
Abstract
Introduction:
Substance P (SP) is a member of the tachykinin family. In the central nervous system, SP participates among others, in the regulation of pain, learning, memory, emotion, and sexual functions. In the periphery, SP affects the gastrointestinal, cardiovascular, and urinary systems. Galanin, similarly to SP, appears to be involved in wide range of physiologic functions, including cognition, waking and sleep, feeding, mood, blood pressure, reproduction, and development, where acts as a trophic factor. The similar distribution of SP-immunoreactive (SP-IR) fibers and galanin-IR perikarya in the human hypothalamus suggests functional interaction between these neuropeptides.
Methods:
We have utilized double-label immunohistochemistry to reveal these putative juxtapositions.
Results:
The majority of galanin-IR neurons receive contacting SP-IR fibers that often cover a significant area of the galaninergic perikarya forming multiple en passant type contacts. These SP-galanin juxtapositions are located mainly in the basal part of the infundibulum/median eminence, populating the basal periventricular region as well as the basal perifornical area.
Discussion:
The density and the morphology of these associations suggest that these contacts are functional synapses and therefore may represent the morphological substrate of the control of SP on multiple functions regulated/modulated by galanin. SP via galanin may modulate anterior pituitary hormone secretion, as contrary to SP, high density of galanin immunoreactivity is present in the median eminence, and by innervating galanin-IR neurons projecting to other parts of the brain, SP can modulate indirectly their activities.
Impact statement
The present study is the first describing juxtapositions between the substance P (SP)-immunoreactive (IR) and galanin-IR neurons in the human hypothalamus. These juxtapositions may be functional synapses and they may represent the morphological substrate of the control of SP on the galaninergic system. SP via galanin may modulate anterior pituitary hormone secretion, as contrary to SP, high density of galanin immunoreactivity is present in the median eminence. Galanin, released into the hypothalamo-hypophyseal circulation, can reach the anterior pituitary and function as a hypophysiotropic substance and regulates anterior pituitary hormone secretion. SP by innervating galanin-IR neurons, which project to other parts of the brain, can modulate indirectly their activities.
Introduction
Substance P (SP) was described almost nine decades ago (von Euler and Gaddum, 1931) and is an undecapeptide that belongs to the tachykinin neuropeptide family along with neurokinin A (NKA) and neurokinin B (NKB). SP is a high affinity substrate of neurokinin 1 (NK1) receptors (Mantyh, 2002), and it also targets neurokinin 2 (NK2) and neurokinin 3 (NK3) receptors with low affinity, which also bind NKA and NKB, respectively.
SP is a neurotransmitter and/or neuromodulator and apart from its well-known role in nociception (Zubrzycka and Janecka, 2000), it is also involved in the physiology of neurogenesis (Park et al., 2007), reinforcement (Huston et al., 1993), respiratory rhythm (Bonham, 1995), vasodilation (Bossaller et al., 1992; Meeking et al., 2000; Schrauwen and Houvenaghel, 1980), and reproduction (Hokfelt et al., 1978; Kalil et al., 2016; Kerdelhue et al., 2006; Lasaga and Debeljuk, 2011); and participates in the pathology of anxiety, stress, mood disorders (Ebner and Singewald, 2006), neurotoxicity, and nausea/emesis (Hesketh, 2001). SP is involved in inflammation and once released it initiates the expression of several cytokines, which in turn induce SP and NK1 receptors [for review see O'Connor et al. (2004)].
Since SP is present in the hypothalamo-hypophyseal axis (Chawla et al., 1997; Dudas and Merchenthaler, 2002, 2006), it is plausible that SP may also modulate neuroendocrine functions by affecting the release of anterior pituitary hormones. SP was the first tachykinin isolated, and this neuropeptide has been implicated in the control of luteinizing hormone (LH) for many years (Lasaga and Debeljuk, 2011). In addition, SP levels in plasma, anterior pituitary, and hypothalamus have been reported to fluctuate in phase with the menstrual cycle or with circulating estradiol levels in the monkey and human (Coiro et al., 1992; Kerdelhue et al., 2000).
