
Abstract
Despite the vital role of seminal plasma (SP) in maintaining sperm function and aiding gamete interaction in many species, SP is usually removed before cryopreservation of stallion sperm to improve cryosurvival of sperm. The present study assessed if the vital sperm functional parameters of genetically superior stallions producing poor quality semen can be enhanced by the supplementation of heterologous SP from the stallion producing high quality semen. Spermatozoa from poor quality semen producing stallions were divided into three aliquots: two aliquots were supplemented with SP obtained from good quality semen producing stallions at the rate of 20% and 30%, respectively, whereas the third aliquot remained as control (0% SP) and incubated at 37°C for 30 minutes. Sperm membrane integrity, mitochondrial membrane potential (MMP), mitochondrial superoxide (mtROS) generation, and intracellular calcium status were assessed at different time intervals during incubation by flow cytometry. It was observed that the dead sperm population increased (p < 0.01) during incubation in both the 20% and 30% SP-supplemented groups. However, no significant changes were observed in MMP in both the control and treatment groups at different time intervals. Interestingly, it was found that sperm mtROS production increased (p < 0.01) during incubation in the SP-supplemented groups compared with the control group. The proportion of live spermatozoa with high intracellular calcium was reduced (p < 0.01) during incubation in the SP-incubated groups. Collectively, heterologous SP addition could not repair the damages caused by the cryopreservation and further resulted in deterioration of semen quality as observed in our study by reducing viability, increasing reactive oxygen species (ROS) production possibly due to high proportion of dead cells, or some factors (yet to be identified) that are inducive of oxidative stress in stallion spermatozoa.
Introduction
It is almost inescapable to encounter substandard quality semen production from breeding stallions since the breeding stallions are selected based on their individual athletic performances and pedigree, rather than their reproductive potential. 1 It has been found that only 20% of the breeding stallions produce semen with good freezability. 2 This is mainly due to higher levels of docosapentaenoic acid, an omega-6 polyunsaturated fatty acids (PUFA) present in plasma membrane of stallion sperm, 3 which makes them susceptible to oxidative damage during cryopreservation. Therefore, the plasma membrane is the primary place of injury during freezing and thawing.
The ultrastructural alterations in the plasma membrane during cryopreservation are not completely reversible on rewarming. 4 Additionally, the reorganization of sperm membrane lipids disrupts the lipid–protein and lipid–lipid interactions essential for the sperm to interact normally with cells of the female reproductive tract and reduces the motility and fertility.5,6 Thus, the possible ways to improve the post-thaw semen quality will hold importance in stallion breeding industry. Various attempts can be made to improve post-thaw quality; one such is the addition of seminal plasma (SP) to the cryopreserved spermatozoa.
SP is an integral component of semen, which is the conglomeration of secretions from the epididymis and accessory sex glands. Detailed analysis of the molecular composition of SP revealed the presence of proteins, ions, lipids, free amino acids, monosaccharides, polyamines, and hormones.7,8 SP plays a significant role in protecting the spermatozoa from harsh environments in the female reproductive tract, prevents premature capacitation (decapacitation factors), provides nutrition, serves as a transportation vehicle for spermatozoa, and influences gamete interaction.9–11
In mares susceptible to persistent breeding-induced endometritis, the addition of minute quantities of SP in insemination dose may be needed for obtaining optimum fertility that implies immunoregulatory and immunomodulating effects of SP on maternal immune system. 12 However, before cryopreservation, most of the SP is removed by centrifugation in order to eliminate its detrimental effects on sperm survival. 12 Despite removal of SP, the sperm survival following thawing and motility remains highly variable, which might be reflective of inherent individual or breed differences as well as absence of some SP-derived moieties that are crucial for the normal sperm functionality.
