
Abstract
Introduction:
Acute central nervous system (CNS) injury, such as stroke, spinal cord injury, or traumatic brain injury can result in dysregulated immune response, and the condition is known as CNS injury-induced immunodeficiency syndrome (CIDS). The endocannabinoid system is an important homeostatic regulator in the CNS and immune system. Activation of cannabinoid 2 receptors (CB2R) on immune cells has been reported to dampen inflammation, suggesting a potential role of CB2R in the peripheral immune response following CNS injury. In this study, we have investigated the effect of CB2R modulation on the peripheral immune response during CIDS.
Materials and Methods:
Experimental CNS injury was induced in C57BL/6 mice through intracerebral injection of the vasopressor peptide, endothelin-1. A selective CB2R agonist (HU308) was used as an early treatment before the onset of CIDS and AM630, a selective CB2R antagonist, was administered as a later-phase therapy to combat the systemic immunodeficiency following the CNS injury. The peripheral immune response to endotoxin was studied 24 h after the CNS injury using intravital microscopy to examine leukocyte activation within the intestinal microcirculation in mice. Brain infarct size, and plasma levels of cytokines and soluble adhesion molecules were measured as additional parameters for the assessment of treatment outcomes.
Results:
Our results showed that early CB2R activation with HU308 reduced brain injury size and restored leukocyte response to endotoxin in the peripheral microcirculation. Late CB2R inhibition with AM630 also improved the peripheral leukocyte response to endotoxin and did not exacerbate the extent of brain injury.
Discussion:
CB2R activation has the potential to mitigate CNS injury as an early treatment by limiting neuroinflammation and preventing the development of CIDS. At the later stage with already-established CIDS, treatment may require dampening CB2R activation to improve the patient's outcome.
Introduction
Central nervous system (CNS) injury is one of the primary causes of disability and mortality worldwide and includes conditions such as stroke, spinal cord injury, and traumatic brain injury.1,2 Globally, stroke ranks the second leading cause of death, 3 the third leading cause of disability,3,4 and up to 85% of stroke patients suffer from complicating medical events during the acute care. 5 Currently, acute stroke therapy is limited to reperfusion approaches and high-end medical support in dedicated stroke units. 6 Stroke patients are highly susceptible to develop infections and around 30% of poststroke infections occur within the first week of stroke. 7
A strong inflammatory response is mounted in the brain immediately after the onset of CNS injury. 2 The damaged brain promotes local immunosuppression to minimize secondary damage to healthy CNS tissue8,9 and subsequent development of systemic immunosuppression (CNS injury-induced immunodeficiency syndrome [CIDS]), 2 which is suggested to be the main reason for infections in patients with acute CNS injury.2,10 Secondary infection associated with post-CNS injury adversely affects the neurological recovery, causes additional invasive medical interventions, and hospitalizations for affected individuals.11–13
The endocannabinoid system (ECS) comprises endocannabinoids, the enzymes involved in the synthesis and degradation of endocannabinoids, and two identified G-protein-coupled receptors—cannabinoid 1 receptor (CB1R) and cannabinoid 2 receptor (CB2R). 14 CB2R is mostly expressed on immune cells.15,16 Activation of CB2R by endocannabinoids and cannabinoid ligands suppresses the production of proinflammatory cytokines. 17 Following injury or in inflammatory conditions, CB2R expression is upregulated, including in the CNS,15,18,19 suggesting the protective role of this receptor under these conditions.
Early treatment with a CB2R agonist suppressed the extent of neurodegeneration during the early proinflammatory phase in a murine stroke model. 19 CB2R activation decreased the size of cerebral infarction in a mouse model of focal ischemia. 20 However, sustained CB2R activation might be responsible for the abovementioned development of CIDS. Consequently, CB2R inhibition might be beneficial in the restoration of the suppressed systemic immune function in the later phase after CNS injury. 21
In the current research, we are investigating the effect of ECS modulation through the CB2R pathway on peripheral immune response following experimental CNS injury in mice. We have hypothesized that targeting CB2R with an agonist as an early treatment of CNS injury will reduce the immunosuppressive response immediately after the CNS injury and will limit the extent of brain injury and severity of CIDS. We have also hypothesized that restoration of the immune function and improvement of the host's ability to escalate an immune response to a challenge can be achieved by suppression of the peripheral CB2R activity after the CNS injury and before the onset of CIDS.
