
Abstract
Introduction:
The endocannabinoid (eCB) system plays a key role in modulating brain development, including myelination processes. Recent studies link a common variant (C385A, rs324420) in the fatty acid amide hydrolase (FAAH) gene to higher circulating eCB levels, lower anxiety, and altered frontolimbic development. Frontolimbic pathways, which demonstrate a protracted maturational course across childhood and adolescence, are associated with anxiety, and are vulnerable to environmental stressors such as trauma exposure. Here, we examined the impact of trauma exposure, FAAH genotype, and anxiety on frontolimbic white matter microstructure in children.
Materials and Methods:
We leveraged baseline data from the Adolescent Brain Cognitive Development (ABCD) study (n=9969; mean±standard deviation age=9.92±0.62 years; 47.1% female). Saliva samples were used for genotyping, and caregivers reported on their child's anxiety symptoms and trauma exposure. Fractional anisotropy (FA), a nonspecific measure of white matter integrity, was estimated for frontolimbic tracts.
Results:
Thirty-six percent of youth experienced one or more potentially traumatic events according to DSM-5 Criterion A (64% controls), and 45% were FAAH A-allele carriers (55% noncarriers). Relative to controls, trauma-exposed youth demonstrated higher anxiety and higher FA of the left uncinate. The FAAH A-allele (vs. CC) was associated with lower FA in the left fornix and left parahippocampal cingulum, and there was an indirect effect of FAAH genotype on anxiety through FA of the left fornix. Moreover, genotype moderated the association between FA of the left cingulum and anxiety.
Conclusions:
Our findings demonstrate distinct effects of trauma exposure and the FAAH C385A variant on frontolimbic pathways and subsequent anxiety risk in preadolescent children. This line of work may provide important insights into neurodevelopmental mechanisms leading to anxiety risk, and potential targets for intervention.
Introduction
The endocannabinoid (eCB) system plays a key role in modulating brain development throughout the life span.1,2 During the prenatal period, the eCB system regulates synaptogenesis, neural progenitor differentiation, and axonal migration. 3 Postnatally, eCB levels change dynamically across childhood and adolescence, and regulate activity in neural circuits, including myelination. 4 In particular, the eCB system modulates the maturation of frontolimbic pathways that are rich in cannabinoid type 1 receptors, and regulate stress- and anxiety-related behaviors.2,5 Cross-species studies show that frontolimbic brain circuitry demonstrates a protracted maturational course across childhood and adolescence, and is sensitive to environmental stress.6–8 Early disruptions to frontolimbic development are thought to have long-term consequences for mental health. 9
One potent form of environmental stress is childhood trauma exposure, such as violence, abuse, or accidental injury. Trauma exposure is common among youth—affecting up to two-thirds of adolescents10,11—and is a chief risk factor for anxiety and other psychopathologies throughout the life span.12,13 Neuroimaging studies demonstrate that childhood trauma exposure is associated with aberrant maturation of frontolimbic circuits, including reduced gray and white matter volumes, and altered microstructural integrity of frontolimbic white matter pathways.14–17 Indeed, diffusion tensor imaging (DTI) studies have been used to measure microstructural integrity of white matter using fractional anisotropy (FA), which is based on the directional diffusion of water molecules in fiber pathways, with values ranging from 0 to 1, wherein 0 indicates isotropic (i.e., nondirectional) and 1 indicates anisotropic (i.e., highly directional) diffusion. 18
DTI studies frequently report altered FA of frontolimbic pathways in adults and youth with histories of childhood trauma exposure, and in youth with post-traumatic stress disorder (PTSD).5,14,18–23 Further, studies of childhood anxiety disorders demonstrate overlapping patterns of altered frontolimbic FA, 24 suggesting that altered maturation of frontolimbic pathways may be a common pathway linking childhood trauma with anxiety risk. For example, alterations in white matter integrity may influence functioning of neuronal circuits implicated in anxiety risk.