Based on our morphological findings that SP-immunoreactive (SP-IR) nerve terminals are present in the median eminence and on galanin-IR neurons in the infundibulum, it is possible that in addition to SP regulating the anterior pituitary hormone section directly via portal circulation, many hypothalamic neuronal circuits can be modulated by SP indirectly, via galanin.
Galanin is a peptide consisting of a chain of 29 amino acids (30 amino acids in humans) (Tatemoto et al., 1983) produced from the cleavage of a 123-amino acid protein known as preprogalanin, which is encoded by the GAL gene (Evans et al., 1993). Galanin is widely expressed in the brain, spinal cord, and gut of humans as well as in other mammalian species.
Galanin signaling occurs through three G protein-coupled receptors (Mitsukawa et al., 2008). Galanin has been implicated in many biologically diverse functions, including nociception, cognition, waking and sleep regulation, feeding, regulation of mood, blood pressure, and reproduction, and it also has roles in development as well as acting as a trophic factor. Galanin appears to have neuroprotective activity as its biosynthesis is increased several fold upon axotomy in the peripheral nervous system and when seizure activity occurs in the brain. It may also promote neurogenesis [for reviews see Hokfelt and Tatemoto (2008); Merchenthaler et al. (1993)]. Galanin is linked to a number of diseases, including Alzheimer's disease, epilepsy as well as depression, eating disorders, depression, and cancer (Berger et al., 2005; Lundstrom et al., 2005).
Galanin is predominantly an inhibitory, hyperpolarizing neuropeptide (Hokfelt et al., 1987) and as such inhibits neurotransmitter release. Galanin is often colocalized with classical neurotransmitters such as acetylcholine, serotonin, and norepinephrine, and also with other neuromodulators such as GnRH, neuropeptide Y, SP, and vasoactive intestinal peptide [for a review see Hokfelt et al. (1987); Melander et al. (1986); Merchenthaler et al. (1993)].
Because of the similar distribution of SP-IR fibers and galanin-IR perikarya in the human hypothalamus (Chawla et al., 1997; Dudas and Merchenthaler, 2002; Kalil et al., 2016; Rance and Young, III, 1991) and the data suggesting functional interaction between these neuropeptides in the peripheral nervous system (Green et al., 1992; Hokfelt et al., 1993; Xu et al., 1990; Yang et al., 2008), the goal of our studies was to identify morphological interactions (close contacts or synapses) between SP- and galanin-IR structures in the human hypothalamus. Because electron microscopy techniques combined with immunolabeling procedures are virtually impossible in humans due to the long postmortem period, we used double-label immunohistochemistry with high-resolution light microscopy to reveal the putative SP-galanin associations in the human hypothalamus.
Materials and Methods
Tissue preparation
Human hypothalamic blocks were obtained from cadavers of three adult women and one man (75 to 87 years of age) within less than a 12-h postmortem period, in accord with the policy of the Institutional Review Board of Lake Erie College of Osteopathic Medicine (LECOM). The clinical records of the individuals did not include any neurological or neuroendocrinological diseases. Hypothalamic blocks were fixed using 0.1 M phosphate-buffered 4% formaldehyde at 4°C for 2–8 weeks. Following fixation, the tissue blocks were trimmed, split along the midsagittal plane, and cryoprotected in 30% sucrose in phosphate buffer containing 0.9% sodium chloride (PBS) and 0.15% sodium azide. Sectioning was performed on a freezing microtome (Model 880; American Optical Corp.).
The 35-micron-thick sections were collected in a sequential order into 24-well plastic tissue culture plates containing PBS with 0.2% sodium azide and then stored at 4°C until processing. Adjoining sections were processed as follows: (1) single-label immunohistochemical detection of SP, (2) single-label detection of galanin, and (3) double-label immunohistochemical detection of SP (first label) and galanin (second label).
