
Abstract
Oxidative stress is one of the main causes of loss of sperm function during chilled storage. The aim of the current study was to evaluate the effects of a fructose-based extender, which was supplemented with catalase or uric acid, on the motility, viability, morphological integrity, and lipid peroxidation (LPO) of Colossoma macropomum spermatozoa. Sperm was diluted in extenders containing catalase (0; 0.1; 0.8; and 1.5 kU/L) or uric acid (0; 0.25; 0.5; and 1.0 mmol/L) and then stored at 4.3 ± 0.6°C for 96 hours. The chilling storage time had more significant and pronounced effects on practically all the measured sperm quality parameters than the different concentrations of both antioxidants added to the extenders. This was true for sperm motility, motility duration, sperm viability, and the percentage of normal spermatozoa. In fact, for all these parameters, values were higher in the extenders supplemented with catalase or uric acid, than those not supplemented with these antioxidants, especially after 96 hours. The LPO process showed an antioxidant-dependent response. In catalase-supplemented extenders thiobarbituric acid reactive substance (TBARS) levels increased gradually and significantly with time, but remained stable during the 96 hours of chilled storage in all samples in which uric acid was added. Despite this, TBARS levels were lower in the extenders supplemented with both catalase and uric acid than in those not having these antioxidants. Inverse correlations were found between sperm motility and the damage in sperm flagella. Our findings suggest that the supplementation of an extender with catalase or uric acid is beneficial and protects fish sperm membranes from damage caused by oxidative stress during low-temperature storage. The extenders containing 0.1 kU/L of catalase and 0.25 mmol/L of uric acid provided effective antioxidant protection for the spermatozoa of this important Amazonian fish.
Introduction
Oxidative stress is commonly recognized as one of the main causes that compromise the morphology and functional capabilities of spermatozoa. Excessive reactive oxygen species (ROS) production results in several deleterious effects in these cells, such as membrane and DNA damage, loss of motility, and fertilization capacity in fish and mammalian species.1–4 Spermatozoa are particularly susceptible to attacks by ROS due to the amount of polyunsaturated fatty acids in their composition, leading to lipid peroxidation (LPO) of the cell plasma membrane and, therefore, decreases the viability of the sperm. 1 Under in vivo conditions, sperm has a widespread and complex antioxidant system that acts as a reducing agent of ROS and, thus, maintains its functionality and viability. 5 Antioxidants are considered the main defense against ROS and include different enzymes and a portfolio of nonenzymatic molecules, such as ascorbic acid, tocopherol, vitamins, carnitine, methionine, reduced glutathione, and uric acid, which maintain a physiological balance between the production and elimination of ROS and its metabolic products. 6
Among the technologies used for sperm cryopreservation, chilling is a simple and inexpensive short-term storage technique that aims to maintain the viability of spermatozoa at temperatures close to 4°C for hours or days.7–9 Preservation through chilling facilitates the transport of sperm between fish farms and simplifies the brood stock management during artificial insemination procedures.
In fish, the success of sperm storage depends on a variety of factors, including the age and individual variability among males, the storage (temperature and oxygen availability) and handling (dilution ratio) conditions, and, chiefly, the use of extenders containing substances that increase the life span of the sperm.10,11 Sperm viability of most fish decreases during chilled storage, which is mainly caused by the overproduction of pro-oxidizing agents by the spermatozoa.12,13 In addition, it is well known that the antioxidant system that is naturally present in fish sperm is inefficient in protecting spermatozoa against oxidizing molecule attacks that occur during storage.14,15 To overcome this challenge, several studies have investigated the supplementation of sperm extenders with antioxidants to mitigate the impacts of ROS during chilled storage in different vertebrates such as bovines, 16 stallions, 17 boars, 18 and fish.5,19
Studies involving teleost fish species have confirmed the presence of enzymatic (superoxide dismutase, glutathione peroxidase and glutathione reductase, catalase, and methionine reductase) and nonenzymatic (mainly uric acid, ascorbic acid, glutathione, α-tocopherol, and ubiquinol) antioxidants in both seminal plasma and spermatozoa. 4 Among the enzymatic molecules, catalase is part of the first line of antioxidant defense that catalyzes the reduction of hydrogen peroxide to water and oxygen. 20 Its concentration in fish seminal plasma and spermatozoa is lower and, therefore, plays only a minor role in in vivo conditions.5,19 Uric acid is one of the main molecules that make up the second line of antioxidant defense and acts to capture active radicals to inhibit the onset of chain reactions. 20 Its concentration in fish sperm, mainly in seminal plasma, is higher and is stable compared to other nonenzymatic antioxidants.5,19,21 Under in vitro conditions, supplementation with catalase and uric acid in fish sperm extenders has proved to be efficient in maintaining or at least mitigating the reduction of sperm motility and sperm integrity during chilled storage.5,19
Despite previous studies conducted with mammalian species showing that high levels of ROS are associated with an increased incidence of sperm morphological abnormalities,15,16,22 this approach has never been used in any fish species, whether marine or freshwater. In the Amazon basin, there is particular interest in the teleost fish Colossoma macropomum, also known as tambaqui. This native species has an important and historical relationship with cultural and economic aspects in both artisanal fisheries and aquaculture, and has a high rate of consumption in Brazil and other South American countries. 23 In contrast, there is concern about the current status of natural stocks of C. macropomum, since its populations have been suffering from overfishing for decades, and is has become increasingly difficult to find large-size fish (≥7 kg), even at distances of over 1000 km from the city of Manaus. 24
In addition, there are still no defined protocols regarding cryopreservation of sperm for this species. In one study, it was revealed that the storage of semen without extenders failed to maintain the viability of spermatozoa, which resulted in dead cells. However, the inclusion of these cells in a fructose-based extender was able to maintain sperm motility for at least 96 hours during storage at the same temperature. 25 This investigation also revealed that the fructose-based extender was more efficient in protecting sperm cells against the increase in coiled flagella damage in this species, compared to glucose-based extenders. 25
Thus, the present study aimed to evaluate the effects of supplementing the extenders with catalase and uric acid on the motility, viability, morphological integrity, and the levels of LPO in C. macropomum spermatozoa stored under chilled conditions.
