
Abstract
Gypsum (CaSO4·2H2O) has been identified at the surface of Mars, by both orbiters and rovers. Because gypsum mostly forms in the presence of liquid water as an essential element for sustaining microbial life and has a low porosity, which is ideal for preserving organic material, it is a promising target to look for signs of past microbial life. In this article, we studied organic matter preservation within gypsum that precipitates in a salt flat or a so-called coastal sabkha located in Qatar. Sabkha's ecosystem is considered a modern analog to evaporitic environments that may have existed on early Mars. We collected the sediment cores in the areas where gypsum is formed and performed DNA analysis to characterize the community of extremophilic microorganisms that is present at the site of gypsum formation. Subsequently, we applied Raman spectroscopy, a technique available on several rovers that are currently exploring Mars, to evaluate which organic molecules can be detected through the translucent gypsum crystals. We showed that organic material can be encapsulated into evaporitic gypsum and detected via Raman microscopy with simple, straightforward sample preparation. The molecular biology data proved useful for assessing to what extent complex Raman spectra can be linked to the original microbial community, dominated by Halobacteria and methanogenic archaea, providing a reference for a signal that may be detected on Mars.
Introduction
Data obtained from NASA's Mars Exploration Rovers (e.g., Spirit and Opportunity) (Squyres et al., 2004; Schopf et al., 2012; García-Florentino et al., 2021), as well as orbiter missions including the Mars Express (e.g., Gendrin et al., 2005) and Mars Reconnaissance Orbiter (e.g., Murchie et al., 2009) have revealed the unambiguous existence of gypsum on the martian surface. Such gypsum likely formed early in the planet's history, in a period for which evidence suggests that conditions at the surface of the planet may have been less inhospitable than today, and compatible with the existence of microbial life. For example, evidence suggests that during the Noachian era between 4100 and 3700 million years ago (Mya), Mars contained a warmer and wetter climate in part due to the presence of its once prominent magnetic field (Véronique et al., 2012).
However, the planetary transformations during the Hesperian era between ca. 3700 Mya and ca. 3000 Mya, which accompanied the loss of the magnetic field, more than likely stunted the evolution of life, preventing it from further diversifying (Connerney et al., 1999). As the presence of gypsum indicates water, which is essential to life, gypsum's existence on Mars lends support for the potential of life. Although gypsum found on Earth is formed through water evaporation from a diverse number of environments (Schopf et al., 2012), on Mars, gypsum formations are often found in secondary fractures and thought to be associated with acidic diagenetic fluids (Yen et al., 2017; Hausrath et al., 2018). However, martian gypsum fracture veins have also been hypothesized to form under nonacidic conditions and directly compared with gypsum veins observed within sabkha environments (Nachon et al., 2014).
In both scenarios, gypsum-forming fluids would exhibit high salinity, leading to the hypothesis that extant halophilic microorganisms may have existed within highly concentrated brine solutions and minerals in the martian environment (Landis, 2001a, 2001b; DasSarma, 2006; Oren, 2014; Oren et al., 2014; DasSarma et al., 2020). In addition, due to low porosity, gypsum may have offered space for organic molecules of microbial origin to be incorporated into a crystal matrix. The possibility to find organic material within gypsum crystals has significant implications for the detection of life on the Red Planet (Edwards et al., 2006; Bibring et al., 2006). There are findings that microbial remnants can be preserved within sulfate deposits across broad range of geological age, for example, in the form of mesoscale stromatolites a few million years old (Allwood et al., 2013) to permineralized cellular microfossils a few 100 million years (Schopf et al., 2012), as well as particulate kerogen in late Miocene to early Pliocene evaporites (Shkolyar and Farmer, 2018), and biomarkers in modern evaporites (Benison, 2019).
The sabkha environments represent a unique opportunity to gain insight into the production and preservation of organic substances originating from halophilic microorganisms similar to those that might have occurred on Mars. Sabkhas may be analogous to past evaporitic surfaces of Mars (e.g., Nachon et al., 2014) due to their arid climate, elevated concentrations of salt, and increased insolation by low-wavelength radiation. Furthermore, sabkhas host extensive assemblages of evaporitic minerals, including gypsum and carbonates (Brauchli et al., 2016) in close association with abundant and diverse hypersaline microbial communities (Diloreto et al., 2019, 2021). It is notable that organic molecules preserved from these microbial communities are associated with functionality. The function of many of the preserved organic molecules may have been to increase microbial survivability under harsh martian conditions. Consequently, these organic molecules have become the focus of the search for life.
A prominent example is β-carotene that can act as a DNA repair agent for cells damaged from high levels of radiation as well as one of few pigments that possess the ability to effectively absorb UV-C radiation, thereby offering protection against environments with harsh UV conditions (Vítek et al., 2009a, 2009b). The detection of this biomolecule has been given a high priority on the European Space Agency's ExoMars missions as an indicator of past organic life (Vago et al., 2006). Past work within sabkha environments has highlighted the presence of organic molecules with astrobiological significance, such as carotenoids, chlorophyll, and scytonemin (Edwards et al., 2006; Stivaletta et al., 2009; Edwards et al., 2010). However, despite the detection of the organic compounds, little work has been performed on the connection between their detection and the microbial ecology responsible for producing them, as well as how those relationships pertain to extant microbial life.
Thus, this study aimed to relate organic molecules trapped within gypsum, and microbial communities at the locations where gypsum is found. In addition, this work aimed to extrapolate what influence this might have on the evaluation of organic molecules that might be found on Mars. Consequently, our objectives were twofold; we aimed to identify dominant species comprising the microbial community present within the gypsum layer at 25 cm depth in a sabkha and to determine if any signature of the microbial community, especially organic molecules was detectable, within the sabkha's gypsum using Raman microscopy.
Raman microscopy was specifically used because the NASA Rover Perseverance (currently active on Mars) as well as the ready-to-be-launched ExoMars rover are equipped with a Raman spectrometer (e.g., Lalla et al., 2022; Lopes-Reyes et al., 2022). The combination of in situ and ex situ techniques employed in this study allows for both the identification of likely candidates for organic material, as well as evaluation of environmental microbial dynamics. Our ultimate goal is to understand the unknown processes that form and trap organic material associated with gypsum in sabkha environments, and implication of the findings for a martian environment.
Materials and Methods
Study site
Gypsum samples were collected from the Dohat Faishakh sabkha in Qatar at a latitude of 25.63682° and a longitude of 50.960533°, shown in Fig. 1, and were extracted from a depth of 25 cm from a core referred to as Dohat Faishakh sabkha (DF). The Dohat Faishakh sabkha is located along the northwestern coast of Qatar, in the northeastern coast of the Arabian Peninsula. The Dohat Faishakh consists of a lagoon, an intertidal zone, and a supratidal zone, which is the sabkha. The lenticular gypsum crystals were found in a subhorizontal orientation within the sediment of the sabkha. These crystals are a component of a buried microbial mat and were found just above blue gray dolomite mud (Fig. 2).