Materials and Methods
Animals
C57BL/6 (6–8 weeks old, male) and DeltaGen CB2R null mice were purchased from Charles River Laboratories International, Inc. (Wilmington, MS, USA) and Jackson Laboratories (Bar Harbor, Maine, USA), respectively. An in-house colony was established for CB2R null mice at the Carleton Animal Care Facility, Dalhousie University, Halifax, Nova Scotia, Canada. Offspring were generated by mating two homozygous CB2R null parents, and male mice reaching 6–8 weeks of age were used for experiments. CB2R null mice were genotyped using the DNA extracted from ear punches using the Accustart II Mouse Genotyping Kit (Quanta Bioscience, MD, USA) following the manufacturer's instructions. Animals were kept on a standard 12-h light/12-h dark cycle in standard room temperature of 22°C and 55–60% humidity. All animal-related procedures were conducted following the guidelines and standards set forth by the Canadian Council for Animal Care and approved by the Dalhousie University Committee on Laboratory Animals.
CNS injury model
Endothelin-1 (ET-1)-induced CNS injury model was used in the study. Endothelin is a potent vasoconstrictive peptide 22 that causes a rapid decrease in cerebral blood flow (CBF),23–25 leading to the formation of the ischemic lesion followed by gradual reperfusion. This situation closely resembles the CBF reduction and reperfusion that occurs in clinical strokes.26,27 Mice were anesthetized and maintained with isoflurane gas (2–5%) throughout the surgery. The animals were placed in a stereotactic frame (Model 900; Kopf, USA) and connected to a facemask delivering isoflurane. A 1-cm vertical skin incision was made in between the bregma and the lambdoid suture, the skin was retracted, and two holes were drilled in the skull, one 2.3 mm posterior and 1.0 mm left of bregma, the other 2.3 mm posterior and 1.0 mm right of bregma. The stereotaxic coordinates were set relative to bregma at (1) 0.0 anterior–posterior (AP), +1.50 medial–lateral (ML), −2 dorsal–ventral (DV) and (2) 0.0 AP, +1.75 ML, −2 DV. Intracerebral injection of ET-1 (Sigma-Aldrich, Canada) was given using a microinjection unit (Model 5000; Kopf) with a Hamilton Neuros® 7001 syringe (Hamilton Company, NV, USA). The incision was sutured with 5-0 polypropylene. The animal was transferred to a heated recovery cage with access to food and water ad libitum. The sham animals in the experiments got skin cut and holes on the skull and sterile vehicle (saline) was injected.
Endotoxin challenge
Intraperitoneal injection of lipopolysaccharide (LPS) endotoxin (5 mg/kg; Sigma-Aldrich, St. Louis, MO, USA) was used to generate endotoxemia in mice.
CB2R pharmacological modulation
A selective synthetic CB2R antagonist AM630 (Tocris Bioscience, Ellisville, MO, USA) was used for the selective inhibition of CB2R. For activation of CB2R, a selective synthetic CB2R agonist HU308 (Tocris Bioscience) was used. AM630 and HU308 were dissolved in 30% and 10% dimethyl sulfoxide (DMSO) vehicle, respectively, and administered as a dose of 2.5 mg/kg through tail vein injection to the mice.
For the CB2R agonist treatment, HU308 was administered 15 min before CNS injury induction and LPS was administered at 22 h of CNS injury induction. For the CB2R antagonist treatment, LPS was administered at 22 h after CNS injury induction and AM630 was administered 15 min before the LPS administration. In both cases, intravital microscopy (IVM) was performed at 24 h of CNS injury induction (Fig. 1). In pilot experiments, DMSO administration did not cause any changes in IVM parameters and brain infarct size compared with CNS-injured mice treated with normal saline.