Immunohistochemistry
Nonspecific staining was blocked in 2.5% normal horse serum (Vector Laboratories, Burlingame, CA). Then, sections were incubated in one of the primary antisera: rabbit anti-galanin serum (dilution 1:10,000; Abcam, Cambridge, MA) and rabbit anti-SP serum (dilution 1:1000; Millipore Sigma, Burlington, MA) for 24 h at 4°C. Incubation in secondary antiserum containing peroxidase-labeled ready-to-use horse anti-rabbit IgG (ImPRESS HRP reagent kit; Vector Laboratories) followed this step for 1 h at room temperature. After three changes of wash in PBS, the sections were immersed into the chromogen solution until sufficient staining was accomplished (Vector SG chromogen kit; Vector Laboratories).
For simultaneous detection of SP and galanin, we used double-label immunohistochemistry. The first signal (SP immunoreactivity) was revealed by using the black Vector SG chromogen (Vector Laboratories), while the second signal (galanin immunoreactivity) was visualized with NovaRED (Vector Laboratories). The main advantage of using chromogens over fluorescence markers is their stability that allows storing the stained sections for extended periods of time. No immunoreaction was observed in control sections where primary antibodies were omitted or replaced by nonimmune rabbit sera at the dilution used for the primary antibodies.
Microscopic analysis
SP-galaninergic juxtapositions were identified using the Olympus BX45 microscope equipped with 100 × oil immersion objective and a digital camera (Olympus DP12). If the neurons and the corresponding processes were larger than the frame of the camera, composite images of the adjacent micrographs were created by Adobe Photoshop software (Adobe Creative Suite 2.0; CS2). To analyze the relationship of galanin-IR neurons with the SP-IR fibers, representative coronal sections, which were identical in each tissue sample, were used. The galaninergic neurons were subdivided into three subclasses: densely innervated neurons (more than five contacting SP fiber varicosities), lightly innervated neurons (one to five contacts), and perikarya that did not receive any contacting SP fibers.
Computer-assisted mapping
Sections were mounted, coverslipped, and scanned on a digital slide scanner (PathScan Enabler IV; Meyer Instruments, Inc.). The galaninergic and SP-IR neurons were denoted on the images using an Olympus BX45 microscope equipped with camera lucida and Adobe Photoshop software (Adobe Creative Suite 2.0; CS2). The distribution of SP-IR and galaninergic perikarya and fibers has been compared with previous studies (Chawla et al., 1997; Dudas, 2017; Dudas and Merchenthaler, 2002, 2006; Dudas et al., 2013) and no significant discrepancy was found between the published and present data.
The three-dimensional (3D) models of the SP and galaninergic systems were created by the computer-aided stacking of the maps of consecutive sections using VoxBlast NT/9x Version 3.0 Light (Vaytek, Image Analysis Facility, University of Iowa). The overlapping areas between the SP and galaninergic systems have been identified by superimposing the 3D maps of these neurotransmitter systems.
Results
SP-IR structures in the human hypothalamus
The hypothalamic SP system has been described by our prior studies (Dudas, 2017; Dudas and Merchenthaler, 2002, 2006; Dudas et al., 2013) that was in general agreement with data by others (Chawla et al., 1997). In essence, SP-IR neurons generally populate the tuberal region of the human hypothalamus, where they form subclusters in the basal perifornical area, dorsomedial subdivision of the ventromedial nucleus, basal part of the periventricular area, and in the infundibular nucleus/median eminence (Fig. 1). Small numbers of SP-IR perikarya can be observed in the periventricular zone of the preoptic area and in the basal part of the posterior hypothalamus. These neurons are generally fusiform, although multipolar cells can also be observed in the medial hypothalamus.
