
Abstract
Cryopreservation of spermatogonial stem cells (SSCs) is an important method to restore and maintain fertility in preadolescent children suffering from cancer. For protection of SSCs from cryoinjury, various antioxidant agents have been used. The aim of this study was to assess the antiapoptotic and antioxidant effects of melatonin in frozen-thawed SSCs. SSCs were isolated from testes of neonatal mice (3–6 days old) and their purities were measured by flow cytometry with promyelocytic leukemia zinc finger protein. After culturing, the cells were frozen in two groups (1) control and (2) melatonin (100 μM) and stored for 1 month. Finally, the cell viability, colonization rate, expression of Bcl-2 and BAX gene, and intracellular reactive oxygen species (ROS) were evaluated after freezing-thawing. Melatonin increased the viability and colonization of SSCs and Bcl-2 gene expression. It also diminished BAX gene expression and intracellular ROS. The results of this study show that melatonin with antioxidant and antiapoptotic effects can be used as an additive for freezing and long-term storage of cells and infertility treatment in the clinic.
Introduction
Treatment success has recently increased considerably in young boys who have cancer before puberty and receive high doses of chemotherapy and radiotherapy. Since spermatogonial stem cells (SSCs) are very sensitive to the toxic effects of chemotherapy and radiotherapy agents, 35% of the children with cancer become infertile later in life.1,2 Therefore, it is necessary to use methods to restore and maintain fertility in these children. Cryopreservation is one of the most common methods. However, since children, contrary to adults, have no sperm for cryopreservation, it seems that extraction of SSCs from the testis tissue before treatment, cryopreservation, and long-term preservation of these cells and finally transplantation of SSCs after recovery can initiate spermatogenesis, sperm production, and fertility in these patients.3–5
Spermatogenesis is a highly coordinated and complex process that starts immediately after puberty in the seminiferous tubules with the aim of correct and accurate transfer of genetic and epigenetic information to the next generation and continues throughout life.6,7 Initiation and continuation of spermatogenesis are controlled and protected by self-renewal and differentiation of SSCs. SSCs are undifferentiated cells that, in addition to the ability of self-renewal and differentiation into daughter cells, have a significant role in producing the next generation and transferring genetic information to it, and are therefore considered unique cells among human stem cells.8–10
Although cryopreservation of SSCs is a useful method for fertility preservation in children who have cancer before puberty, this process is associated with biochemical changes of the cell plasma membrane, impaired cellular function, increased reactive oxygen species (ROS), DNA fragmentation, decreased cell activity and survival, and finally reduced quality and fertility of SSCs.11,12
Therefore, to minimize the injuries associated with cryopreservation, addition of cryoprotectants to the freezing medium is necessary. So far, different antioxidants have been used for cryoprotection.13–15 Despite the use of different cryoprotectants as antioxidant agents to the SSC freezing medium, introducing a suitable factor is still debated. Melatonin is a hormone secreted by the pineal gland in the body. It has different characteristics, including antioxidant, anti-inflammatory, and antiapoptotic properties, in different types of stem cells.16,17 The role of melatonin has been reported as an effective antioxidant agent in scavenging free radicals.18,19
There are reports of the role of melatonin as an appropriate cryoprotectant in the protection of the testis tissue and sperm.20–22 According to the above, the use of an optimum freezing method for the long-term preservation of SSCs is an important issue in the studies. This study was conducted to investigate the effect of melatonin as an antioxidant in the cryopreservation medium of SSCs for the long-term preservation of SSCs.
Materials and Methods
Maintaining the animals
Male BALB/c mice (3–6 days old) were purchased from the Pharmacology Animal Center, Tehran University of Medical Sciences. The animals were kept under standard laboratory conditions of a 12-h light/12-h dark cycle at 22°C–25°C. All animal experimental methods were approved by the Animal Ethics Committee of Tehran University of Medical Sciences.
Isolation of SSCs
SSCs were isolated using two-stage enzymatic digestion according to the method proposed by Kanatsu-Shinohara with a slight modification. 23 First, anesthesia was administered using 0.05 mg/kg ketamine (Sigma-Aldrich, St. Louis, MO); then, the removed testes were immediately transferred to dishes containing phosphate-buffered saline (PBS; Sigma-Aldrich). Under sterile conditions, the samples were washed in PBS supplemented with 1% penicillin/streptomycin (Sigma-Aldrich). In the first step of enzymatic digestion, the minced testes were transferred to a digestion medium containing 5 μg/mL DNase (Sigma-Aldrich), 1 mg/mL collagenase type IV (Gibco, CA), and 1 mg/mL hyaluronidase (Sigma-Aldrich). The cell-enzyme mixture was incubated at 37°C for 20 minutes with 5% CO2 and then pipetted gently every 5 minutes.
Centrifugation was done at 1500 g for 5 minutes to remove interstitial cells and maintain seminiferous tubules. The second stage of enzymatic digestion was performed using the same method and digestion solution. Finally, the viability of the cells was evaluated with a hemocytometer using 0.04% trypan blue.
Enrichment of SSCs
Initially, the differential plating technique was applied to separate somatic cells and enrich SSCs. 24 The cell suspension, which was collected from lectin covered dishes after 2 hours, contained purified SSCs. The purity of the collected SSCs was assessed by flow cytometry using the promyelocytic leukemia zinc finger protein (PLZF) marker. Briefly, 105 cells were incubated in 100 μL PBS/FBS and 10 μL primary antibody (1:100; ab104854; Abcam, Cambridge) at 4°C for 1 hour. After washing twice with PBS, the cells were incubated in 100 μL PBS/FBS and 10 μL secondary antibody at 4°C for 1 hour. Fluorescein isothiocyanate (FITC) goat anti-rabbit IgG H&L (ab6717; Abcam, United Kingdom) was used as the secondary antibody. The control cells were not treated with antibodies. Finally, the cells were kept in a dark room on ice and the purity percentage was determined by flow cytometry.
