
Abstract
Rationale:
There is little information regarding the allergen content of milk feeds in the preterm population. Previous studies have not performed a broad analysis of the allergenic peptide content and protease activity of milk feeds in this population.
Methods:
To evaluate feasibility, we initially performed mass spectrometry on 4 human milk (HM) samples (2 term and 2 preterm) from the Mommy's Milk Human Milk Biorepository (HMB) and analyzed the results against the University of Nebraska FASTA database and UniProt for a total of 2,211 protein sequences. We then further analyzed five samples from the Microbiome, Atopy, and Prematurity (MAP) study including peptidomic and protease activity analysis.
Results:
Each HMB sample had between 806 and 1,007 proteins, with 37–44 nonhuman proteins/sample encompassing 26 plant and animal species. In the preterm MAP samples, 784 digested nonhuman proteins were identified, 30 were nonbovine in origin. Proteins from 23 different species including aeroallergens, food, and contact allergens were identified. Protease activity was highest in HM samples without human milk fortifier and lowest in preterm formula.
Conclusions:
These findings represent the first preterm milk feed mass spectrometry and protease analysis with identification of known allergenic proteins to food, contact, and aeroallergens. These results raise questions of whether the composition of milk feeds in the neonatal intensive care unit impact the development of atopic disease in the preterm population and whether the complex interaction between allergens, proteases, and other HM components can serve to induce sensitization or tolerance to allergens in infants.
Clinical Trial Registration Number: NCT04835935
Introduction
There has been increasing recognition of the protective role of human milk (HM) and early food exposure in the development of atopic conditions such as eczema, food allergy, and asthma.1–4 Asthma is the most prevalent chronic disease in children, affecting >300 million people worldwide 5 and disproportionately affects preterm infants. In one study, infants born before 37 weeks (moderate to late preterm) were 50% more likely to develop asthma, and infants born before 32 weeks (extremely to very preterm) were three times as likely to develop asthma. 6 Lower rates of atopy in term infants are associated with exclusive breastfeeding for the first 4 months of life, 7 lack of antibiotic exposure (either maternal intrapartum or early in infancy), 8 vaginal delivery, 9 and furry pets in the home. 10 Preterm infants spend this critical period in the neonatal intensive care unit (NICU), and some are exposed to antibiotics and cesarean section delivery but most importantly, almost all experience most of their nutrition via a nasogastric tube or a bottle containing previously frozen maternal or donor breast milk, instead of fresh breast milk via breastfeeding.
Recommendations regarding the introduction of allergenic foods to infants has shifted from avoiding allergenic foods until 1 year of age to early introduction before 6 months of age, as the latter has been shown to be associated with a decreased risk of food allergy during a critical window of the infant's immune development. 4 This approach is thought to reduce the induction of type 2 inflammation that is responsible for allergic conditions. Given that early exposure to food proteins can alter the development of food allergies later in life, early nutrition has become an area of interest in studying the pathogenesis of atopic disease. 11 Over 70% of peanut reactions occur on the first known exposure, indicating prior sensitization, perhaps transcutaneously, through inhalation, or through HM.12,13 Because HM is often the primary source of nutrition in infancy, it has been postulated that it could be a source for allergen introduction. 13 Although immunoglobulin E (IgE)-mediated reactions to HM are rare, they do occur, demonstrating its immunogenic nature. 14
Airborne allergens from house dust mite have also been found in HM at similar quantities to food allergens.15,16 One dust mite protein, Der p1, has demonstrated both Toll-like receptor agonist and protease activities, which could potentially initiate allergic immune responses. 16 In addition to food and environmental allergens, HM also contains many other bioactive substances, including endogenous proteases and protease inhibitors, immunoglobulins, soluble receptors, cytokines, HM oligosaccharides, fatty acids, and microbes.11,17–19
HM proteomics studies have utilized different methods for protein identification, including Western blotting, enzyme-linked immunosorbent assay, and mass spectrometry, which may account for heterogeneity between experiment results.20–22 Mass spectrometry can determine the protein content in HM; however, there are few studies that are primarily focused on allergenic proteins. 11 Studies utilizing mass spectrometry, which allows for a broader untargeted search for proteins, have identified 1,200–1,600 total proteins in HM, with most nonhuman proteins being derived from cow's milk and one study also identifying dog, horse, cat, chicken, and rice proteins.14,23
The development of tolerance versus sensitization to allergens is complex and depends on the interaction and often multidirectional relationship between many different factors, such as maternal history, milk composition, gut immunology, and microbiome and external environment. 17 Allergen shedding in HM may be a way to educate the infant's immune system and modulate allergy risk in the infant. 24 Owing to newer techniques in proteomic and peptidomic analysis, and the paucity of data regarding the presence of allergens in HM, we developed a study on a subset of milk samples to investigate allergenic peptide content and protease activity of milk feeds in the preterm infant population.