Owing to the critical role of SP in maintaining normal functionality of spermatozoa during transportation in the male and female reproductive tract, numerous studies have been carried out to incorporate SP in semen before cryopreservation as well as in frozen–thawed semen.13–15 However, the results are not consistent and highly variable. While few studies found that supplementation of SP had beneficial effects on cryosurvival,11–14 other studies reported that SP addition had no beneficial effects on sperm.15,16 The variations among these studies might be explained by the source of SP.
It is reported that proteomic composition of SP from high-fertile males differed significantly from that of the low-fertile males.17–19 During the process of centrifugation of semen to obtain sperm pellet, SP is removed and some of the SP proteins present on sperm surface are also removed. Therefore, if known quantities of SP from stallion producing high quality semen are incorporated after freezing and thawing of stallion spermatozoa, it might be possible to improve the post-thaw sperm quality. In this regard, we hypothesized that the vital sperm functional parameters of genetically superior stallions producing poor quality semen can be enhanced by the supplementation of heterologous SP from the stallion producing high quality semen.
Materials and Methods
The current study was conducted at the Theriogenology Laboratory, Southern Regional Station of ICAR-National Dairy Research Institute, Bengaluru, Karnataka, India. Prior approval of the Institutional Animal Ethics Committee (Ethical approval No. IXX15812) was obtained for all the experimental procedures and performed in accordance with relevant guidelines and regulations.
Source of semen and sperm preparation
Semen samples used in the present experiment were collected from six adult stallions (high genetic merit with poor quality semen; age 4–6 years) that were maintained at Equine Production Campus, ICAR-National Research Centre on Equines, Bikaner, Rajasthan, India. No separate provision for the light or feed has been provided to the stallions involved in the study. Semen collection was performed early in the morning hours using a mare in estrus as a dummy. Semen collection and sperm preparation methods were followed as previously described by Soni et al., 20 with little modification. Semen was collected using a Colorado model artificial vagina lubricated with liquid paraffin, prewarmed to 45°C–50°C, and fitted with an inline filter to separate the gel fraction. To reduce the extragonadal sperm reserves, semen was collected over a period of four consecutive days before the commencement of the actual experiment.
The stallions were given 2 days of sexual rest before the actual commencement of the experiment. After that, two ejaculates were collected from each stallion. Immediately after collection, ejaculates were transported to the laboratory and kept at 37°C until processing. Sperm motility and concentration parameters were evaluated for all the six stallions. Sperm motility was <40% in poor quality semen producing stallions, whereas it was >70% in stallions producing high quality semen. The semen samples were centrifuged at 650 g for 5 minutes to obtain the sperm pellet, and the supernatant was discarded. The sperm pellet was further extended in a secondary freezing extender (6.0 g glucose, 11.00 g lactose, 0.37 g EDTA, 0.37 g sodium citrate, 0.12 g sodium bicarbonate, 10% egg yolk, 100,000 IU penicillin, and 0.10 g streptomycin) containing the 5% of dimethyl formamide as cryoprotectant. The extended semen was subjected to cooling for 3 hours in a cooling cabinet at 4°C, and the semen was cryopreserved as per the protocols as previously described. 20
SP preparation
SP was obtained from three stallions that consistently produced good quality semen with a greater cryotolerance of spermatozoa. The SP was prepared as per the protocols previously described by Talluri et al., 8 with slight modifications. Briefly, immediately after semen collection, the gel portion of the semen was removed through a sterilized gauze filter. The remaining gel-free semen was immediately centrifuged at 3000 g for 15 minutes to separate the SP from the spermatozoa, and the supernatant was aspirated into sterile tubes. The harvested SP was checked again under microscope for the presence of any spermatozoa; if spermatozoa were detected, then the samples were recentrifuged at 4500 g and the procedure was repeated until the SP became free of spermatozoa. Sperm-free SP was filtered into 15 mL tubes using a 0.2 μm membrane filter before further use.