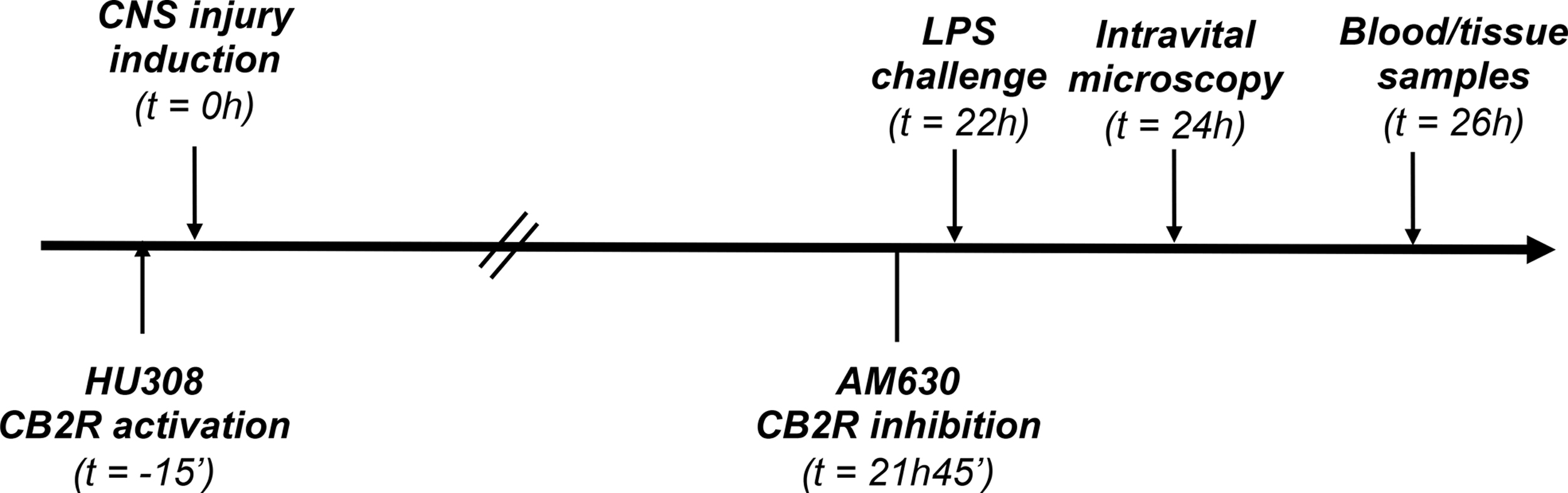
Experimental timeline for CB2R modulation treatments. CB2R, cannabinoid 2 receptor.
Intestinal IVM
Animals were anesthetized by intraperitoneal injection of sodium pentobarbital (90 mg/kg, 54 mg/mL; Ceva Sante Animale, Montreal, Quebec, Canada). Laparotomy was performed and a part of the small intestine, terminal ileum, was exteriorized onto a heated platform of a specially designed apparatus for IVM. The animal was placed on its side, the terminal ileum on the platform was covered with a coverslip, and perfused with continuous thermostat-controlled (37°C) saline to maintain the physiological conditions of temperature and hydration. The heating pad with the intravital stage containing the animal and the intestine on the platform was placed under an epifluorescent microscope to observe intestinal leukocyte activation and capillary perfusion.
A black and white DAGE CCD video camera (DAGE MTI, Inc., Michigan City, IN, USA) was used for recording videos directly onto a computer through an analog-to-digital video converter (DFG/USB2PRO; The Imaging Source, Germany) and image-capturing software (IC capture; The Imaging Source, Germany).
Two hundred microliters of 0.05% rhodamine-6G solution and 200 μL 5% fluorescein isothiocyanate–bovine serum albumin was given to mice by tail vein injection to visualize leukocytes and microvascular perfusion, respectively. Six visual fields comprising nonbranching, submucosal collecting venules, measuring at least 300 μm in length, and another six visual fields containing similar postcapillary venules were observed and recorded for 30 sec each in every animal. Leukocytes that remained adherent to the endothelial cells during the 30 sec were considered as adherent leukocytes. Six visual fields of the intestinal muscle layer and six fields of the mucosa were selected and recorded for 30 sec for each field. Postcapillary venules were categorized as V1 (>50 μm in diameter) and V3 (<50 μm in diameter) venules. Functional capillary density (FCD) was analyzed using ImageJ to evaluate vascular perfusion. FCD was defined as the total length of capillaries with observable erythrocyte perfusion to a predetermined rectangular field.
Assessment of infarct volume
Blood was collected from mice by cardiac puncture under general anesthesia and animals were euthanized by cervical dislocation. The brain was extracted postmortem. The brain was placed in a three-dimensional printed brain matrix with slots for surgical blades to be placed in, cooled at −20°C for 10 min, sliced into 1-mm thickness, and stained with 2% 2,3,5-triphenyltetrazolium chloride (TTC) for 12 min at 37°C. The slices were digitally scanned, and the infarct sizes were analyzed using ImageJ (version 1.47v). Infarct volume was estimated based on the thickness of each slice.
Plasma cytokine analysis
Plasma levels of proinflammatory cytokines and adhesion molecules were measured using the 12-plex Magnetic Immunoassay Kit (Luminex® Assays; R&D Systems, Minneapolis, MN, USA) following the manufacturer's instructions. A Bio-Plex® 200 System and Bio-Plex Manager 6.1 software (Bio-Rad, Mississauga, Canada) were used for analyzing specific cytokine analytes.