Materials and Methods
Ethical statement
The experimental procedures conducted in the present study were in accordance with the ethical principles adopted by the National Council for the Control of Animal Experimentation (CONCEA) and were approved by the Ethics Committee for Animal Use in Experiments of Nilton Lins University (CEUA protocol No. 007/2016).
Brood stock and hormonal treatment
Mature male C. macropomum was obtained from the Experimental Aquaculture Station of the Federal University of Amazonas (UFAM), located 40 km from Manaus, Amazonas, Brazil (02°38’56.1” S; 60°03’14.7” W). The brood stock was kept in 600 m2 earthen ponds (1 fish/12 m2) and fed thrice a week with extruded commercial feed (40% crude protein, Confiança®, Amazonas, Brazil), at 2% of the total biomass. The fish were sampled during the breeding season (September–December) of 2019, and those that released sperm after a light abdominal massage in the cranial-caudal direction were selected for the experimental trials and then acclimated in two indoor circular tanks (2 m3) with continuous water supply from the same source as the brood stock, for 6 hours, until sperm collection. During this period, water quality was monitored, and the following results were obtained: dissolved oxygen = 6.0 ± 0.6 mg/L, pH = 6.8 ± 0.3, and temperature = 27 ± 1°C.
Hormonal induction and sperm collection
After the acclimation period in the indoor circular tanks, C. macropomum males were intraperitoneally injected with a single dose (2.0 mg/kg) of crude carp pituitary extract (Danubio, Santa Catarina, Brazil) to increase the sperm volume to the level necessary for the analyses. After 11 hours of hormonal application, fish were anesthetized with eugenol (76 mg/L) 26 for 2.42 ± 0.42 minutes. The urogenital region of each male was dried with a paper towel, and contamination of sperm with blood, feces, urine, or mucus was carefully avoided during stripping.
A mean volume of 2.5 ± 1.0 mL of sperm was collected from each of the fish and immediately assessed for sperm motility using light microscopy with 10 × magnification (Nikon Eclipse E100-LED, Melville, NY, USA). Sperm samples with motility of ≥90% after activation (2 μL of sperm were diluted with 198 μL of distilled water—1:100) were selected to compose a pool (10 mL), which was designed to prevent the effect of individual variability of sperm quality and concentrations 27 that is observed in C. macropomum. 28 Thereafter, fresh pooled sperm samples were distributed among the treatments, evaluated for sperm motility and motility duration, and transported to the laboratory at a low temperature (5.1 ± 0.2°C). To assess sperm concentration, a 2 μL of pooled fresh sperm was fixed in 1998 μL of buffered formal-saline solution (1:1000), and an aliquot (12 μL) of this diluted solution was placed in a hemocytometer (Neubauer Improved) to perform the sperm counts (in triplicate).
Extenders
A fructose-based extender (previously developed by our research group) was used as a standard extender, 25 and was supplemented with different concentrations of catalase (0; 0.1; 0.8; and 1.5 kU/L) or uric acid (0; 0.25; 0.5; and 1.0 mmol/L) as antioxidants. The concentrations of catalase and uric acid (Sigma Aldrich, San Luis, MO, USA) were defined according to the results previously found for fish spermatozoa stored at 4°C.5,19 The pH of the extenders was adjusted with a 2 mM sodium hydroxide solution to 7.6 ± 0.01 and osmolality to 300 ± 0.01 mOsmol/kg, according to Pastrana. 29
Experimental design
In the present study, two independent experiments were conducted to assess the protective effects of catalase or uric acid added to a standard (fructose-based) extender.
In Experiment 1, the collected sperm from seven males (3.1 ± 1.3 kg) was pooled and diluted into the extender with the different concentrations of catalase in the proportion of 1:10 (100 μL sperm: 900 μL extender) in 1.5 mL microcentrifuge tubes, with a final cell density of 2.4 ± 1.3 × 109 spermatozoa/mL. For Experiment 2, the pooled sperm collected from ten different males (2.5 ± 0.2 kg) was diluted into 1.5 mL microcentrifuge tubes containing the same standard extender with different concentrations of uric acid in the proportion of 1:10 (100 μL sperm: 900 μL extender), but with a final cell density of 5.8 ± 1.7 × 108 spermatozoa/mL.
In both experiments, the diluted samples from all the treatments (catalase or uric acid) were allocated in microtubes arranged in a 4 × 3 factorial scheme, whereby the first factor corresponded to the concentrations of antioxidants and the second factor to the three chilled storage times (0, 48, and 96 hours). The experimental unit was characterized by a closed microtube containing 1000 μL of diluted sperm, with seven replicates per treatment. Sperm samples were kept in a refrigerator at 4.3 ± 0.6°C, and the sperm motility (%), the motility duration (s), sperm cell viability (%), sperm morphology (%), and thiobarbituric acid reactive substance (TBARS) levels (nmol/sperm cells) were assessed at time 0 hour (immediately after dilution) and after 48 and 96 hours of storage.
Sperm motility assessment
To assess sperm motility in chilled samples, an aliquot (2 μL) was placed on a histological slide and diluted (1:10) with distilled water (28.6 ± 0.5°C). The motility was subjectively assessed by an experienced researcher 30 under a preset optical (Nikon Eclipse E100-LED, Melville, NY, USA) microscope (10 × magnification), and the percentage of motile cells exhibiting forward movement was recorded. 14 To assess duration of the motility (in seconds), a timer was triggered at the moment of sperm cell activation and counting stopped at the full arrest of motility for all spermatozoa in the optical field. These analyses were conducted in triplicate.