Culture of SSCs
Culture medium containing Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% FBS (Life Technologies), 10 ng/mL leukemia inhibitory factor (LIF; Sigma, Haverhill), 10 ng/mL basic fibroblast growth factor (Peprotech, Rocky Hill, NJ), 0.1 mM β-mercaptoethanol (Sigma-Aldrich), 10 ng/mL glial cell line-derived neurotrophic factor (GDNF; Sigma-Aldrich), 100 U/mL penicillin (Sigma-Aldrich, Darmstadt), and 100 μg/mL streptomycin (Sigma, Germany) was added to purified cells (2 × 105 cell/cm2). To reach optimal confluence, all cultures were kept in a humidified 5% CO incubator at 37°C for 2 weeks. During incubation, the medium was changed every 2–3 days. 24
Identification of melatonin optimum concentration
Before the freezing process of SSCs, the optimum concentration of melatonin for adding to the basic freezing medium was determined using the methyl thiazol tetrazolium (MTT) assay, in treatment groups at doses 50, 100, and 150 μM/mL melatonin. The MTT assay was carried out as mentioned earlier, and the optical density was measured at 540 nm using an Elisa Reader. 25
Freezing–thawing process
Freezing of SSCs was done according to a protocol described by Izadyar et al. 26 After cell collection, the cell suspension (0.5 mL) was transferred to a 1.8-mL freezing vial (Cryovial, Nunc, Denmark) and an equal volume of basic freezing medium, including 10% dimethyl sulfoxide (DMSO), DMEM, and 10% FBS, was slowly added to the cells. After complete mixing, the cryovials were rapidly and directly transferred to a −80°C freezer overnight. Then, the frozen vials were immersed in liquid nitrogen for 1 month.
The treated group was frozen in the basic freezing medium containing 100 μM/mL melatonin, while the control group was frozen in the basic freezing medium without melatonin. For thawing, the cryovials were kept at room temperature for 30 seconds and then quickly placed in a 37°C water bath for 2 minutes. The thawed cells were transferred to a falcon tube containing DMEM and 10% FBS. Finally, cells were centrifuged at 1500 g for 5 minutes. After discarding the supernatant solution, the cell pellet was cultured to perform further techniques.
Assessment of cell viability in frozen-thawed SSCs
To define the effect of melatonin (100 μM) on cell viability, the MTT assay was performed on cultured cells after the freezing–thawing process. First, 100 μL DMEM and 10 μL MTT were added to SSCs (2 × 105 cells/mL) in each well of 96-well plates, and then the mixture was incubated at 37°C for 4 hours. Then, the MTT solution in each plate was replaced with 100 mL DMSO (Sigma, Germany). The optical density was measured at 540 nm using an ELISA reader (BioTek Instruments).
Assessment of intracellular ROS in frozen-thawed SSCs
To measure intracellular ROS, 2′,7′-dichlorofluorescein diacetate (DCFH-DA; Sigma-Aldrich) was used as a fluorescent oxidative probe. The cell suspension was incubated with 10 μL of 2′,7′-dichlorofluorescin diacetate at 37°C for 25 minutes. Next, the fluorescence intensity of DCFH-DA was assessed between 500 and 530 nm wavelengths using a BD FACScan flow cytometer (Becton Dickinson, San Jose, CA).
Assessment of malondialdehyde and superoxide dismutase in frozen-thawed SSCs
The malondialdehyde (MDA) assay kit (Zellbio GmbH, Germany) with thiobarbituric acid (TBA) reaction was used to measure the lipid peroxidation level in SSCs. It was performed according to the manufacturer's instructions. Briefly, the reagents were added to SSCs and the prepared mixture was then heated in a boiling water bath for 1 hour. After pipetting the supernatant into the microplate, the absorbance of the pink color product was read at 535 nm to determine the level of MDA.
The superoxide dismutase (SOD) level of SSCs was measured using the SOD assay kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer's guidelines. The absorbance was also measured at 550 nm using an enzyme-linked immunosorbent assay plate reader.
Assessment of PLZF and ID4 proteins expression in frozen-thawed SSCs
Immunocytochemistry was done to determine the effect of melatonin (100 μM) on markers related to the proliferation of SSCs. First, the SSCs were fixed with paraformaldehyde 4% (Sigma-Aldrich) and permeabilized with 0.1% Triton X-100 (Sigma-Aldrich) followed by blocking for 1 hour with 10% goat serum (Sigma-Aldrich). Then, the samples were incubated with inhibitor of DNA binding 4 (ID4) (1:100; PA5-26976; Invitrogen) and PLZF (1:100; ab104854; Abcam, Cambridge) antibodies overnight, followed by a 2-hour exposure to secondary antibody FITC (ab6717; Abcam, United Kingdom). In addition, the nuclei were stained with 4,6-diamid-ino-2-phenylindole (DAPI, 1 g/mL. Finally, the slides were observed under a fluorescence microscope (Olympus LX71, Japan).
Assessment of BAX and Bcl-2 protein expression in frozen-thawed SSCs
Protein extraction from SSCs was done using TriPure Isolation Reagent (Roche, Germany). Electrophoresis was performed to separate 20 μg of the total protein for each sample. Then, the proteins were transferred into 10.5% and 12.5% gradient sodium dodecyl sulfate (SDS)–polyacrylamide gel (BioRad Laboratories, Hercules, CA) and polyvinylidene difluoride (PVDF) membrane (Roche) and blocked with 5% nonfat dry milk (Carnation, CA). Subsequently, the samples were incubated with suitable primary antibodies against BAX (sc23959, sec mouse: sc516102) (1:1000) and Bcl2 (sc7382, sec mouse: sc516102) (1:1000) at 4°C overnight. A nitrocellulose membrane was incubated for 2 hours after adding appropriate secondary antibodies (HRP conjugated goat anti-rabbit) (Abcam). Finally, the expression of the proteins was assessed using enhanced chemiluminescence.
Assessment of SSCs colonization in frozen-thawed SSCs
After the freezing–thawing process, SSCs (2 × 105 cell/cm2) were cultured in DMEM containing 10% FBS, 100 U/mL penicillin, 100 mg/mL streptomycin, 10 ng/mL LIF, and 10 mg/mL GDNF in 5% CO2 at 37°C for 1 week. The SSC colonies were observed and captured using an inverted microscope. Then, the number and diameter of the colonies were assessed using the Image J software.