Methods
Sample collection
Four HM samples (two from mothers of term infants and two from mothers of preterm infants) were analyzed from the Mommy's Milk Human Milk Biorepository (HMB) to evaluate the feasibility of analyzing HM samples by untargeted mass spectrometry (Table 1). 25 Following informed consent, women provided 50 mL up to a full pump of expressed breast milk (convenience sample). Participants were interviewed about their sociodemographic characteristics, pregnancy history, dietary intake using a standard questionnaire (NutritionQuest), and breastfeeding behaviors. Sample collections occurred at UC San Diego, community sites, or the participant's home. HM samples were stored and shipped on ice within 24 hours of collection to the Mommy's Milk lab, where the sample was aliquoted and stored at −80°C until requested for study analysis.
Maternal Dietary History: Mommy's Milk Human Milk Repository Samples
We also analyzed five HM samples and one formula sample (J11) from mothers and preterm infants recruited into the Microbiome, Atopy, and Prematurity (MAP) pilot study. 26 The MAP study population recruited 48 preterm infants, ≤34 weeks, and their mothers, from Jacobs Medical Center at UCSD NICU and Scripps Memorial La Jolla NICU. At birth, prenatal (maternal antibiotics and diet, pregnancy morbidities, smoking, pet ownership, family history of asthma, and other social and demographic information) and perinatal (method of delivery, need for resuscitation) factors/exposures were documented at the time of enrollment. Parents were given a History and Allergy Questionnaire at enrollment, which asked about family history of asthma, smoking, allergies, medication, and dietary history during pregnancy and postnatally (Table 2 and Supplementary Table S1).
Maternal Dietary History: Microbiome, Atopy, and Prematurity Samples
Milk, stool, and saliva samples from the study participants were collected weekly and stored immediately at 4°C and transferred to −80°C within 36 hours postcollection. For this pilot study, first week milk samples were analyzed from six infant/parent couplets. Because most preterm infants require fortification to support their growth and development and mothers may not have adequate milk supply, half of the milk samples (3/6) analyzed from the MAP study contained formula-based fortifiers, and all but one sample (J9) contained donor milk. To control for additives in fortifiers, an additional two samples each of regular and hydrolyzed HMF (Enfamil human milk fortifier acidified liquid; Mead Johnson) and one formula sample (Enfamil premature; Mead Johnson) were analyzed.
This research was performed in accordance with the ethical principles for medical research involving human subjects outlined in the Declaration of Helsinki. The study protocol was approved by the University of California, San Diego's Human Research Protections Program IRB No. 181711.
Sample preparation for proteomic analysis
Milk samples were thawed on ice before preparation for proteomic analysis. Milk samples were not skimmed. Guanidine-HCl was added to 2 μL of milk sample to achieve a final concentration of 6 M. The samples were boiled for 10 minutes followed by 5 minutes cooling at room temperature. The boiling and cooling cycle was repeated three times. The proteins were precipitated with the addition of methanol to a final volume of 90% followed by vortex and centrifugation at RCFmax 20,800 × g on a benchtop microfuge for 10 minutes at 25°C. The soluble fraction was removed by flipping the tube onto an absorbent surface and tapping to remove any liquid.
The pellet was suspended in 200 μL of 8 M urea made in 100 mM Tris pH 8.0. Tris (2-carboxyethyl) phosphine and chloro-acetamide were added to final concentrations of 10 and 40 mM, respectively, and the mixture was vortexed for 5 minutes. Three volumes of 50 mM Tris pH 8.0 was added to the sample to reduce the final urea concentration to 2 M. Trypsin was added in a 1:50 protein ratio and incubated at 37°C for 12 hours. The solution was then acidified using trifluoroacetic acid (TFA) (0.5% TFA final concentration) and mixed. Samples were desalted using 100 mg C18-StageTips (Thermo Scientific™ Pierce™ C18 Tips, 100 μL bed Catalog No. 8778) as described by the manufacturer protocol. The peptide concentration of the samples was measured using bicinchoninic acid assay after resuspension in sample loading buffer and a total of 0.5 μg was injected for each label-free quantification run. 27
Sample preparation for peptidomic analysis
To remove high molecular weight milk proteins, 100 μL of HM was mixed with 900 μL of methanol and vortexed for 5 seconds. The samples were kept at 25°C for 30 minutes followed by centrifugation at RCFmax 15,294 × g for 10 minutes at 25°C. Five hundred microliters of supernatant was transferred to a fresh tube dried in a vacuum centrifuge. The samples were hydrated in 0.5 mL of 0.5% formic acid and 5% acetonitrile (ACN) solution and desalted using a Sep-PAK C18 1cc Vac (Waters Corp., Milford MA) according to the manufacturer's protocol with the exception that 40% ACN was used to elute peptides. The eluents were dried in speed-vac in preparation for mass spectrometry analysis. A summary of the samples and the type of analysis they underwent is listed in Table 3.