Neutrophil isolation and count
Polymorphonuclear neutrophils (PMNs) were isolated from femurs and tibias of mice. Bone marrow cells were flushed out with Hank's Balanced Salt Solutions (HBSS) (1 mM ethylenediaminetetraacetic acid, 10 mM 4-[2-hydroxyethyl]-1-piperazineethanesulfonic acid, 2% fetal bovine serum), cells were centrifuged at 300×g for 10 min at 4°C and resuspended in HBSS. PMNs were isolated from single-cell suspensions of bone marrow by negative selection using the EasySep™ Mouse Neutrophil Enrichment Kit and manual EasySep magnet (StemCell Technologies, Vancouver, Canada) following the manufacturer's instructions. The number and purity of the isolated neutrophils were assessed using a compound microscope.
Data analyses and statistics
Statistical analyses were performed using GraphPad Prism 6.0 (GraphPad Software, Inc., La Jolla, CA, USA) and resulting data were presented as mean±standard deviation (SD). The normality of the data was evaluated using Kolmogorov–Smirnov test. One-way analysis of variance (ANOVA) was used to analyze the differences between groups and group-wise comparisons were done using the Newman–Keuls test. Unless otherwise stated, data comparing two groups were assessed by a two-tailed t-test. The significance level was considered at p<0.05.
Results
CB2R null mice
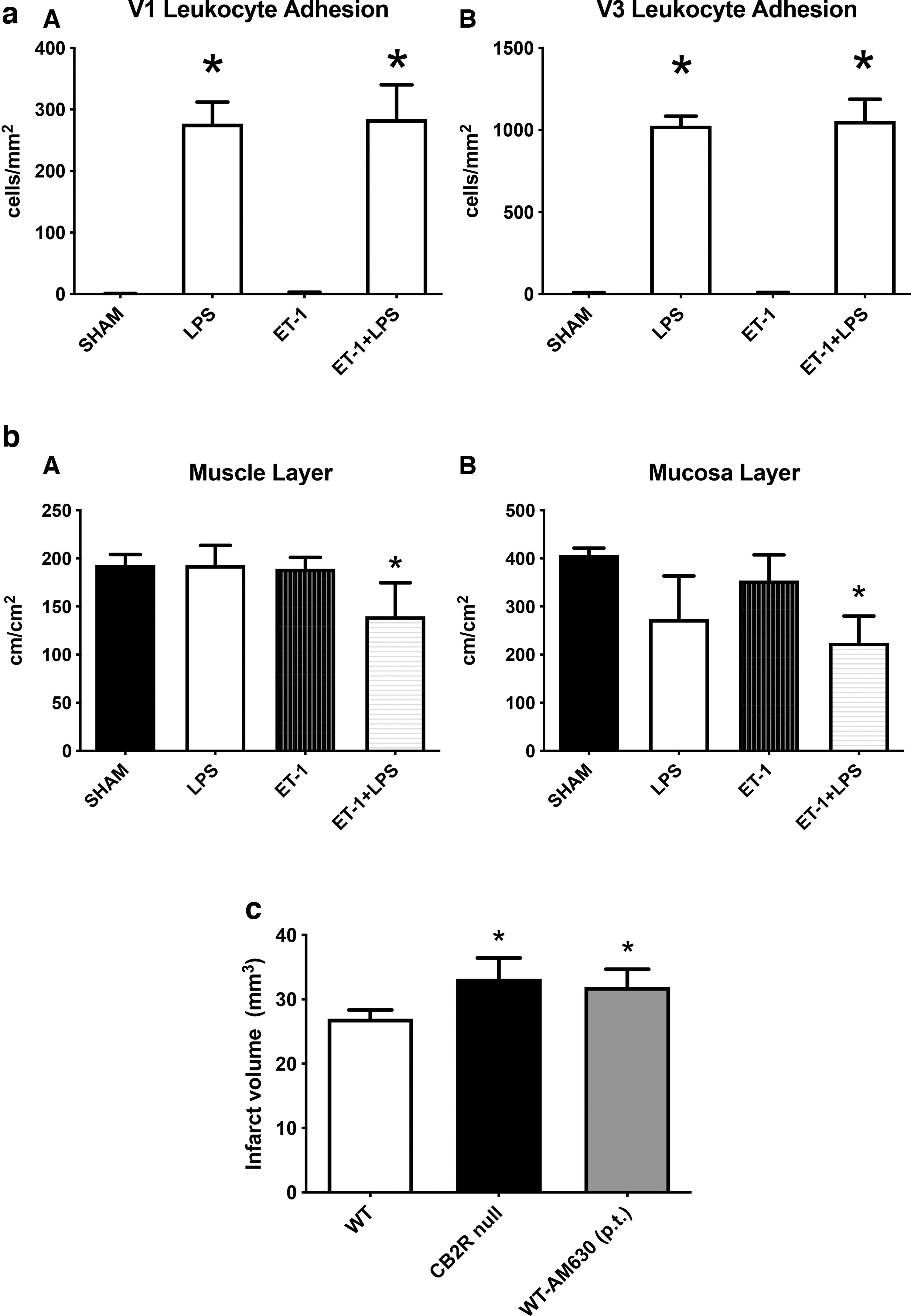
Sham animals and animals with CNS injury displayed minimal numbers of adherent leukocytes in collecting and postcapillary venules of the small intestine, which was significantly increased upon LPS administration (Fig. 2aA and B). Endotoxin challenge in the null mice with CNS injury showed similar levels of adherent leukocytes to that of LPS-challenged null mice without CNS injury. CNS injury did not make significant changes in the FCD in muscle and mucosal villi (Fig. 2bA and B) compared with the sham animals. Endotoxemia in animals with CNS injury significantly reduced the FCD of both regions of the small intestine compared with that of the sham animals. CNS injury significantly increased the size of infarct volume both in CB2R null mice and in mice where CB2R was pharmacologically inhibited by AM630 (Fig. 2c).