Statistical analysis
The results were analyzed using the GraphPad Prism 7.0 using one-way ANOVA and the Tukey post hoc tests. p-Values <0.05 were considered statistically significant (p ≤ 0.05). The data are expressed as mean ± standard deviation.
Results
Determining the purity of SSCs
After the separation of SSCs of 4–5 neonatal mice (BALB/C), enzymatic digestion, and differential plating, the purity of SSCs (2 × 105 cell/cm2) was investigated with flow cytometry using PLZF as a marker for undifferentiated cells. As expected, 98.32% of the cells were positive for PLZF (Fig. 1).
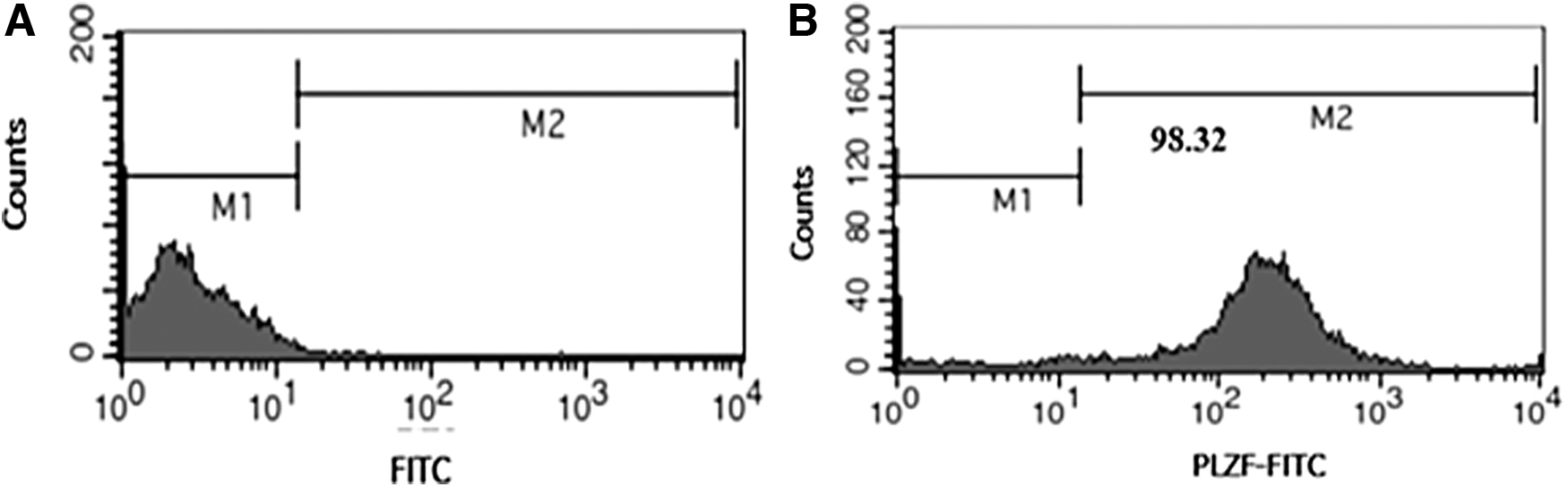
Results of flow cytometry indicating the purity percentage of SSCs with PLZF marker. M1: marker-negative cells, M2: marker-positive cells.
Identification of optimum concentration of melatonin
The MTT assay was used to determine the optimum concentration of melatonin for adding to the basic freezing medium. The results indicated that the survival rate was significantly higher in the SSCs treated with 100 μM melatonin (95.83% ± 0.88) compared to the control group (65.40% ± 0.06) and other melatonin groups at doses of 50 μM (68.13% ± 0.98) and 150 μM (62.90% ± 0.86) (p < 0.001). The survival rate in the melatonin group of 100 μM was similar to the fresh group (98.40% ± 0.26) (p < 0.001). Thus, 100 μM melatonin was considered the optimum concentration (Fig. 2).
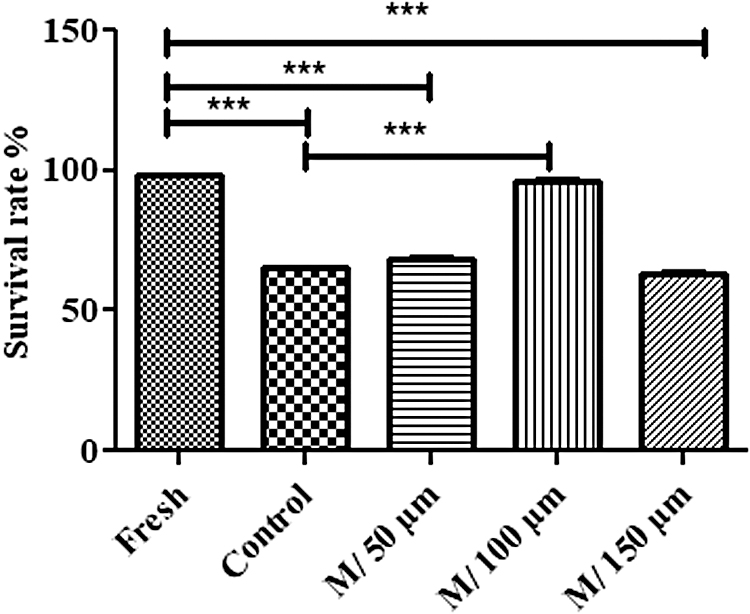
Determining the optimum concentration of melatonin by MTT assay. According to these results, 100 μM of melatonin was selected as the optimum concentration. The experiment was performed in triplicate and repeated thrice. The results are reported as mean ± SE. ***p < 0.001. MTT, methyl thiazol tetrazolium.
Effect of melatonin on viability in frozen-thawed SSCs
The results of the MTT assay (Sigma-Aldrich) technique showed that the freezing–thawing process markedly decreased the viability of SSCs in the control group (65.17 ± 0.31) compared to the fresh group (98 ± 0.15) (p < 0.001). The results also showed that the viability of SSCs frozen with melatonin (100 μM) was significantly higher (96.77 ± 0.18) compared to the control group (p < 0.001) (Fig. 3).