Sperm cell viability test
Plasma membrane viability of the spermatozoa was assessed in accordance with the method adapted from Garner et al. 31 First, chilled sperm was diluted in the proportion of 1:40 (5 μL of sperm: 195 μL of potassium phosphate buffer) to reduce sperm concentration and facilitate sperm count. Thereafter, 50 μL of diluted sperm was added to an Eppendorf microtube and mixed with 1 μL of an 8.7 mmol/L 5-carboxyfluorescein diacetate solution in 0.4% dimethyl sulfoxide (diluted in 50 mmol/L buffer, pH 7.0). After 4 minutes at room temperature, 3 μL of a 10.4 mmol/L propidium iodide solution in phosphate buffer was added and mixed into the microtube.
After 1 minute, 5 μL of stained sperm was placed on a histological slide, covered with a coverslip, and assessed under a fluorescence microscope (Leica, DM 2000, Wetzlar, Germany) at 40 × magnification. Blue and green light filters were used alternately to identify cells with intact membranes (spermatozoa that fluoresced green) and cells with damaged membranes (spermatozoa that fluoresced red), respectively.
From each experimental unit, two histological slides were prepared, and 100–400 spermatozoa were counted per slide. The number of viable spermatozoa was expressed as a percentage of all observed cells.
Spermatozoa morphology
Fresh sperm was fixed in a buffered formal saline solution in the proportion of 1:1000 (2 μL of sperm: 1998 μL of solution), whereas the diluted sperm was fixed in a proportion of 1:100 (10 μL of sperm: 990 μL of solution). The buffered fixative solution was prepared by dissolving 1.8% NaCl (150 mL), 4.3% Na2HPO4 (71.4 mL), 4.5% KH2PO4 (28.6 mL), and 37% commercial solution of formaldehyde (62.5 mL in 500 mL distilled water). 32 Afterward, 50 μL of this dilution was placed on a histological slide and immediately stained with a 5 μL aliquot of Bengal Rose dye. 33
One hundred spermatozoa were assessed on each slide using optical microscopy (100 × magnification, Nikon Eclipse E100-LED, Melville, NY, USA), with three replicates per experimental unit. Spermatozoa showing integrity of the head, mid-piece, and flagellum structures, as well as a standard shape, were considered as being cells with normal morphology. 25 The abnormal or damaged cells were classified according to the presence of a detached head, degenerated head, degenerated mid-piece, detached flagellum, fractured flagellum, simple bent flagellum, and coiled and strongly coiled flagellum. The number of spermatozoa with normal or abnormal morphology was expressed as a percentage of all observed cells.
Sperm LPO test
The LPO levels, expressed as the concentration of TBARS, were measured in fresh and diluted sperm, in accordance with the method adapted from Draper and Hadley. 34 To prepare the homogenate, fresh sperm was centrifuged (refrigerated centrifuge 5030R, Eppendorf, Hamburg, Germany) at 106 × g for 10 minutes at 4°C to remove seminal plasma. The obtained pellet (spermatozoa) was resuspended in 50 mmol/L potassium phosphate buffer, pH 7.0, containing 0.5 mmol/L ethylenediaminetetraacetic acid, and washed twice in the same buffer.35,36 The chilled sperm was centrifuged, and the extender compounds were discarded. The pellet was rediluted in phosphate buffer and washed twice in the same buffer. An aliquot (300 μL) of homogenate was added to a test tube containing 350 μL of 10% sodium dodecyl sulfate, 350 μL of 10% trichloroacetic acid (TCA), and 1.0 mL of 0.67% thiobarbituric acid. The reaction medium was incubated at 100°C for 30 minutes and kept at room temperature for 15 minutes. Thereafter, the absorbances of the samples were detected using a spectrophotometer (Biospectro, SP-22, São Paulo, Brazil) at a wavelength of 532 nm. 1,1,3,3-tetraethoxypropane (Chem-Impex International, IL, USA) was used in the preparation of a standard curve of malondialdehyde, with a concentration range of 0.5–50 nmol/L. All analyses were conducted in triplicate.
Statistical analyses
Normality and the homogeneity of variance of all data were first assessed using the Shapiro–Wilk and Bartlett tests, respectively. Data from Experiments 1 (motility duration) and 2 (sperm motility, normal spermatozoa, coiled and strongly coiled flagellum, and TBARS) were transformed by square root to reach the parametric assumptions. We used two-way analysis of variance followed by Tukey's test to identify the statistical differences between the treatments (catalase or uric acid concentration) and period of storage (0, 48, and 96 hours). In addition, a simple linear regression analysis was applied to test the relationship between sperm motility (dependent variable) with coiled flagellum and strongly coiled flagellum (independent variables). In all the analyses, a significance level of α ≤ 0.05 was considered. All data are presented as mean ± standard deviation.
Results
The chilling storage time had more significant and pronounced effects on practically all the measured parameters related to sperm quality of C. macropomum than the different concentrations of both catalase and uric acid added to the fructose-based extenders, as follows.
Sperm motility
The motility of C. macropomum sperm decreased gradually (p < 0.05) in all samples during the chilled storage, irrespective of the presence or absence of catalase in the extenders. This pattern was more evident after 96 hours when the motile spermatozoa represented only 30% of those found in the initial evaluation (Table 1). No significant differences in the sperm motility were observed among samples supplemented with catalase during each chilled storage period. However, a higher percentage of motile spermatozoa (p < 0.05) was observed in samples supplemented with this enzyme (73 ± 0.8% and 31 ± 2% at 48 and 96 hours, respectively) compared to those found in catalase-free samples (62 ± 4% and 16 ± 5% at 48 and 96 hours, respectively; Table 1).
Quality Parameters (Mean ± Standard Deviation) of Colossoma macropomum Sperm Throughout Chilling Storage at 4°C in a Catalase-Supplemented Extender
Data were analyzed using two-way ANOVA followed by Tukey's test. Distinctive capital letters in the same time and lowercase letters among chilling times indicate significant statistical difference (p < 0.05).
ANOVA, analysis of variance.
No differences were found in the duration of motility of spermatozoa incubated with the different concentrations of catalase added to the fructose-based extenders, as well as compared to samples without the presence of this enzyme. However, a significant decrease (p < 0.05) in this parameter was found in all the samples (with and without catalase) only after 96 hours of chilled storage (Table 1).
