. The prebiotic synthesis of AMP (adenosine monophosphate), ADP (adenosine diphosphate), and ATP have been demonstrated, although whether sufficient quantities could have been generated to offset ATP's instability remains uncertain (Kolesnikov et al., 2008; Pinna et al., 2022). Alternative molecules have been proposed as potential precursors to ATP, including thioesters (de Duve, 1998; Goldford et al., 2017; Sanden et al., 2020), pyrophosphate (Lipmann, 1984; Holm and Baltscheffsky, 2011), and acetyl phosphate (Whicher et al., 2018).
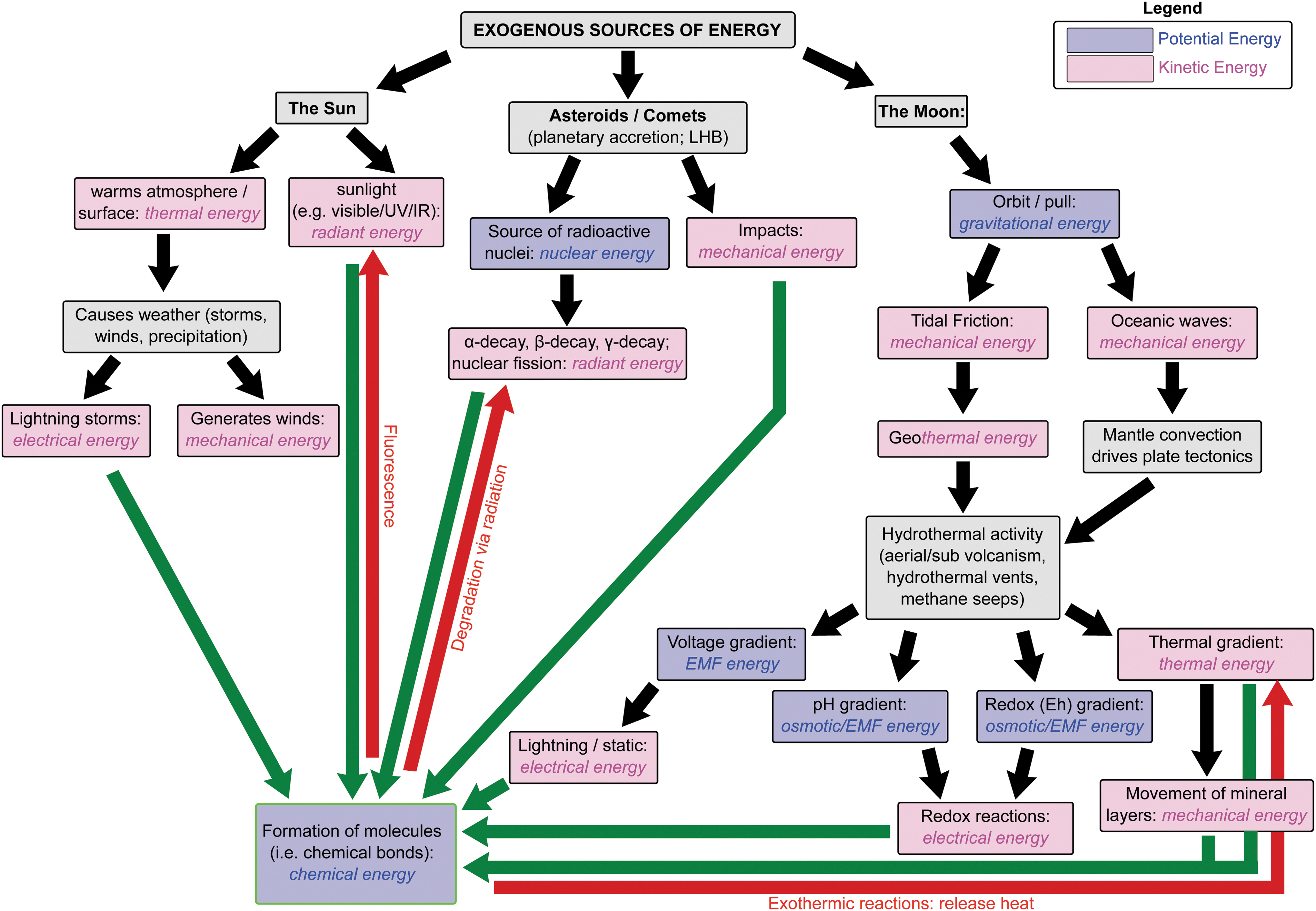
A summary of the origins and transformation of energy sources that helped shape early Earth, including the formation of biomolecules relevant for abiogenesis. EMF stands for electron motive force. LHB stands for Late Heavy Bombardment. Green arrows indicate reactions that lead to the formation of biomolecules; red, their degradation and transformation of their chemical energy into another energy source.
Geochemical gradients were vital to the initial creation of disequilibria which could serve as a source of potential energy for the origins of life. Gradients occur between two adjacent environments with significantly different geochemical parameters (e.g., temperature, pH, salinity, etc.) that are separated by some sort of semipermeable physical barrier that prevents rapid mixing and gradient dissipation. Because they drive ATP synthesis in cells via cellular respiration, pH gradients are the most vital for terran life (see Chapter 6.2.1). However, some organisms use Na+ gradients (Gemperli et al., 2003), a pathway that may have evolved first (Mulkidjanian et al., 2008).
On prebiotic Earth, pH gradients could have been generated along the semipermeable membranes of lipid vesicles (Bonfio et al., 2018; Dalai and Sahai, 2019) or semiporous rocks (Russell and Hall, 1997). If the gradient was maintained long enough, a current could have been generated that reduced CO2 by metal catalysts akin to biological CO2 reduction (Hudson et al., 2020). As pH gradients are maintained within serpentinizing alkaline vents, it might be possible that such systems were important for the emergence and/or persistence of life (see Chapter 4.3.2.1).
The most abundant elements within all life-forms are carbon (C), hydrogen (H), oxygen (O), nitrogen (N), phosphorus (P), and sulfur (S) (see Chapter 2.2.2). CHONPS elements are also some of the most abundant elements in the Solar System (see Chapter 3.1.1). Although their availability for prebiotic reactions on early Earth would have varied significantly depending on the geochemical environment, CHONPS species were likely relevant for the earliest forms of life. Indeed, elevated concentrations of these elements have been detected in an ancient, surface hot spring environment from the Pilbara Craton of Western Australia, where the oldest, most convincing evidence of life has been found in 3.5 billion-year-old rocks (Van Kranendonk et al., 2021).
Carbon
On early Earth, most carbon would have existed in oxidized forms: CO2 in the atmosphere or carbonate (CO3 2-) in the early oceans. The long-term geochemical cycling of CO2 and carbonate (i.e., the carbonate-silicate cycle) modulates Earth's climate over geologic timescales (see Chapter 3.4.2.2). To a lesser extent, carbon monoxide (CO) and possibly (in trace amounts) methane (CH4) could have been sourced from volcanic emissions, impact events, or photochemistry (see Table 4.2; Harman et al., 2015). However, any methane generated would have been unstable and readily destroyed by UV irradiation. While a myriad of chemical processes could have transformed gaseous CO2, CO, and CH4 into organic matter (Chapter 4.2.3), important prebiotic molecules may have also been delivered to early Earth via impactors (see Chapter 3.3.4).
Summary of Endogenous Sources and Sinks of Simple Reactive Molecules That Facilitate Organic Synthesis of Biologically Relevant Compounds
Summary of Endogenous Sources and Sinks of Simple Reactive Molecules That Facilitate Organic Synthesis of Biologically Relevant Compounds
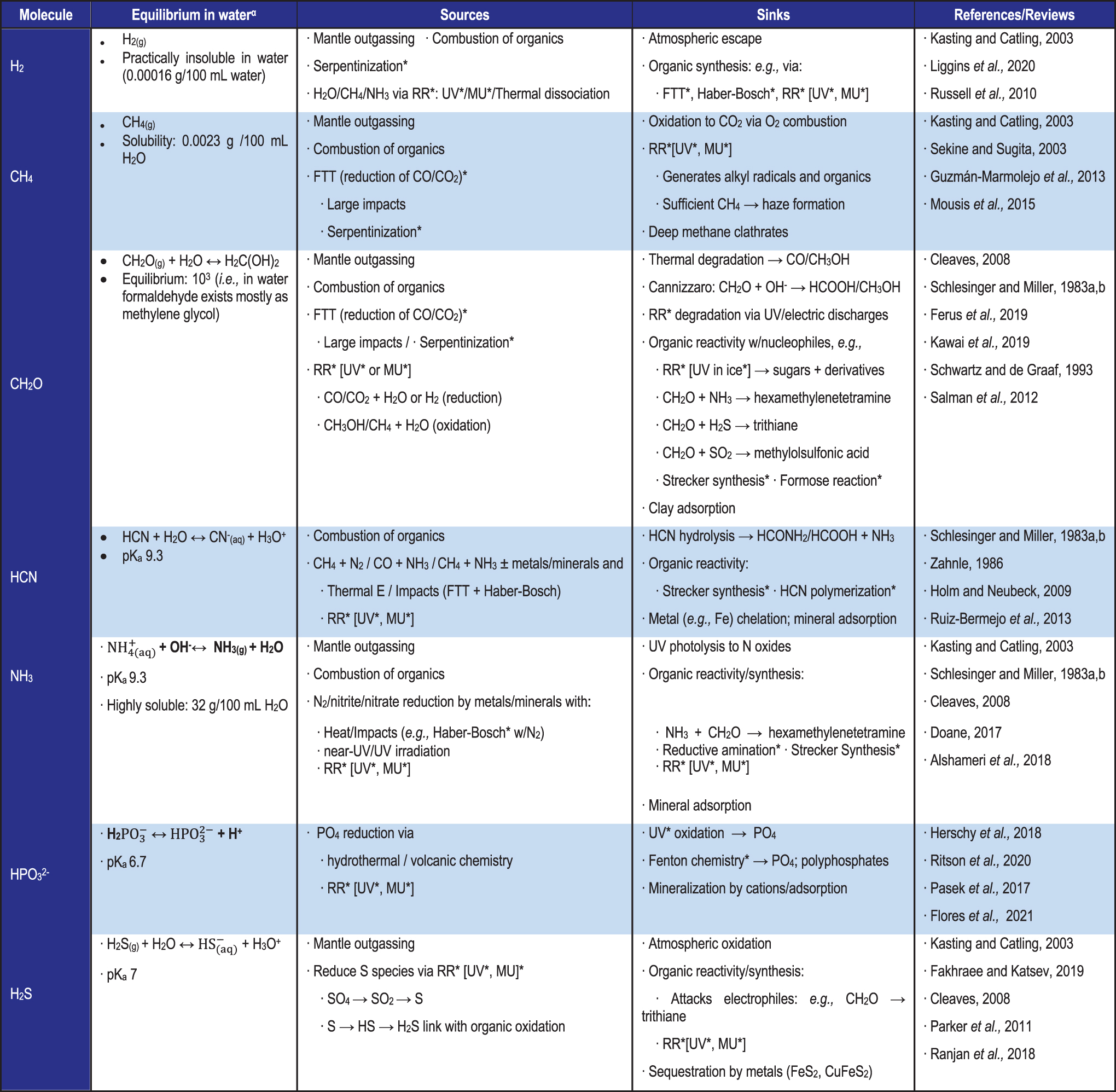
αThe chemistry for molecules that exist in equilibrium with its conjugate acid/base in solution (and the corresponding pKa of the species) is shown. Note that when the pH of a solution is equal to that of the pKa, the species is 50% deprotonated; thus, the pH has a strong influence on the reactivity of the molecules (e.g., charged species are electron-rich (e.g., SH-) and are better at attacking electron-deficient molecules (e.g., CH2O) in reactions). RR is radical recombination; FTT is Fischer–Tropsch type synthesis; MU is Miller–Urey chemistry (i.e., electric discharges/lightning). *See Chapter 4.2.3 for a detailed description and Figure 4.7 for the mechanism of each reaction.