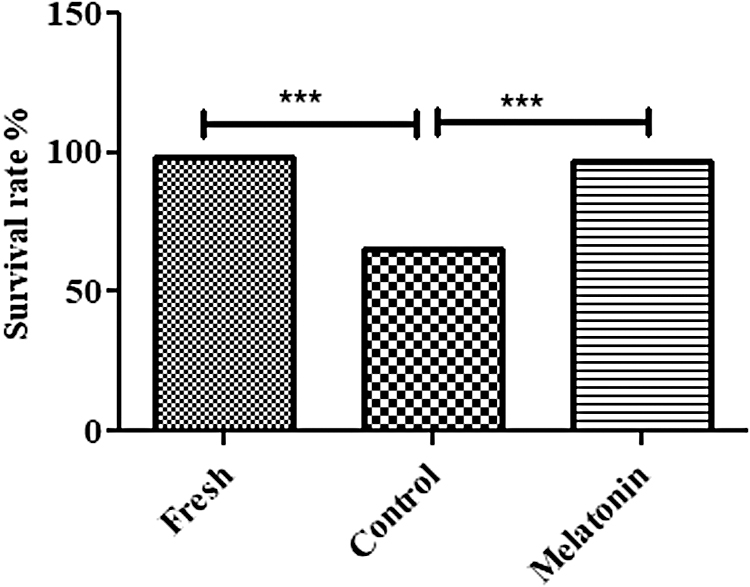
Analysis of cell viability after the thawing process by using the MTT. Viability percentage in melatonin group was significantly more than other groups and close to the fresh group. The experiment was performed in triplicate and repeated thrice (there are 2 × 105 cells/cm2 of SSCs in each group). The results are reported as mean ± SE, ***p < 0.001.
Effect of melatonin on intracellular ROS in frozen-thawed SSCs
The level of intracellular ROS was assayed in fresh, control (cryopreservation), and cryopreservation with melatonin groups using flow cytometry after thawing. The results of this experiment revealed that the freezing–thawing process significantly increased the intracellular ROS production in the control group (48.50% ± 0.28) versus the fresh group (22.87% ± 0.78) (p < 0.001). The level of ROS production decreased significantly in the cryopreservation group with melatonin (100 μM) (24.83% ± 0.12) compared to the control group (p < 0.001) (Fig. 4).
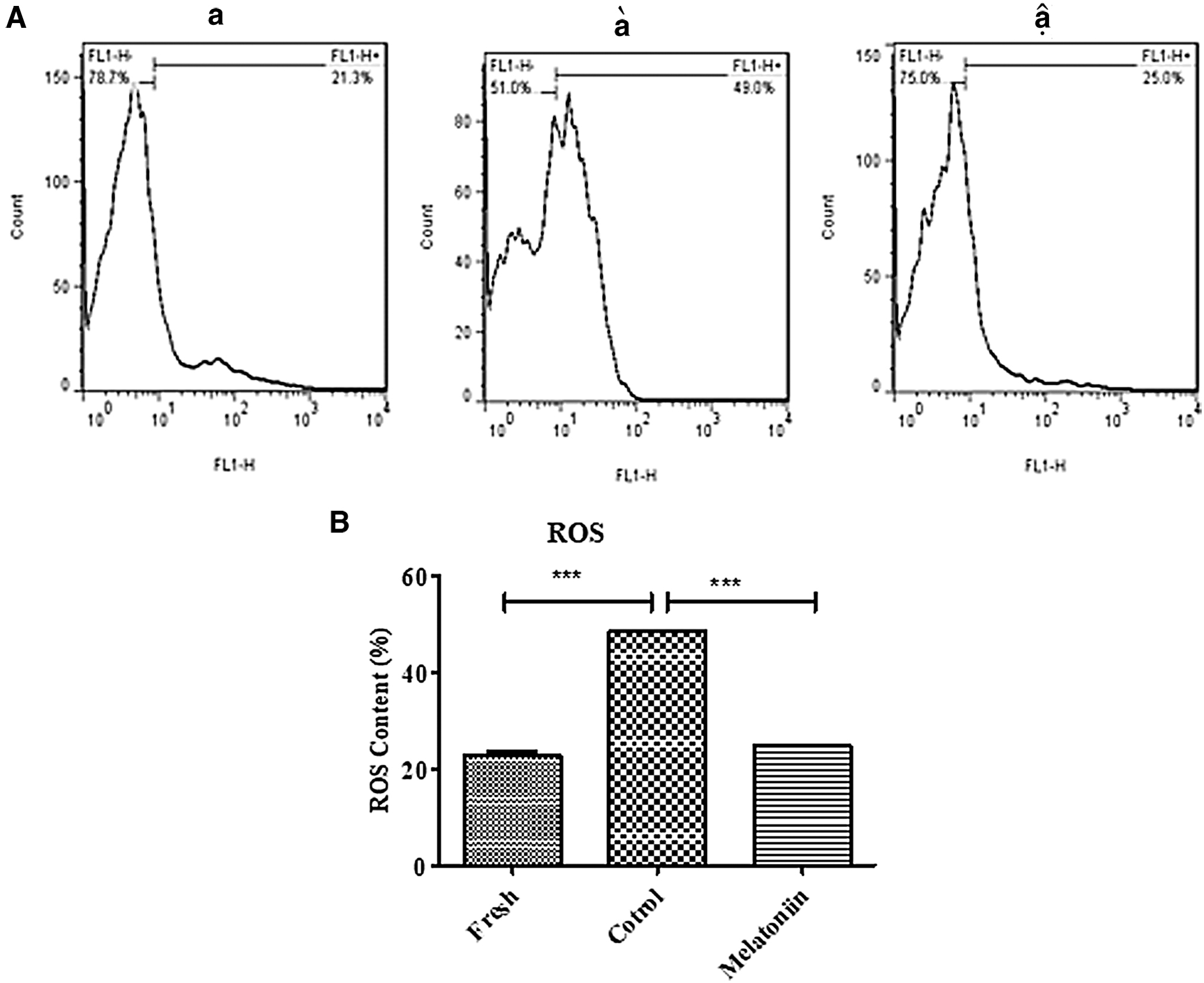
Effect of melatonin on levels of MDA and SOD in frozen-thawed SSCs
At this stage, the levels of MDA and SOD were evaluated in three groups of SSCs, including fresh, control (cryopreservation), and cryopreservation with melatonin using flow cytometry after freezing–thawing. The level of MDA production, as a lipid peroxidation end product, was measured using the TBA method. The level of MDA increased significantly in the control group (202.6 μM ± 4.35) compared to the fresh group (152.3 μM ± 8.25) (p < 0.01). The results indicated that the freezing medium containing melatonin (100 μM) could effectively decrease (145.4 μM ± 9.94) the level of MDA compared to the control group (p < 0.01) (Fig. 5A).