Sampling site location and its coordinates in the Dohat Faishakh sabkha in Qatar. Credit Google Earth 2023.

The lenticular gypsum crystals were isolated from the 25 cm layer of the cores and were found to be elongated along their crystallographic ac-plane and ranged in length from 5 to 7 cm, as shown in Fig. 2. The samples were stored at 4°C after their collection during fieldwork in the sabkha. In the laboratory, the gypsum samples were stored at −20°C until preparation, in which the crystals' surfaces were sterilized to remove any organic material not contained within the gypsum.
To sterilize gypsum crystals, they were rinsed with de-ionized water and scrubbed to remove large patches of mud and debris. Once physically clean, the crystals were submerged into 0.5 M hydrocloric acid (HCl) for 15 min to force the neutralization of any surface contaminant, which was basic in nature. The crystals were then successively submerged for a period of 1 min in 2% (weight per volume [w/v]) sodium dodecyl sulfate (SDS) (CAS: 151-21-3; Sigma-Aldrich), 0.02% (w/v) taurocholic acid (TA) (CAS: 345909-26-4; Sigma-Aldrich), and de-ionized water. Following this, the crystals were dried by air. This procedure was adapted from Rosenzweig et al. (2000) using a lower concentration of HCl to prevent dissolution of gypsum crystals.
To ensure that no mass loss occurred when submerging the crystals into the 0.5 M HCl sample, weights were measured before and after. SDS was used to denature proteins, which could be present on the surface of the sample, whereas TA was used to solubilize any fats present on the surface of the crystals. Sterilization of the crystals with HCl and SDS was preformed to ensure that any Raman signal detected during the analysis was a direct result of molecules present in the crystal matrix, as opposed to those present on the crystal surface or from any possible contamination. All the samples were then stored in an opaque sterile bag at −20°C. Gypsum samples were fractured along cleavage plans to not only create a fresh interior surface for analysis but also to ensure that samples were of appropriate thickness (<5 mm) for Raman analysis.
Pigment extraction
Up to 10 g of sample was isolated from 22.5 to 25.5 cm from cores extracted from the sabkha (Fig. 2) for pigment extraction. We considered the core layer at 22.5 cm as a reference layer, which has no gypsum crystals. In addition, we extracted pigments from two samples from microbial mats from the Khor Al-Adaid sabkha at the South of Qatar (green and orange sample labels) dominated by aerobic and anaerobic phototrophs, respectively (Diloreto et al., 2019, 2021). Pigments were extracted to act as an internal reference of potential biomarkers and provide a basis for comparison to any signals detected within gypsum.
Samples were collected in the field and stored at −80°C until pigment extraction could be performed. Pigment extraction via sonication was executed using an extraction solution of 7:2 v/v mixture of acetone/methanol for 15 min to efficiently extract and prevent damage of common pigments, such as chlorophyll and carotenoids (e.g., Ruivo et al., 2014). After extraction, samples were filtered through 0.2 μm polytetrafluoroethylene syringe filters and stored at −20°C in the dark until analyzed. For Raman analysis, pigment samples were pipetted onto a glass microslide and air-dried in the dark.
Microbial community analysis
Up to 5 g of sample was isolated from depths of 5, 7, 22.5, and 25.5 cm from the DF core and a replicate core extracted from the sabkha (Fig. 2) for amplicon sequencing of the 16S rRNA gene. Samples were directly collected in the field and stored in individual microcentrifuge tubes and brought back to the laboratory where they were kept at −80°C until DNA extraction could be performed. DNA extraction was performed using a MoBio powerbiofilm DNA isolation kit as per the manufacturer's instructions. DNA was stored at −20°C until sequenced. 16S rRNA amplicon sequencing of the V4 variable used a combined bacterial and archaeal primer set. This consisted of the 341F forward primer (CCTACGGGNGGCWGCAG) and 785R reverse primer (GACTACHVGGGTATCTAATCC). Polymerase chain reaction (PCR) was performed using the HotStarTaq Plus Master Mix Kit (Qiagen) under the following conditions: 94°C for 3 min, followed by 28 cycles of 94°C for 30 s, 53°C for 40 s, and 72°C for 1 min, followed by a final elongation step at 72°C for 5 min.
Post-amplification PCR products quality checked using a 2% agarose gel, which was pooled and then purified using AMPure XP Beads. Amplicon sequencing was performed using a combination of Quantitative Insights into Microbial Ecology (QIIME) (Caporaso et al., 2010) and Usearch version 10.0 (Edgar, 2010). Analyzed sequences were joined and demultiplexed in QIIME due to file size. Primers were stripped and sequences <200 base pairs were removed using Usearch. Sequences were also filtered and dereplicated using Usearch. To assign operational taxonomic units (OTUs), dereplicated sequences were denoised using the UNOISE algorithm (Edgar, 2016a) and clustered at 100% similarity creating zero distance operational taxonomic units (ZOTUs) (Edgar, 2018). Taxonomy of assigned ZOTUs was classified using the SINTAX algorithm (Edgar, 2016b) with RDPII database as a reference.
Raman microscopy analysis
Sample analysis was performed using an NTEGRA Spectra system (NT-MDT, Zelenograd/Moscow, Russia) coupled with a Solar TII spectrometer. Analysis was carried out in the upright confocal configuration using a 532 nm laser at 8.7 mV. Spectra were collected with 100 × objective and 0.7 numerical aperture with a spectral resolution of 1.4 cm−1 for 600 lines/mm grating using a high-resolution charged-coupled device (CCD) ANDOR Newton 970 CCD camera with a resolution of 1600 × 200 pixels cooled to −70°C. Imaging areas of interest was performed using a photomultiplier tube. The instrument was calibrated using a Silicon-wafer standard, and the Si-peak is located at wavelength of 520 cm−1. All measurements were performed under ambient conditions. Areas of interest among several gypsum crystals were subjected to two-dimensional (2D) mapping.