Life on Earth is composed mostly of H and O in the form of water, which makes up ≥70% of cellular mass. Given the properties of water (see Chapter 2.2.2.1) and its abundance on Earth, H and O were likely critical for the origins of life. Although water would have been the dominant source of H and O, UV splitting of H-containing species (i.e., H2O, CH4, NH3, H2S), serpentinization reactions (see Chapter 4.1.4), volcanic outgassing (see Chapter 3.4.3.4), and impact events provided numerous sources of H2 (Zahnle et al., 2020). Atmospheric H2 is readily lost to space through thermal escape; if the H is sourced from H2O, the residual O2 remains behind (see Chapter 3.4.1.5). Gaseous oxygen (O2) could have also been generated via photolysis of oxygen-containing compounds other than water (i.e., CO, CO2, N2O, NO2, NO3, SO2).
Due to the understood redox state of the mantle, volcanism is not expected to have been a significant source of O2 on early Earth (see Chapter 4.1.4). Rather than accumulating in the atmosphere, photolysis-generated O2 would have been consumed via oxidation of reduced gases. Calculations suggest that the volcanic emission of H2 alone was more than sufficient to quench O2 generated via H2O splitting (Kasting and Catling, 2003); O2 would have also been lost due to oxidation of crustal materials. Overall, models estimate that in the Hadean O2 surface levels were ∼10−13 present atmospheric level (PAL) (Kasting and Catling, 2003). It has been predicted that H2 could have made up at least ∼1% of the Hadean atmosphere (Kasting and Catling, 2003; Tian et al., 2005; Liggins et al., 2020).
Nitrogen
Nitrogen was likely critical for the emergence of life, as it is a key component of nucleic acids and proteins (see Chapter 2.2.3). Like the modern atmosphere, N2 would have been the dominant N-containing species on early Earth, with trace amounts of NH3 also present (Chapter 4.1.4). Radical chemistry (see Chapter 4.2.3.8) and/or fixed N species (e.g., nitrate (NO3 2-), nitrite (NO2 2-), or ammonia/ammonium (NH3/NH4 +)) would have been required for abiotic synthesis of N-containing biomolecules. Abiotic nitrogen fixation could have occurred via lightning, mineral redox, photolysis, or impact events (Table 4.2; Doane, 2017).
The conversion of atmospheric N2 and NOx - to ammonia (NH3) on early Earth is highly relevant to origins-of-life chemistry given that (i) amine groups (R-NH2) are ubiquitous in living matter, whereas those with nitroso groups (R-N = O), nitro groups (R-NO2), and nitrite/nitrate esters (R-O-N = O/R-O-NO2) are rarely found in life; and (ii) life uses ammonia as the precursor for N-containing molecules. While ammonia is produced via the irradiation of N2 adsorbed onto dried titanium-oxides, continued irradiation oxidizes ammonia to N2 and NOx -, hindering it from accumulating above trace levels in Earth's atmosphere (Kasting and Catling, 2003; Doane, 2017). However, if the minerals were submerged in water, the resulting ammonia could have been readily sequestered to deeper waters, inhibiting further photolysis (Hirakawa et al., 2017; Mateo-Marti et al., 2019). Alternatively, ammonia could have formed in deeper waters via hydrothermal reduction of nitrate, nitrite, and N2 in the presence of Fe-containing minerals (Smirnov et al., 2008; Nishizawa et al., 2021).
Green rust as well as dissolved ferrous iron (Fe2+) or copper (Cu2+) can also reduce nitrite and nitrate to ammonia under mild, alkaline conditions (pH >7, ∼25°C; Cho et al., 2010). Nitrite and nitrate are generated efficiently via lightning through an N2-rich atmosphere (Cleaves et al., 2008; Wong et al., 2017). Models accounting for the various sources and sinks of nitrate and nitrite built by Ranjan and colleagues postulate that the dissolved NOx - concentrations in the Hadean ocean would have been <1 μM (pH 6.5–8; T = 0–50°C), with significantly less nitrite due to its reactivity, compared to ∼40 μM nitrate in modern oceans (Olsen et al., 2016; Ranjan et al., 2019). Significant amounts of ammonia may have also been delivered to early Earth via comets and asteroids (Pizzarello et al., 2011; Altwegg et al., 2020).
Phosphorus
Since phosphate is rapidly sequestered into insoluble precipitates by dissolved cations that were likely common in Earth's early oceans (i.e., Ca2+, Fe2+, Fe3+, Al3+, or Mg2+), it remains unclear what the most prominent source of phosphorus may have been (Pasek et al., 2017). Early Earth's oceans on average contained an estimated 0.04–0.13 μM of dissolved phosphate (Jones et al., 2015), which is an order of magnitude less than that in modern oceans (typically 0.2–4 μM; Patey et al., 2008) and orders less than that in modern cells (0.5–10 mM; Bevington et al., 1986). This solubility issue is commonly dubbed the “phosphate problem.”
The phosphate problem could have been resolved in solutions with (i) anions that bind cations other than phosphate preferentially (e.g., carbonate-rich waters that sequester Ca2+; Toner and Catling, 2019), (ii) organic acids or other solubilizing agents that liberate precipitated phosphate (Burcar et al., 2016), or (iii) minerals that adsorb phosphate and later release it in concentrated batches or polymerize it with co-adsorbed organics (Barge et al., 2014; Flores et al., 2021). The reduced form of phosphate (i.e., phosphite, HPO3 2-) is more soluble and is relatively stable under early Earth conditions (see Table 4.2). Oxidation of phosphite via Fenton chemistry generates phosphate and polyphosphates, the latter of which can phosphorylate organics in aqueous solutions (see Chapter 4.2.3.9); polyphosphates can also be generated from phosphate minerals subjected to lightning (Çalışkanoğlu et al., 2023). Alternatively, iron-rich meteorites may have delivered a significant amount of the phosphorus mineral schreibersite ([Fe,Ni]3P) that upon corrosion can phosphorylate organics (Gull et al., 2015). Notably, organophosphate bonds break in aqueous solutions subjected to UV irradiation—this process would have been key for recycling phosphorus on prebiotic Earth (Farr et al., 2023).
Sulfur
On Hadean Earth, S existed mostly as sulfur dioxide (SO2) and, to a lesser degree, hydrogen sulfide (H2S) or polysulfur (S8). In the absence of oxygen, sulfate (SO4 2-), the most common form of S on modern Earth, would have been rare (Crowe et al., 2014; Fakhraee and Katsev, 2019). Upon dissolution into Earth's early oceans, SO2 formed sulfurous acid (H2SO3) and existed in equilibrium with bisulfite (HSO3 -) and sulfite (SO3 2-). The equilibrium was pH dependent with strongly acidic solutions (pH <1.85) favoring SO2 and neutral/alkaline (pH >7) favoring sulfite.
Unlike modern Earth, where sulfite species are rapidly oxidized to sulfate, sulfite could have accumulated on early Earth since its only major sink would have been oxidation to sulfate via UV irradiation (Ranjan et al., 2018). This oxidation mechanism can be coupled with reduction of HCN to form biologically relevant organics (Ritson et al., 2018; Xu et al., 2018). Meanwhile, sulfide from dissolved H2S would have been consumed by organic electrophiles and metals forming organic thiols (R-SH) and metal sulfides. Mineralization of organic thiols would have regenerated dissolved sulfide (HS-) and the production of sulfite (SO3 2-) (Fakhraee and Katsev, 2019). Models predict that (bi)sulfite species were more abundant (μM at 25°C; mM at 0°C; nM at 50°C) than dissolved sulfide (nM range 0–50°C; Ranjan et al., 2018).
Chemical mechanisms that form biologically relevant monomers
Prebiotic synthesis of primitive biopolymers and their monomeric precursors stems from the reactivity of CHONPS molecules that would have been readily available on Hadean Earth. However, as discussed in Chapter 4.2.2, the most stable form of CHONPS elements required for biosynthesis was relatively unreactive, oxidized molecules (i.e., CO2, CO3 2-, H2O, N2, PO4, SO3 2-, SO2). Thus, a continuous source of reactive and reduced molecules was needed (Table 4.2) for high yields of biologically relevant compounds to have formed (Fig. 4.7). This section will discuss several abiotic chemical mechanisms that may have been important for the origins of life.
Serpentinization (Fig. 4.7A)
The serpentinization reaction is the hydrolysis of a mineral within a rock, generating H2 as a by-product. Serpentinization most often occurs with ultramafic minerals that are common in Earth's mantle (see Chapter 4.1.2), and it may have also served as a source for H2 on early Earth (Sleep et al., 2004) or on other worlds (see Chapter 7.1). The production of H2 from ongoing serpentinization has been observed in the field including at Lost City Hydrothermal Field (Kelley et al., 2005), Rainbow Hydrothermal Field (Charlou et al., 2002), and the Oman ophiolite (Neal and Stanger, 1983). Serpentinization generates OH- anions that would have resulted in pH gradients with early Earth's slightly acidic waters, fueling CO2 reduction to methane and potentially ATP production for the earliest forms of life (Martin et al., 2008; Russell et al., 2010; Sojo et al., 2016; Hudson et al., 2020).
Haber-Bosch process (Fig. 4.7B)
In the Haber-Bosch reaction, nitrogen (N2) and hydrogen-containing gases (usually H2 or CH4) are combined under high temperatures (400–650°C) and pressures (150–300 bar) to form ammonia (Rodriguez et al., 2012). This reaction requires dry conditions (i.e., the absence of water) and the presence of a strongly reducing metal catalyst, typically zero-valent iron (Fe0) or ferrous iron (Fe2+).
The prebiotic relevance of Haber-Bosch reactions remains uncertain given that the required conditions would have been rare on early Earth. The Haber-Bosch process still could have served as a minor source of ammonia during impact events where the requisite dry, high temperatures and high pressures could occur (Shimamura et al., 2017). Impacts by stony-iron meteorites would be particularly conducive to this chemistry, given that they are enriched in the catalysts that can drive the reaction such as reduced iron and nickel alloys and the minerals magnetite (Fe3O4; source of Fe2+), wüstite (FeO; source of Fe0), and schreibersite ([Fe,Ni]3P; source of Fe0) (Rubin and Ma, 2017).
Strecker synthesis (Fig. 4.7C)
Strecker synthesis occurs when ammonia, cyanide, and an aldehyde combine to form α-amino acids (Fig. 4.6). It begins with condensation of the ammonia and aldehyde to form an imine, which is then attacked by the cyanide anion. The cyano group can then be hydrated, generating a carboxylic acid. The Strecker synthesis has been posited to have occurred in the primitive ocean (Miller and Van Trump, 1981) and to have been the major source for α-amino acids within meteorites (Burton et al., 2012).