Regarding the use of uric acid, a similar pattern was evidenced in the motility of sperm in C. macropomum, that is, a gradual and significant (p < 0.05) decrease was observed according to the chilled storage time, even with the presence or not of this antioxidant (Table 2). No significant differences in the sperm motility were observed in any of the samples (with and without uric acid) during each chilled storage time. However, a higher percentage of motile spermatozoa (p < 0.05) was observed in samples supplemented with uric acid (73 ± 2% and 18 ± 6% at 48 and 96 hours, respectively), compared to samples without this antioxidant (57 ± 4% and 8 ± 2% at 48 and 96 hours, respectively; Table 2).
Quality Parameters (Mean ± Standard Deviation) of Colossoma macropomum Sperm Throughout Chilling Storage at 4°C in a Uric Acid-Supplemented Extender
Data were analyzed using two-way ANOVA followed by Tukey's test. Distinctive capital letters at the same time and lowercase letters among chilling times indicate significant statistical difference (p < 0.05).
The motility duration of spermatozoa was not influenced (p > 0.05) by the absence or presence of different uric acid concentrations in the extenders. However, the time of storage promoted a significant decrease (p < 0.05) in the motility duration after 48 and 96 hours of chilling, with a reduction of 15 and 29 seconds, respectively, in relation to the initial evaluation (Table 2).
Sperm viability
Sperm cell viability was influenced by an interaction between the catalase concentration of the extenders and storage time (p < 0.05). A significant decrease of 32% in viable sperm cells was observed in all treatments after 48 hours of storage compared to the initial evaluation (0 hour). In contrast, after 96 hours of chilling, the sperm cell viability (63 ± 6%) in the presence of catalase was higher (p < 0.05) than those without (49 ± 6%) this antioxidant enzyme (Table 1).
In extenders supplemented with uric acid, the sperm cell viability was also influenced by an interaction between the concentration of this antioxidant and the storage time (p < 0.05). After 96 hours, sperm viability decreased 23% and 40% in the extenders with and without uric acid supplementation, respectively, compared to the initial evaluation. In addition, at 96 hours sperm cell viability was greater (p < 0.05) in samples containing uric acid (53 ± 7%), compared to those when this antioxidant was not added to the extender (36 ± 5%; Table 2).
Sperm morphology
The different concentrations of catalase added to the extenders did not affect (p > 0.05) the normal morphology of spermatozoa in up to 48 hours of chilled storage, even compared to the catalase-free samples (Fig. 1a). Although an interaction effect (p < 0.05) between the catalase concentration of the extender and the storage period was observed, significant decreases (p < 0.05) in the proportion of normal spermatozoa were only observed after 96 hours of chilled storage, with the presence of cells with normal morphology (81 ± 1%) in the catalase-supplemented extenders being significantly higher than in the extenders without (70 ± 2%) this enzyme (Table 1).

Light micrographs of Colossoma macropomum spermatozoon.