Spectra were accumulated from 30 to 200 s to keep an intensity of peaks in the range between 30,000 and 50,000 arbitrary units. We used reference materials of β-carotene (synthetic β-carotene, CAS: 7235-40-7; Sigma-Aldrich) and gypsum (RUFF ID: R040029) as a comparison for the analysis. As some mat materials underwent extraction of bacterial pigments, spectra of the air-dried acetone:methanol extract were collected as a reference. Finally, we extracted pigments from collected microbial mats with documented gypsum and a known microbial community from two layers (e.g., Diloreto et al., 2019, 2021). One was a green-colored hypersaline mat (green) dominated by cyanobacteria, whereas the other orange-colored mat (orange) was dominated by anoxygenic phototrophs (Diloreto et al., 2021)
Raman images were processed using the microscope's native software, NT-MDT NOVA, while Raman spectra were processed using a combination of this software and OriginLab. To remove fluctuations in the 2D Raman maps as a result of interactions between the instruments piezometer and the sample's non-ideal (non-flat) surfaces, a three-point leveling operation was performed. After 2D images were corrected, each region of interest (ROI) in the spectrum of organic material 1 (1007 and 1313 cm−1) and organic material 2 (1007 and 1636 cm−1) was isolated by subtraction of the spectral regions. Once isolated, each ROI was used to create a single image layer, which was overlain to create a final image showing spectral ROIs and their spatial associations within the sample.
For purely spectral data, to gain better signal-to-noise ratio, the spectra were processed using the Savitzky–Golay approach to smooth the spectra data (Luo et al., 2005). For the spectra, the 15 points adjacent to each datapoint were used for the creation of the lines of regression required for the analysis and subsequent smoothing. This method preserves the features of the data, such as peak height and width, most effectively preventing attenuation of the peaks. To assess the veracity of the Raman peaks that had been identified, a fast Fourier transform filter was applied to all data sets to remove high-frequency noise while leaving the true signal unaltered. After filtering Raman spectra, we performed an 8- to 10-point baseline correction and identified peaks using the OriginLab peak analyzer algorithm.
Results
Microbial community composition
The microbial communities above and within gypsum-forming layers, 22.5 and 25.5 cm, were examined at the class level (Fig. 3). At a depth of 22.5 cm, the dominant classes include Actinobacteria (10.67%), Clostridia (10.58%), Cyanobacteria (11.82%), and Halobacteria (25.65%). At a depth of 25.5 cm, the dominant classes include Clostridia (10.25%), Halobacteria (49.92%), and Nitrospira (8.90%). A majority of ZOTUs fall within both layers consist of the class Halobacteria. Replicate samples from gypsum-forming depths show the dominant classes at the 22.5 and 25.5 cm depths to be Methanobacteria (22.03% and 25.17%), Bacteroidia (5.89% and 10.59%), and Flavobacteria (12.71% and 5.41%), respectively. A significant portion of ZOTUs in replicate samples is <1% abundance composing 26.34% of the 22.5 cm replicate and 15.70% of the 22.5 cm replicate.
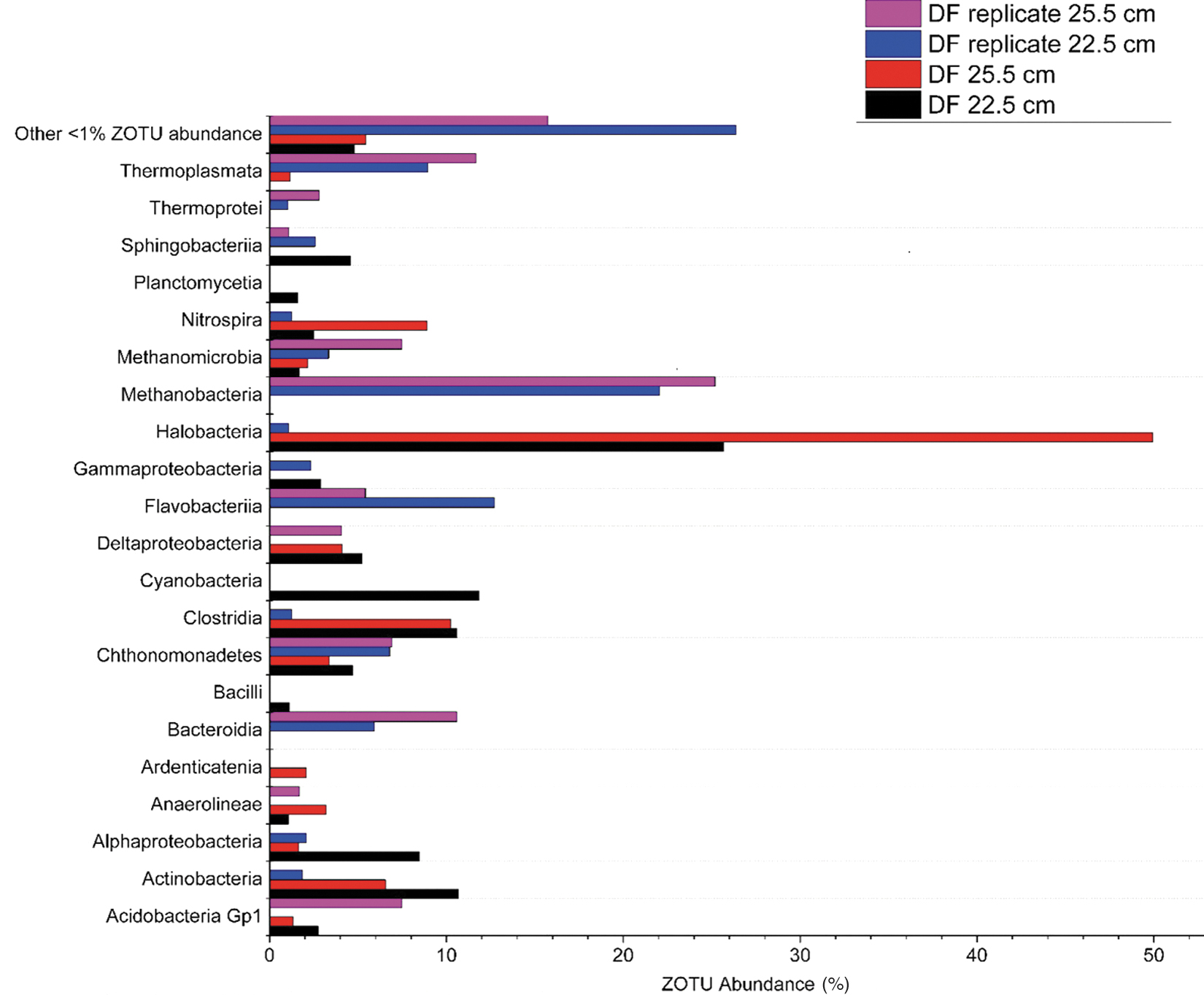
Microbial community composition at the class level at a depth of 22.5 cm, just before the layer the gypsum was extracted from, and microbial community composition at the class level at a depth of 25.5 cm, the layer the gypsum was extracted from with replicate samples.