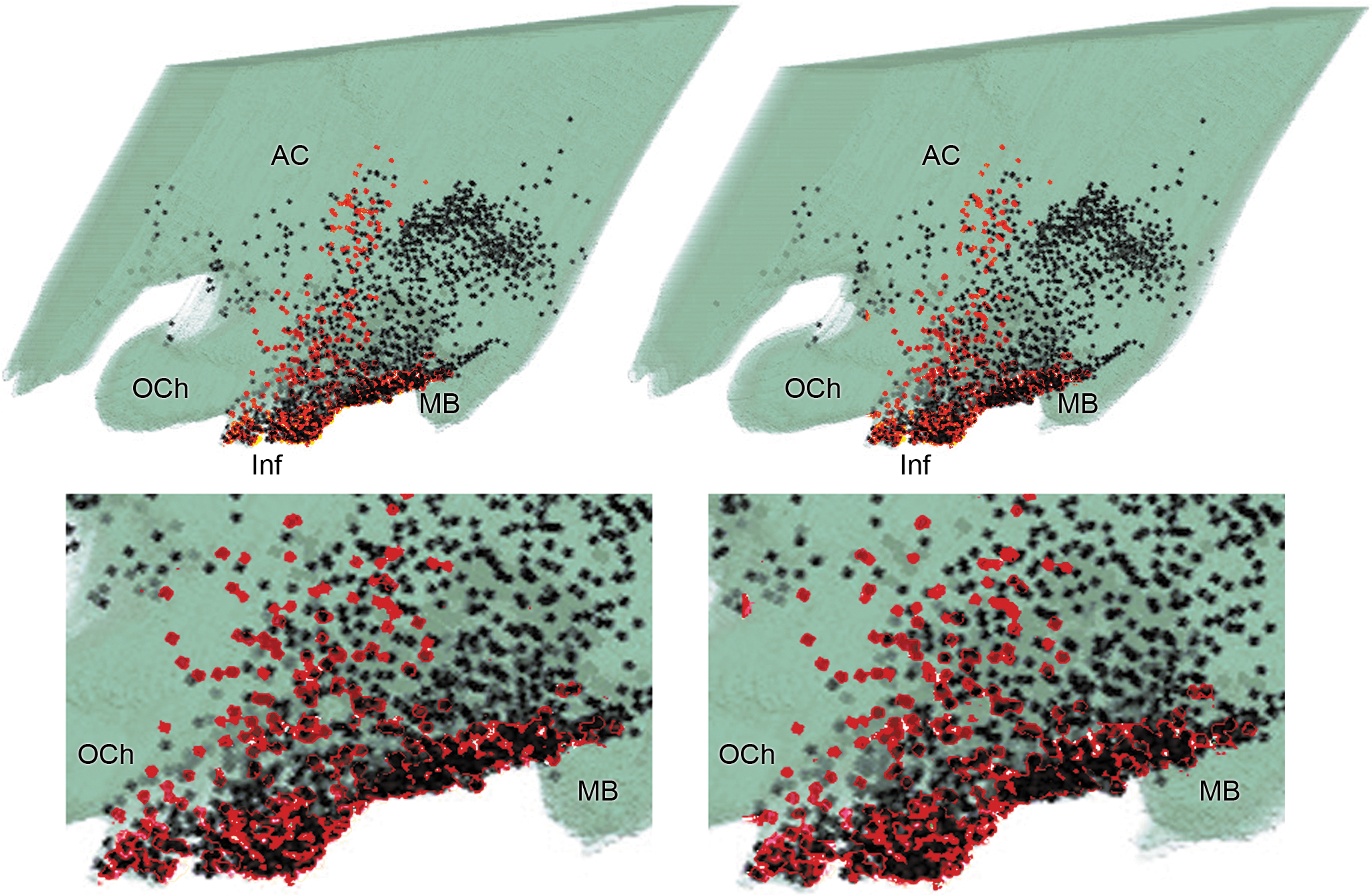
Stereoscopic image of the human diencephalon reconstituted from 30-μm -hick sections, illustrating the distribution of galaninergic neurons (black dots) and SP-IR perikarya (red dots). Lower inserts illustrate the infundibular area with higher magnification. Stereoscopic images can be seen using U or parallel vision. The eyes are relaxed to look into the distance until the pair of the images fuse, and then refocused by the brain. With this technique, a 3D hypothalamus can be seen on the figure, floating in front of the paper, with the immunolabeled perikarya in it at different depths marked by the dots. The optimal viewing distance is about 50 cm from the paper surface (average reading distance). A stereoscopic viewer, or stereoscope, is suggested to readers unfamiliar with U or parallel vision (a stereoscope is an inexpensive device for viewing a stereoscopic pair of separate images, depicting left-eye and right-eye views of the same scene, as a single 3D image). 3D, three-dimensional; AC, anterior commissure; Inf, infundibulum; MB, mammillary body; OCh, optic chiasm; SP-IR, substance P-immunoreactive.