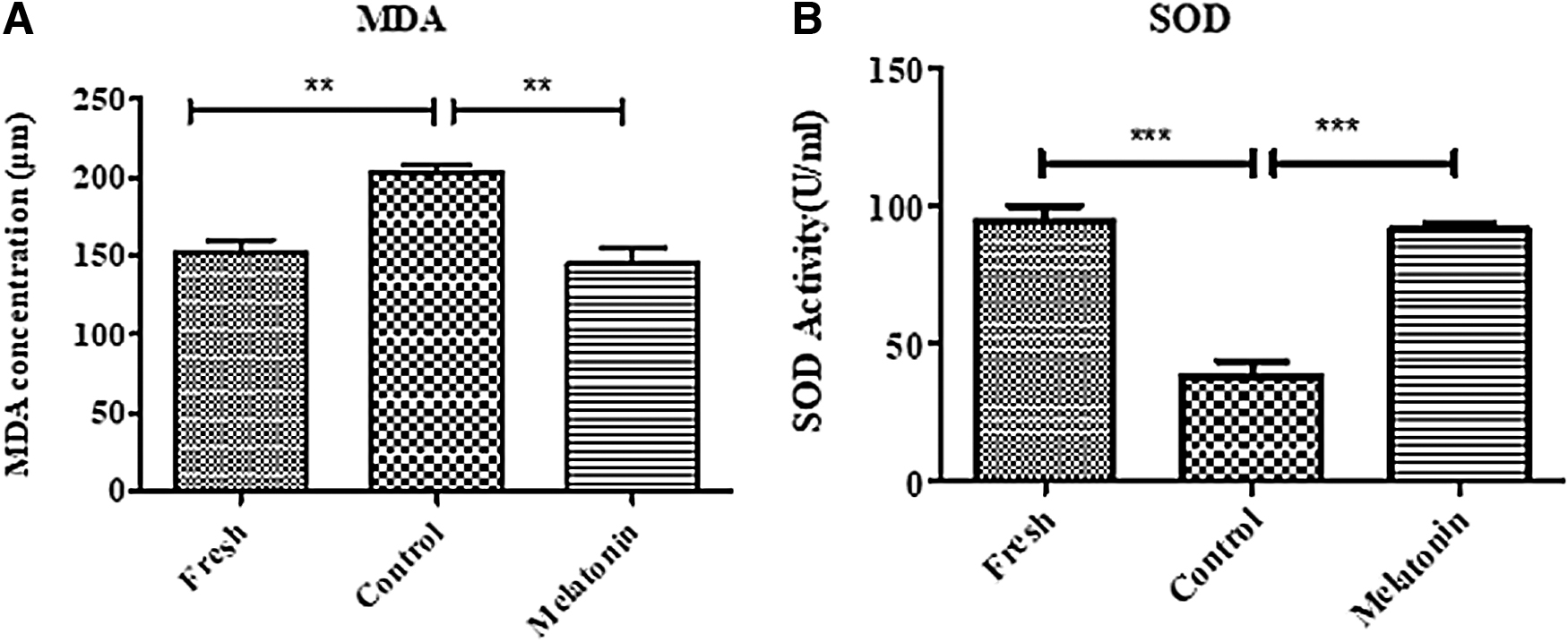
Analysis of MDA
The results of SOD evaluation showed that the level of SOD decreased significantly in the control group (38.48 U/mL ±5.52) compared to the fresh group (94.33 U/mL ±6.15) (p < 0.001). Moreover, melatonin (100 μM) could effectively increase (91.43 U/mL ±2.42) the level of SOD compared to the control group (p < 0.001; Fig. 5B).
Effect of melatonin on colonization of frozen-thawed SSCs
One week after culturing the thawed SSCs, the images of the colonies were captured using an inverted microscope and the diameter and number of colonies were measured using the Image J software (Fig. 6A). Data analysis showed that melatonin (100 μM) could effectively increase (195.9 μm ± 6.06) the diameter of SSC colonies in comparison with the control group (97.59 μm ± 1.62) (p ≤ 0.0001). The number of colonies was significantly larger in SSCs treated with melatonin (100 μM) (4.66 numb ±0.33) compared to the control group (2.33 numb ±0.33) (p < 0.01) (Fig. 6B, C).