Two layers at 5 and 7 cm within the DF core and DF core replicate, above the gypsum-forming portion of the core, were also examined at the class level (Fig. 4). Microbial community composition of the DF core at 5 cm shows high abundances of Cytophagia (23.40%), Actinobacteria (9.75%), Alphaproteobacteria (6.03%), and some Halobacteria (6.9%). The replicate sample of 25 cm shows a different composition with a majority of ZOTUs assigned to Bacilli (33.14%), Cyanobacteria (12.21%), and Sphingobacteria (11.09%). Microbial community composition at the 7 cm depth consists of Gammaproteobacteria (22.59%), Alphaproteobacteria (14.65%), Bacilli (14.39%), Halobacteria (7.67%), and Cyanobacteria (10.19%). The microbial community of the replicate sample at this depth is composed of Bacteroidia (14.64%), Gammaproteobacteria (10.89%), Methanobacteria (8.54%), Thermoprotei (9.56%), and Halobacteria (6.62%).
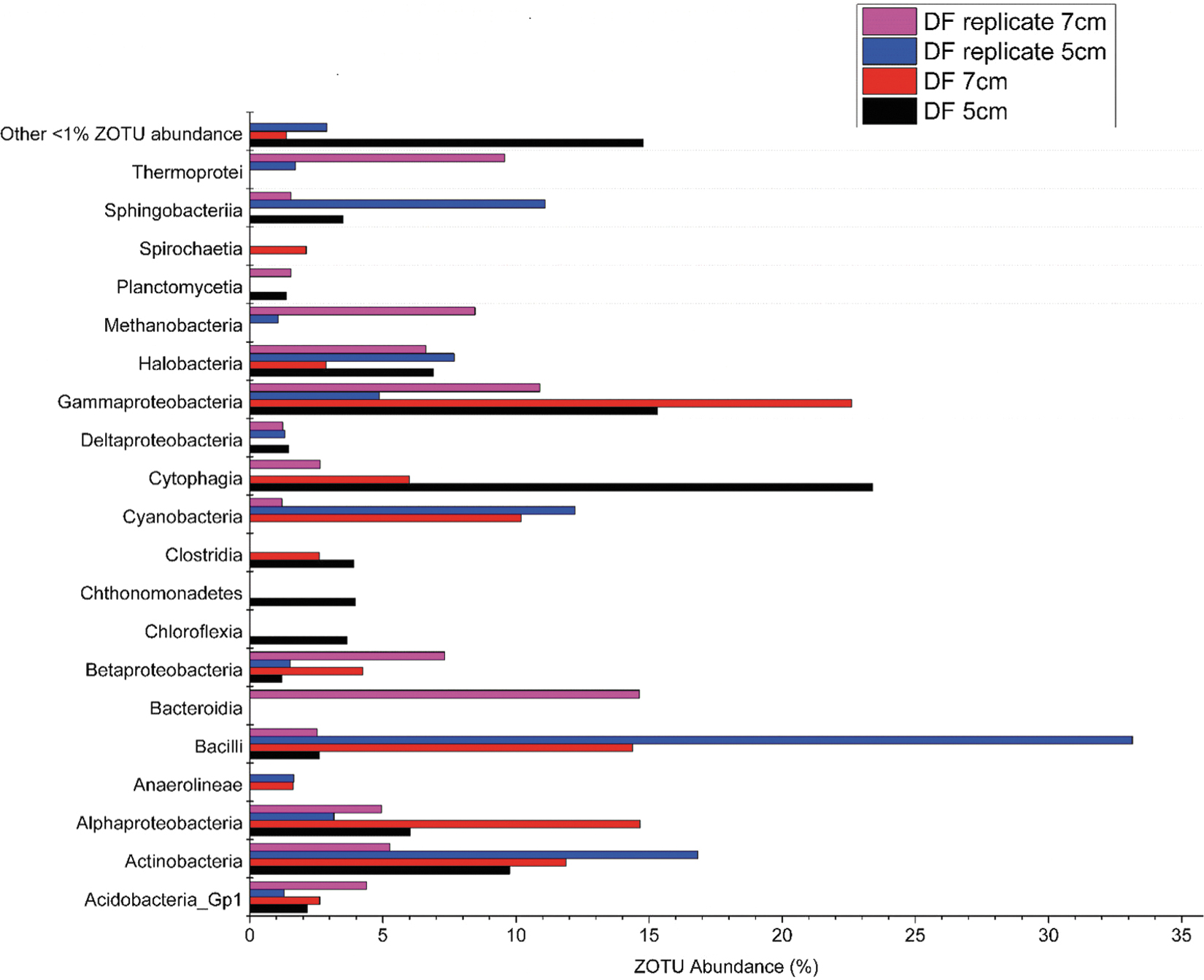
Microbial community composition of upper layers of the DF cores, ≈18–20 cm above the layer the gypsum was extracted from. Microbial community composition is shown at the class level with replicates.
Raman maps show two distinct morphologies of organic material embedded in the gypsum matrix (Fig. 5A, B). The Raman spectra for the sabkha gypsum matrix when compared with a reference show identical characteristic peaks. The gypsum standard (RUFF ID: R040029) shows an identifying δ peak at 1007 cm−1 and a v 3 peak at 1140 cm−1 with lower amplitude v 2 peaks at 492 and 412 cm−1 and a v 4 peak at 620 cm−1 with lower intensity (Fig. 6 and Table 1). The sabkha gypsum exhibits wavelengths at the same peak positions of 412, 492, 620, 1007, and 1140 cm−1. Observed organic material exhibits a variety of peaks that do not coincide with any of the characteristic gypsum peaks (Fig. 6).