Sperm preparation and treatment distribution
Sperm preparation for flow cytometry analysis was performed as per Kumaresan et al. 21 Cryopreserved semen straws were thawed at 37°C for 30 seconds and assessed for individual motility. Semen samples were washed using sperm-Tyrode's Albumen Lactate Pyruvate (TALP) medium (3.1 mM KCl, 100 mM NaCl, 0.29 mM NaH2PO4, 25 mM NaHCO3, 2.0 mM CaCl2, 21.6 mM C3H5NaO3, and 1.5 mM MgCl2) for two times at 300 g for 3 minutes before experiments. Spermatozoa from each ejaculate were divided into three aliquots. The first aliquot was considered as control (with no addition of SP), whereas to the second and third aliquots, SP was supplemented at 20% and 30%, respectively, and incubated at 37°C. Different sperm phenotypic characteristics were evaluated using flow cytometry in a time-dependent manner (0, 10, and 30 minutes).
Flow cytometric analysis of sperm phenotypic characteristics
Flow cytometry was used to assess various sperm functional attributes, namely viability, mitochondrial membrane potential (MMP), mitochondrial superoxide (mtROS) generation, and intracellular calcium status. Representative flow cytometry plots of these parameters are shown in Supplementary Figure S1. All the flow cytometry-based sperm functional parameter assessments were performed using CytoFLEX-S flow cytometer (Beckman Coulter Life Sciences, Indianapolis, IN) equipped with near UV (375 nm), blue (488 nm), and red (638) laser. The fluorescence of SYBR-14, Fluo-3, and JC-1 monomer was detected with a band-pass filter 525/40 nm, and the fluorescence of propidium iodide (PI), MitoSOX Red, and JC-1 aggregate was measured using a band-pass filter of 585/42 nm. A minimum of 10,000 events were assessed for each sample. Data acquisitions and analyses were made using CytExpert software (Beckman Coulter Life Sciences).
Plasma membrane integrity
Sperm plasma membrane integrity was assessed using a combination of fluorochromes SYBR-14 and PI (Invitrogen, Thermo Fisher Scientific) as per the procedure given by Kumaresan et al. 21 The SYBR-14 working solution was prepared by diluting the stock in dimethyl sulfoxide (1:50), and a volume of 1.2 μL was added to 2 × 106 cells suspended in sperm TALP. The sperm suspension was incubated at 37°C for 10 minutes in the dark, followed by the addition of 1 μL of PI (2.4 mM). The samples were subjected to flow cytometric analysis after 2 minutes of PI addition. Both the membrane intact and membrane-compromised spermatozoa were assessed and are expressed in percentage.
Mitochondrial membrane potential
MMP of spermatozoa was assessed using 5,5,6,6′-tetrachloro-1,1′,3,3′ tetraethylbenzimi-dazoylcarbocyanine iodide (JC-1) fluorochrome (Invitrogen, Thermo Fischer Scientific) as described by Mocé and Graham, 22 with minor modifications. Briefly, 5.6 μL of 0.2 mM JC-1 solution was added to 2 million spermatozoa in 300 μL sp-TALP and incubated for 30 minutes at 37°C in the dark, followed by analyzing in a flow cytometer. The proportion of spermatozoa with high and low MMP was assessed.
mtROS generation
Mitochondria-specific reactive oxygen generation (mtROS) was estimated using mitochondria-specific superoxide fluorescent probe MitoSOX Red (MitoSOX) (Thermo Fischer Scientific), as previously described by Kumaresan et al. 21 To 2 × 106 sperm in 300 μL sp-TALP suspension, 1 μL of MitoSOX Red (500 μM) dye was added and incubated for 30 minutes at 37°C in the dark. Thereafter, flow cytometric assessment of the samples was carried out.