A network of SP-IR fiber varicosities can be found in the basal infundibulum, often in the close proximity of the portal vessels. Periventricularly arranged SP-IR axonal varicosities populate the preoptic and tuberal regions. IR fibers can also be observed in the basal part of the lateral hypothalamus over the optic tract, along the diagonal band of Broca and surrounding the fornix and the anterior commissure and the fornix. Occasional fiber varicosities can be detected in the paraventricular nucleus. SP-IR fiber varicosities often abut on SP-IR perikarya as well as on fusiform neurons that are apparently not SP-IR in the basal part of the infundibulum.
Galanin-IR structures in the human hypothalamus
The distribution of the galanin-IR neurons in the human hypothalamus has been previously detailed in our preceding studies (Dudas and Merchenthaler, 2004, 2006; Merchenthaler et al., 2010, 2013). Concisely, the majority of the galanin-IR perikarya populate the infundibulum/median eminence, particularly the infundibular nucleus in humans (Fig. 1). In addition, a significant number of galanin-IR perikarya are present periventricularly in the tuberal region that extends to the preoptic region and the posterior hypothalamus as well. The basal perifornical area, the ventromedial and dorsomedial subdivisions of the ventromedial nucleus, and the paraventricular nucleus also contain galaninergic neurons.
In addition to the posterior periventricular cell group, galanin-IR neurons are also present superior to the mammillary nuclei in the posterior hypothalamus. Dense network of galanin-IR fiber varicosities and occasional nerve terminals can be observed in the infundibulum/median eminence as well as periventricularly extending through the entire hypothalamus, while the lamina terminalis, diagonal band of Broca, and the basal perifornical area also contain smaller number of galaninergic fibers.
SP-galanin associations
The majority of galaninergic neurons receive contacting SP-IR fibers in the human hypothalamus, typically forming en passant type associations where SP-IR fibers form multiple contacts with galaninergic neurons while passing by. These contacts can be confirmed by high-magnification microscopical analysis with 100 × oil immersion objective revealing no gaps between the contacting elements. Most of these juxtapositions can be observed in the infundibulum/median eminence (Fig. 2A, B, F–H, J, K) and in the basal periventricular area of the tuberal region (Fig. 2E, I) where SP-IR fiber varicosities frequently surround the galaninergic perikarya and the emanating axons/dendrites typically following the contours of the target neurons.

Juxtapositions between the SP-IR (black) and galanin-IR (brown) neural elements are revealed using double-label immunohistochemistry in the human diencephalon. The SP-IR fiber varicosities often abut on the galaninergic neurons forming numerous en passant type contacts. Asterisks on the coronal sections of the diencephalon in the corners of the micrographs denote the positions of the demonstrated galaninergic cells. The thickness of the sections is 30 μm. Scale bar: 20 μm. These juxtapositions (arrows) are typically located in the infundibulum/median eminence
These basket-like encasements that cover a significant amount of surface area of the perikarya, with more than five contacts/cell, are characteristic of the 23% of the counted galaninergic neurons. Numerous galaninergic perikarya appear to be lightly innervated by SP-IR fibers, receiving one to five contacts (48% of the counted neurons), while 29% of the counted galanin-IR neurons do not appear to be associated with SP-IR fiber varicosities. Numerous SP-galaninergic contacts can be observed in the basal perifornical area of the tuberal area (Fig. 2C, D), while SP-IR fibers typically avoid galaninergic neurons in the posterior hypothalamus. The morphology and distribution of these SP-galaninergic juxtapositions do not show significant gender/age differences when comparing the individual samples.