In all treatments, morphological abnormalities were observed in the head (detached = 2.0 ± 1.0% and degenerated = 1.0 ± 0.4%), mid piece (degenerated = 4.0 ± 1.0%), and flagellum (simple bent = 4.0 ± 0.4%, coiled = 3.0 ± 1.0%, strongly coiled = 2.0 ± 1.0%, fractured = 1.4 ± 0.2%, and detached = 1.0 ± 0.3%). The chilling storage period increased (p < 0.05) the occurrence of cells with coiled and strongly coiled flagellum only after 96 hours (Fig. 1b, c). The incidence of spermatozoa with coiled flagellum was greater (p < 0.05) in the extenders not supplemented with catalase compared to those supplemented with catalase (Fig. 2a). In addition, the percentage of spermatozoa displaying strongly coiled flagellum doubled (p < 0.05) in the catalase-free extender, compared to those supplemented with this antioxidant enzyme (Fig. 2b).
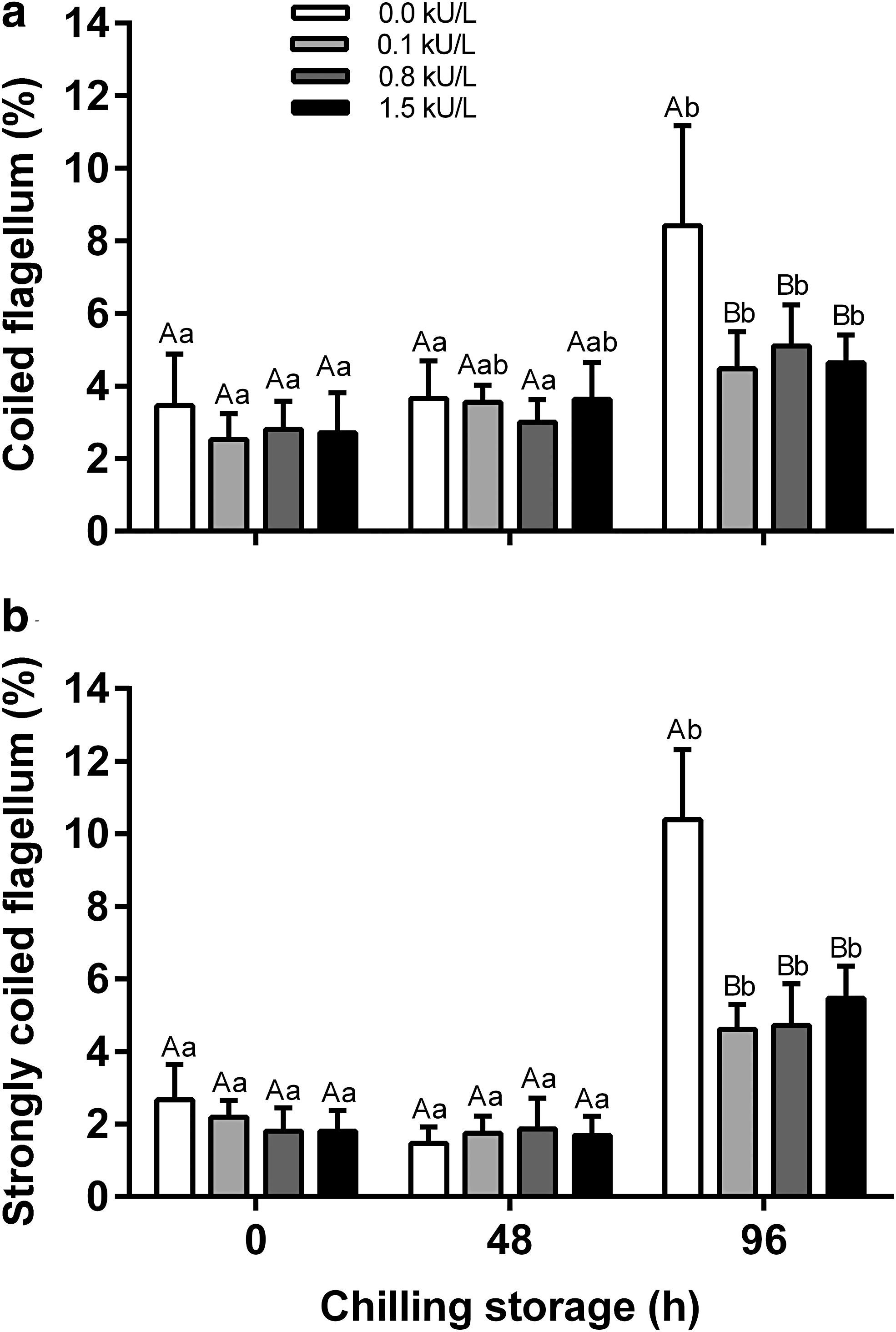
Percentage of Colossoma macropomum spermatozoa displaying coiled flagellum and strongly coiled flagellum after 96 hours of chilled storage at 4°C in a catalase-supplemented extender. Data were analyzed using two-way ANOVA:
Concerning the use of uric acid in the fructose-based extenders, a similar response to the addition of catalase was observed, that is, the different concentrations of uric acid did not affect (p > 0.05) the normal morphology of spermatozoa in up to 48 hours of chilled storage, even compared to samples lacking this antioxidant (Table 2). In contrast, significant decreases (p < 0.05) in the percentage of normal spermatozoa were observed only after 96 hours of chilled storage, in which the presence of cells with normal morphology was 12% higher (p < 0.05) in uric acid-supplemented extenders than in the samples kept in uric acid-free extenders (Table 2).
In all extenders, morphological abnormalities were observed in the head (detached = 1.2 ± 0.4% and degenerated = 1.0 ± 0.3%), mid piece (degenerated = 4.0 ± 0.5%), and flagellum (simple bent = 4.0 ± 0.6%, coiled = 3.0 ± 0.5%, strongly coiled = 1.4 ± 0.4%, fractured = 1.2 ± 0.3%, and free = 1.2 ± 0.3%). Similarly, for the samples supplemented with catalase, the chilled storage time increased (p < 0.05) only the incidence of cells with coiled and strongly coiled flagellum (Fig. 1b, c). These increments were evident at 96 hours of chilled storage, in which the percentage of spermatozoa with coiled flagellum was 25% and 63% higher in samples kept in extenders with and without uric acid, respectively, compared to 48 hours of chilled storage (Fig. 3a). The percentage of spermatozoa showing strongly coiled flagellum was 2-fold higher (p < 0.05) after 96 hours in sperm stored in the uric acid-free extender (10 ± 0.7%) compared to samples kept in extenders supplemented with uric acid (5 ± 1%; Fig. 3b).
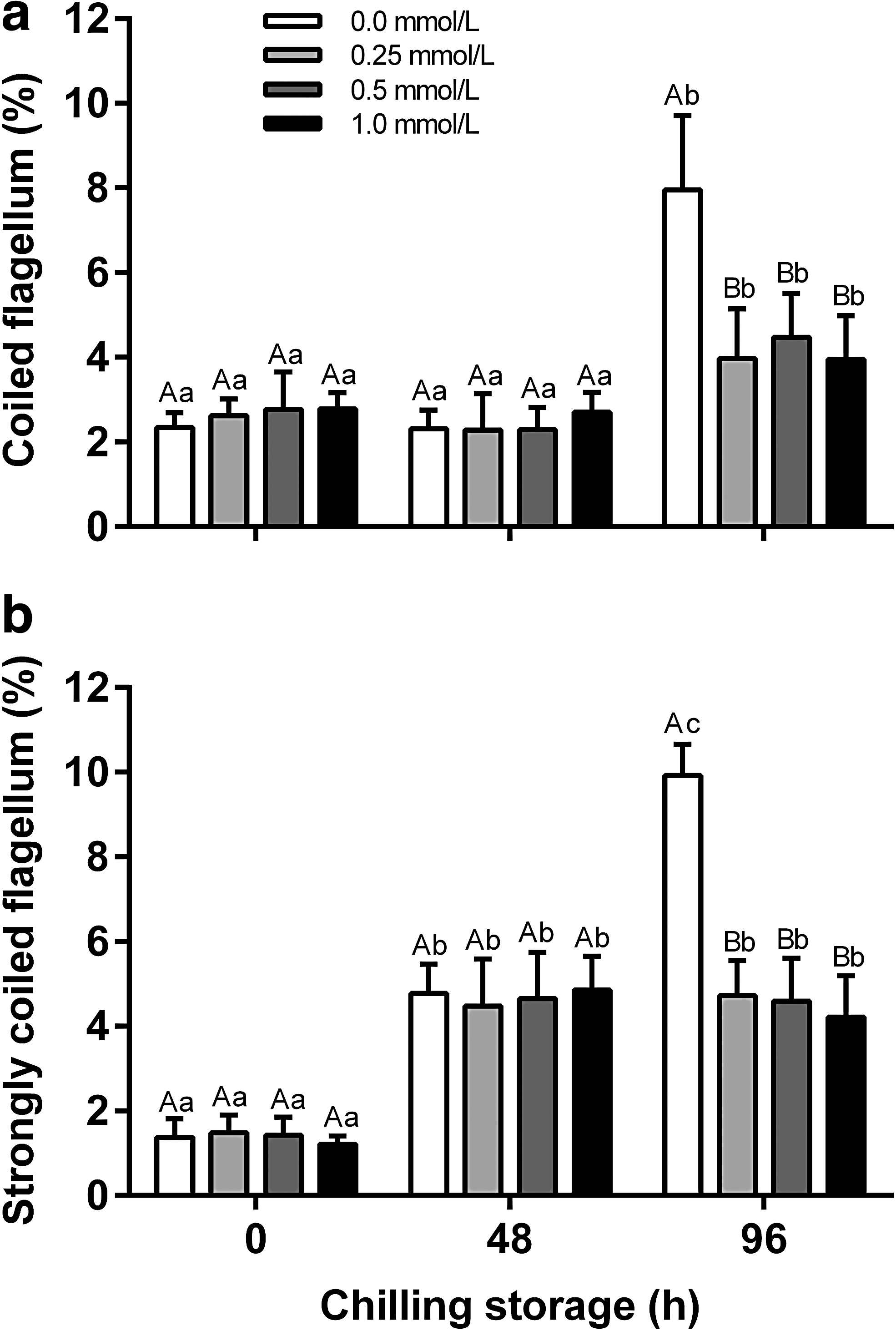
Percentage of Colossoma macropomum spermatozoa displaying coiled flagellum and strongly coiled flagellum after 96 hours of chilled storage at 4°C in a uric acid-supplemented extender. Data were analyzed using two-way ANOVA:
Lipid peroxidation