Milk Sample Composition and Type of Analysis
Fortifier: Enfamil Human milk fortifier (nonhydrolyzed). All MAP samples (J7–J12) contain donor milk aside from J9.
HM, human milk; MAP, Microbiome, Atopy, and Prematurity.
Liquid chromatography with tandem mass spectrometry
Trypsin-digested peptides were analyzed by ultrahigh-pressure liquid chromatography (UPLC) coupled with tandem mass spectroscopy (LC-MS/MS) using nanospray ionization. The nanospray ionization experiments were performed using a Orbitrap fusion Lumos hybrid mass spectrometer (Thermo) interfaced with nano-scale reversed-phase UPLC (Thermo Dionex UltiMate™ 3000 RSLC nano System) using a 25 cm, 75-μm ID glass capillary packed with 1.7-μm C18 (130) BEHTM beads (Waters Corp.). Peptides were eluted from the C18 column into the mass spectrometer using a linear gradient (5–80%) of ACN at a flow rate of 375 μL/min for 2 hours. The buffers used to create the ACN gradient were as follows: Buffer A (98% H2O, 2% ACN, 0.1% formic acid) and Buffer B (100% ACN, 0.1% formic acid).
Mass spectrometer parameters are as follows: an MS1 survey scan using the orbitrap detector (mass range [m/z]: 400–1,500 [using quadrupole isolation], 120,000 resolution setting, spray voltage of 2,200 V, ion transfer tube temperature of 275°C, AGC target of 400,000, and maximum injection time of 50 milliseconds) was followed by data-dependent scans (top speed for most intense ions with charge state set to only include +2–5 ions, and 5-second exclusion time, whereas selecting ions with minimal intensities of 50,000 at which the collision event was carried out in the high-energy collision cell [HCD Collision Energy of 30%]). Fragment masses were analyzed in the ion trap mass analyzer (with ion trap scan rate of turbo, first mass m/z was 100, AGC target 5000, and maximum injection time of 35 milliseconds). Protein identification and label-free quantification was carried out using Peaks Studio 8.5 (Bioinformatics Solutions, Inc.) All label-free quantifications were normalized to total ion count for each sample.
Analysis was performed in two separate runs—the first included the samples from the breast milk repository, and the second included the samples from the MAP study, formula, and fortifier. Database searches were carried out against a reference database that included FASTA protein sequences of known protein allergens from the University of Nebraska (www.allergenonline.org/; used version 19, published February 10, 2019) that was combined with human proteome UniProt sequences using Peaks 8.5 (Bioinformatics Solutions) search engine. This database included known allergenic peptides and all predicted human proteome. The hits were filtered at 1% false discovery rate (FDR) before being considered for further analysis. All positive peptide sequence results were verified by blasting original sequences against UniProt (https://www.uniprot.org/blast/) and NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi) to confirm accuracy. Results where at least one of the two of the databases did not have >70% identity to the labeled species-specific protein were discarded.
In addition, an experiment to address false hits that may arise from our analysis using a Hela cell line confirms that the nonhuman protein/peptide sequences are in fact nonhuman. By running a control analysis using Hela cells, a human cell line, we are able to identify proteins that are either human or part of the analysis process. We then filtered for nonhuman proteins and compared them with our samples to identify false hits (Supplementary Table S2). This process further assures that nonhuman proteins identified in the samples are in fact nonhuman and not a contaminant from the analysis process itself. Only one false hit (identified by [?] in Supplementary Table S2.) Salmo Salar (salmon) Enolase found in the Hela cell line was found in our samples. Potential false hit identified are given in Table 4.
List of Pertinent Bovine Peptides and All Non-Human Nonbovine Peptides Stratified by Sample Proportion in Fortifiers, Human Milk Biorepository, and Microbiome, Atopy, and Prematurity Samples
Bolding denotes known protien nomenclature.
Total number of samples: 14 (4 from Mommy's Milk Human Milk Repository, 6 from Microbiome, Atopy, and Prematurity Study, 4 Human Milk Fortifier Samples).
(?) Salmo salar may represent a “false hit” based on secondary analysis. Summarized in Supplementary Table S2.
PEAKS Sequence search parameters utilized and peptide overlays in the HTML format are extracted from Peaks analysis. PSM (peptide-spectrum-match) scores, sequence coverage of the identified proteins, number of peptides identified, protein IDs, MS/MS spectra of the nonhuman peptide sequences reported in Pride database and sequences can be downloaded (Project Accession: LPXD033473). Information regarding PEAKS search parameters are given in Supplementary Data.
Protease activity
Samples were diluted 50-fold in 50 mM Tris-HCl, pH 9.0, 150 mM NaCl, and assayed with 25 μM RR-AMC (sc-281540; Santa Cruz Biotechnology) in triplicate wells on a black 384-well plate. The final volume in each well was 30 μL and the assay was performed at 25°C. Activity was monitored for 2 hours on a BioTek HTX plate reader with excitation of 360 nm and emission of 460 nm. Activity was reported as the change in relative change in fluorescence units per second.