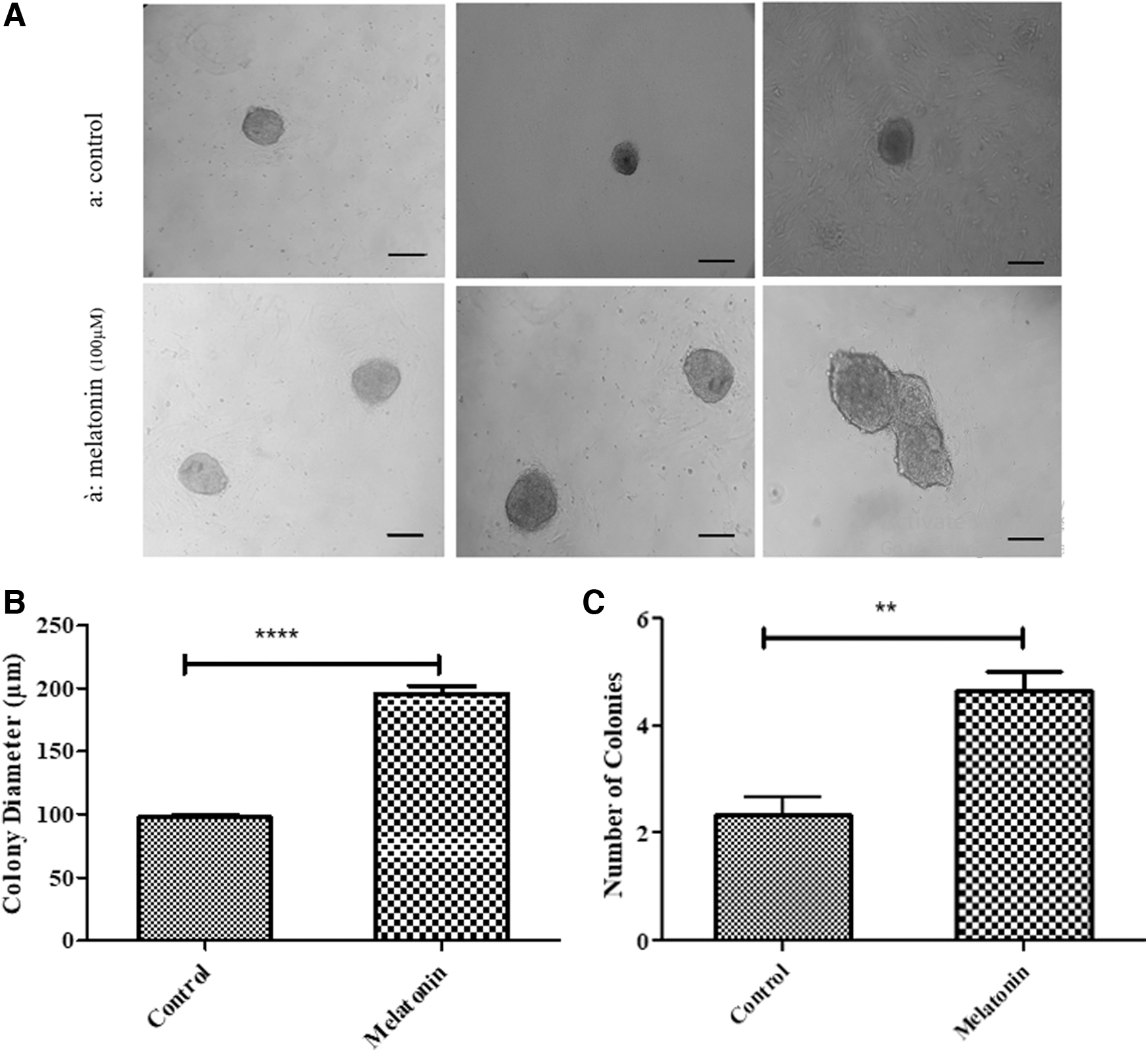
(
Effect of melatonin on expression of BAX and Bcl-2 in frozen-thawed SSCs
Western blot analysis was performed to investigate the expression of proteins related to apoptosis after culturing the thawed SSCs. According to the results, cryopreservation with melatonin (100 μM) significantly increased (1.45 ± 0.06) the expression of Bcl-2 protein relative to the control group (1.09 ± 0.06) (p ≤ 0.05) (Fig. 7A, B). Analysis of the BAX protein also revealed a marked reduction in the melatonin (100 μM) group (0.35 ± 0.02) versus the control group (0.47 ± 0.02) (p ≤ 0.05) (Fig. 7C, D).
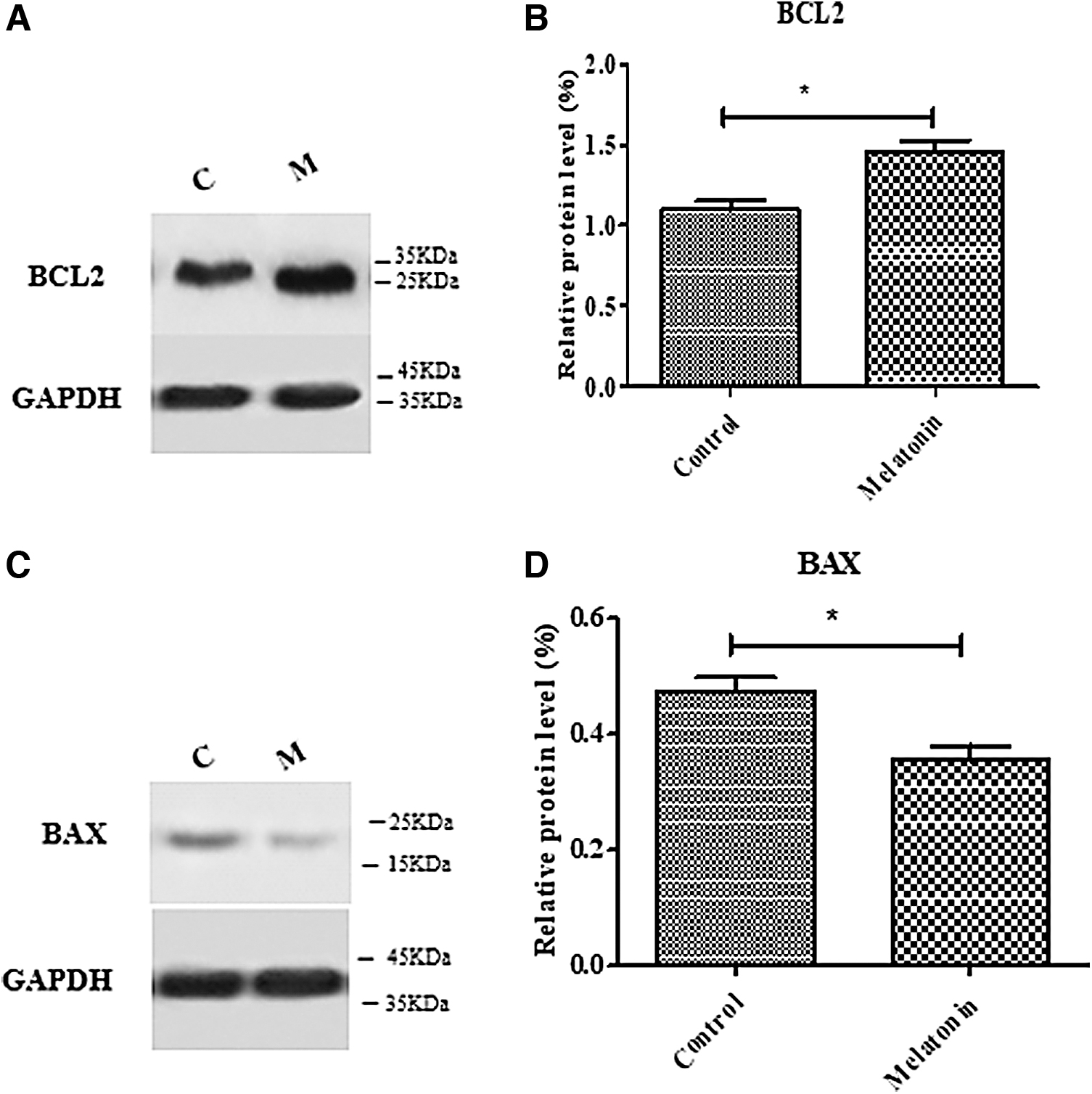
Effect of melatonin on expression of ID4 and PLZF in frozen-thawed SSCs
The expression of ID4 and PLZF proteins as markers of proliferation was evaluated in SSCs after culturing the thawed SSCs using immunocytochemistry. The results of immunocytochemistry analysis indicated that the percentage of ID4 expression increased significantly in SSCs treated with melatonin (100 μM) (72.93% ± 1.85) compared to control cells (41.01% ± 4.14) (p < 0.01) (Fig. 8A, B). Similarly, the expression of the PLZF proliferation marker also increased significantly in the cryopreservation group with melatonin (100 μM) (65.70% ± 2.32) compared to the control group (38.73% ± 1.83) (p < 0.001) (Fig. 8C, D). Collectively, the results indicated that melatonin (100 μM) induced the proliferation process in mouse SSCs after the freezing–thawing process.