The TBARS concentration in spermatozoa was not influenced (p > 0.05) by the different concentrations of catalase supplementing the extenders during each chilled storage time. In contrast, although a gradual and significant increase was observed with time, lower TBARS values (p < 0.05) were found in spermatozoa maintained in the presence of catalase after 48 and 96 hours of chilled storage (0.65 ± 0.06 nmol/108 cells and 1.09 ± 0.08 nmol/108 cells, respectively) compared to the catalase-free samples (1.03 ± 0.08 nmol/108 cells and 2.25 ± 0.24 nmol/108 cells, respectively) assayed for the same periods (Table 3).
Concentration (Mean ± Standard Deviation) of Thiobarbituric Acid Reactive Substances in Colossoma macropomum Sperm After 96 hours Chilling Storage at 4°C in a Fructose-Based Extender Supplemented with Different Concentrations of Catalase or Uric Acid
Data were analyzed using two-way ANOVA, and all pairwise were compared by post hoc Tukey's test. Distinctive capital letters at the same time and lowercase letters among chilling times indicate significant statistical differences (p < 0.05).
TBARS, thiobarbituric acid reactive substance.
The concentration of uric acid in the extender and the storage period also affected the TBARS levels produced in the spermatozoa. The levels of this biomarker increased (p < 0.05) three and four times in the extender without this antioxidant after 48 and 96 hours of chilled storage, respectively, compared to the initial evaluation. In contrast, the TBARS levels remained stable (p > 0.05) in all samples kept in the uric acid-supplemented extenders during the 96 hours of chilled storage (Table 3).
Correlation between sperm motility and flagellum damage
In both experiments and in all the extenders (with and without antioxidants), a negative linear relationship between sperm motility and the number of spermatozoa with flagellum damage was observed. In those samples kept in an antioxidant-free extender, this relationship was observed in spermatozoa with coiled flagellum (R 2 = 0.74; p < 0.001) and strongly coiled flagellum (R 2 = 0.86; p < 0.001; Fig. 4).
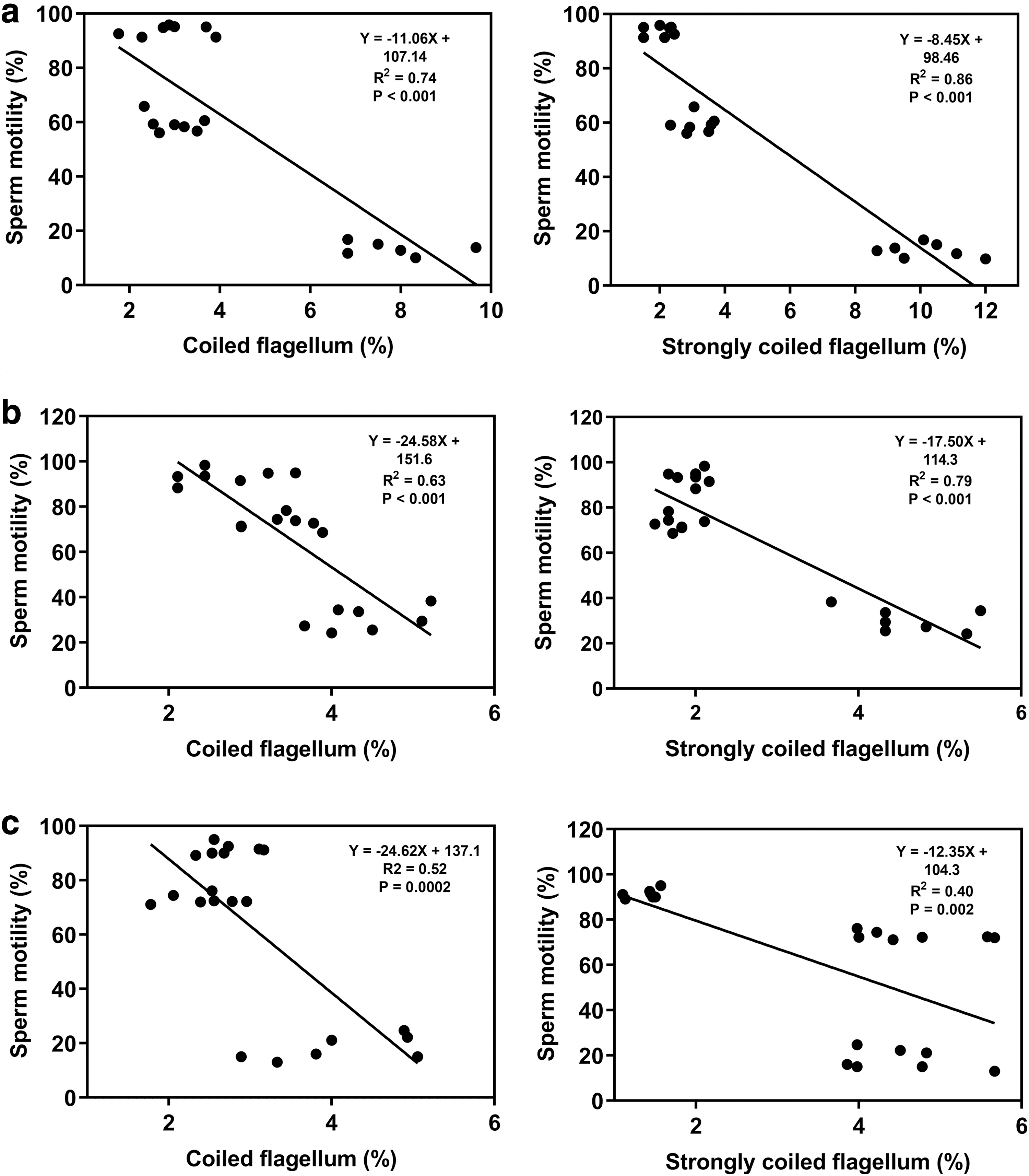
Simple linear regressions between sperm motility and flagellum damage in Colossoma macropomum spermatozoa after 96 hours of storage at 4°C.
In catalase-supplemented extenders, this negative linear relationship was stronger in spermatozoa with strongly coiled flagellum (R 2 = 0.79; p < 0.001) than in those cells with coiled flagellum (R 2 = 0.63; p < 0.001). However, although also significative, this negative linear relationship between sperm motility and the number of spermatozoa with coiled flagellum (R 2 = 0.52; p = 0.0002) and strongly coiled flagellum (R 2 = 0.40; p = 0.002) was weaker in samples maintained in uric acid-supplemented extenders compared to those without any antioxidant or supplemented with catalase (Fig. 4).
Discussion
In our study, we showed that supplementation of a fructose-based extender with enzymatic (catalase) or nonenzymatic (uric acid) antioxidant can protect the sperm cells of C. macropomum against damage caused during chilled storage. In addition, we demonstrated that the decreased sperm motility observed during chilled storage is largely explained by increases in flagellum damage caused by ROS attacks.