Emerging data indicate that the eCB system serves as “stress buffer,” and may protect against stress-related changes in frontolimbic circuitry and elevated anxiety risk.25,26 For example, a common variant (C385A) in the gene encoding fatty acid amide hydrolase (FAAH)—the enzyme that catabolizes eCBs—is linked to lower central binding to the FAAH enzyme, higher circulating eCB levels, elevated resting-state functional connectivity within frontolimbic circuitry, and lower anxiety.27,28 Similarly, a cross-species study by Mayo et al 29 found that the FAAH A-allele offered protection against stress-related decreases in eCB concentrations and increases in negative effect. Conversely, low eCB signaling is implicated in the pathophysiology of anxiety and other stress-related pathologies. 30 Behavioral or pharmacological interventions that boost eCB signaling are an area of active investigation for the prevention and treatment of anxiety and other mental disorders.31–34
Although recent studies suggest that eCB signaling may protect against the negative effects of stress, few studies have focused on developing populations. This is a critical gap given that frontolimbic pathways develop across childhood and adolescence, corresponding with a sharp uptick in the onset of anxiety and other stress-related pathologies. 35 Further, preclinical evidence suggests that the eCB system changes dynamically across development, 36 and that early life stress blunts eCB signaling in frontolimbic regions (e.g., the hippocampus), which persists until adulthood. 37
Given these developmental changes, it is difficult to extrapolate findings from adult studies to youth. Understanding of how the eCB system and stress shape frontolimbic circuitry during development may open up novel approaches for preventing psychopathology before it begins. To our knowledge, only two studies have explored the impact of FAAH genotype on anxiety and frontolimbic brain circuitry in children and/or adolescents. In an elegant cross-species study, Gee et al. linked the FAAH C385A variant to lower anxiety and higher FA of the uncincate fasciculus; however, these main effects only emerged after age 12. 38
A recent study by Sisk et al 39 explored effects of FAAH on anxiety and large-scale resting-state brain networks in the Adolescent Brain Cognitive Development (ABCD) study, an ongoing neuroimaging study of >10,000 youth across 21 sites in the United States. 40 In their analysis of 3,109 9- to 11-year-old children from the ABCD study, Sisk et al 39 identified main effects of FAAH on connectivity of a distributed brain network, which included frontolimbic regions. Consistent with the earlier study by Gee et al., there were no main effects of FAAH genotype on anxiety in children (<12 years old). There was, however, an interaction between FAAH and connectivity in predicting anxiety scores, such that youth with the CC genotype with connectivity that more closely resembled that of A-alleles showed lower anxiety. 39 However, no studies have tested whether FAAH protects against the effects of childhood trauma exposure on frontolimbic white matter pathways.
Here, we leveraged the ABCD data set to examine the impact of the FAAH C385A polymorphism, childhood trauma exposure, and anxiety and microstructure of frontolimbic white matter pathways in 9- to 11-year-old children. We tested for both main and interactive effects of FAAH genotype and trauma, and additionally tested for overlapping effects of anxiety symptoms on frontolimbic pathways.
We hypothesized that (1) compared with control youth, trauma-exposed children would demonstrate elevated anxiety symptoms and differences in white matter integrity in frontolimbic regions, following prior work summarized above. Given evidence that effects of FAAH on anxiety and FA of the uncincate do not emerge until age 12, 39 we did not expect to find main effects of FAAH on anxiety or frontolimbic FA in this preadolescent sample. We did, however, hypothesize that (2) the FAAH C385A (associated with elevated eCB concentrations) would protect against the effects of trauma exposure on anxiety and frontolimbic FA (i.e., trauma-by-FAAH interaction), and (3) frontolimbic FA would interact with FAAH to predict anxiety in youth, suggesting a possible brain phenotype linking anxiety with eCB signaling. 38
Of note, the ABCD study was not designed with in-depth measures of traumatic experiences and trauma-related symptoms. However, as one of the largest “population neuroscience” studies of brain development, the ABCD cohort provides a unique opportunity to examine the impact of trauma exposure and genetic variation in FAAH on the brain in a community-based sample.
Materials and Methods
Participants, trauma, and anxiety
This study leveraged data from 9969 children from the ABCD study (Table 1 and Supplementary Data).41,42 All participants provided written informed consent or oral assent and all study procedures were approved by the local and central institutional review boards. Caregivers reported on their child's trauma exposure using the PTSD Criterion A Traumatic Events Screener 43 from The Kiddie Schedule for Affective Disorders and Schizophrenia (K-SADS) for DSM-5, 44 and anxiety symptoms using the Child Behavior Checklist (CBCL). 48 We used the DSM-5–oriented anxiety subscale to measure continuous anxiety scores (standardized t-scores), and considered low- and high-anxiety groups based on cutoff scores (≥60). 45
Sample Demographics: Overall, by Trauma Group, and Genotype
Bold numbers indicate significant group differences (p < 0.05). Annual family income is reported in US Dollars.