Early CB2R activation in CNS injury in wild-type mice
Adherent leukocytes were minimal in numbers in collecting and postcapillary venules of the small intestine of sham animals (Fig. 3aA and B). Animals with CNS injury had a similarly low level of adherent leukocytes as the sham animals. CNS-injured animals demonstrated a significant reduction in leukocyte adherence after LPS administration when compared with endotoxemia animals without CNS injury. Treatment with HU308 before CNS injury and endotoxin challenge prevented the decrease in the number of adhering leukocytes in collecting venules when compared with the untreated animal group with CNS injury and endotoxemia.
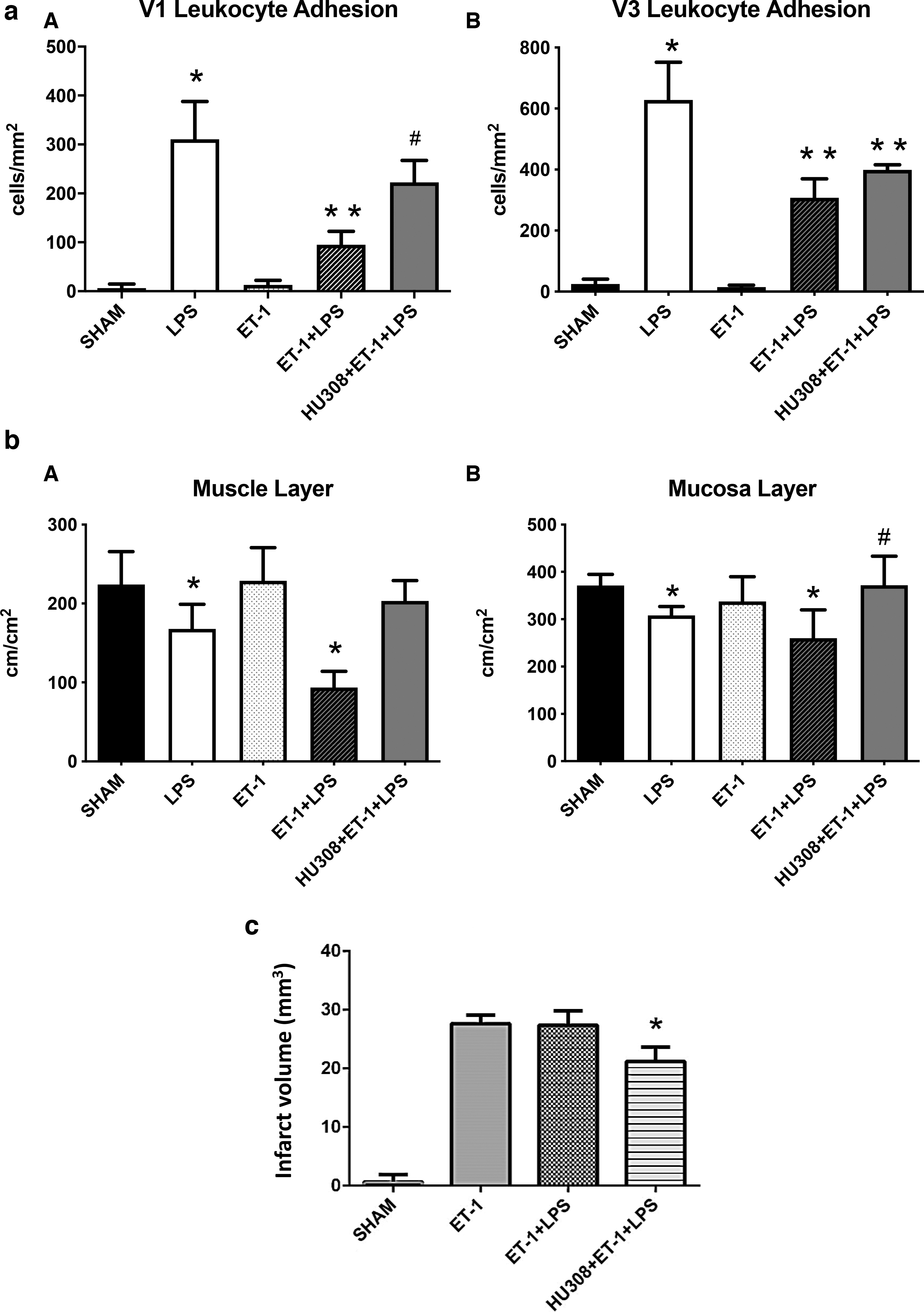
Endotoxemia in animals with CNS injury caused a significant reduction of the FCD in muscle and mucosal layers compared with that of the sham group (Fig. 3bA and B). CB2R activation with HU308 significantly improved the FCD of mucosa when compared with the mice with CNS injury and endotoxemia without the cannabinoid treatment.