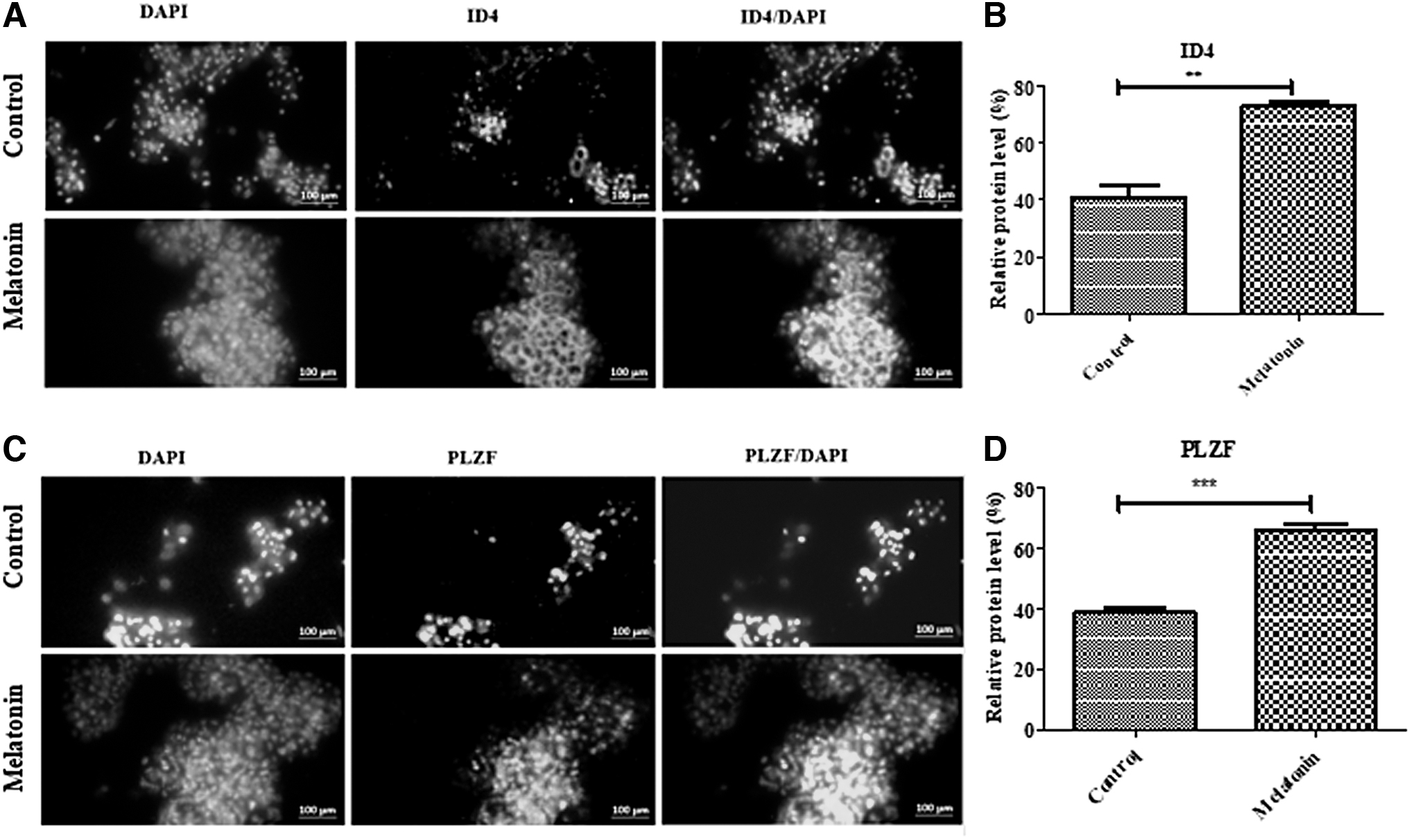
Discussion
The culture and cryopreservation of SSCs as an appropriate technique for long-term preservation of SSCs and restoration of fertility in young men undergoing cancer treatment has been the subject of a number of studies.27,28 The normal function of SSCs decreases after the freeze-thaw process due to cryoinjuries. Therefore, it is necessary to add cryoprotectants to the freezing medium of SSCs.25,29 In this study, melatonin was used in the freezing medium as an antioxidant and a cryoprotectant to reduce cryoinjuries and their effect on the function of cells after the freeze-thaw process was assessed.