Results
Proteomic feasibility study from the Mommy's Milk HMB samples
To determine the feasibility of detecting allergenic peptides/proteins in HM, we performed mass spectrometry on 4 (2 term and 2 preterm) trypsin-digested breast milk samples (R1, R2, R3, R4) and utilized the University of Nebraska FASTA and UniProt databases for a total of 2,211 sequences for comparison. Each sample had between 806 and 1,007 peptides with 28–38 nonhuman proteins per sample encompassing 23 different plant and animal species (Table 4). We detected peptides from various food, venom/salivary, and airborne sources. The highest proportion of nonhuman peptides was identified as cow, yak, cat, horse, dog, pig, and dust mite. There were no appreciable differences between term and preterm samples in terms of total protein content. One sample accounted for >50% of the nonhuman peptide variability (R4).
In their dietary histories, all mothers ingested fish, shellfish, nuts, and wheat. Two of the four mothers did not consume cow's milk (dairy), although bovine peptides were found in all samples. Two of the four mothers did not ingest egg, yet egg protein was detected in both those samples. Of interest, the one mother who drank almond milk was the only one who did not have almond detected.
Proteomic pilot study from the MAP samples
Once the protocol was validated with the samples from the HMB repository, a subsequent mass spectroscopy run was performed on samples from the MAP study including five HM samples, one formula sample, two regular milk fortifier samples, and two hydrolyzed milk fortifier samples. All the milk samples except for one (J9) were a mix of maternal expressed breast milk or donor breast milk and/or HMF. J9 contained maternal expressed breast milk only.