Sham animals did not show the presence of cellular death in the brain tissue (Fig. 3c). ET-1 injection caused cerebral infarction, both in the presence and absence of endotoxin challenge. Activation of CB2R with HU308 in animals with CNS injury and endotoxemia significantly reduced the extent of brain injury compared with the untreated animal groups with CNS injury and endotoxemia.
Late CB2R inhibition after CNS injury in wild-type mice
Inhibition of CB2R by AM630 in CNS-injured animals and later on challenging the animals with endotoxin significantly restored the number of adherent leukocytes in collecting venules when compared with untreated animals under similar conditions (CNS injury and endotoxemia) (Fig. 4aA and B).
AM630 administration significantly improved the FCD in the muscle layers of the small intestine when compared with the sham animals. FCD of the mucosal layer was increased to a small extent upon treatment with AM630, but the change was not significant compared with the FCD of the intestinal mucosa of the sham animals (Fig. 4bA and B).
Inhibition of CB2R in animals with CNS injury and endotoxemia did not increase the infarct volume compared with the untreated group with CNS injury (Fig. 4c).
At 24 h of induction of CNS injury, the average total neutrophil count in the bone marrow was significantly reduced compared with the sham animals, which was elevated upon administration of AM630 (Fig. 4d).
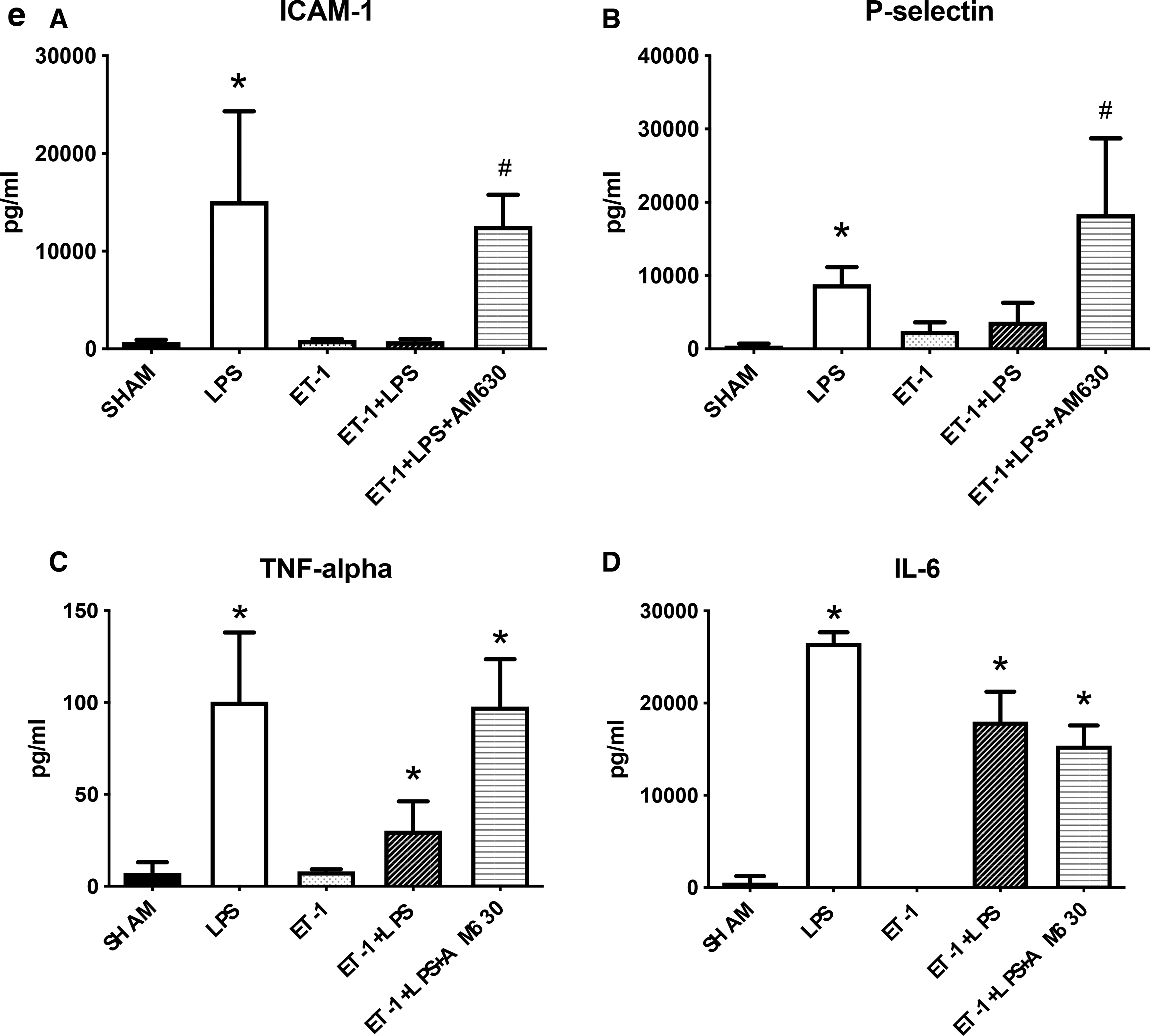
LPS-administered mice had significantly higher levels of P-selectin, intercellular adhesion molecule 1 (ICAM-1), tumor nuclear factor alpha (TNF-α), and interleukin-6 (IL-6) in plasma compared with the sham group (Fig. 4e). Treatment with AM630 in animals with CNS injury and endotoxemia caused significant elevation of P-selectin and ICAM-1 levels when compared with the untreated animals with endotoxemia and CNS injury.
Discussion
Our study has demonstrated that early CB2R activation with HU308 before stroke lessens the extent of brain lesions in a murine stroke model and reduces the severity of subsequent systemic immune suppression during CIDS, whereas CB2R inhibition with AM630 at the later phase of CIDS restores peripheral immune function without causing any exacerbation of the brain injury at the observed time point.