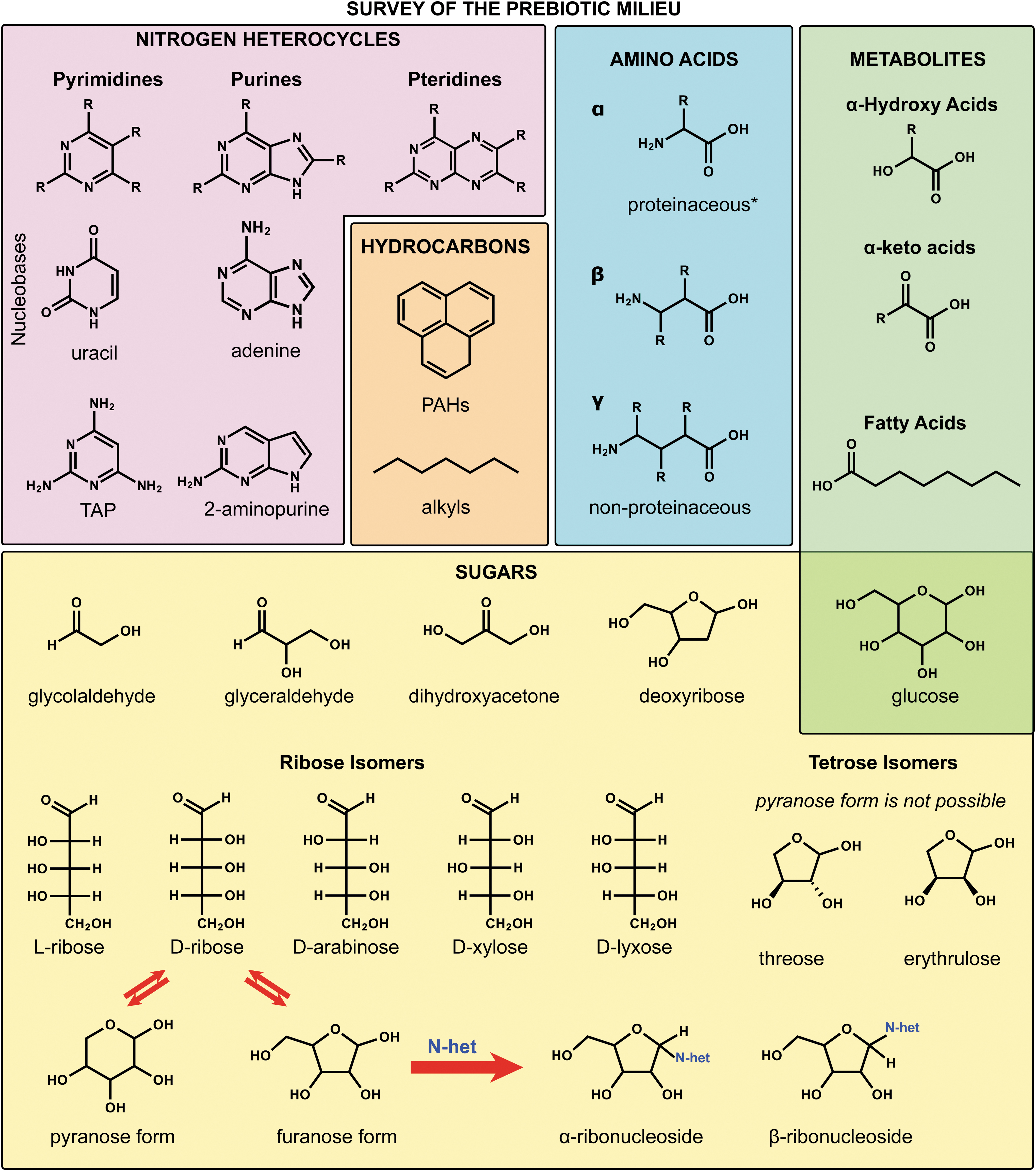
Possible molecular building blocks relevant to the origins of life, including molecular building blocks: nucleobases, amino acids, hydroxy acids, fatty acids, and sugars. N-het is short for N-heterocycle. The reaction with the N-heterocycle and ribofuranose show how the reaction can produce two nucleoside isomers. *Proteinaceous amino acids are those used by life to make proteins.
Reductive amination is a process by which substituted amines such as amino acids can be generated via α-keto acids (Muchowska et al., 2017; Barge et al., 2020). This reaction can occur at relatively low temperatures (i.e., 23–70°C) and is initialized with the condensation of an amine or ammonia (NH3) onto an aldehyde/ketone (R2C = O) to form an imine intermediate (i.e., R2C = NH) that is then further reduced to the amine product (i.e., HR2C-NH2). Iron is typically used as the reductant (i.e., electron source) and water as a proton source.
Fischer–Tropsch type (FTT) synthesis (Fig. 4.7E)
Fischer–Tropsch reactions can efficiently generate liquid hydrocarbons (e.g., alkanes, alkenes, alcohols) from hydrogen and carbon monoxide in the gas phase. This process is performed with metal catalysts at high temperatures and pressures (typically 200–350°C; 5–275 bar; Speight, 2015). When conducted in aqueous solutions and/or in the presence of other gases such as NH3 or H2S, the reactions are known as Fischer–Tropsch type (FTT). Such reactions have been proposed as potential abiotic sources of fatty acids (with up to 18-carbon-long side chains; McCollom et al., 1999), amino acids, nucleobases (Fig. 4.6), and hydroxy acids, among other organics in various planetary environments, including within the deep subsurface of the crust, hydrothermal vents, meteorites, and the solar nebula (Kress and Tielens, 2001; Simoneit, 2004). FTT chemistry may have also been the source of organic matter found in the oldest Archean rocks, making it difficult to discern the biogenicity of potential microfossils (Lindsay et al., 2005).
Formose reaction (Fig. 4.7F)
The formose reaction (Butlerov, 1861) describes the polymerization of formaldehyde (CH2O) and is the leading mechanism proposed for the prebiotic origin of sugars, including ribose (the sugar in RNA), glucose (critical for modern metabolism), and other sugar-like compounds, namely, sugar alcohols, sugar acids, and sugar polymers (i.e., polysaccharides). Degradation of these molecules leads to the formation of a complex mixture which includes hydroxy acids relevant to metabolism (i.e., metabolites; Weber, 1992; Omran et al., 2020). The initial (and slowest) step of the mechanism is the condensation of two formaldehydes to form glycolaldehyde (Fig. 4.7F). This induction period is sped up when trace amounts of glycolaldehyde or higher-weight sugars are present in solution (Kitadai and Maruyama, 2018).
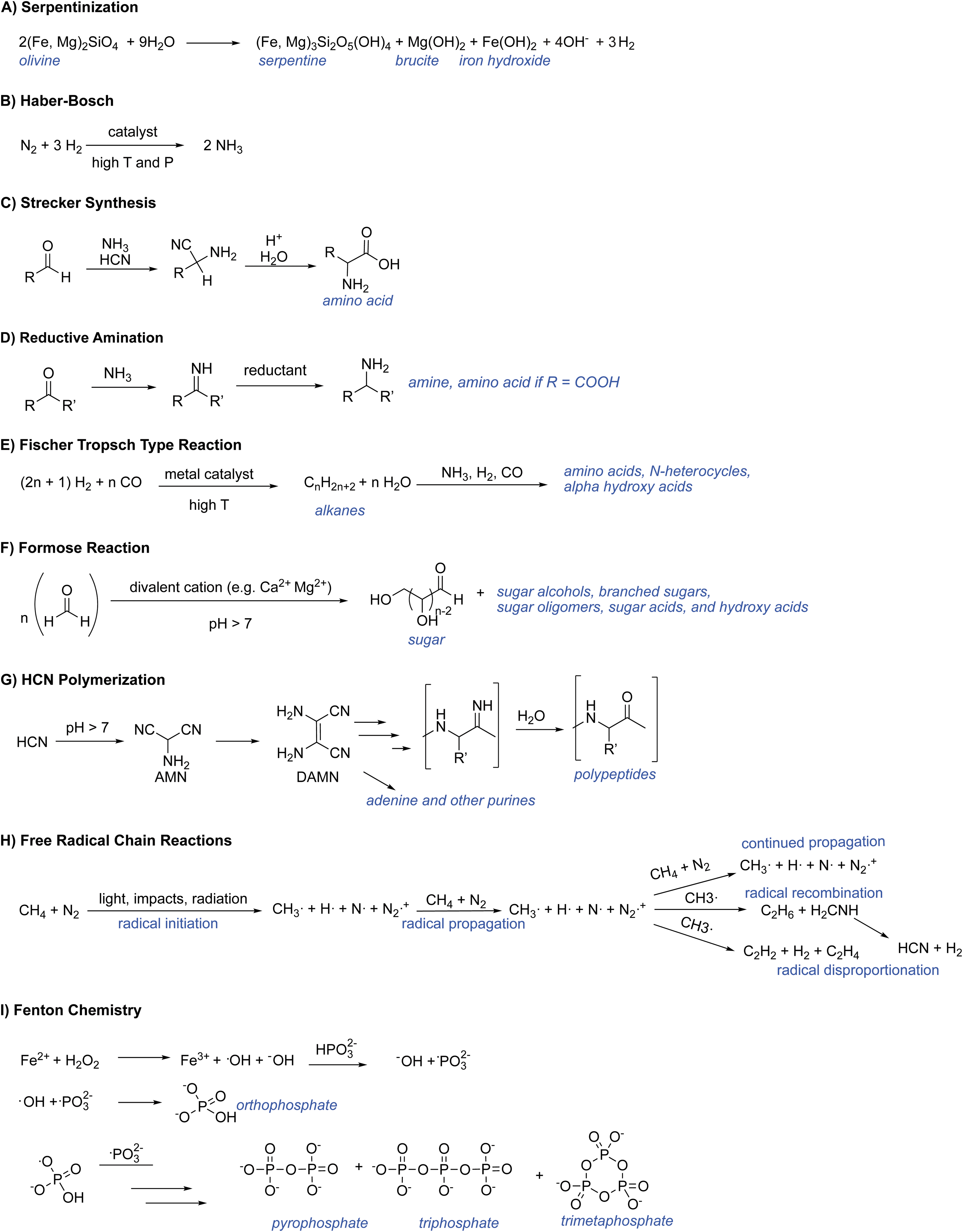
Chemical mechanisms relevant for transforming simple reactive molecules to biologically relevant compounds.
The formose reaction requires higher temperatures (60–80°C), basic conditions (usually pH 10–11), and a catalyst—commonly Ca2+, Mg2+, however other divalent (e.g., Ba2+, Sr2+, Pb2+), monovalent (e.g., Na+, K+) and trivalent cations (e.g., Fe3+), as well as organic bases (e.g., pyridine) (Weber, 1992; Kopetzki and Antonietti, 2011) can also catalyze the reaction. The relevance of the formose reaction for abiogenesis has been questioned given that, if not stopped quickly, it results in the degradation and polymerization of the sugars, forming insoluble tar (Shapiro, 1988). Moreover, ribose is generated in relatively low yields among a large assortment of other side products (including ribose isomers, see Fig. 4.6). Notably, the presence of inorganic reagents, namely borates and, to a much lesser extent, silicates, has been shown to stabilize cis-diols including pentoses like ribose (Lambert et al., 2010; Nitta et al., 2016). Alternatively, specific reagents present within the solution could have sequestered ribose (Springsteen and Joyce, 2004). While the formose reaction has been invoked as a potential source for sugars and sugar derivatives found in meteorites and irradiated interstellar ice analogs (Cooper and Rios, 2016; Meinert et al., 2016; Furukawa et al., 2019), it should be noted that other (lesser known) mechanisms may be at play (Nuevo et al., 2018). It also has been proposed that a “glyoxylate scenario,” rather than formose reactions, may have produced the first sugars, given it can yield ribose directly as a product (Eschenmoser et al., 2007; Fialho et al., 2018). This mechanism is in stark contrast with the formose reaction wherein a plethora of products are generated.