We performed proteomic and peptidomic analysis on the MAP samples and HMFs. We identified a total of 784 digested nonhuman proteins, 754 that were bovine in origin and 30 nonbovine in origin. Hydrolyzed HMF samples had significantly fewer proteins (average of 48) compared with nonhydrolyzed HMF (average of 264), HM (average of 256), and formula (average of 236) (Supplementary Fig. S1).
In total, we identified proteins from 23 different species, including aeroallergens, food and contact allergens (Table 4). For quality control purposes, peptide alignment maps were made for two common allergens, β-lactoglobulin and cat albumin. The maps demonstrated consistent overlap, thereby supporting that these proteins were indeed identified and digested similarly between samples (Supplementary Fig. S2).
Bovine peptides were the most numerous of the nonhuman peptides detected in HM samples with and without HMF. Specific allergenic bovine peptides (β-lactoglobulin, α- and β-casein, α-lactalbumin) were in the highest relative quantification in regular fortifier and formula, intermediate in hydrolyzed fortifier, and lowest in HM samples (Supplementary Fig. S3).
Peptidomics studies of the MAP samples identified peptides that were generally <40 amino acids in length (Fig. 1). Most nonbovine peptides were digested (28/33); of those 28 nonbovine digested peptides, 26 were found in samples with HM and fortifier, which contrasts with only a few (2/28) being found in the formula sample.