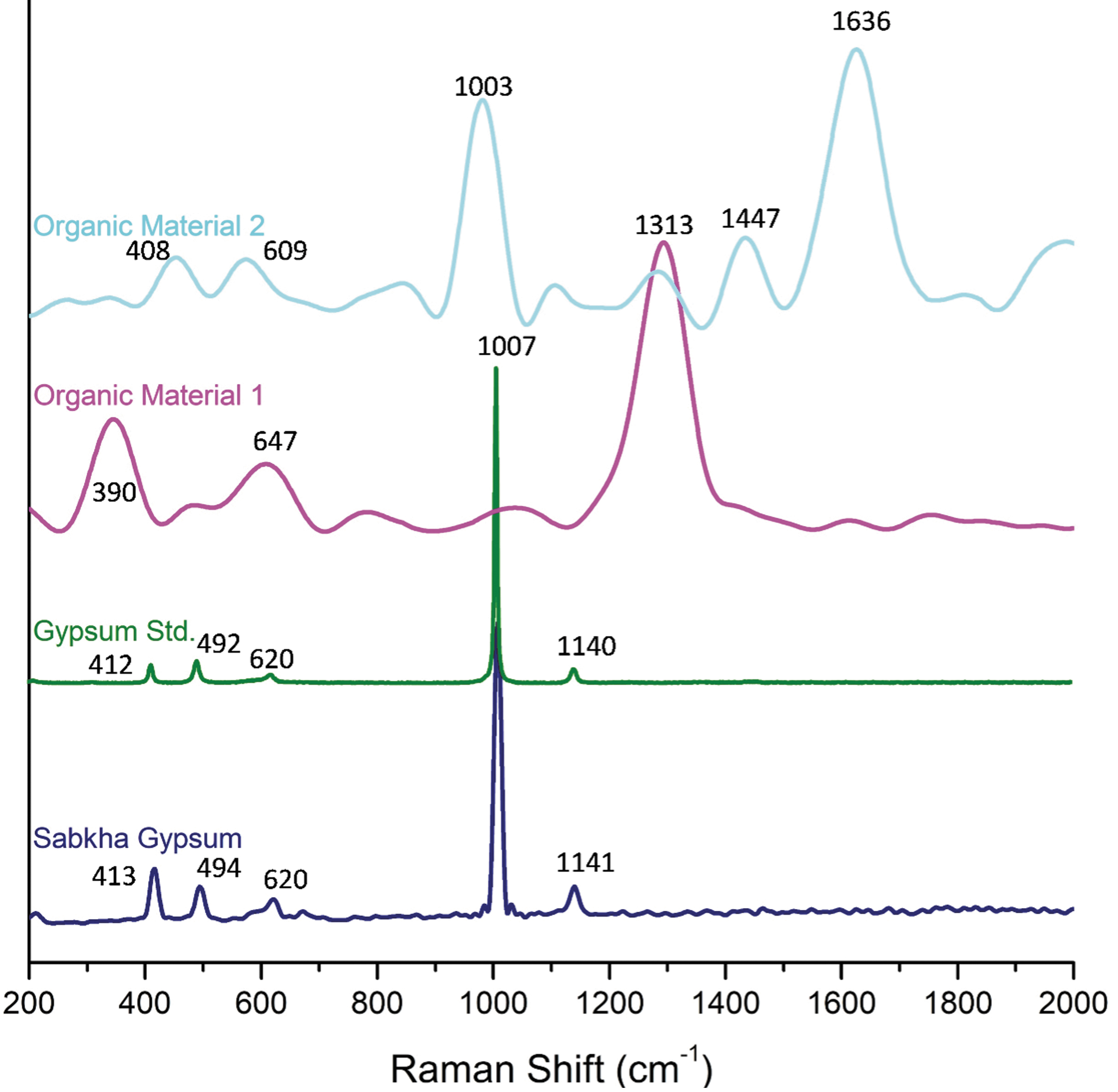
Raman spectra of sabkha gypsum, gypsum reference, and the two different types of organic material detected.
Raman Wavenumber Positions of Gypsum
This study.
These are indicative of different bond wavenumbers that occur in the 600–850, 1250–1300, and 1500–1650 cm−1 ranges across all samples and are generally associated with C, N, and H bonds (Table 2). In addition, by differentially processing the signals that generate the 2D map by subtraction, the spatial association between the organic material (1313 and 1636 cm−1) and gypsum (1007 cm−1) was shown. For both types of organic material, they appear to be incorporated into the gypsum matrix (Fig. 5C–F). This is also evident in the light microscope images, where the examined particles are surrounded by higher surfaces, existing in a “pocket”-like area (Fig. 5A, B).
Main Raman Wavenumber Positions for Samples and Reference Organic Compounds
Bond origin of wavenumber and relative strength of intensity are given if available.
w = weak intensity; m = medium intensity; s = strong intensity.
Analysis of Raman spectrum of pure synthetic β-carotene revealed two distinct peaks at 1513 cm−1 (v 1) and 1155 cm−1 (v 2) and showed a peak at 1010 cm−1 (δ) (Fig. 7). The Raman spectrum of the methanol:acetone extraction solution, which can be used to rule out overlapping peaks from the green mat, orange mat, and 25.5 cm extracts, exhibits two peaks at 1414 cm−1 and a weaker peak at 454 cm−1. In addition to the peaks characteristic of the acetone-methanol extract, the Raman spectrum of the extracted microbial mat hosted gypsum at a depth of 25.5 cm has peaks at 1525, 1019, 611, and 441 cm−1. The Raman spectrum of the sample from the green mat had a distinct peak at 1559 cm−1 and the peaks at 522 and 889 cm−1. The Raman spectrum of the sample of orange mat had two peaks at 1124 and 1484 cm−1, as well as three peaks at 976, 1259, and 1578 cm−1.
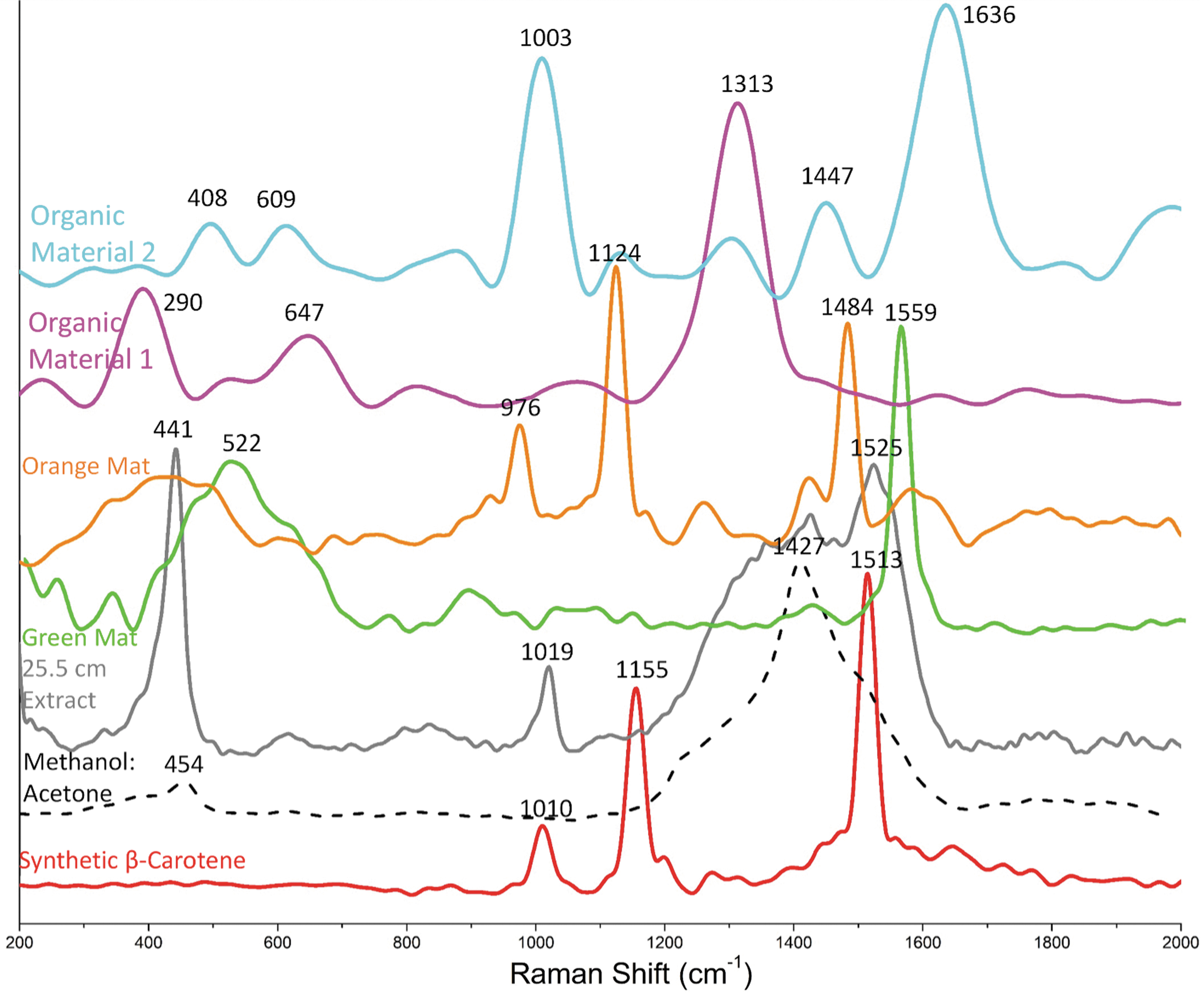
Raman spectra of a β-carotene standard, dried acetone–methanol pigment extract, extracted material from the green mat, the orange mat, the gypsum-forming layer of the current sabkha, as well as embedded organic material within the sabkha gypsum. Reference spectra for gypsum, β-carotene, and the acetone:methanol extract included.