Hydrogen cyanide (HCN) is a weak acid (pKa 9.2) and in aqueous solutions will exist in equilibrium with the cyanide anion: HCN ↔ CN- + H+. HCN has a boiling point of only 25.6°C at low pH, but under more basic conditions (pH >8) the proportion of cyanide anion (CN-) increases, and at pH >9.2 it is the dominant form. At high-enough concentrations, CN- readily undergoes polymerization even at mild conditions (i.e., 1 bar, 25°C) to form a range of compounds including hydroxy acids, amino acids, N-heterocycles (e.g., purines, pyrimidines, and pteridines; Fig. 4.6), and polypeptides (Voet and Schwartz, 1982; Marín-Yaseli et al., 2015). Although a catalyst is not needed for this reaction, NH4 + and O2 can facilitate it (Marín-Yaseli et al., 2018). The ability for HCN polymerization to generate biochemical building blocks has led some researchers to suggest that HCN could facilitate abiogenesis on planetary bodies (Matthews, 2004; Matthews and Minard, 2006; Sasselov et al., 2020).
Sufficiently high HCN concentrations (>0.01 M) would be difficult to achieve on early Earth, especially considering the high reactivity of the CN- anion with aldehydes. Solutions containing lower concentrations undergo hydrolysis to form formamide (HCONH2) and formic acid (HCOOH) (Sanchez et al., 1967). Mechanisms by which CN- may have been concentrated include freezing of dilute CN- solutions (Menor-Salván and Marín-Yaseli, 2012) or precipitation over time with dissolved metals and subsequent release (Ruiz-Bermejo et al., 2013). HCN, or products of HCN polymerization, may have also been delivered by comets (Biver et al., 2002) and meteorites (Callahan et al., 2011; Smith et al., 2019).
Free-radical chain reactions (Fig. 4.7H)
Reactions involving free radicals typically have low to no activation energy. Consequently, radicals can generate a variety of products, including polymers, from otherwise stable, less reactive molecules (e.g., CO2, N2, etc.). Free-radical chain reactions are characterized by three phases. First, the chain is initiated by the formation of a radical species, a process that requires significant energy. Common sources for the initiation phase include thermal energy (e.g., induced by impact events), ionizing radiation (e.g., UV light, X-rays, γ-rays, α-particles, β-particles), and electric currents (e.g., lightning, redox reactions). Second, propagation occurs due to the interaction of radicals with other atoms, molecules, or ions to form more radicals either via hydrogen abstraction (removal of •H) or addition of the radical to the double/triple bond of another compound. Finally, termination occurs when radicals react with other radicals to form nonradicals either via radical recombination or radical disproportionation (see Fig. 4.7H).
Radicals, being short-lived, exist in significantly lower concentrations than nonradicals. Consequently, the termination phase only occurs after many propagation cycles and explains how a wide range of organic compounds can be derived from radical chain reactions involving simple volatiles (e.g., CO2, CH3OH, CO, or CH4). If other molecules are present (e.g., N2, H2O), an even wider array of products can be generated. For example, UV irradiated ices containing volatiles form a diverse array of organic compounds including nucleobases, sugars, hydroxy acids, and amino acids (Fig. 4.6; see Chapter 3.3). In addition, radicals can also efficiently transform stable molecules (e.g., benzene, naphthalene) into more reactive compounds. For a review on the organics that can be generated from irradiated ices see (Sandford et al., 2020).
Fenton reaction (Fig. 4.7I)
The classic Fenton reaction describes the generation of hydroxyl radicals (•OH) during the oxidation of ferrous iron (Fe2+) by hydrogen peroxide (H2O2). Once formed, the hydroxyl radicals oxidize inorganic ions and organic molecules. Similar processes could have occurred on prebiotic Earth given that its oceans were enriched in ferrous iron. Potential prebiotic sources for peroxide include (i) oxidation of water by pyrite and nickel disulfides (Borda et al., 2001); (ii) irradiation of atmospheric gases (Kasting et al., 1989); and (iii) water oxidation at water-rock interfaces subjected to stress (Balk et al., 2009).
Fenton reactions can occur at room temperature and proceed most efficiently within acidic solutions (pH 2–4) which promote the dissolution of minerals. Fenton-like reactions under alternative conditions have been carried out (i.e., using Cu2+, Mn2+, Co2+, Ni2+, V2+, Ti2+, Cr2+), suggesting this mechanism may have been prevalent at acidic black smoker vents (pH 2–4) where these metals can accumulate alongside peroxide (Borda et al., 2001; Kelley et al., 2002; Balk et al., 2009; Tivey, 2015). Notably, Fenton reactions may have been key to resolving the phosphate problem (see Chapter 4.2.2.4), as they could have oxidized soluble phosphorus species, phosphite (HPO3 2-) and hypophosphite (H2PO2 -) to polyphosphates whose high-energy bonds could phosphorylate organics (Fig 4.7I; Pasek et al., 2008, 2017; Gull et al., 2015) or served as energetic precursors to ATP (Lipmann, 1984; Holm and Baltscheffsky, 2011).
Miller–Urey spark discharge reactions
In 1953, Stanley L. Miller and Harold Urey revolutionized origins-of-life research when they demonstrated that biologically relevant organic compounds could be generated from electric discharges through a reducing atmosphere (e.g., CH4-NH3-H2-H2O) over water—simulating the chemistry that may occur on Earth during a thunderstorm or volcanic eruption (Miller, 1953). Miller–Urey reactions generate a complex mixture of organics that includes hydroxy nitriles/amides/acids, amino nitriles/amides/acids, aldehydes, HCN, HCN polymers, nucleobases, and more (McCollom, 2013; Parker et al., 2016; Ferus et al., 2017). Although initial experiments showed that a N2-CO2 atmosphere did not facilitate the formation of organics or any detectable amino acids (Schlesinger and Miller, 1983b), subsequent work showed that organics are generated when the reaction occurs over a pH buffered solution (Cleaves et al., 2008; Rodriguez et al., 2019). When the starting gases interact with a spark discharge, radicals form and recombine into new molecules. Dissolution of these organics into an aqueous solution can lead to subsequent reactions such as HCN polymerization and the Strecker synthesis.
Researchers have noted that the products generated from Miller–Urey chemistry are similar to those formed from UV irradiation through a reducing atmosphere simulating both the formation of Titan and Triton tholins (Cleaves et al., 2014). Furthermore, the distribution of organics detected in meteorites and via FTT synthesis has also been shown to be similar to Miller–Urey mixtures (McCollom, 2013). These studies demonstrate that different energy sources may not have a significant impact on the resulting organics generated via these complex processes.
The prebiotic synthesis of biologically relevant macromolecules
Several biomolecules are critical for terran life, namely, (i) DNA/RNA to store information, (ii) proteins to catalyze reactions, (iii) a phospholipid membrane to buffer cellular processes, and (iv) polysaccharides for structural support and energy storage (see Chapter 2.2.3 for details). Whether each molecule was needed for abiogenesis or was the product of biological evolution remains debated, although the idea that biomolecular polymers and their functional capabilities are the result of their coevolution and mutual relationships has gained favor in recent years (see Frenkel-Pinter et al., 2020 and references therein).
Informational molecules: nucleic acids and their precursors
The informational polymers used by extant life are the nucleic acids DNA and RNA (see Chapter 2.2.3.1). It is hypothesized that RNA may have emerged first, serving as a dual genetic-enzymatic biomolecule (i.e., the RNA World hypothesis; Gilbert, 1986) given (i) a wide array of ribonucleotides (RNAs) function as universal metabolic cofactors (Fig. 4.8C) and (ii) catalytic RNAs carry out various critical roles in the cell (Saad, 2018). Alternatively, RNA could have preceded DNA but still have been the product of evolution stemming from a different informational molecule (i.e., pre-RNA/proto-RNA; Robertson and Joyce, 2012; Hud et al., 2013; Cleaves et al., 2019). However, it is not certain whether (proto)life transitioned from one genetic system to another (i.e., a genetic takeover) or whether various genetic systems (i.e., pre-RNAs, RNA, DNA) could have coexisted and coevolved (Bhowmik and Krishnamurthy, 2019; Xu et al., 2020).
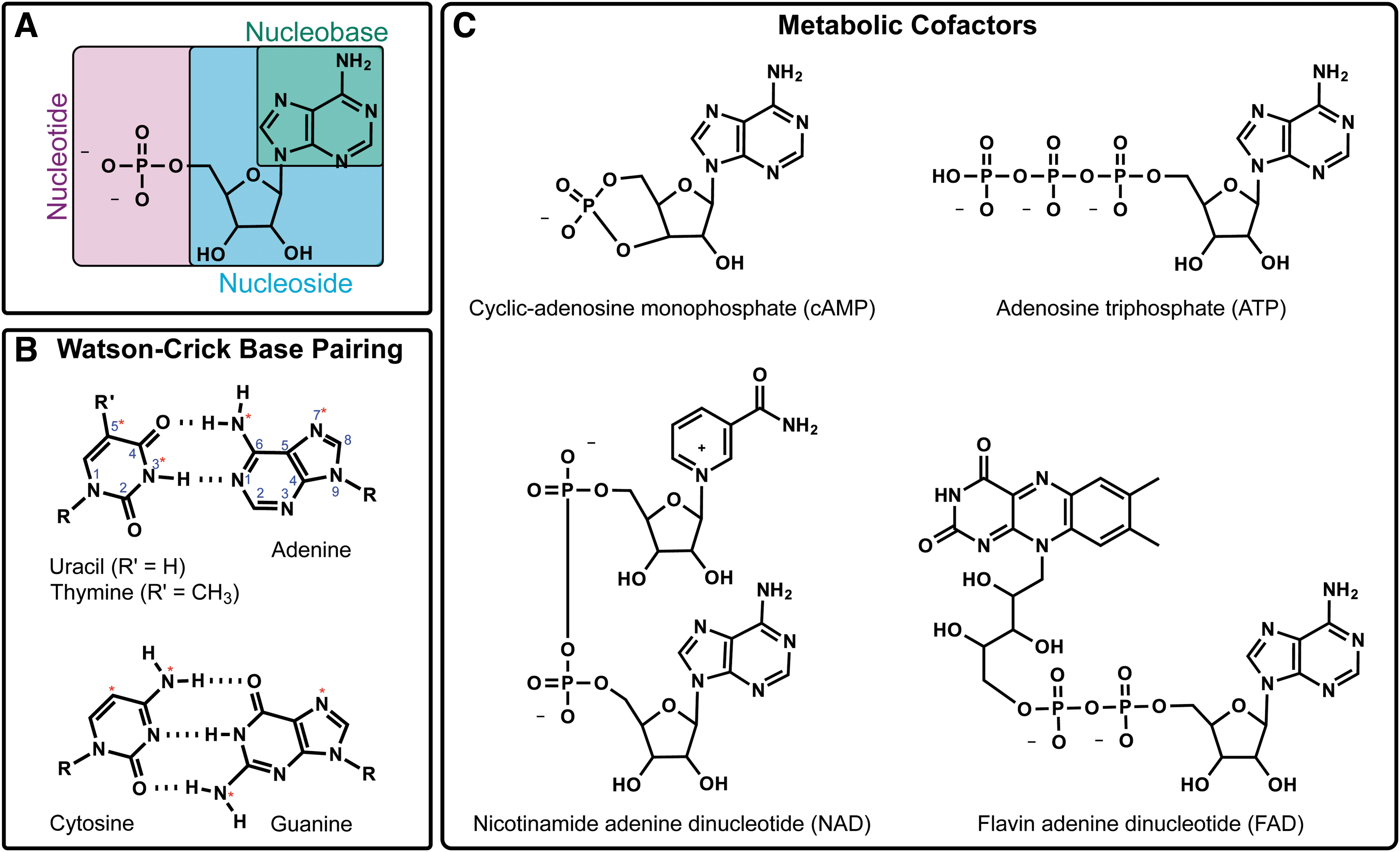
A primer to nucleic acids. (
Figure 4.9 showcases how pre-RNAs could have used alternative molecules for each of the three units (the nucleobase, sugar, and phosphate) within nucleic acids. Researchers have suggested a variety of prebiotic mechanisms for the formation of (non)canonical nucleobases, including cyanide polymerization (see Chapter 4.2.3.7), and others (Menor-Salván, 2009; Ferus et al., 2017). Ribose/deoxyribose sugars may have been derived from the formose reaction (see Chapter 4.2.3.6) or alternative pathways (Eschenmoser, 2007; Roche et al., 2022).