PLZF, as one of the most common markers of SSCs, has an important role in self-renewal regulation of SSCs.15,30,31 After enzymatic digestion and isolation of SSCs, flow cytometry with the PLZF marker was used to identify SSCs. In this study, flow cytometry data showed that 98.32% of the cells expressed PLZF, which was consistent with previous studies.32,33 As reported by other studies, the high percentage of PLZF expression in these cells indicated that the isolated cells were in the early stages of development and the differentiation process was not started yet.
Although cryopreservation is the best method for long-term preservation of stem cells, it results in the formation of intracellular ice crystals and cell injury without the use of proper cryoprotectants.34,35 Cryoprotectants used to reduce cryoinjury are divided into intracellular and extracellular cryoprotectants. According to reports, intracellular cryoprotectants with a low-molecular weight prevent cell injury through penetrating the cell membrane and extracellular cryoprotectants prevent cell dehydration during cryopreservation by producing a sheath around the cells. Therefore, cryoprotectants use two different mechanisms to reduce cryoinjury. 36
In this study, the results of adding melatonin to the freezing medium showed that cryopreservation of SSCs with 100 μM melatonin improved cell viability compared to the control group. Another study investigating the role of melatonin as a cryoprotectant also found that melatonin improved cell viability after thawing.20,37,38 The exact mechanism by which melatonin protects cells from freezing injury is unclear, but in this study, cryopreservation of SSCs with melatonin resulted in higher viability and proliferation rate and lower apoptosis rate compared to the control group, so we speculate that it may delay extracellular ice crystallization, thereby reducing freezing injury. 36
Freezing the cell at very low temperatures increases the ROS production. Cell injuries like lipid peroxidation, protein oxidation, and nucleic acid injury occur when ROS production exceeds the cell physiological level. It has been reported that adding different antioxidants to the freezing medium can reduce ROS production.34,39,40 For this reason, melatonin was added to the freezing medium to reduce ROS production in this study. The results showed a marked reduction in intracellular ROS production in the freezing medium containing melatonin compared to the control group. Several studies investigating the role of different antioxidants on the intracellular ROS production rate have found a reduction in intracellular ROS production after the freeze-thaw process.13–15
Interestingly, the results of this study regarding intracellular ROS production were more favorable compared to previous studies. Moreover, many researchers found that melatonin, as a potent antioxidant, could reduce oxidative stress and protect the sperm against cryoinjury through scavenging the produced ROS.41,42 In fact, it can be stated that melatonin helps to eliminate ROS, probably through neutralizing toxic free radicals and increasing antioxidant enzymes, including SOD.
The results of this study regarding the antiapoptotic effect of melatonin in the freezing medium showed a significant increase in the expression of the BCL2 antiapoptotic gene and a significant decrease in the BAX apoptotic gene in the melatonin group. Deng et al. found that melatonin reduced apoptosis through increasing the expression of BCL2 and reducing the expression of BAX in sheep spermatogonial cells. 43 Furthermore, another study found that melatonin treatment markedly increased the expression of the BCL2 gene and reduced the expression of the BAX gene in cultured bovine fetus. 44 In addition, some studies found that adding antioxidants like vitamin E, catalase, α-tocopherol, and quercetin to the freezing environment was effective in reducing the rate of mouse SSC apoptosis.13,14,34
Molecular studies suggest that the PLZF and ID4 markers have an important role in regulation of self-renewal and proliferation of SSCs.45–47 The exact mechanism of PLZF in self-renewal regulation of SSCs is not clear, but it has been hypothesized that PLZF inhibits the expression of c-kit (differentiation gene) directly and indirectly. 48 C-kit is a class III receptor tyrosine kinase that is only expressed in differentiated SSCs and is considered a specific marker for differentiation of these cells.10,46,47 In this study, the effect of melatonin on proliferation of SSCs was assessed using PLZF and ID4. The results showed that melatonin increased the expression of PLZF and ID4 proteins in the cells after the freeze-thaw process. A study by Navid et al. also confirmed the effect of melatonin on the proliferation of these cells. 19
Melatonin plays a role in the regulation of many biological activities of the reproductive cells through receptor-dependent and receptor-independent mechanisms. 22 The MT1 receptors are found in some reproductive system structures. For this reason, several studies have evaluated the melatonin membrane receptor-dependent pathway in the testes and ovaries. In the receptor-independent pathway, melatonin directly eliminates free radicals and neutralizes hydroxyl (OH•) and peroxynitrite anion (–ONOO) as very toxic agents. 49
In addition, metabolites like cyclic 3-hydroxymelatonin, acetyl-N 2 -formyl-5-methoxykynuramine (AFMK) and N¹-acetyl-5-methoxykynuramine (AMK), which are produced as a result of the integration between melatonin and free radicals, are capable of eliminating free radicals. 50 In general, we believe that melatonin, as a cryoprotectant, protects SSCs from cryoinjuries. Although the melatonin exact mechanism is not clear, we believe that melatonin, as a cryoprotectant, may facilitate the elimination of free radicals through a mechanism independent of receptors. The results of this study report on antioxidant and antiapoptotic effects of melatonin. 51
According to the results of this study, melatonin may be a suitable candidate for freezing, long-term cell preservation, and infertility treatment in the clinical setting, considering its antioxidant and antiapoptotic properties, as well its effects on increased colonization and cell viability of SSCs.
Conclusion
According to the results of this study, it can be concluded that melatonin as a cryoprotectant with antioxidant and antiapoptotic effects can be used for freezing and long-term storage of cells and infertility treatment in the clinic.
Footnotes
Authors' Contributions
M.K. and A.H. designed experiment and M.K. and T.N. performed cell culture. M.K., S.K., and T.N. performed Western blot and M.K. and M.S. performed analyses. M.K. and S.S. edited article. M.K. supervised the research and all authors read and approved the final article.
Ethical Approval and Consent to Participate
This article does not contain any study with human participants performed by any of the authors. All experiments involving the use of animals were approved by the Institutional Animal Care and Use Committee (IACUC) of the Tehran University of Medical Science. All applicable international, national, and institutional guidelines for the care and use of animals were followed.
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.