Within gypsum crystals, two different morphological types of organic material were observed: the first type (1) was light colored and more spherical (Figs. 6 and 7), while the second type (2) was black colored with an elongated lens shape (Figs. 6 and 7). The first type of organic material had a distinct peak at 1313 cm−1, two peaks at 647 and 390 cm−1, and two weaker peaks at 818 and 1068 cm−1. The second type of organic material had two high intensity peaks at 1003 and 1636 cm−1, three medium intensity peaks at 498, 609, and 1447 cm−1, as well as three low-intensity peaks at 875, 1130, and 1303 cm−1.
Sabkha's microbial communities and their astrobiological significance
Ultimately, microbial communities are one of the possible sources of organic material, including biomarkers, that would be preserved within gypsum or other mineralogical phases on the martian surface. Thus, microbial community dynamics across Earth analogs is fundamental information that can be used to improve our understanding and interpretation of biosignatures detected on Mars missions. Most environments located on Earth that have astrobiological significance are extreme in nature and include such places as the Canadian High Arctic (Edwards et al., 2005), the Atacama Desert (Vítek et al., 2017; Wierzchos et al., 2018), basins in Italy and Poland (Němečková et al., 2020, 2021), and sabkha environments (Edwards et al., 2006).
Although microbial community analysis has been performed on some of these environments, the focus of the studies was phototrophic organisms (e.g., Němečková et al., 2021) due to the production of pigments, as the key astrobiological biomarker targets, including β-carotene. However, gypsum from this study is found in large abundances buried at a depth of 25 cm, well outside the habitable zone for photosynthetic organisms, suggesting that a different source of microbial community may be involved, and other organic material may be of interest.
Microbial investigations into sabkha environments are less prolific than most Mars analog environments currently studied, with many studies only postulating microbial community composition based on detection of key biochemicals, such as chlorophylls, carotenoids, and scytonemin (Edwards et al., 2006) rather than direct molecular analysis. Descriptions of microbes responsible for production of detectable organic material within these studies are limited to cyanobacteria and nondescript halophilic extremophiles. Data from the current study provide much needed details, and results show that the microbial community within the 25.5 cm gypsum layer is dominated by halophilic archaea (Halobacteria ≈50%); a striking contrast from gypsum endolithic communities dominated by cyanobacterial species.
However, it should be noted that the preceding layer at a depth of 22.5 cm showed a ZOTU abundance of 11.82% for cyanobacterial species. The presence of cyanobacterial ZOTUs at this depth suggests that organic material from the surface may be preserved as burial occurs. Overall, the microbial community described in the sabkha more closely resembles that of halite endolithic communities documented within the Atacama Desert, where Halobacteria dominated (Wierzchos et al., 2018).
It has been previously postulated that halophilic microbes could (or still may) have existed under similar conditions in ancient martian environments (Landis, 2001a, 2001b). Thus, the prolific nature of halophilic archaea is in agreement with previous studies. Specifically, they may have been present in highly concentrated brine solutions due to the lower vapor pressure and freezing point of highly saline solutions in a martian environment (Landis, 2001a, 2001b). Such conditions would be similar to those within the Qatari sabkhas, which can reach salinities of >100 ppt (Diloreto et al., 2019) and create highly saturated conditions with respect to mineralogical phases, such as gypsum. In turn, the crystals of gypsum could have served as a protective environment against desiccation, rapid temperature fluctuations, and exposure to UV radiation for extreme halophiles (Parnell and Baron, 2004), which may also form endolithic communities. Consequently, detection of extant life based on extreme halophilic microorganisms has been proposed (DasSarma, 2006; Oren, 2014; Oren et al., 2014; DasSarma et al., 2020).
Although samples indicated increased abundances of halophilic bacteria, there was also spatial variation of the microbial community composition in replicate samples. Analysis of replicate samples from the same gypsum-forming depths showed a much lower concentration of Halobacteria, which was replaced by significant increases in the abundance of methanogenic archaea, such as Methanobacteria. Methanogenic archaea are not uncommon members of hypersaline microbial mat communities (e.g., López-López et al., 2013: Spring et al., 2015; Fernandez et al., 2016) and are also sources for trapped organic material resulting in potential biomarker targets. Specifically, methanogenic organisms and the lipid biomarkers they produce have been previously proposed as potential astrobiological signatures (e.g., Cheng et al., 2017; Newman et al., 2020). Raman microscopic mapping data collected in this study corroborate these hypotheses that extreme organisms may have sought refuge in brine solutions and crystals of gypsum, which could have acted as an organic material reservoir.