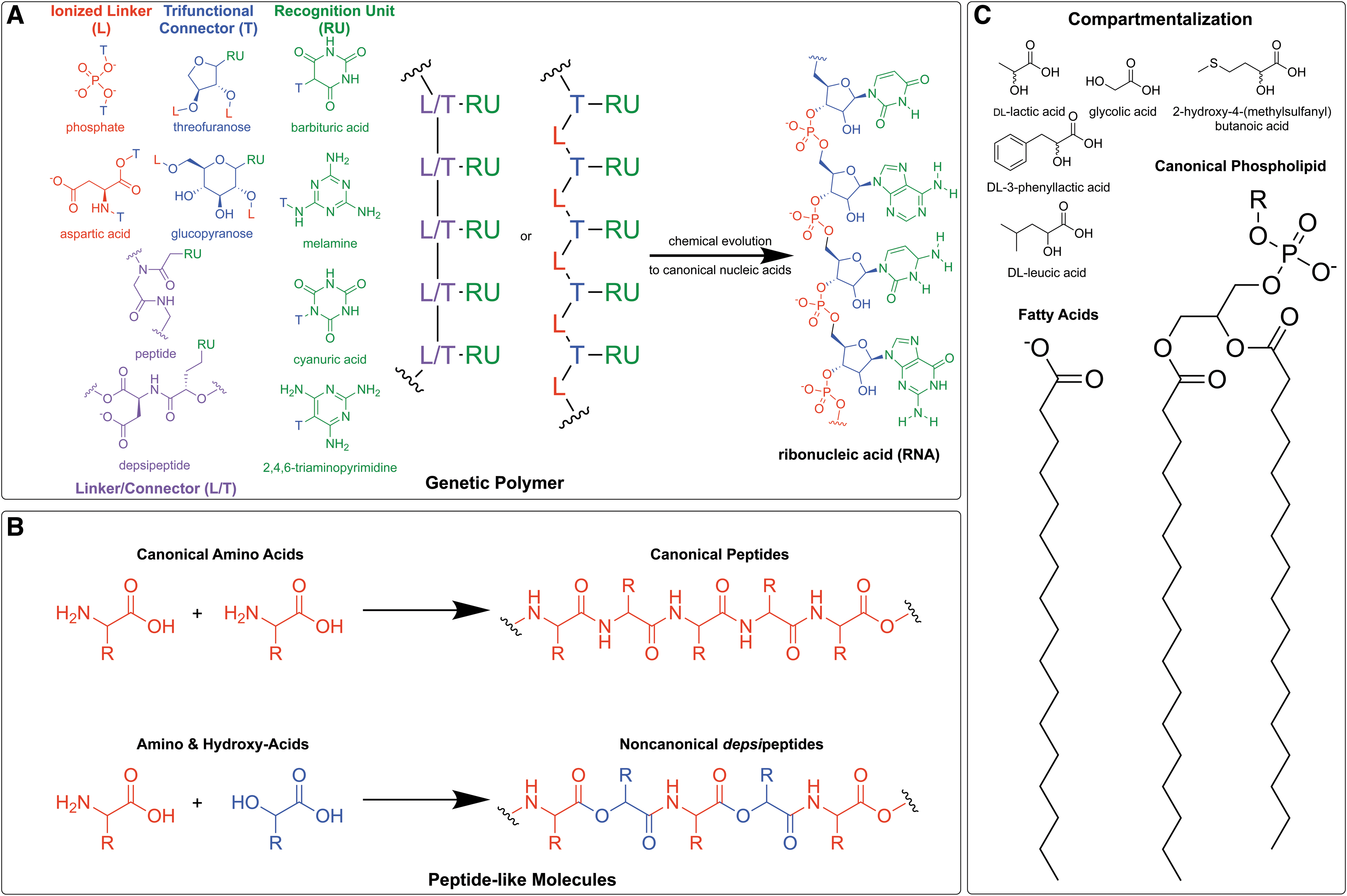
Alternative or nonstandard biomolecules with prebiotic significance. Sets of molecules that could have had prebiotic significance as either facilitators or precursors of canonical, or modern, biomolecules. (
The prebiotic synthesis of nucleotides (a combination of the nucleobase, sugar, and phosphate units) can be accomplished by one of two different strategies (Fig. 4.10). In the first, the three units are produced separately and then combined via condensation reactions that are typically thermodynamically unfavorable in water (see Chapter 4.2.4.3; Fuller et al., 1972), though it may be more favorable for noncanonical nucleobases (Kolb et al., 1994; Fialho et al., 2020). This could involve sugar condensation onto nucleobases followed by phosphorylation. Alternatively, cyclic phosphorylated sugars in solution could have reacted with purine nucleobases, generating the corresponding nucleotides (Kim and Benner, 2017). In the second strategy (i.e., the indirect approach), the molecular units are formed simultaneously on a single scaffold via low molecular weight compounds (Powner et al., 2009; Becker et al., 2019; Kruse et al., 2020). Alternatively, solutions containing primed pyrimidines such as those with a formyl (R-CHO) or vinyl (R-CH = CH) group can readily undergo aldol reactions with surrounding sugars forming purine ribonucleosides (Becker et al., 2019) and pyrimidine deoxyribonucleosides (Teichert et al., 2019).
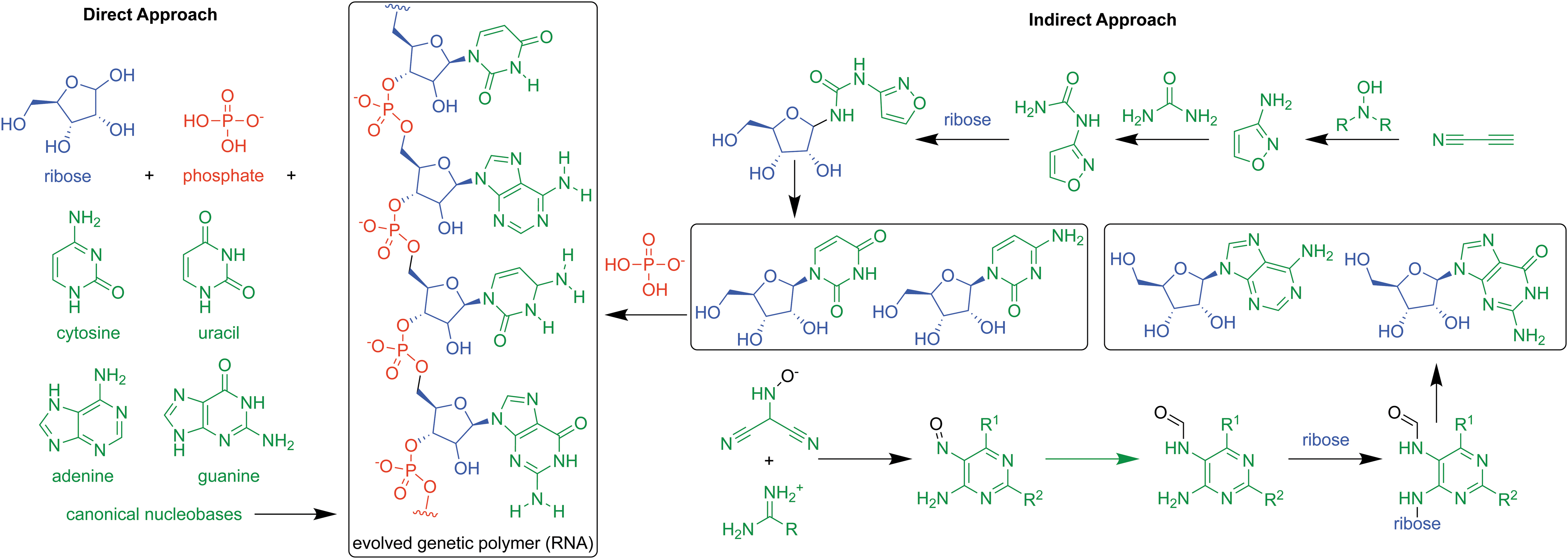
Prebiotic synthesis pathways of nucleotides and RNA via the direct (left) and indirect (right) approaches. In the direct approach, sugar, phosphate, and nucleobase components are formed separately and react with each other directly to form nucleotide monomers, which can then polymerize to form RNA. In the indirect approach, atoms destined for the final nucleotide are built onto chemical intermediates in a series of reactions in one-pot (i.e., all together in one solution) before undergoing phosphorylation to form the requisite nucleotides. In either case, polymerization of the resulting nucleotides remains a separate stage in chemical evolution. Figure adapted from Becker et al. (2019).
Although phosphorylation of nucleosides is thermodynamically unfavorable in aqueous solutions (see Chapter 4.2.2), the reaction has been shown to occur (i) in the presence of activated phosphates, namely, diamidophosphate (Gibard et al., 2019), trimetaphosphate (Kolb and Orgel, 1996), or phosphate radicals generated from schreibersite (a meteoritic mineral, [Fe,Ni]3P) corrosion (Gull et al., 2015); and (ii) in solutions with sufficiently low water activity (i.e., formamide, urea-rich solvents, or solutions subjected to wet-dry cycles) (Gull et al., 2015).
Researchers have also proposed informational systems based on alternative polymers. Metabolic cycles and other forms of complex chemical systems could have allowed for information storage, transfer, and replication (Segré et al., 2000). For example, amyloids, which are aggregates of polypeptides arranged in multiple β-sheets held together via hydrogen bonds, can store information, catalyze reactions, self-propagate, and (unlike RNA) are relatively stable in harsh aqueous solutions (Frenkel-Pinter et al., 2020).