We utilized null mice in our study to validate our therapeutic target, that is, CB2R. We did not see any change in the systemic immune response in CB2R null mice with CNS injury compared with CB2R null animals without CNS injury following endotoxin challenge. The lack of suppression of the systemic immune response indicated that CB2R null animals did not develop CIDS. In contrast, knocking out CB2R or pharmacologically inhibiting the CB2R by AM630 resulted in exacerbation of brain damage after CNS injury, indicative of the potentially protective role of CB2R in the brain following a CNS injury.
Intracerebral injection of ET-1 induced CIDS in experimental wild-type animals in our study. A pathological decrease in the peripheral immune response was confirmed by reduced leukocyte adhesion in the intestinal microcirculation, and reduced levels of soluble adhesion molecules (ICAM-1 and P-selectin) and inflammatory cytokines (TNF-α and IL-6) in endotoxin-challenged animals with CNS injury compared with the endotoxin-challenged animals without CNS injury. HU308 administration before the onset of CNS injury restored the leukocyte activation in the intestinal microcirculation of mice with CNS injury, indicative of diminution of the extent of immunosuppression in animals suffering from CIDS. Peripheral immunosuppression is a consequence of an initial hyperinflammatory state within the brain after CNS injury.2,10 If a treatment approach is capable of combating this inflammatory response within the CNS at an early stage of CNS injury, it will subsequently reduce the extent of systemic immunosuppression throughout the later pathophysiological stages.
Some previous studies reported the attenuation of the activated immune response within the brain upon CB2R stimulation. In a rodent cerebral ischemia/reperfusion injury model, treatment with selective CB2R agonists, O-1966 and O-3853, suppressed the severity of immune activation within the CNS, which was evident by reduced leukocyte–endothelial interaction in the pial vessels of CNS-injured mice. 20 In another study, the selective CB2R agonists, JWH133 and O-1966, decreased leukocyte–endothelial interactions in pial vessels and cortical postcapillary venules in an LPS-induced encephalitis model in mice and reduced the extent of neuroinflammation. 28 Selective activation of CB2R attenuated the firm leukocyte–endothelial interaction in the venous microvessels on the surface of the brain in experimental autoimmune encephalomyelitis in mice, thus reduced the hyperactive immune state. 29 These studies provide evidence for CB2R-mediated attenuation of the hyperinflammatory state within the CNS after a CNS injury by activating CB2R, which might explain the reduced systemic immunosuppression following experimental stroke observed in our study.