The standard (fructose based) extender used in our study has been refined by our research group for use as a specific extender for C. macropomum spermatozoa. This extender contains energy sources for spermatozoa metabolism, controlled pH and osmolality, and the addition of antibiotics to prevent bacterial growth. 25 However, studies have demonstrated that the storage of fish spermatozoa under refrigeration leads to excessive production of oxidizing agents, which are highly reactive and cause negative impacts on the sperm viability. 13 Antioxidants naturally present in fish sperm are not enough to protect the sperm during in vitro storage; thus, supplementation with exogenous antioxidants in extenders is recommended to maintain the viability and quality of spermatozoa.5,19
Colossoma macropomum seminal plasma contains about 0.0027 kU/L of catalase and 115 μmol/L of uric acid 29 ; therefore, the concentrations we added in the extender were 37–555- and 2–8.7-times higher than those naturally found in the seminal plasma of the species, respectively. Such antioxidant supplementation helped by controlling the production of oxidizing agents and maintained spermatozoa quality during chilled storage, which indicates that the spermatozoa antioxidant systems were insufficient for protecting against ROS attacks under such conditions.
Sperm motility is one of the most important variables utilized to assess sperm quality and has been associated with the success of fertilization. 37 In our study, the supplementation of catalase (0.1–1.5 kU/L) or uric acid (0.25–1.0 mmol/L) in the standard extender helped to maintain the motility of the sperm after the chilled storage. In a similar investigation, Lahnsteiner and Mansour 4 demonstrated that a catalase-supplemented extender was beneficial in maintaining the sperm motility of S. trutta, Perca fluviatilis, and Lota lota during refrigeration for 72 hours. In relation to uric acid, the concentration of 0.25 and 0.5 mmol/L had a positive effect on the sperm motility of S. trutta under chilled storage. 19 The catalase and uric acid-supplemented extenders were efficient in minimizing the toxicity of ROS generated by sperm cells during refrigeration and protected spermatozoa against oxidative stress and its resulting damage.
Motility duration of spermatozoa is also an important variable that is measured to determine sperm quality, since this period should be sufficient for sperm to reach the oocyte micropyle before it closes. 38 The excessive production of ROS leads to a LPO of cells, destroying sperm plasma membrane and compromising cellular function. 1 In fact, by adding catalase or uric acid to the extender, we observed a higher proportion of spermatozoa with an intact and viable membrane at the end of chilled storage. Lahnsteiner et al. 19 also observed an increase in sperm membrane integrity in spermatozoa of S. trutta in the presence of catalase after 72 hours of incubation. In a similar study, the sperm membrane integrity of P. fluviatilis, A. alburnus, and L. lota was also improved in the presence of these two antioxidants. 5 This higher percentage of spermatozoa with a viable membrane in our study is a result of the supplemental antioxidants helping to minimize the plasma membrane damage caused by ROS attacks and more efficiently protecting the cell structure.