Number of types of potentially traumatic events experienced according to DSM-5 Criterion A.
CBCL, Child Behavior Checklist; DTI, diffusion tensor imaging; FAAH, fatty acid amide hydrolase; SD, standard deviation.
FAAH genotype
We focused on the FAAH C385A polymorphism (rs324420). Following prior work,39,46,47 we characterized participants as A-alleles (i.e., AA or AC) or CC homozygotes. Importantly, FAAH genotype groups did not differ in trauma exposure (Table 1).
Frontolimbic white matter tracts
We focused on FA of the following ten frontolimbic white matter tracts (five tracts/hemisphere): fornix, uncinate fasciculus, cingulum bundle, parahippocampal cingulum, and inferior frontal occipital fasciculus (Fig. 1).
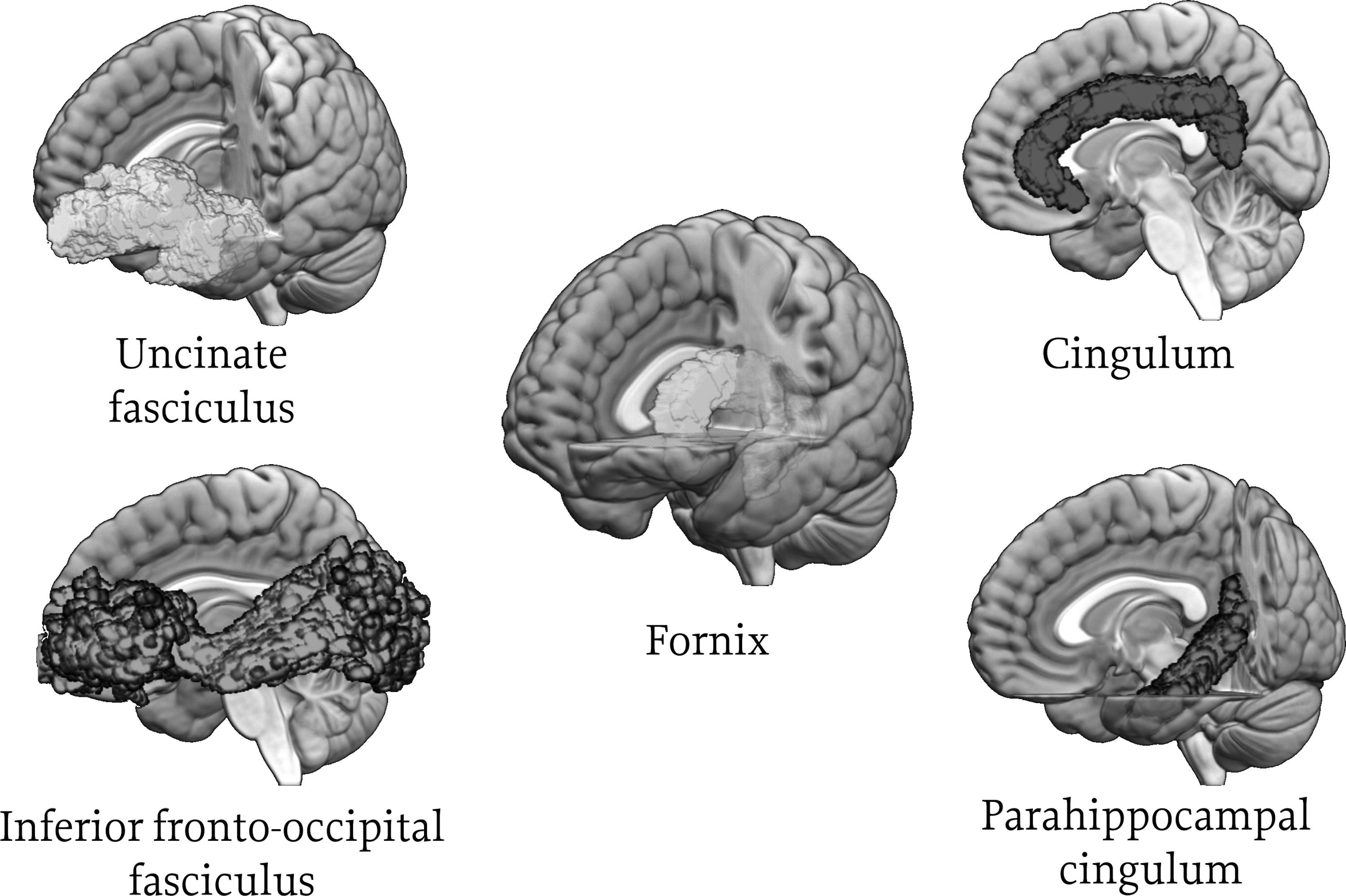
Frontolimbic white matter tracts of interest. Includes fornix (connecting the hippocampus and hypothalamus), uncinate fasciculus (connecting the inferior frontal lobe and anterior temporal lobe), cingulum bundle (connecting the cingulate gyrus and entorhinal cortex [cingulate portion]), parahippocampal cingulum (connecting the cingulate gyrus and entorhinal cortex [parahippocampal portion]), and inferior frontal occipital fasciculus (connecting the occipital lobe and frontal lobe).