Polypeptides (Fig. 4.9B), or proteins, serve as the catalytic polymers in extant life and are composed of individual monomers called amino acids (see Chapter 2.2.3.2). For proteins, modern life utilizes exclusively 20 α-amino acids in ribosomal synthesis, although non-proteinaceous amino acids (e.g., β-, γ-, and α-amino acids) are used to carry out a myriad of functions. Abiotic reactions can form β-, γ-, and other non-proteinaceous α-amino acids; thus the chemical reasoning behind the selection for the 20 encoded amino acids is an open question. One possibility is that the proteinaceous α-amino acids have a relatively easier time oligomerizing compared to their similar, non-proteinaceous counterparts (Frenkel-Pinter et al., 2019).
Amino acids are hypothesized to have formed prebiotically through several different pathways, including Miller–Urey chemistry, HCN polymerization, the Strecker synthesis, FTT reactions, and reductive amination (see Chapter 4.2.3); they may have also been delivered to early Earth by meteorites (Burton et al., 2012). The difficulty in polymerizing amino acids in aqueous solutions has led some researchers to suggest that proteins are the product of chemical evolution and that the first catalysts may have been alternative compounds such as Fe/Fe-S minerals or organic clusters (see Chapter 4.2.6 for details). Other suggestions include polymers of hydroxy acids (i.e., polyesters), hydroxy and amino acids (i.e., depsipeptides, Fig. 4.9B), or mercaptoacids and amino acids (i.e., thiodepsipeptides) (Forsythe et al., 2015; Mamajanov and Cody, 2017; Frenkel-Pinter et al., 2022).
Polymerization of organic molecules
Terran biopolymers are formed through condensation linkages, which are thermodynamically unfavorable in water. Polymerization could have been promoted by low water activity induced by wet-dry cycling, freeze-thaw cycling, hydration of inorganic salts, or the presence of brines (Da Silva et al., 2015; Forsythe et al., 2015; Campbell et al., 2019). Alternatively, polymerization could have been facilitated via deprotonation of the amine via mineral adsorption (Kitadai et al., 2017), with the help of radicals (Pasek et al., 2008) or condensing agents such as cyanamide (Ibanez et al., 1971), or by activating the monomers via phosphorylation or thiolation (adding a thiol group forming a thioester) (Weber and Orgel, 1979; Burcar et al., 2015). Polypeptide polymers may have also been generated via replacement of hydroxy acids in polyesters with amino acids during wet-dry cycling (Forsythe et al., 2015) or due to polymerization of amino amides (NH2-CHR-CONH2) (Brack, 2007).
Studies into how prebiotic oligomers/polymers fold, especially their tertiary and quaternary structures, and their catalytic capabilities remains understudied (Frenkel-Pinter et al., 2020). However, an understanding of the overall structures that prebiotic polymers can take on—and the stability of these structures under fluctuating environmental conditions—could inform the selective pressures which resulted in life's specificity for its building blocks (i.e., the biological nucleobases and D-ribofuranose isomer; see Fig. 4.6) for nucleic acids and the proteinaceous amino acids for proteins.
Role and formation of compartments
Modern life is composed of cells whose metabolic reactions and materials are enclosed within membranes. Encapsulation was important for chemical evolution of self-replicators as it prevents their diffusion into the environment and minimizes the possibility of a takeover by parasitic molecules (i.e., molecules with no catalytic activity; Bansho et al., 2016). The cells of modern life rely on a bilayer membrane of phospholipids (Fig. 4.9C; see Chapter 2.2.3.3), molecules containing two long carbon-chain fatty acids, glycerol, and phosphate. The prebiotic synthesis of modern phospholipids containing two hydrocarbon chains has not yet been shown, prompting some to propose that the earliest membranes used simpler molecules such as single-chain fatty acids to form vesicles (Apel et al., 2002; Hanczyc et al., 2003; Deamer, 2017). Single-chain fatty acids would require divalent cation chelators or mixed lipids to enable heat and pH stability (Adamala and Szostak, 2013; Sarkar, et al., 2020).
Hydroxy acids—a critical component of fatty acids, polyesters, and phospholipids—would have been delivered by meteorites (Lai et al., 2019) or generated via mineral-facilitated reduction of α-ketoacids (Barge et al., 2020), FTT synthesis (see Chapter 4.2.3.5), the formose reaction (see Chapter 4.2.3.6), or Miller-Urey chemistry (see Chapter 4.2.3.10). Glycerol, a key linker between the fatty acid and phosphate group, could have been formed by the formose reaction, meteoritic delivery, impact events, or synthesis in interstellar ice (Kaiser et al., 2015; Zellner et al., 2020). Phosphorylation of glycerol to form potential phospholipid precursors has also been demonstrated in eutectic or alternative solvents (Dalai and Sahai, 2019).
Liquid-liquid phase separation may be an important aspect of primitive compartmentalization given that it is critical for membraneless organelles and is an important process in the regulation and function of modern cells (e.g., chromatin; Yoshizawa et al., 2020). A variety of membraneless compartments generated from phase separation have been investigated including coacervates (i.e., aqueous phase colloids enriched in molecules/ions; Frankel et al., 2016), aqueous two-phase systems (Cakmak and Keating, 2017), polyester (polymers of hydroxy acids) microdroplets (Jia et al., 2019), and supercritical carbon dioxide (Mayer et al., 2017). Geological structures on early Earth could also have served as precursor membranes. For example, porous cavities in rocks at hydrothermal sites could facilitate thermophoresis (i.e., transportation of molecules driven by a temperature gradient), or synthesis and accumulation of genetic materials (Kreysing et al., 2015). Alternatively, iron-sulfide bubbles precipitated at submarine vents or mineral sheets could have concentrated organics and served as a source of electrical (redox) or mechanical energy for driving prebiotic reactions, respectively (Russell and Hall, 1997; Hansma, 2013).
The origin of homochirality
Chirality is a property of asymmetry observed in some molecules, including those used in biology. Enantiomers are chiral compounds that have the same chemical formula and connectivity but are mirror structures and are non-superposable, meaning they cannot precisely overlap one another (see Chapter 2.2.3.5). Why and how modern life ended up choosing right-handed sugars and left-handed amino acids instead of their mirror counterparts remains an active area of research (Blackmond, 2019). Rather than being a requirement for abiogenesis, it is possible that the tendency for life to utilize one enantiomer over another originated at a later stage of (proto)life's development. While mixed chirality polymers are not as functional, nor in some cases as stable (Brack, 1987), both enantiomers could have served different functions for primitive life. On the other hand, too much enantiomeric diversity could lead to enantiomeric inhibition impeding polymerization and enzyme activity (Schmidt et al., 1997; Maury, 2000).
Amino acids and sugars with a slight excess of the L and D enantiomers, respectively, have been found in primitive meteorites, leading some researchers to suggest an astrophysical origin for homochirality (Chapter 3.3.5). Alternatively, terrestrial mechanisms for promoting homochirality have been demonstrated. For example, D- and L-amino acids preferentially catalyze the synthesis of enantiomeric sugars having an excess of L and D isomers, respectively (Córdova et al., 2005; Sadagopan, 2019). Similarly, D-pentose sugars promote the synthesis of L-amino acids (Wagner et al., 2017). Enantiomeric excess could have also been enhanced due to differences in the solubility between crystals of pure and mixed enantiomers generated during a chemical reaction (Breslow and Cheng, 2009; Hazen and Sverjensky, 2010; Blackmond, 2019). Selective adsorption of organics onto a chiral crystal surface (e.g., quartz, specific calcite faces) can also enhance enantiomeric excess (for a review, see Lee et al., 2022).
The origin of metabolism
Metabolism is an essential feature of biology wherein life extracts energy from carbon compounds to generate and store energy, typically via the production of ATP (Chapter 6.2.1). In aerobic organisms, the most important energy-generating pathway is the citric acid cycle. In anaerobes, it is the Wood–Ljungdahl pathway and fermentation. Each cycle generates ATP by breaking down pyruvate. These metabolic cycles, and thus pyruvate, are thought to be ancient as they are present throughout the Tree of Life (Camprubi et al., 2017; Muchowska et al., 2017). On a prebiotic Earth, protometabolic reactions could have been catalyzed by proto-enzymes, which can be grouped as either a “pre-protein” or a “non-protein” type.
Pre-protein catalysts
Pre-protein catalysts are molecules that are critical to the function of modern-day proteins that are hypothesized to have been capable of catalyzing reactions alone, without their protein envelope, on prebiotic Earth. If true, pre-protein catalysts were encased within a protein only later in the evolution of life, perhaps to ensure the catalyst functioned despite environmental fluctuations (White, 1976). The most favored pre-protein catalysts are cofactors (i.e., a molecule that binds to and activates proteins) such as nicotinamide adenine dinucleotide (NAD+; Fig. 4.8C) or metallic clusters (such as the Fe-S clusters in metalloproteins; Bonfio et al., 2018; Sanden et al., 2020). Researchers have postulated that enzymatic Fe-S clusters are an ancient remnant of the earliest life-forms that relied on Fe-S minerals to catalyze reactions (i.e., the Iron-Sulfur World hypothesis; Wächtershäuser, 1990; Goldford et al., 2017).
Non-protein catalysts
Non-protein catalysts are catalysts unrelated to the current protein/enzyme used (e.g., a mineral or a metal; Lazcano and Miller, 1999). Metals and minerals have been shown to facilitate various metabolically relevant reactions including CO2 reduction, reductive amination, and steps of the reverse citric acid cycle (Muchowska et al., 2017; Barge et al., 2020; Weber et al., 2022). Per the Iron-Sulfur World hypothesis, iron-sulfur minerals are predicted to have been critical for abiogenesis given their abundance on early Earth and structural similarity to Fe-S clusters, which are ubiquitous in life (Wächtershäuser, 1990; Nitschke et al., 2013; Bonfio et al., 2018). Researchers have also reported the use of metal catalysis for protometabolic reactions under conditions simulating alkaline hydrothermal vents (Kitadai et al., 2019).
Protometabolic cycles and networks
Chemical reactions in isolation would not have been sufficient to facilitate the jump from chemistry to life. Indeed, life sustains itself via a series of linked chemical reactions where the product of the first reaction is the starting material for the second, and so on until the desired product (i.e., ATP) is generated (Fig. 4.11). On early Earth, a series of linked reactions could have been considered a protometabolic cycle. Research has demonstrated that large metabolic networks can be generated from 10 enzymatic functions, suggesting that the formation of a few unspecific catalysts would have been sufficient for producing protometabolic cycles/networks, consisting of many linked reactions (Goldman et al., 2012).