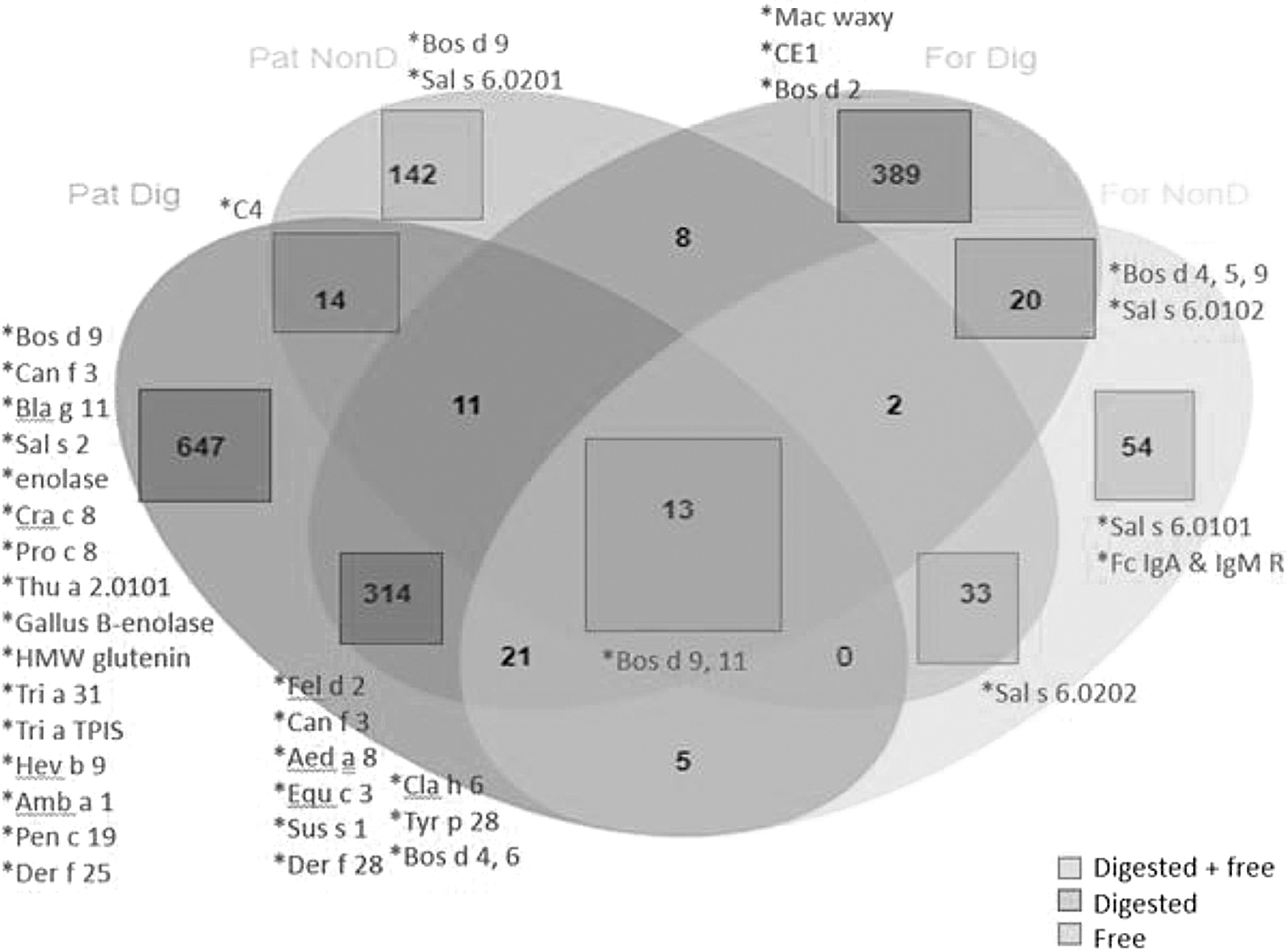
Distribution of free and digested peptides in MAP samples. For Dig = formula sample digested; For NonD = formula sample nondigested; Pat Dig = patient samples digested; Pat NonD = patient samples nondigested. MAP, Microbiome, Atopy, and Prematurity.
Protease activity
Upon analysis of the amino acids at the N- and C- termini of peptides, we discovered that there was a high frequency of proline (P) and glutamine (Q) residues in the MAP samples. This was not surprising, as human casein is the major protein in these samples, with residue composition entailing 17% proline and 11% glutamine. Samples J8–J11 are notable as they have peptides that were cleaved after lysine (K) and arginine (R) at the N-terminal side of peptides. This is not seen for J7 and J12 (Fig. 2). To quantify protease activity in these HM samples, we assayed the samples with the fluorogenic substrates, Arg-Arg-AMC. We found that the most abundant protease activity was in pure HM sample (J9), whereas the formula-only sample (J11) had the lowest activity (Fig. 3).
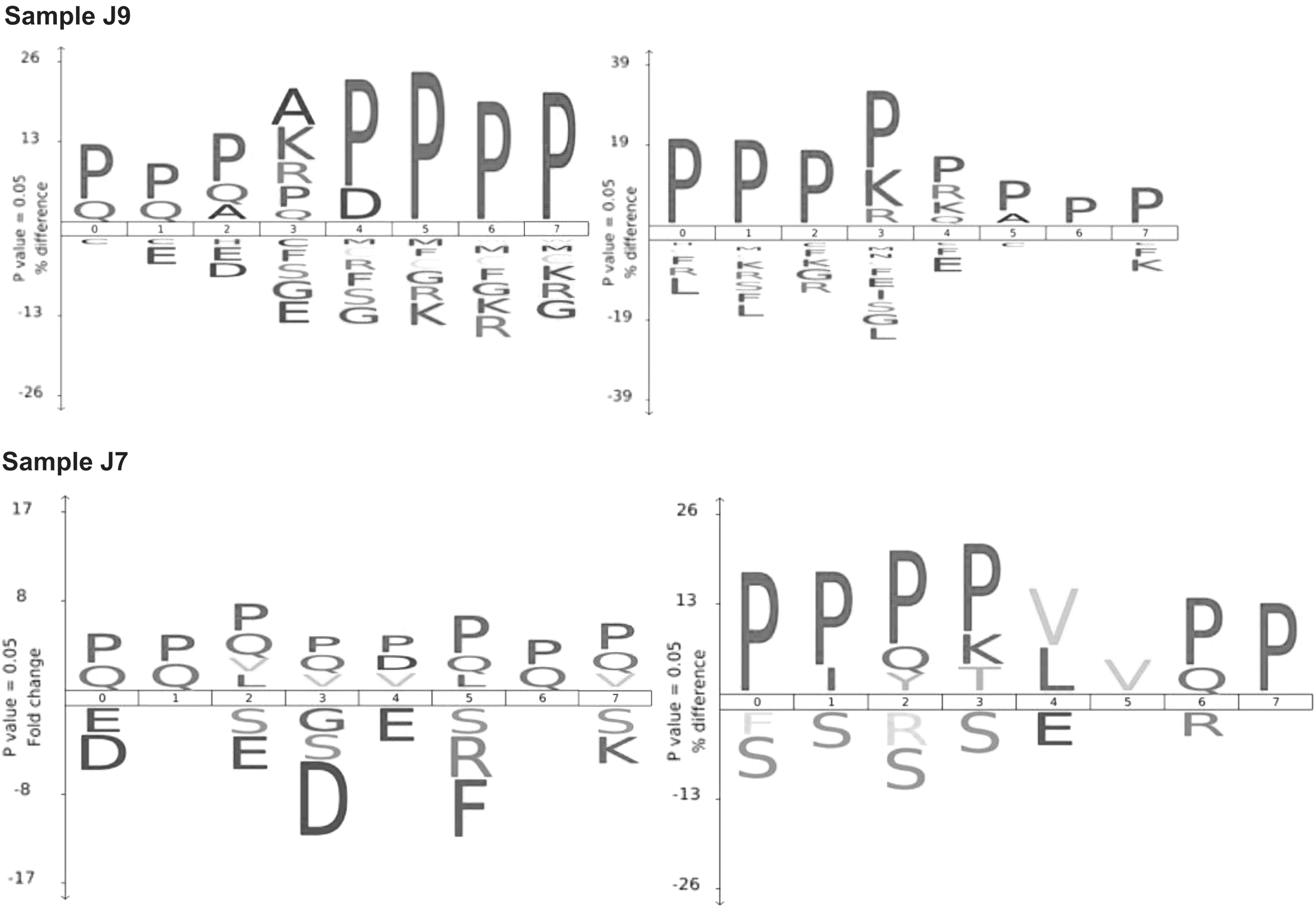
Protease activity in MAP samples. Frequency of amino acids at the four positions or either side of the cleaved bond (positions 0 to 7 where cleavage occurs between 3 and 4). The
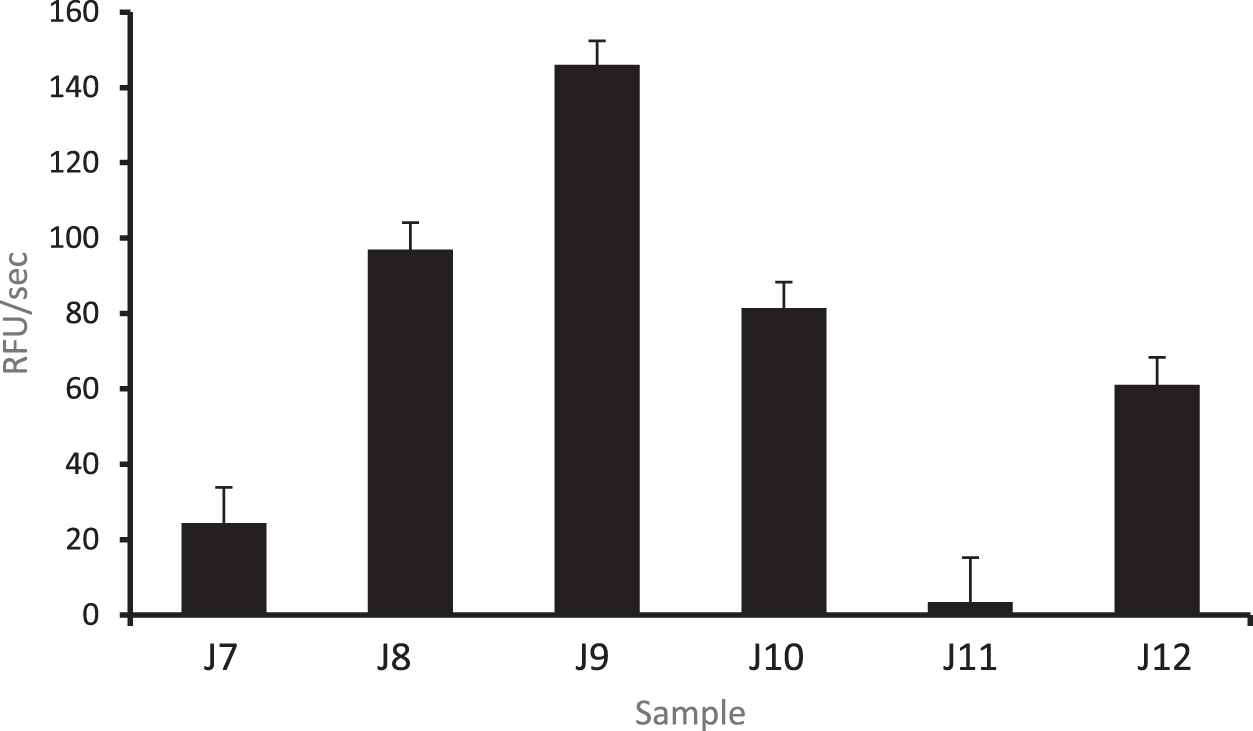
Protease activity in MAP samples. Proteolytic activity in milk samples using the fluorogenic dipeptide substrates Arg-Arg-AMC. J9: Maternal expressed breast milk only. J11: Fortified formula. MAP, Microbiome, Atopy, and Prematurity.