Sperm morphology should be considered as an indicator of fish sperm quality, since spermatozoa abnormalities are associated with a decrease in motility and fertilization capacity. 38 However, most of the studies dealing with in vitro storage of fish sperm have neglected this variable. Previous studies with mammals have shown that an increase in the incidence of morphological abnormalities of sperm was associated to the high levels of ROS produced in these cells.15,22 Likewise, we found a significant increase in the percentage of spermatozoa with flagellum damage in sperm kept in the antioxidant-free extender. This result is in accordance with other investigations in which the supplementation of extenders with antioxidants decreases the incidence of morphological abnormalities of spermatozoa in stallions, 39 dogs, 15 bovines, 16 and rams 40 after chilled storage. Nonetheless, in fish, the mechanism leading to the increase in abnormalities in the flagellum of spermatozoa during chilled storage is not yet fully understood.
In a previous study evaluating in detail the spermatozoa morphology as an indication of sperm viability in C. macropomum, we demonstrated that the fructose-based extender was more efficient in protecting sperm cells against an increase in coiled flagellum damage compared to glucose-based extenders. 25 In this study, progress has been made to improve the positive effects of the fructose-based extender by supplementing it with catalase or uric acid. Thus, the efficient protection of the sperm membrane from ROS attacks during low-temperature storage led to greater structural stability of cells and, consequently, less flagellum alterations.
In our study, the increased TBARS levels observed in sperm kept in the extender without antioxidants contrasted with the samples supplemented with catalase or uric acid, which is in accordance with the findings of Lahnsteiner et al., 19 who observed augmented TBARS levels in spermatozoa of S. trutta kept in extenders without antioxidants. This indicates that this antioxidant mitigated oxidative stress during chilled storage for 72 hours. Lipid peroxides have a toxic effect on cells, which alters the composition, structure, and dynamics of the plasma membrane, causes cell degradation, and triggers loss of sperm functionality.1,41 In fact, in the case of C. macropomum sperm, our results show that there is no need for supplementation with high concentrations of catalase or uric acid, since a slightly higher concentration than that found naturally occurring in the seminal plasma was enough to provide full protection to the cells, with no difference among the tested concentrations.
The relationship between morphological damage and sperm motility, which are considered indicators of sperm fertilizing ability, has been well documented in mammals, but not in fish. Studies on humans, bovines, equines, and canines show that increased spermatozoa abnormalities are associated with decreases in sperm motility.15,16,39,42,43 Among the morphological abnormalities, flagellum damage has been more frequently correlated with motility decline than damage observed in the head and mid piece.42,44 It has also been demonstrated that the morphological abnormalities in stallion spermatozoa, such as a detached head, mid piece damage, and premature germ cell, were correlated with a decline in sperm motility, but coiled flagellum was the specific damage that was negatively associated with fertilization ability. 45
Our study demonstrated, for the first time in fish, that decreases in sperm motility during chilled storage are largely explained by increases in the number of spermatozoa with coiled and strongly coiled flagellum. The plasma membrane of sperm is a primary structure that is attacked by ROS during chilled storage and leads to morphological abnormalities, especially axonemal damages. 46 Structural axoneme dysfunctions have an adverse effect on sperm motility, since the flagellum can no longer perform the sliding mechanism needed for propagation of flagellar waves and consequent cell movement. 44 The flagellum damage that we observed herein probably occurred because the axonemal structure was disrupted by the overproduction of ROS, leading to a decline of sperm motility during chilled storage. Supplementation of the extender with catalase or uric acid is therefore beneficial for stabilizing the sperm viability and can be used to improve semen storage techniques.
Finally, further investigations in other tropical fishes are needed to understand the mechanisms involving the use of antioxidants in the protection of sperm viability, either for production in artificial (aquaculture) systems or for conservation plans addressed to key freshwater species such as C. macropomum in the Amazon region.
Conclusion
The association between oxidative stress and the incidence of morphological damage to spermatozoa flagellum is described here for C. macropomum spermatozoa and shows that in vitro chilled storage conditions promote morphological damages, especially in the flagellum, with a decrease in sperm motility that is related to an increasing number of spermatozoa with coiled and strongly coiled flagellum. In addition, our findings indicate the supplementation of extenders with catalase (0.1 kU/L) or uric acid (0.25 mmol/L) as a reliable procedure to protect fish sperm cell membranes from damage caused by ROS attacks during low temperature storage. Thus, we believe that supplementation of antioxidants in the extender medium can aid the establishment of a successful protocol for C. macropomum sperm cryopreservation, with applications in both brood stock management and conservation plans for the endangered wild populations.
Footnotes
Acknowledgments
The authors thank the technicians from Experimental Farm (FAEXP) of the Federal University of Amazonas (UFAM) for the assistance provided during experiments. The authors also thank the Laboratory of Ecophysiology and Molecular Evolution (LEEM) of the National Institute of Amazonian Research (INPA) for allowing the use of microscopy facilities.
Authors' Contributions
Y.M.P.: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing—original draft, Writing—review and editing. J.L.M.: Conceptualization, Investigation, Methodology, Writing—review and editing. A.P.d.A.: Conceptualization, Investigation, Methodology, Writing—review and editing. F.B.P.S.: Methodology and Writing—review. E.S.L.: Conceptualization, Investigation, Resources, Writing—review and editing. L.D.R.A.: Methodology and Writing—review. R.O.S.d.S.: Methodology and Writing—review. C.C.G.: Data curation, Methodology, Writing—review and editing. D.P.S.J.: Resources and Writing—review. L.G.: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing—original draft, Writing—review and editing.
Data Availability
The data generated and analyzed during this study are available from the corresponding author upon request.
Ethical Approval
The authors declare that all applicable international, national, and/or institutional guidelines for sampling, care, and experimental use of animals for the study have been followed, and all necessary approvals by the Ethics Committee for Animal Use in Experiments of Nilton Lins University (Protocol CEUA No. 007/2016) have been obtained.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by the Brazilian National Council for Scientific and Technological Development—CNPq (Project No. 421121/2018-1). Y.M.P. was granted a PhD scholarship from the Research Support Foundation of Amazonas State (FAPEAM). L.G. and E.S.L are research fellows from CNPq (Process No. 310463/2018-1 and 405992/2013-0).