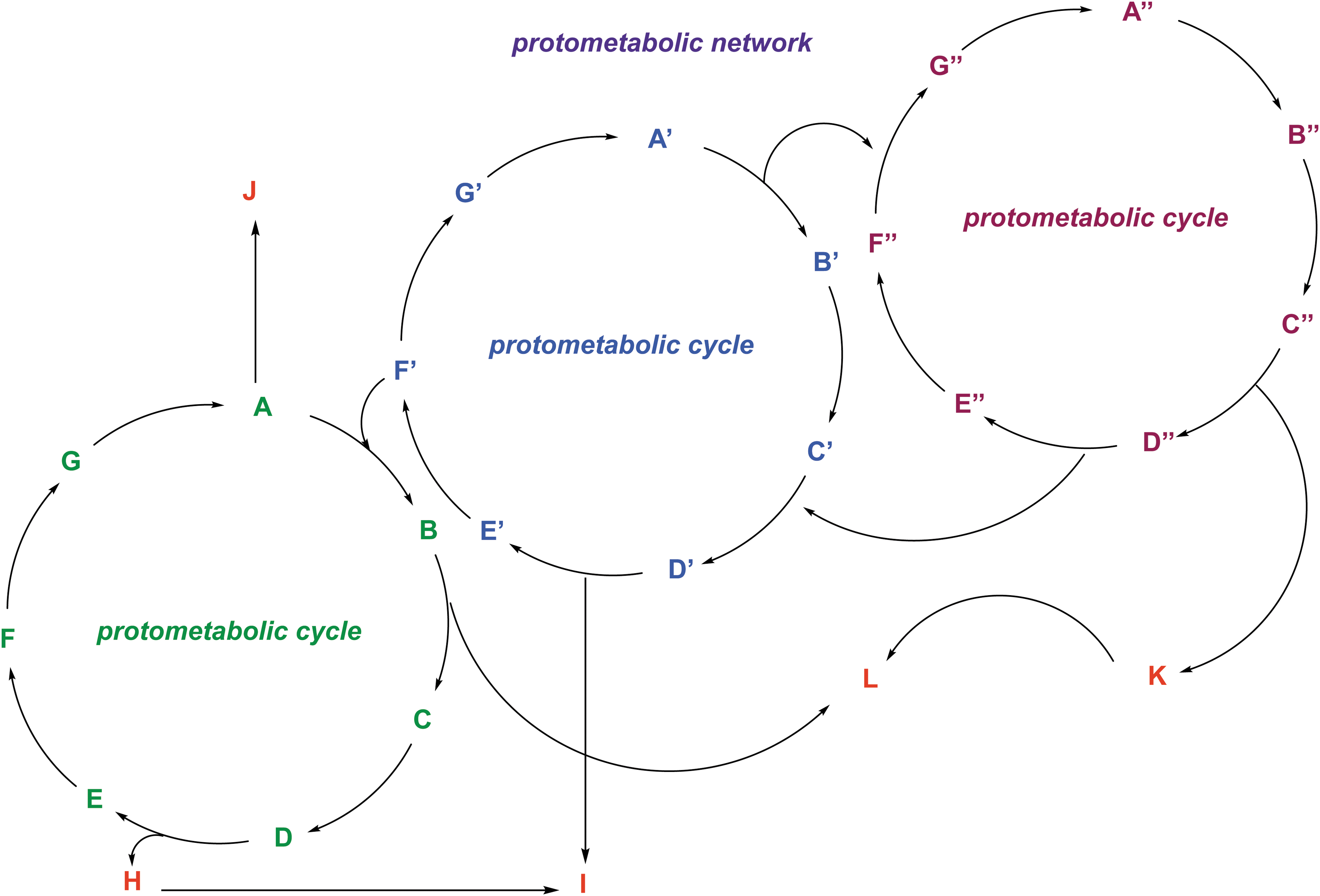
An example of a hypothetical protometabolic network composed of three protometabolic cycles (green, blue, purple). A protometabolic cycle consists of two or more reactions whose products are designated by letters and which feed into the subsequent reaction. The cycles form a network when their end products (i.e., the red letters) feed forward to a different metabolic cycle.
While protometabolic cycles could have facilitated the formation of organics needed to sustain/initiate life, such prebiotic reactions were probably inefficient with low product yields. Low yields are prohibitive toward forming more complex, larger organic molecules and sustaining protometabolic cycles, as both rely on the products generated via multiple inefficient reactions. One proposed solution is autocatalytic cycles (Preiner et al., 2019). Autocatalysis is the catalysis of a reaction by one of its products, creating a positive feedback loop. Some examples of autocatalytic reactions include the Soai reaction (which leads to nearly homochiral products; Soai et al., 1995) as well as FTT and formose reactions.
Like protometabolic cycles, autocatalytic cycles can be combined to form complex networks. If the network can be sustained via a set of ambient “food,” the network can be referred to as a “reflexively autocatalytic food-generated” (RAF) set. To fit this definition, the system must be catalytically closed (i.e., all of the reactions are able to react within the system) and self-sustaining (Hordjick et al., 2019).
Environmental Considerations for the Origins of Life on Earth
Astrobiologists have different perspectives on the specific environmental conditions necessary for abiogenesis. Some important constraints to consider include salinity, temperature, pH, and organometallic interactions. This section gives an overview of how these geochemical parameters could have impacted prebiotic chemistry (Chapter 4.3.1) and the environments on early Earth where the geophysical conditions could be conducive to the origins of life (Chapter 4.3.2).
Conditions conducive to prebiotic chemistry and the origins of life
The presence of salts
Salts can form complexes with organics, facilitating reactions and even promoting selectivity in complex reactions (e.g., formation of cis-diols like ribose in the formose reaction; Chapter 4.2.3.6). By coordinating with water, salts also lower the water activity of an aqueous solution, promoting polymerization (see Chapter 4.2.4.3). However, dissolved ions could also have introduced new hurdles to the origins of life: cations could inhibit phosphorylation (see Chapter 4.2.2.2) and vesicle formation by chelating with phosphates [HPO4 2-] and forming soaps/crystal aggregates with carboxylates [R-COO-], respectively (Milshteyn et al., 2018). Although cations disrupt the formation of some membrane precursors (Apel and Deamer, 2005), high salt concentrations do not disrupt vesicle formation from mixtures of amphiphiles (e.g., fatty acids, hydroxy acids; Jordan et al., 2019).
Suggestions that the ionic concentration of modern cells reflects the chemical composition of the environment in which life first evolved argue against marine environments for abiogenesis (Mulkidjanian et al., 2012). This argument is based on two related observations: (i) regulating intracellular salinity (i.e., osmoregulation) requires complex biomolecules, and (ii) intracellular trends of K+ (100 mM) and Na+ (10 mM) are opposite to that of marine waters, which have about 40 times as much Na+ (460 mM) than K+ (10 mM). If protocells had similar Na+ and K+ as modern-day cells, protocells in a marine environment would have had to constantly expend energy to maintain their Na+ and K+ concentrations from equalizing with surrounding waters. Moreover, if salinity was not tightly regulated, fluctuations in the environment could have caused cells to rapidly shrink or expand and rupture. Environments more closely aligned with cellular composition include surficial hot springs where K+ concentrations can exceed that of Na+ (Mulkidjanian et al., 2012).
High versus low temperatures
Elevated temperatures can increase reaction yields and rates of prebiotic reactions (e.g., formose and FTT reactions; see Chapter 4.2.3). Early Earth's surface was relatively hot following primary accretion and continued bombardment from impactors (see Chapter 3.4.3.1 and Chapter 4.1.1), and genomic evidence suggests some of the earliest organisms may have been thermophiles, although this claim remains controversial (Forterre, 2015).
Higher temperatures can also promote organic degradation and induce nonproductive condensation reactions, which together could have driven the formation of insoluble, unreactive tar. Lower temperatures better preserve organic molecules and promote condensation reactions and the catalytic activity of proto-enzymes (Attwater et al., 2010). As ice crystals grow, they tend to exclude dissolved impurities (e.g., organics or salts) via a process known as eutectic freezing. The residual eutectic fluids are significantly enriched in dissolved compounds that can facilitate organic synthesis (Menor-Salván and Marín-Yaseli, 2012).
Acidic versus basic pH
Mildly acidic conditions could have been beneficial for the origins of life given that under such conditions (i) the RNA phosphodiester bond is most stable at pH 4–5, (ii) peptide bonds are most stable at pH 4–7.5, with pH 6 being optimum, (iii) a plethora of isolated ribozymes are more active, (iv) RNA polymerization via wet/dry cycles is more efficient, and (v) phosphate minerals slowly dissolve, releasing phosphate (Bernhardt and Tate, 2012). Alternatively, alkaline conditions may have been important for other aspects of abiogenesis, as many other prebiotically important reactions are favored in alkaline solutions (e.g., the formose reaction and HCN polymerization; see Chapter 4.2.3).
Importance of minerals
Water on Earth has been exposed to a rocky surface throughout geologic history. Thus, minerals were present for prebiotic reactions and may have even been critical for abiogenesis. Indeed, minerals can promote organic preservation, facilitate reactions and polymerization (e.g., Ferris 2005; Kitadai et al., 2017), enhance enantiomeric excess via their inherent chirality (see Chapter 4.2.5), and concentrate otherwise dilute reactants onto a catalytic surface (Salman et al., 2012; Alshameri et al., 2018; Flores et al., 2021).
While adsorption can promote reactions, it limits the mobility of organics, which inhibits their reactivity in solution. For example, when organic material was added to a hot spring pool, it quickly adsorbed onto clays and never became encapsulated into the dissolved lipid vesicles (Deamer et al., 2006). Importantly, in a fluctuating environment, geochemical changes could lead to the rapid release of adsorbed species (Ruiz-Bermejo et al., 2013).
Environments proposed as sites for abiogenesis
Submarine hydrothermal vents
Several types of hydrothermal vents could have existed on early Earth, including black smoker vents (high temperatures of 350–400°C, acidic with pH 2–5, metal and sulfide rich), white smoker vents (similar temperatures and pH as black smokers, Zn-enriched, Fe and sulfide depleted), diffuse vents (moderate temperatures of 50–200°C, acidic to alkaline, metal depleted), and alkaline vents (mild temperatures of 40–75°C, alkaline with pH 10–12, sulfide/metal depleted). Acidic vents can form wherever a magma chamber is near oceanic sediments. Typically, this formation can occur at mid-ocean ridges where seafloor spreading or subduction zones develop, but they can also form at volcanic hot spots (e.g., at Lōʻihi Seamount). Alkaline vents can form wherever ongoing serpentinization of ultramafic rock or (to a lesser extent) mafic rock takes place (see Chapter 4.2.3.1). While most hydrothermal vents form in deeper waters (typically thousands of meters), they have also been observed in shallow-sea settings (5–212 m depth), where there is sufficient sunlight for photosynthesis (Gilhooly et al., 2014; Price and Giovannelli, 2017). For a review on hydrothermal vent chemistry, see Kelley et al. (2002) and Tivey (2015).
As the interior of Hadean Earth was hotter, the young planet is thought to have been more geologically active, thus promoting the formation of acidic hydrothermal vents at the seafloor. Alkaline vents were also likely widespread during the Hadean since the crust and mantle would have been less differentiated, resulting in an ultramafic Hadean seafloor that could more readily undergo serpentinization (Sleep et al., 2004). Although acidic hydrothermal vents have several useful properties for abiogenesis, the cooler alkaline vents have since gained favor in hypotheses concerning the origins of life on Earth as they have (i) longer lifetimes (up to 30,000 years; Früh-Green et al., 2003), (ii) higher concentrations of H2 (Russell et al., 2014), (iii) alkaline pH and high concentrations of Ca2+ and Mg2+, which can facilitate the formose reaction (Kopetzki and Antonietti, 2011), and (iv) cooler temperatures, normally within the range of 40–75°C (Kelley et al., 2002).