Discussion
To our knowledge, this is the first combined HM mass spectrometry and protease analysis using clinically relevant NICU preterm milk samples. Although the detection of food peptides in the milk samples is interesting and somewhat expected, the size, breadth, and variety of food and aeroallergens as well as other nonfood peptides is fascinating, especially when comparing HM with cow's milk formula. Furthermore, we found differential protease activity between the samples, with the highest being in maternal expressed breast milk alone, without fortifier (J9), and the lowest in preterm formula (J11).
The presence of allergen peptides in HM may be linked to the development of allergy, especially in susceptible individuals. In one study, there was an increase in atopy in children who were breastfed by atopic mothers who had high HM dust mite (Der p 1) levels; this was not noted in the offspring of mothers without atopic history, regardless of Der p 1 level in HM. 28 In addition, in food allergy, mother's avoiding cow's milk had lower breast milk beta-lactoglobulin-specific IgA, and their children had lower serum beta-lactoglobulin and casein IgG1 and β-lactoglobulin IgG4 with higher rate of cow's milk allergy. 29 In our analysis, bovine peptides were the most numerous of the nonhuman peptides detected in the HM-only samples (without formula or fortifier) and specific allergenic bovine peptides (β-lactoglobulin, α- and β-caseins, α-lactalbumin) were found in the highest relative quantification in regular fortifier (F) and formula sample (J11) and lowest in the HM sample (J9) (Supplementary Fig. S3).
Multiple other common food allergens have been identified in HM studies. Studies have demonstrated dose–response phenomenon between maternal egg intake and concentration in the breast milk, 30 with one study demonstrating a relation to infant serology whereby for each additional egg ingestion, HM ovalbumin concentration increased by 25% and infant egg-specific IgG4 increased by 22%. 31 However the presence of ovalbumin and ovomucoid in HM vary greatly in different studies (8.3–78%).30–32 Egg protein (β-enolase) was found in our analysis, although specific ovalbumin and ovomucoid peptides were not identified. Regarding peanut protein, a small study demonstrated peanut allergen (Ara h 6) in HM that was functional and IgE-reactive as evidenced by in vitro assays and the observation that administration to mice lead to partial oral tolerance. 33 Peanut protein was not identified in our analysis. With wheat protein, gliadin was detected in 67.5–100% breast milk samples in two studies.21,34 Multiple different wheat peptides were identified in our samples, but not gliadin. Lack of these peptides may be explained by the type of milk produced in the first week after preterm birth.
Preterm milk has a higher protein content but different peptide compositions compared with term milk. 35 Milk composition may change over time and longitudinal, prospective, preterm HM proteome analyses are needed. Moreover, there is little known regarding how proteins ingested by the mother cross over from the maternal bloodstream into the breast milk and then into the infant's blood, although prior studies indicate gastrointestinal rather than mammary epithelial permeability may be playing a primary role in determining presence of food allergens in HM. 31 There is also evidence that β-lactoglobulin secretion in HM varies widely after cow's milk ingestion in mothers of infants with cow's milk allergy.20,21,36 Taken together, maternal allergy status and inter-women kinetic factors may play a role in the presence of food allergen in HM and the subsequent development of allergies in infants; however this is an area of research that needs further exploration.
Antigen-presenting cells introduce processed allergens to T-helper lymphocytes and proceed down a type 2 inflammatory pathway in allergic conditions.37,38 Thus, how allergens are cleaved in HM may be important in allergy development. We demonstrated that many bovine peptides are found digested (original protein size >40 amino acids) and free (original protein <40 amino acids), in the HM plus fortifier or formula samples, indicating that there are a variety of different parent proteins. Conversely, HM samples without fortifier had more free bovine peptides, supporting that cow milk-derived peptides in HM originated from smaller proteins. Interestingly, cow's milk allergy is one of the first to appear in infants and the majority of those are sensitized to caseins, which may be able to cross the gastrointestinal border relatively intact and larger in size, as they coagulate in acidic conditions and may be less susceptible to proteolysis.23,39 Whether our findings of smaller bovine peptides in HM are protective for the development of milk allergy warrants further investigation.