Intracellular calcium status
Intracellular calcium concentrations in spermatozoa were assessed using Fluo-3 AM dye (Thermo Fischer Scientific), as described by Nag et al. 23 Briefly, 3 μL of Fluo-3 was added to 2 × 106 sperm in 300 μL sp-TALP suspension and incubated at 37°C in dark condition for 30 minutes. Following incubation, 1 μL of PI (2.4 mM) was added and the sample was further incubated for 2 minutes before subjecting to flow cytometric analysis. The sperm population was classified as live and dead spermatozoa with high and low calcium levels.
Statistical analyses
All the statistical analyses were performed using GraphPad Prism V8.4.3. Mixed effects model analysis of variance was used to identify the best combination of SP concentration and incubation time for significant difference in sperm functional attributes between the control and treatment groups. Tukey's multiple comparison test was carried out to study the effect of addition of SP on various seminal parameters.
Results
The results for the effects of different doses of SP in various sperm functional quality at various time intervals are shown in Figures 1–4.
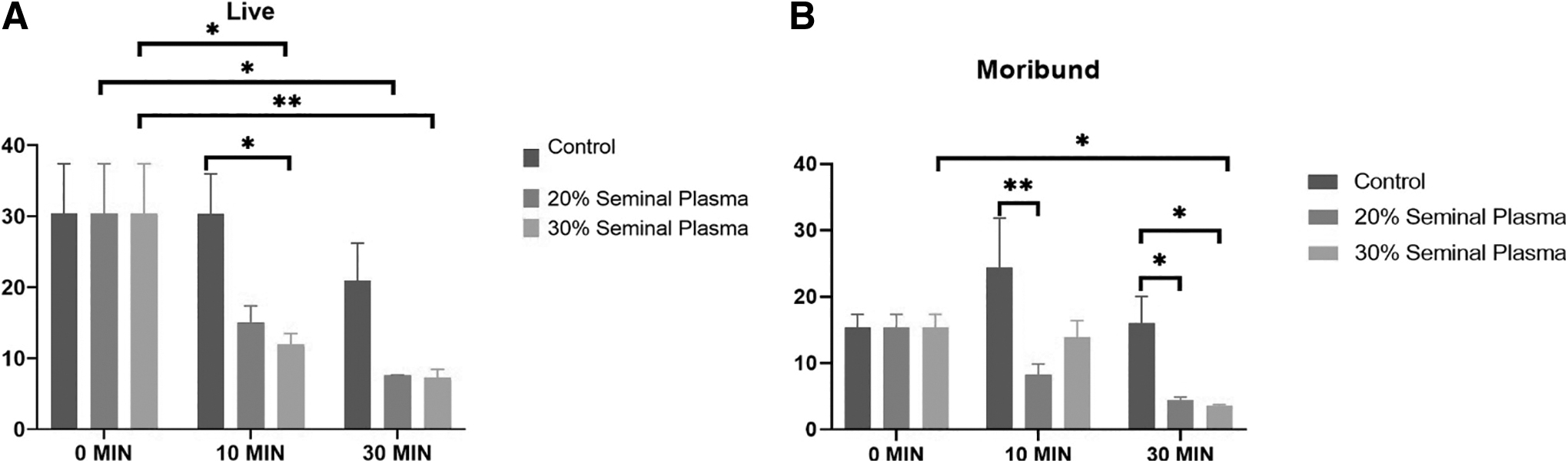
Dynamics of stallion sperm viability up on supplementation of the SP (Control, 20%, and 30%) at various time intervals (0, 10, and 30 minutes) to the post-thawed stallion semen.