Hydrothermal vents display several characteristics that could have been conducive for the emergence of life because they have (i) porous structures, which provided containment of reactants in a manner analogous to a protocell, (ii) pH gradients (Δ pH = 3–5) that can drive reactions and are key to metabolism (Sojo et al., 2016), (iii) thermal gradients which have been shown to promote the replication of DNA (Mast and Braun, 2010), and (iv) proliferation of catalytic metals/minerals that can drive organic synthesis (Barge et al., 2020). However, there are several limitations to submarine hydrothermal vents as settings for the origins of life such as the following: (i) wet/dry cycling (which promotes condensation reactions) is not possible, although this may not have been an issue for shallow vents (Barge and Price, 2022) or if condensation reactions proceeded via different mechanisms (see Chapter 4.2.4.3); (ii) for alkaline vents, a combination of high pH and elevated Ca2+ and Mg2+ concentrations favor the formation of insoluble phosphate minerals and destabilize RNA (Bernhardt and Tate, 2012); and (iii) regulating against high salinity may have been difficult for any protocell (see Chapter 4.3.1.1).
Deep subsurface beneath oceanic crust
The deep subsurface (i.e., kilometers beneath the Hadean crust) may have contained an abundance of H2 formed via serpentinization, mineral catalysts, and elevated temperatures. An origin of life in the deep subsurface (Gold, 1992) would have benefited from incredibly stable conditions (remaining unchanged for millions of years) compared to the more ephemeral hot springs or hydrothermal vents. In fact, the deep crust may have been the only early Earth environment protected from impact events (Maher and Stevenson, 1988; Grimm and Marchi, 2018). Additionally, the deep subsurface was ubiquitous on early Earth with organics that, unlike those produced in the bulk ocean, could be more easily concentrated. However, complex chemical systems, gradients, and mixing, which are critical for establishing metabolism and driving chemical reactions away from equilibrium (see Chapter 4.2.1), would have been hindered in the deep subsurface. Additionally, the deep subsurface would have had limited access to organics brought by exogenous sources or generated at the surface.
Terrestrial hot springs
Terrestrial hot springs were first broadly proposed as a potential site for the origins of life by Charles Darwin and have been widely considered since (see Damer and Deamer, 2020 and references therein). Importantly, hot springs differ from deep sea hydrothermal vents in several ways as they (i) act as an interface between atmosphere/minerals/fluids and allow for wet/dry cycling (Damer and Deamer, 2015); (ii) concentrate and make available important prebiotic minerals (borates, nitrates, and phosphates) through water/rock interactions, evaporation, wet-dry cycling, or low pH (Burcar et al., 2016; Ranjan et al., 2019; Steller et al., 2019); (iii) have an abundance of clay minerals that can concentrate and polymerize biologically relevant monomers for the origins of life (Ferris, 2005; Hazen and Sverjensky, 2010); (iv) are exposed to solar and cosmic radiation that may aid in prebiotic synthesis (Ranjan and Sasselov, 2016); and (v) exhibit steep and complex chemical/temperature/pH gradients through mixing zones, thus forming different aqueous bodies with unique chemistries and creating “pools of innovation” to foster the origins of life (Van Kranendonk et al., 2017, 2021). However, some of these benefits may also be detrimental because (i) the low pH may destroy polymers by hydrolyzing bonds (Mungi and Rajamani et al., 2015), (ii) clays could adsorb organics and remove them from solution, therefore impeding reactions (Deamer et al., 2006), and (iii) solar and cosmic radiation could destroy organic molecules (Phoenix et al., 2006; Farr et al., 2023).
Hot spring pools may have been present on early Earth as evidenced by the discovery of geyserite (a mineral only produced in surficial hot springs) interbedded within the 3.5 Ga stromatolite fossils of the Pilbara Craton, Western Australia (Djokic et al., 2017). Further evidence of terrestrial hydrothermal pools stem from ∼4.3 Ga zircons from Jack Hills, Australia, whose rare earth elements and isotopic content suggest that these terrestrial pools may have been fed by relatively oxidized fluids that facilitate the dissolution of metals (e.g., Fe2+, Fe3+, Mn2+, and Zn2+) that can drive prebiotic chemistry (Trail and McCollom, 2023). In contrast to hot springs of today which exist over a granitic/felsic crust that is relatively depleted in Fe/Mg and enriched in silica and Na/K (Fig. 4.1), Hadean hot springs over a basaltic bedrock, such as those on Hawaii or Iceland, would have been enriched in magnesium and iron.
Other environments
Other environmental niches have been proposed as containing favorable components for the origins of life on Earth, including the shallow subsurface sedimentary environment (Westall et al., 2018). Pumice rafts and microlayers of organic “oily slicks” on the ocean surface could have accumulated suspended mineral grains and served as havens for origins-of-life chemistry (Brasier et al., 2011). Shallow pools on exposed land masses (e.g., basaltic islands) may have formed from rainfall; these pools may have promoted condensation reactions via repeated aerosol formation from splashing (Ruiz-Bermejo et al., 2007; Nam et al., 2017) or formation of semi-anhydrous solvents upon significant evaporation (Burcar et al., 2016). Furthermore, tide pools could have accumulated radioactive detrital grains, which may have aided in prebiotic reactions (Adam, 2007) or concentrated organics upon freezing (see Chapter 4.3.1.2).
It is important to remember that these systems do not work in isolation, and there is a degree of mixing that creates unique and complex settings (Stüeken et al., 2013). Deep sea hydrothermal vents may appear as isolated systems, but clay minerals can transport organics concentrated on the ocean's surface to the seafloor (Kennedy et al., 2002). Thus, multiple environmental combinations may have been required to facilitate the emergence of life. Moreover, different environments could have hosted their own genesis events (i.e., life might have arisen multiple times on early Earth).
Conclusions: From Prebiotic Chemistry to the Formation of a Protocell
Although the division between life and nonlife is blurry (see Chapter 2.3.2), one possible intermediate structure may have been the assembly of a primitive cell-like compartment called a protocell. A protocell is, by definition, a self-sustaining compartment that could have accomplished a number of essential functions including genetic replication, division, and metabolism. In essence, a protocell may have simply been a primitive compartment containing a replicating genetic molecule and catalytic molecules to perform metabolism, with the ability to grow and divide through external stressors or environmental changes. A number of potential protocell models have been proposed, including fatty-acid bilayer vesicles or membraneless droplets (Martin, 2019; Chandru et al., 2020). However, there is no specific agreed-upon protocell definition or structure (Lopez and Fiore, 2019). Notably, a protocell is distinct from a primitive prebiotic compartment in that it was a direct precursor to modern cells, with artifacts of its existence found in modern biology.
The process from which a protocell evolved into a modern cell was likely facilitated by selective chemical evolution, a subset of Darwinian evolution (Higgs, 2017). Selective chemical evolution of any population results in accumulation of only a subset of the initial population which possesses greater survivability/persistence either through a more stable structure or an efficient/effective function that improves its own survivability (Ruiz-Mirazo et al., 2004). In the laboratory, selective chemical evolution has been observed on various systems, ranging from primitive chemicals and polymers (Forsythe et al., 2015; Mamajanov, 2019), to modern biopolymers (Tagami et al., 2017; Kroiss et al., 2019), to populations of compartments (Bansho et al., 2016; Mizuuchi and Ichihashi, 2020). On primitive Earth, the environment would have supplied selective pressures, perhaps including fluctuating temperatures, mineral surfaces, radiation from sunlight or radioactive elements in Earth, changes in salinity/pH, or even physical processes such as oceanic waves or emission from fumaroles (Maruyama et al., 2019). These processes could have resulted in the emergence of protocells that (i) contained structures that were more robust and persistent to environmental perturbations (Cakmak et al., 2020; Sarkar et al., 2020), (ii) contained catalytic polymers that could efficiently synthesize important reactants or catalyze essential reactions (Müller, 2009; Fares et al., 2020), (iii) could self-assemble into novel structures capable of adsorbing essential molecules or facilitating essential chemical reactions (Tena-Solsona et al., 2018; Jia and Fraccia, 2020), or (iv) might have itself evolved at a greater rate than previous iterations (Matsumura et al., 2016). As such, more investigation into protocells or primitive compartments with these characteristics, or even novel emergent characteristics, is warranted to understand how prebiotic chemistry evolved into cellular biology (see Jia et al., 2021 for a detailed discussion).
As Darwinian evolution is a process that occurs across a number of timescales and on different types of systems, including the population level in modern biology (see Chapter 5 for a discussion on the evolution of life on Earth), it is likely that chemical evolution drove the selection of a specific subset of the entire population of potential primitive chemicals on early Earth, favoring those that could either (i) assemble and function within a protocellular structure or (ii) catalyze or participate in essential chemical reactions. Constant changes of the geological environment likely resulted in the degradation of functionally evolved species (Chandru et al., 2020). These changes may have eliminated several primitive chemicals or structures for which there is no biological record in modern life. Thus, further prebiotic chemistry research must also equally focus on molecular species that are no longer present in modern biology.
Footnotes
Acknowledgments
LPI Contribution No. 3004. LPI is operated by USRA under a cooperative agreement with the Science Mission Directorate of the National Aeronautics and Space Administration. The authors would like to thank the lead editors of the Astrobiology Primer 3.0, Dr. Micah Schaible and Dr. Nadia Szeinbaum, for providing edits and feedback on this manuscript and, along with Dr. George Tan, for organizing and leading efforts related to the Primer as a whole. We would also like to thank Dr. Muammar Mansor (Universität Tübingen), Dr. Martin van Kranendonk (University of New South Wales), Dr. Moran Frenkel-Pinter (Georgia Tech), and Dr. Philip Bevilacqua (Penn State) for their insightful comments and edits which together improved the manuscript. Work by LER, JMW, and NYH was carried out at the Jet Propulsion Laboratory, California Institute of Technology, under a contract with NASA (80NM0018D0004). We would like to thank Dr. Laurie Barge (JPL) for her support and helpful discussions. Work by NYH at Ludwig-Maximilians-Universität München was supported via the CRC 235 Emergence of Life grant. TZJ is faculty at the Earth-Life Science Institute (ELSI) of the Tokyo Institute of Technology, which is sponsored by a grant from the Japan Ministry of Education, Culture, Sports, Science and Technology (MEXT) as part of the World Premier International Research Center Initiative (WPI). TZJ is further supported by Mizuho Foundation for the Promotion of Sciences, Japan Society for the Promotion of Science (JSPS) Grants-in-aid 18K14354 and 21K14746, project grants from the Astrobiology Center Program of Japan National Institutes of Natural Sciences (NINS) (Grant Number AB311021, AB021008), the Tokyo Institute of Technology Yoshinori Ohsumi Fund for Fundamental Research, and by the Assistant Staffing Program by the Gender Equality Section, Diversity Promotion Office, Tokyo Institute of Technology. TZJ would like to acknowledge Dr. Niraja V. Bapat for the initial invitation to contribute to the Astrobiology Primer 3.0, which was partially facilitated by a grant from the ELSI Origins Network, which was supported by a grant from the John Templeton Foundation. The opinions expressed in this publication are those of the authors and do not necessarily reflect the views of the John Templeton Foundation. LHS is supported by the Australia Research Council Discovery Project grant 180103204 awarded to M. Van Kranendonk. LHS would like to acknowledge and pay their respects to the Bedegal people, who are the traditional custodians of the land on which this research took place. © 2023 All rights reserved.
Author Contributions
LER was editor of the chapter, managed the team, and developed the structure of the chapter. All authors contributed to the writing and editing of the entire document.
Associate Editors: Sherry Cady and Christopher McKay