Assuming that most of these HM peptides were generated by proteases, we examined the amino acid sequence in the proteins that ended up being the substrate for cleavage. Sample J9 (pure maternal expressed breast milk) from the MAP study showed the highest protease activity. This sample was also from the mother/infant dyad pair at the lowest gestational age of all the samples (27.4 weeks). Previous proteomic studies have shown differences in the presence of proteases and protease inhibitors in term HM between allergenic and nonallergenic mothers. 40 It is thought that there is a complex interplay between allergens, proteases, and protease inhibitors in the pathogenesis of atopy. Whether the level of protease activity affects the development of allergies is an active area of research, and protease inhibitors are being evaluated as potential therapeutic agents to treat asthma and other atopic conditions. 41
There are several limitations to this study. The first is the small sample size. Our aim was to first determine the feasibility of detecting HM peptides in a variety of preterm milk samples before committing to a large-scale analysis. The second major limitation is that our subject dietary history did not include the temporal relationship of specific food ingestion and sample collection, or quantification of food consumption. Thus, secretion kinetics cannot be concluded, and contamination/inadvertent consumption is an issue with the self-reported dietary histories. In this study, we manually curated the data and attempted to delete peptides that had poor correlation with the protein databases as discussed. However, given the technical and high-throughput nature of mass spectrometry, contamination cannot absolutely be ruled out.
The sensitive nature of mass spectrometry entails the risk of identifying contaminant proteins introduced during collection, transport, or processing, and the inability to discern protein origin is an established issue with use of this technology. In addition, proteins may be present owing to contamination in the primary food source, not yet well-elucidated individual kinetics with respect to physiologic food protein metabolism and passage into the breast milk, or a combination of the above. Although it is outside the scope of this analysis, closer analysis of maternal diet and timing of consumption, along with paired serum samples, may help to determine the kinetics of HM peptides and the origin of contamination (dietary, sample collection, or via mass spectrometry) that could account for the detection of proteins that are not reported in the diet.
In our methods, technical artifact may occur because of sample carry over from chromatography and sample contamination during sample preparations, or false hits when analysis of mass spectrometry data is being carried out. However, the typical contaminant that is often detected in mass spectrometry sample analysis are keratin components and not the particular specific antigen proteins we detected. To minimize sample carry over, we performed blank runs between each sample. In this analysis, we used some of the most stringent settings for FDRs to ensure that we are not falsely detecting peptides. Certain unexpected protein sources, like yak and crayfish, may also be miscategorized by the database given evolutionary similarities and potential protein homology among species (i.e., yak and cow; and crayfish, shrimp, and dust mite, respectively).
Sample-based limitations include the lack of “pure” samples from MAP because most feeds contained maternal expressed breast milk with fortifier or pooled donor HM. This limits our ability to determine the specific source of protein antigen. However, because a large proportion of preterm infants receive supplementation, donor milk, or formula, our results reflect the real-world setting in the NICU and the infants' potential cumulative exposure. Furthermore, HMB samples did not contain donor milk or fortifiers. Finally, we did not manually blast all proteins against NCBI and Uniprot databases, only those that were positively identified, so it is possible that there were false negatives and proteins were not identified owing to inaccurate database sequences.
In conclusion, the detection of allergenic peptides and protease activity in preterm milk samples raises important questions about how exposures and nutrition in the NICU and early life may impact the development of atopy in preterm infants. Ultimately, whether HM can serve to induce allergic sensitization or tolerance in an infant is an area of research that needs much further exploration.
Data Availability
Mass Spectroscopy raw files are publicly available in the Pride Database Information for Nonhuman Peptide Sequences.
Project Name: Allergen Shedding in Human Milk from Mothers of Preterm Infants.
Project accession: PXD033473.
Footnotes
Acknowledgments
The authors thank enrolled patients and their families for their participation in the study. The authors also thank the nursing staff from both study NICUs for their assistance in sample collection.
Authors' Contributions
S.A.L., S.L.L., B.G., M.G., A.O., S.B.-T., K.L. designed the study. S.L., A.A., K.S.E., K.B., S.B.-T., D.M., K.L. collected clinical data. M.G., A.O., S.L., A.A., K.S.E., K.B., S.B.-T., D.M., K.L. processed the data and performed bioinformatic analyses. S.A.L., S.L.L., M.G., A.O., K.S.E., S.B.-T., K.L. wrote the article. S.A.L., S.L.L., B.G., M.G., A.O., K.S.E., C.C., K.B., S.B.-T., K.L. provided conceptual advice and edited the article. All authors have approved the final version of this article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was funded in part by a MOMI (Mother-Milk-Infant) Seeds pilot grant from UC San Diego's Mother-Milk-Infant Center of Research Excellence (MOMI CORE).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.