Treatment with HU308 resulted in a significant decrease in the brain infarct size in animals with CNS injury compared with untreated stroke animals, indicative of the neuroprotective role of CB2R agonists in CNS injury. Similar neuroprotective effects were reported by Zhang et al., where administration of O-1966 decreased cerebral infarction in mouse focal ischemia/reperfusion model. 20 Treatment with AM1241, a CB2R agonist, restrained neurodegeneration in the middle cerebral artery occlusion stroke model in rats. 19 Neuroprotective effects of CB2R agonists are primarily exerted through their anti-inflammatory properties. 30 Downregulation of inflammatory cytokines caused by CB2R agonists leads to a reduced invasion of inflammatory cells in the CNS after stroke.20,31 CB2R activation also prevents further disruption of the blood–brain barrier (BBB) caused by CNS injury. 20 However, other mechanisms may also be involved with CB2R agonist-associated neuroprotection. Suppression of microglial activation.32,33 and protection of CNS tissue from oxidative damage 34 are some of the other effects exerted by CB2R agonists contributing to neuroprotection.
The other therapeutic approach used in our study was to reverse CIDS by inhibiting CB2R activity using the CB2R inverse agonist, AM630. Administration of AM630 at the later phase of the injury restored the leukocyte activation and capillary perfusion in the small intestine of animals with CNS injury compared with stroke animals without the treatment. These observations complied with our previous study using the hypoxic–ischemic (HI) stroke model of mice, where CB2R inhibition caused similar changes in gut microcirculation. 21 The present study included some additional outcomes for the assessment of immune function, such as the plasma levels of soluble adhesion molecules, proinflammatory cytokines, and bone marrow neutrophil count. During inflammation, leukocytes are activated, followed by the induction of expression of adhesion molecules on the surfaces of endothelium and leukocytes, promoting leukocyte adhesion on the endothelial surface.35,36
Restoration of plasma levels of P-selectin, ICAM-1, and TNF-α in animals receiving AM630 therapy are additional indicators of restoration of the immune response. The reduction in bone marrow neutrophil count observed in animals with CNS injury in our study complies with the finding of a preclinical study, where CNS injury reduced NK cell counts in mice, making them vulnerable to septicemia. 37 A clinical study on ischemic stroke patients also reported lymphocytopenia to be most detectable among the NK cell subset. 38 Inhibition of CB2R with AM630 increased the number of neutrophils close to the level of sham animals, indicative of a potential reversal of the immune state back to homeostasis.
AM630 administration did not have any detrimental impact on the infarct size in our study. Peripheral inhibition of CB2R activity and enhancement of the immune function includes the risk of exacerbation of CNS damage due to dysfunctional BBB and penetration of peripheral immune cells into the brain through the damaged BBB. 39 However, our data in the present study with ET-1-induced CNS injury and the previous study with HI-induced CNS injury, 21 suggests that CB2R inhibition treatment may not exacerbate the magnitude of the brain injury during CIDS. However, we cannot exclude potential exacerbation of CNS damage at later time points or with different doses of the CB2R antagonist.
However, the combination of CB2R agonist and CB2R antagonist treatment could be another potential therapeutic approach for combating peripheral immunosuppression following a stroke, which also needs to be investigated.
Limitations
The experiments in the current study were conducted using young, healthy mice, whereas patients with stroke are likely to be older individuals with comorbidities.40,41 Sex variance also contributes to the pathophysiology and outcome after acute CNS injury.42,43 Studies suggest stroke as one of the top three causes of death in women. 44 There are also other lines of evidence suggesting variations between sex in microglia, dendritic cells, and neutrophils, potentially drawing more differences in sex-associated pathophysiological consequences of CNS injury.9,45 Also, there are known sex differences in the ECS of animals, with female rodents being more sensitive than males to the effects of cannabinoids. 46
Conclusion
Our work has demonstrated that CB2R modulation is a viable pharmacological target to combat the immunosuppression following CNS injury. Depending on the timing of the treatment, both activation and inhibition of CB2R can be potentially beneficial for an adequate immune response to a pathogen or an injury. While the obtained data are promising, further investigation of the effect of CB2R modulation on long-term neurological outcomes and survival is necessary before considering transferring any of these findings to the bedside.
Footnotes
Acknowledgment
The authors are thankful to Dr. Jean Marshall, Professor, Department of Microbiology and Immunology, Dalhousie University, Halifax, Nova Scotia, Canada for giving us access to Bio-Plex 200 System of her laboratory for the plasma cytokine analysis.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.