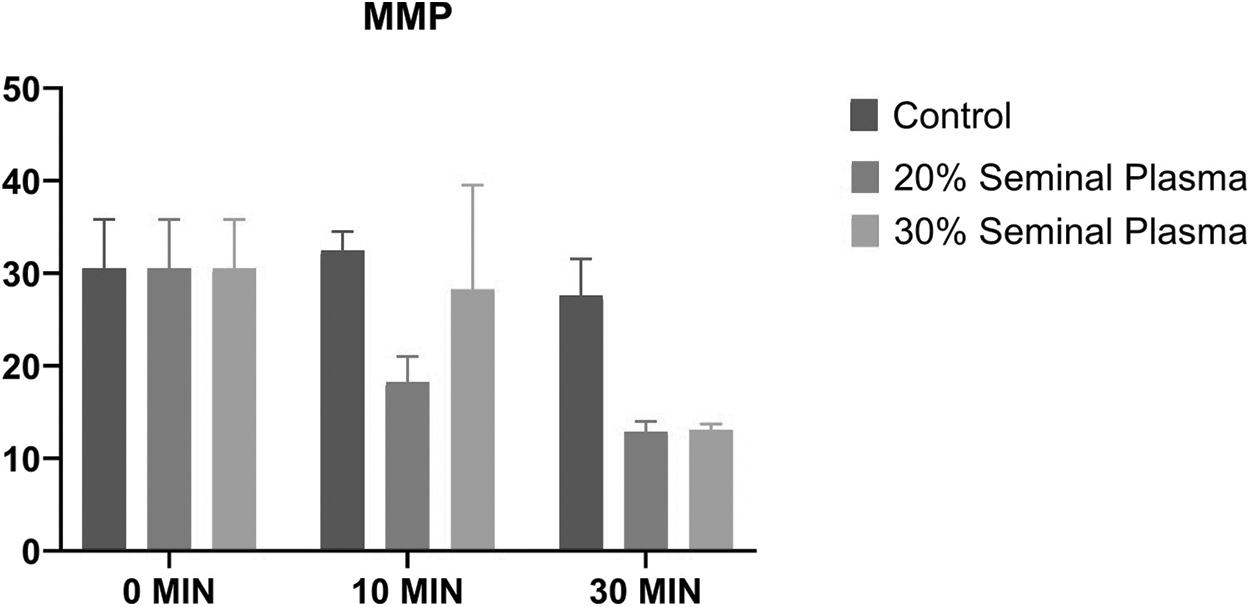
Dynamics of stallion sperm MMP up on supplementation of the SP (Control, 20%, and 30%) at various time intervals (0, 10, and 30 minutes) to the post-thawed stallion semen. MMP, mitochondrial membrane potential.
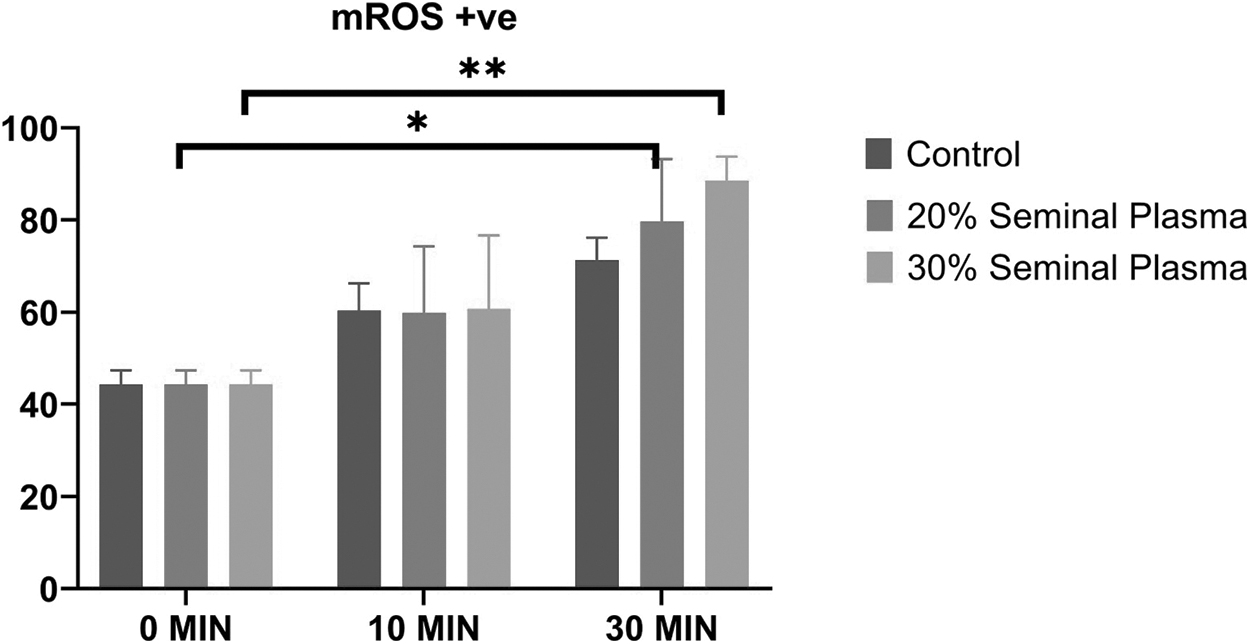
Dynamics of stallion sperm mitochondrial ROS levels up on supplementation of the SP (Control, 20%, and 30%) at various time intervals (0, 10, and 30 minutes) to the post-thawed stallion semen. (*indicates p < 0.05 and **indicates p < 0.01). ROS, reactive oxygen species.
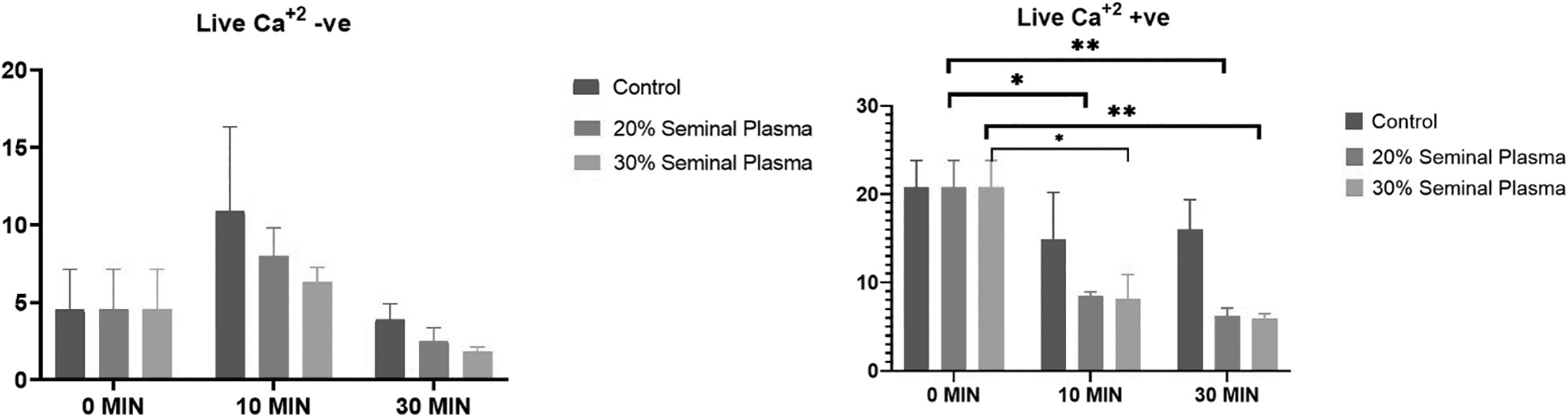
Dynamics of stallion sperm intracellular calcium levels up on supplementation of the SP (Control, 20%, and 30%) at various time intervals (0, 10, and 30 minutes) to the post-thawed stallion semen (*indicates p < 0.05 and **indicates p < 0.01).