Discussion
The distribution of the SP- and galanin-IR structures in the human hypothalamus is in good agreement with our previous studies (Dudas and Merchenthaler, 2004, 2006; Luu et al., 2020; Merchenthaler et al., 2010, 2013; Uhlman et al., 2019). Although the morphology of the SP-galanin juxtapositions reported in this article appears to support our suggestion that these associations represent functional synapses, due to the long postmortem period before preserving these antigens, we were unable to verify the synaptic structures by electron microscopy. However, the density of the abutting SP-IR axonal varicosities on the surface of galanin-IR neurons strongly suggests that these neurotransmitter systems indeed communicate at certain hypothalamic regions, through which the SP may modulate the activity of hypophysiotropic and nonhypophysiotropic neurons, that is, the activity of neurons regulating anterior pituitary hormone release and the activity of neurons interacting with other neurons participating in different neuronal circuits and functions.
SP and galanin are two neuropeptides with diverse, and many times opposite, functions in both the central and peripheral nervous systems. For example, acting in the hypothalamus, both galanin and SP increase the food intake and galanin regulates glucose metabolism and can alleviate symptoms of type 2 diabetes (Fang et al., 2020; Karagiannides et al., 2008) in rodents. SP stimulates the appetite and promotes weight gain in rodents and its effect is independent of leptin (Karagiannides et al., 2008); however, others reported opposite effects in rodents (Dib, 1999) and birds (Pauliukonis et al., 2020). In addition to their effect on food intake, both galanin and SP participate in gonad regulation by modulating LH secretion. Galanin stimulates LH secretion (Lopez et al., 1991), while SP can stimulate or inhibit it depending on the hormonal status (Kalil et al., 2016).
Outside of the hypothalamus, both peptides are involved in the regulation of pain, but while galanin plays an inhibitory role in pain processing (Hobson et al., 2010; Mitsukawa et al., 2010; Xu et al., 2010), SP is an important factor of pain perception by transmitting pain information from primary afferents to the central nervous system. SP is also released from peripheral nerve terminals of sensory nerve fibers in the skin, joints, and muscles and it is involved in neurogenic inflammation [for reviews see Feickert and Burckhardt (2019); Mashaghi et al. (2016); Schank and Heilig (2017); Zieglgansberger (2019)].
Galanin as an inhibitory peptide has been shown to weaken learning and cognition (Mitsukawa et al., 2010), decreases seizure threshold (Mitsukawa et al., 2008), and therefore acts as an anticonvulsant (Mazarati, 2004; Zhang et al., 2009), while SP causes anxiety, stress (Ebner and Singewald, 2006), and reinforcement (Huston et al., 1993). Galanin seems to have antidepressant activity by stimulating the hypothalamo/pituitary/adrenal axis resulting in elevated glucocorticoid secretion (Kuteeva et al., 2010). SP, however, induces anxiety and depression (Ebner and Singewald, 2006).
The role of galanin and SP in Alzheimer's disease and addiction is rather controversial. Hyperinnervation of cholinergic neurons by galanin in the septum of Alzheimer's disease (AD) patients (Counts et al., 2003, 2008a, 2008b) may result in reduced cholinergic transmission (Counts et al., 2008a; Mitsukawa et al., 2008) affecting cognition and memory, but it is also possible that galanin hyperinnervation may reduce differentiated functions of these cholinergic basal forebrain neurons and help them to recover from the toxic effect of beta-amyloid (Aβ) (Ding et al., 2006).
SP may also play an important role in AD since reduced levels of SP were found in certain brain areas and spinal fluid of AD patients. In addition to its neuroprotective properties, it was recently demonstrated that SP is able to stimulate nonamyloidogenic APP processing, thereby reducing the possibility of generation of toxic Aβ peptides in the brain. Recent studies, using in vitro and in vivo models, have also shown that the neuroprotective role of SP against Aβ could be related to its ability to modulate Kv channel currents [for reviews see Campolongo et al. (2013); Severini et al. (2016)].
Both galanin and SP play a role in addiction. Galanin is involved in repeated alcohol intake. Along with this effect, galanin has been shown to play a role in addiction to nicotine and opiates (Genders et al., 2020; Picciotto, 2010). SP and its principal receptor NK1 play a specific role in the behavioral response to opioids and stress that may help to initiate and maintain addictive behavior. In animal models, the NK1 receptor is required for opioids to produce their rewarding and motivational effects. SP neurotransmission is also implicated in the behavioral response to stress, and in the process of drug sensitization potentially contributing to vulnerability to addiction or relapse [for a review see Commons (2010)].