Gypsum as biomarker reservoirs
The high spatial resolution (1 μm) of Raman spectra collected from the gypsum from sabkhas shows a clear distinction between the gypsum matrix and entrapped organic material. We revealed that the organic material was trapped based on sample preparation and chemical image analysis of the 2D Raman maps. To elaborate, sample preparation involved removing any surface contamination from the gypsum in addition to exposing a fresh surface, ensuring that the exposed surface was from within the gypsum itself. Thus, the presence of the organic matter molecules in these samples may only reflect that the organic matter was trapped within the crystal.
The spectral signals vary between both the sabkha hosted material and the pigments extracted from microbial mats. This is due to the highly dynamic sabkha environment and diverse microbial community (Diloreto et al., 2019, 2021), as well as the matrix itself. It is well documented that biomarkers are not only simply trapped within minerals but also adsorbed to and interact with their mineral hosts (Röling et al., 2015). Minerals, such as gypsum, can interact with organic molecules based on both phases surficial chirality, which has been hypothesized to result in “sabre-gypsum” (Babel, 1990; Allwood et al., 2013). In addition, sulfate minerals can cause oxidation of organic biomarkers when analyzed, as well as strongly adsorb DNA if they contain silica (e.g., jarosite) (Navarro-González et al., 2010; Lewis et al., 2015; Röling et al., 2015).
These reactions are complex in their nature, depend on mineralogy and formational environment and the factors such as pH, ionic strength, and redox, which can influence ligand chemistry, and as a result cause a complex Raman signature. Although the mineral host can preserve and may chemically interact with biomarkers, studies using similar Raman spectroscopy methods, as implemented here, have shown that biomarkers are discernable in concentrations as low as 0.1 μg/g while embedded fully in transparent or translucent minerals (2.5 mm thick calcite, 40 mm thick halite) (Hooischurr et al., 2016).
In conjunction with mineral–biomarker interactions, the microbial community forming biomarkers is an important aspect contributing to the complexity of the Raman spectra. The microbial community not only determines the biomolecules produced but also governs transformations of organic material during the life of the microbial mat (Diloreto et al., 2021). Furthermore, microbial communities vary not only between mats at different locations but also at different depths within the mats. Regarding extant microbial communities, this suggests that it is important to examine the microbial community of Earth analogs to determine what original organic material is being trapped and preserved to predict biomolecular end-members that might be identified in a martian environment.
Implications for ExoMars missions and the detection of life
One proof that life once existed on Mars would be the detection of biomarkers, which are distinct organic molecules left behind by microorganisms inside the materials that were formed in the past. Among the possible Earth analogs of early Mars, evaporitic systems are a model environment having high salinity, a richness of halophilic and methanogenic organisms, and carbonate and sulfate mineral precipitation. All of these conditions are favorable to the preservation of organic material, and surveys for biomarkers need to reflect these conditions. Despite the fact that evaporitic systems have these recurring characteristics, they do not significantly narrow down targets for biomarker investigations. This is caused by high degree of diversity among halophiles and methanogens and the organic material they produce.
Among these organisms, there are several groups of organic material that can eventually transform into precursor biomarkers, including carotenoids, chlorophylls, phycobiliprotein, scytonemin, mycosporine-like amino acids, pulvinic acid derivatives, and anthraquinones (Jehlicka and Oren 2013; Jehlička et al., 2014). Despite many possible biomarkers originating from microbes, there is a consensus that one of the signs of the existence of past life on Mars may be the presence of β-carotene in gypsum deposits (Vítek et al., 2009a). The presence of this biomolecule would indicate that there once existed life-forms, which produced this DNA repair agent and employed its UV-resistant properties in their survival strategy (Patel et al., 2004).
One challenge to overcome is to find a technique that is capable to detect at high resolution in situ biomolecules trapped within gypsum. Several studies investigating the ability of Raman spectroscopy to effectively identify β-carotene within a gypsum sample have used samples that were either created in the laboratory by spiking with synthetic β-carotene levels (Vítek et al., 2009a, 2009b) or specifically selected from the natural environment due to the physical appearance of endolithic communities within the sample (Jorge-Villar et al., 2005, 2011).
Our previous studies that collected Raman data on microbial mats and exopolymeric substances (EPS) extracted from these environments demonstrated strong signatures for carotenoid compounds (e.g., Paulo and Dittrich, 2013; Diloreto et al., 2021). In our study, the primary indicator of β-carotene in the samples is a distinct v 1 peak at 1513 cm−1, as well as v 2 peak at 1141 cm−1, and δ peak at 1010 cm−1. Although as expected for an complicated environmental matrix we did not observe these specific peaks, there were distinct peaks between 1500 and 1550 cm−1 assigned to C = C stretching as well as 1150–1170 cm−1 assigned to C-C stretching and 1000–1020 cm−1 assigned to C-CH3 deformation associated with carotenoids. These peak positions were pronounced in the spectrum of organic material 2, as well as extracts from the green mat, orange mat, and 25.5 cm within the DF core.
Several previous studies such as Jehlicka and Oren (2013), Jehlička et al. (2014), and Vítek et al. (2017) have demonstrated that the peaks of organic material can be grouped, for example, carotenoids from 1500 to 1550 cm−1, chlorophyll from >1550 to 1600 cm−1, and phycobiliproteins from >1600 to 1650 cm−1. Given the suggested Raman groups for organic materials, we assigned the organic peaks in the spectrum from sabkha samples to carotenoids. However, the peak wavenumbers present in the aforementioned samples may also coincide with those of chlorophyll, in particular chlorophyll a, which might also be preserved within gypsum.
Specifically, both Raman spectrum of organic material 2 and the green mat exhibited peak at the wavenumber >1550 cm−1 falling outside the wavenumber position for C = C stretching of carotenoids, but within the wavenumber position for C-C stretching characteristic of chlorophyll a. These peak positions in conjunction with high abundances of cyanobacterial ZOTUs suggest that chlorophyll is also preserved within sabkha gypsum. The previous laboratory study of carotenoids inclusions in evaporites showed similar Raman band positions of carotenoids from haloarchaea Halobacterium salinarum, Halorubrum sodomense, and Salinibacter ruber (Bacteroidetes), namely at 1508/1150/991 cm−1 (Jehlička et al., 2018).
Interestingly, organic material 1 exhibited different peak positions when compared with other organic material. Spectrum from organic material 1 exhibited no peaks >1313 cm−1 making it unlikely to be organic material associated with carotenoids. Organic material 1 exhibited Raman activity at 1313, 1068, 818, 647, and 390 cm−1, which could be indicative of chlorophyll as a result of N-C bending, C-C-C or N-C-C bending, and CH deformation or O-C-O vibrations, respectively. In addition, these peak positions might be indicative of pulvinic acid derivatives. Considering the multitude and complexity of the wavenumber positions, the identification of specific organic molecules requires more information. Thus, it is crucial to refer to the microbial community closely associated with a layer where gypsum was found to further delineate the spectral results. Afterward, the link between organic molecules identified by Raman spectroscopy and microbial community description can be identified based on known spectra and information from previous studies.