Sperm viability analysis revealed that there was significant (p = 0.01) reduction in viability in the 20% SP group after an incubation of 30 minutes (Fig. 1). We also found significant reduction in viability in the 30% SP group after an incubation for 10 minutes (p = 0.04) and 30 minutes (p = 0.01). Moribund sperm population significantly (p = 0.044) decreased in the 30% SP group following an incubation of 30 minutes. There was also decrease in moribund sperm population in the 20% SP group but it was not significant. Following an incubation of 10 minutes, there was significant (p = 0.006) reduction in moribund population between the control and 20% SP groups. In addition, at 30 minutes of incubation, significant reduction in moribund population was observed in the control group versus the 20% SP group (p = 0.05) and in the control group versus the 30% SP group (p = 0.03). The dead sperm population significantly increased after incubation with 20% SP for 30 minutes (p = 0.01) as well as after incubation with 30% SP for 30 minutes (p = 0.01).
Assessment of sperm MMP in the control, 20% SP, and 30% SP groups revealed no significant changes in MMP in between the control group and the two treatment groups at different time intervals (Fig. 2). In addition, sperm MMP within each group was found to be nonsignificant when analyzed at different time intervals. The production of sperm mitochondrial reactive oxygen species (mtROS) was assessed in the control, 20%, and 30% SP-supplemented groups, and it was found that there was significant (p = 0.036) increase in mtROS production in the 20% SP group after 30 minutes of incubation (Fig. 3). Also, there was a significant (p = 0.0084) increase in mtROS production in the 30% SP group after 30 minutes of incubation. We observed no significant difference in mtROS production between the control, 20%, and 30% SP-supplemented groups at different time intervals.
The live sperm with low calcium population within each group was found to be nonsignificant when analyzed at different time intervals (Fig. 4). The live sperm with high calcium in 20% SP was significantly reduced upon incubation for 10 minutes (p = 0.019) as well as for 30 minutes (p = 0.006). Similarly, in the 30% group, there was significant reduction in live sperm with high calcium when assessed at 10 minutes (p = 0.02) and 30 minutes (p = 0.006). The dead sperm with high calcium was increased upon incubation with 20% SP for 10 minutes; however, 30% SP did not alter the dead positive population significantly. The dead sperm with low calcium in 20% SP was significantly increased upon incubation for 10 minutes (p = 0.016) as well as 30 minutes (p = 0.04). Similarly, in the 30% group, there was significant increase in dead sperm with low calcium when assessed at 10 minutes (p = 0.01) and 30 minutes (p = 0.03).
Discussion
In the present study, we reconstituted frozen–thawed spermatozoa (from stallion producing semen with poor cryotolerance) with heterologous SP (from stallions with higher sperm cryotolerance) and evaluated crucial sperm functional parameters in a time-dependent manner. During the period of incubation, the viability of stallion spermatozoa declined in the control group as well as in the 20% and 30% SP groups. However, the sperm viability was significantly reduced in the SP-supplemented groups upon incubation for 30 minutes. The decline in viability in the control group was nonsignificant. This reflects the deleterious effect of SP on sperm viability in a time-dependent manner. This finding corroborates the earlier studies conducted by Šichtař et al., 24 who reported that there was no improvement in viability after addition of SP upon incubation for 30 minutes. However, in contrast, De Andrade et al. 14 reported that the addition of 20% SP increased the proportion of sperm with intact plasma membrane.
We also observed that the moribund sperm population significantly declined in the 20% and 30% SP-supplemented groups following incubation for 30 minutes compared with the control group. In the control group, there was a nonsignificant increase in moribund population following incubation for 10 minutes, indicating that higher population of live spermatozoa were converted to a moribund state. At 30 minutes of incubation, the moribund population was less in the control group, but in the SP-enriched groups, there was sharp decline in moribund sperm populations at 10 minutes as well as 30 minutes.