Based on these diverse, many times complementary, effects of galanin and SP on various hypothalamic functions, it appears that these two neurotransmitter systems interact with each other. Galanin may play an important role in the regulation of anterior pituitary hormone secretion either directly, acting as a hypophysiotropic peptide released into the hypophyseal portal circulation in the median eminence, or indirectly, acting as a neurotransmitter/neuromodulator by innervating other hypophysiotropic neuronal populations, including luteinizing hormone-releasing hormone (LHRH), growth hormone-releasing hormone (GHRH), somatostatin, thyrotropin-releasing hormone, and corticotropin-releasing hormone [reviewed in Merchenthaler et al. (1993)]. Indeed, the dense accumulation of galanin in the external zone of the median eminence and the findings that these terminals release galanin into the hypophyseal portal circulation and regulate anterior pituitary hormone secretion (Lopez et al., 1991; Merchenthaler, 1991) strongly suggest that galanin acts as a hypophysiotropic hormone. However, considering the dense communication of galanin with other hypothalamic and extrahypothalamic neuronal systems (Merchenthaler et al., 1993) also supports the role of galanin as a neurotransmitter/neuromodulator.
SP may also play a role in the regulation of anterior pituitary hormone secretion, however, because of the sparse presence of SP in nerve terminals in the median eminence and the low concentrations of SP in the hypophyseal portal blood, it is likely that SP does not play a significant role as a hypophysiotropic peptide, rather, it regulates anterior pituitary hormone secretion indirectly, for example, via galanin. Although both peptides are released into the hypophyseal portal blood and regulate anterior pituitary hormone secretion, their concentrations in the hypophyseal portal blood are quite different. In the ewe, collection of portal blood is relatively simple compared with other species. In this species, the concentration of SP in portal blood is 4.9 pg/mL and the concentration of galanin is up to 1700 pg/mL depending on the hormonal status of the animal (Clarke et al., 1993). These large differences correspond well with the immunocytochemical data showing only a few SP-IR nerve terminals in the median eminence compared with a dense accumulation of galanin-IR nerve terminals in this region [(Merchenthaler, 2010) for galanin and (Kalil et al., 2016; Mikkelsen et al., 1989) for SP].
The concentrations of the classical hypophysiotropic peptides are in the 40–100 pg/mL range. For example, the concentration of GHRH is 40–60 pg/mL, the concentration of somatostatin is 80–100 pg/mL (Fletcher et al., 1996), but the concentration of LHRH in human portal blood, collected via transpharyngeal surgery, is high, 2000 pg/mL (Antunes et al., 1978), but significantly lower in rams: 30–100 pg/mL (Caraty and Locatelli, 1988). The low concentration of SP in the portal blood suggests that the efficacy of SP in regulating anterior pituitary hormone secretion directly in the pituitary is low and the hypothalamic SP functions primarily as a neurotransmitter/neuromodulator affecting the activity of other neurotransmitter systems, including galanin. Subsequently, galanin can be released into the portal circulation and/or innervate other neurotransmitter-containing neurons with diverse intra-and extrahypothalamic connections. Our morphological findings regarding SP-IR nerve terminals densely contacting galanin-IR perikarya and nerve processes in the human hypothalamus provide strong support for this hypothesis. By interacting with galanin, as well as many other neurotransmitters, SP may have a more widespread regulatory capacity than acting alone on different neurotransmitter systems.
Conclusion
In conclusion, the present study has revealed that the majority of galaninergic neurons are closely associated with contacting SP-IR fiber varicosities in the basal part of the human hypothalamus. These juxtapositions appear to be functional synapses and they may represent the morphological substrate of the control of SP on the galaninergic system in humans.
Footnotes
Acknowledgment
The authors are indebted to Mr. George Grignol for the technical support.
Compliance with Ethical Standards
Authors' Contributions
B.D.: Study design, immunohistochemistry, interpretation of the results, and article preparation. I.M.: Interpretation of the results and article preparation.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.