Despite the harsh nature, sabkhas host thriving microbial communities forming microbial mats that are also rich in microbial diversity (Diloreto et al., 2019, 2021). Notably, similar hypersaline communities may represent extant microorganisms due to their adaptability and resilience to extreme environmental conditions. The microbial community composition within the sabkha mats is diverse, but the near-gypsum and gypsum-forming layers of the sabkha are dominated by Halobacteria and Methanobacteria. Within Halobacteria, there exist several genera such as Halococcus, Halobacterium, and Natronomonas that have the potential to produce well-documented halophilic carotenoids such as bacterioruberin and variations of β-carotene (Jehlicka and Oren 2013; Jehlička et al., 2014). In addition, there is a striking similarity in microbial community composition, morphology, coloration, and structure between endolithic samples examined from the Atacama Desert (Vítek et al., 2010, 2017; Wierzchos et al., 2018) and sabkha microbial mats (Diloreto et al., 2019, 2021) where carotenoids, chlorophyll, and scytonemin have been directly observed.
Consequently, based on our data, we can distinguish between the gypsum matrix and organic material, and we observed the complex spectra obtained from the organic material. Methanobacteria also consist of several genera such as Methanothermos, Methanothermobacter, and Methanobrevibacter capable of producing organic material of interest mainly from archaeal lipids (e.g., Cheng et al., 2017; Newman et al., 2020). More specifically, lipids such as archaeols, sn-2-hydroxyarchaeol, and other glycerol diethers are organic molecules that transform into conventional biomarkers through geological time and were characterized by UV Raman spectroscopy in carbonate microbialites as the broad G peak around 1600 cm−1 (Brocks and Pearson, 2005).
Overall, the combination of Raman spectra coupled with amplicon sequencing suggests that the primary organic material preserved within gypsum are carotenoids, chlorophylls, and archaeal lipids, suggesting that any biomarkers preserved on Mars would be diagenetic products of these precursors. In terms of β-carotene, the geological biomarker equivalents would be β-carotene or γ-carotene, whereas other carotenoids compounds such as those identified by Raman spectra in this study would result in other saturated carotenes such as lycopene or aromatic carotenoids such as chlorobactane, okenane, isorenieratane, renieratene, or renierapurpurin (Brocks et al., 2005: Brocks and Schaeffer, 2008; Lee and Brocks, 2011).
For chlorophyll compounds, biomarker targets include geoporphyrins, geochlorins, or hopanoid compounds (Woolley et al., 1998; Brocks et al., 2003; Schinteie and Brocks, 2017), while methanogenic archaeal biomarkers mainly consist of compounds such as crocetanes, methylhopanes, pentamethylicosane, and crenarchaeol (Brocks et al., 2005; Schinteie and Brocks, 2017; Cheng et al., 2017: Newman et al., 2020). Finally, some studies have observed biomarkers indicative of general hypersaline conditions, such as squalene and biphytane (Schinteie and Brocks, 2017).
Interestingly, another consideration that has not been taken into account in the literature is the reprecipitation of gypsum itself, which is documented in such environments (Edwards et al., 2006) and could create separate or multiple organic material preservation events (Jehlička et al., 2018). This could greatly influence potential biomarker targets. This is clear in the amplicon sequencing and Raman spectrum data from this study where organisms no longer alive have left behind organic material (i.e., carotenoids and chlorophyll) at a depth where accumulation of the mats may have taken several decades.
Extrapolating our findings to a martian environment where any biomarkers would be of geologically significant age presents an important consideration for the evaluation of martian samples. Gypsum deposits have been hypothesized to have undergone multiple diagenetic events under various environmental conditions (Yen et al., 2017; Hausrath et al., 2018; Vaniman et al., 2018). These variations in environmental conditions and variations in the geochemistry of gypsum-forming fluids (Yen et al., 2017) could result in different microbial communities leaving behind sets of biomarkers at different times, resulting in complexities in biomarker signals.
Conclusions
Implementing Raman microscopy, DNA sequencing, and laboratory experiments, this study suggests that biomolecules produced by microbes can be trapped within a gypsum matrix and detected by Raman microscopy. The combination of field, laboratory, and Raman microscopy data has shown that minimal sample preparation is required to detect biomarker signals within a gypsum mineral matrix by Raman microscopy. The data in this study also highlighted the trapping of carotenoid, chlorophyll, and likely archaeal lipid organic molecules that can be converted to biomarkers over geological time. However, from a practical perspective, many of these biomarkers are identified with isotopic techniques, gas chromatography, or high-precision liquid chromatography, which is not feasible on a Mars mission. There are limited studies on the Raman signature of these compounds within astrobiological context, and future work should focus on a combination of high resolution and enhanced techniques to delineate complex biomarker signatures.
This study demonstrated that interpretation of the highly complex Raman signal could be aided through a hybrid approach by coupling the spectrum with microbial community analysis to help confirm or exclude certain organic molecules. Due to the high diversity of microbial communities in many Earth analogs, the signal of biomarkers trapped within a mineral matrix is quite complex, and it requires additional techniques to be unambiguously interpreted. Future work can focus on a similar combination of microbiological techniques for molecular characteristics and high spectral resolution analysis of biomarkers that allow for differentiation of single biomarkers. One such type of analysis is high-performance liquid chromatography, which could be coupled with high-resolution and functional metagenomics (e.g., metatranscriptomics or whole-genome sequencing) of Earth analogs to develop a microbe biomarker Raman fingerprint, which may be an important step based on the complex formational history of martian gypsum.
As remains from an extinct or extant species on Mars can be preserved within the gypsum matrix, it is important to understand their analogous Earth sources and how microbial remains may persist in mineralogical reservoirs.
Footnotes
Authors' Contributions
M.D. and M.S.A. formulated the concept of this study. Z.D., M.D., and T.R.R.B. were responsible for field work and collection of samples. Z.D. and M.D. were responsible for DNA analysis. Z.D., M.D., and M.S.A. were responsible for laboratory preparation and Raman analysis of samples. M.D. and T.R.R.B. secured funding for the project. Z.D. and M.D. prepared the article, and all authors contributed to the discussions and revisions.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors would like to acknowledge the Qatar National Research Fund (a member of Qatar Foundation) for the Grant NPRP12S-0313-190349. M.D. was also supported by the National Sciences and Engineering Research Council of Canada (NSERC Discovery Grant) and the Canada Foundation for Innovation and Ontario Research Fund (Leaders Opportunity Fund, Grant No. 22404).
Abbreviations Used
Associate Editor: Michael C. Storrie-Lombardi