The decreased moribund sperm population cannot be considered as a positive effect of SP addition because the dead sperm population also increased significantly in the 20% SP-supplemented group after 30 minutes (p = 0.01) as well as in the 30% SP-supplemented group after 30 minutes (p = 0.01). This might be due to the presence of unidentified factors present in SP resulting to rapid shift from viable to moribund and eventually to dead spermatozoa (SP induced transition of viable → moribund → dead spermatozoa). The importance of moribund and dead spermatozoa should be considered as these are a potent source of production of reactive oxygen species that has deleterious effects on functionality of viable spermatozoa population. 25
mtROS assessment revealed that there was significant increase in reactive oxygen species (ROS) production in the 20% and 30% SP groups upon incubation for 30 minutes. The possible reason for the increased level of ROS in both treatment groups may be due to higher proportion of dead spermatozoa that acts as a vessel for ROS generation and release. Thus, the addition of SP in stallion sperm increased the ROS rather than the ameliorate oxidative stress. Al-Essawe et al. 15 reported that the sperm treated with SP from good freezer stallions had the lowest proportion of sperm with high MMP.
The proportion of live sperm with low intracellular calcium did not vary significantly between the control and treatment groups. In addition, within each group, the difference was found to be nonsignificant. However, the proportion of live sperm with high calcium in both the 20% and 30% SP-supplemented groups was significantly reduced at 10 minutes as well as 30 minutes of incubation. However, this difference was nonsignificant in the control group. However, the proportion of dead sperm with low intracellular calcium increased significantly in the 20% and 30% SP-supplemented groups at 10 and 30 minutes of incubation. Collectively, live high-calcium sperm population was decreased, whereas dead low-calcium sperm population was increased upon incubation with SP (shift from live high calcium to dead low calcium). In addition, the dead sperm population also increased.
Sperm with low calcium are destined to die, but the sperm with high calcium are the surviving population. Although this hypothesis was explained and reported in dog and boar spermatozoa earlier, this can be applicable to stallion sperm as well.26,27 Therefore, this study proves that the stallion sperm can survive with high calcium concentrations in their cytosol such as dog and boar sperm. Recently, we reported that calcium binding proteins were upregulated in the SP of stallion producing high-motile sperm. 19 In the present study, we also used SP from high quality semen-producing bulls. Therefore, the calcium binding proteins from the SP that was added in this study could have been responsible for the higher proportion of low-calcium sperm probably by causing calcium efflux.
Altogether, our study was an attempt to assess if supplementation of heterologous SP of stallions with good semen quality to the spermatozoa of stallions producing low quality semen had any beneficial effects on sperm functions. Flow cytometric assessment revealed no improvement in semen quality after SP supplementation in a time-dependent manner. Collectively, SP addition could not repair the damages caused by the cryopreservation and further resulted in deterioration of semen quality as observed in our study by reducing viability, increasing ROS production due to higher proportion of dead cells, or possibly due to presence of some factors (yet to be identified) that are inducive of oxidative stress in stallion spermatozoa. Further research need be conducted to unravel the molecular composition of stallion SP and their putative roles in sperm function and fertilizing potential. Therefore, the molecules that are detrimental to sperm viability and ROS generation can be identified and removed/inhibited, which might help in enriching SP with molecules beneficial for maintaining normal sperm functionality.
Footnotes
Acknowledgments
The authors thank the Director, ICAR-National Dairy Research Institute, India, for providing necessary facilities for carrying out this research. The authors are thankful to the Equine production Campus, ICAR-National Research Centre on Equines, Bikaner, Rajasthan for providing stallion semen.
Data Availability Statement
Data will be made available by the corresponding author upon reasonable request.
Author Disclosure Statement
All authors hereby declare that there is no conflict of interest among the authors and co-authors. All the authors are aware of submission and agreed to be listed as co-authors and sequence of the authorship.
Funding Information
This work was supported by the Indian Council of Agricultural Research under the Indian Council of Agricultural Research-Post-Doctoral Fellowship Programme (ICAR-PDF) granted to Dr Thirumala Rao Talluri under the mentorship of Dr A Kumaresan.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.