Statistical analyses
Descriptive statistics were used to examine the prevalence of trauma exposure and distribution in FAAH genotype across the sample. We tested for effects of trauma and FAAH on demographic variables and anxiety symptoms. Variables that showed significant associations with trauma and/or FAAH genotype were included as covariates in subsequent analyses. Regression was used to test for main effects of FAAH genotype, trauma-by-FAAH, and anxiety symptoms on FA for each white matter tract, controlling for multiple comparisons. Following a recent article outlining best practices for control of covariates in the ABCD data set, 48 we used a tiered approach to covariates. We also conducted follow-up analyses to test for differences among the three gene groups (AA, AC, and CC). Mediation and moderation analyses were conducted for white matter pathways showing associations with both anxiety and FAAH genotype. See Supplementary Data for further details.
Results
Trauma exposure and FAAH
More than one-third (35.7%) of youth were exposed to one or more types of potentially traumatic events according to DSM-5 Criterion A. Trauma and control (no-trauma) groups differed in race/ethnicity, maternal education, and annual family income (Table 1). Therefore, these variables were included as “tier 2” covariates in subsequent regression models. Forty-five percent of youth were A-allele carriers; 55% were noncarriers.
Anxiety
Trauma-exposed youth showed higher anxiety as compared with control youth (Table 1). Similarly, trauma-exposed youth were more likely to exceed the cutoff for anxiety (Table 1). Therefore, anxiety was included as a “tier 3” covariate in subsequent regression analyses. There was no effect of FAAH (Table 1) or trauma-by-FAAH interaction on anxiety symptoms, nor proportion of children with elevated anxiety (p=0.39). There were also no differences between gene groups (AA, AC, and CC) in anxiety symptoms (p's>0.5).
Frontolimbic white matter
Main effects of trauma
With the inclusion of tier 1 covariates (age, sex at birth), trauma was positively associated with FA of the right cingulum bundle [overall model: F(3, 9452)=53.34, p<0.001; β=0.002, t=2.13, p=0.033], the left uncinate fasciculus [F(3, 9455)=10.52, p<0.001; β=0.002, t=3.1, p=0.002], and the right uncinate fasciculus [F(3, 9454)=10.19, p<0.001; β=0.001, t=2.05, p=0.04]. With the addition of tier 2 covariates (race, household income, maternal education), trauma exposure was positively associated with FA of the left fornix [overall model: F(6, 8633)=19.85, p<0.001; β=0.001, t=2.03, p=0.042], right fornix [F(6, 8633)=15.26, p<0.001; β=0.002, t=2.25, p=0.025], right parahippocampal cingulum [F(6, 8633)=6.13, p<0.001; β=0.002, t=2.36, p=0.018], and left uncinate [F(6, 8633)=6.06, p<0.001; β=0.002, t=2.87, p=0.004]. The left uncinate was the only tract that passed False Discovery Rate (FDR) correction and that remained significant when adding the tier 3 covariate [i.e., anxiety; overall model: F(7, 8631)=5.38, p<0.001; β=0.002, t=2.87, p=0.004; see Fig. 2a]. Trauma exposure was not associated with FA of other pathways.
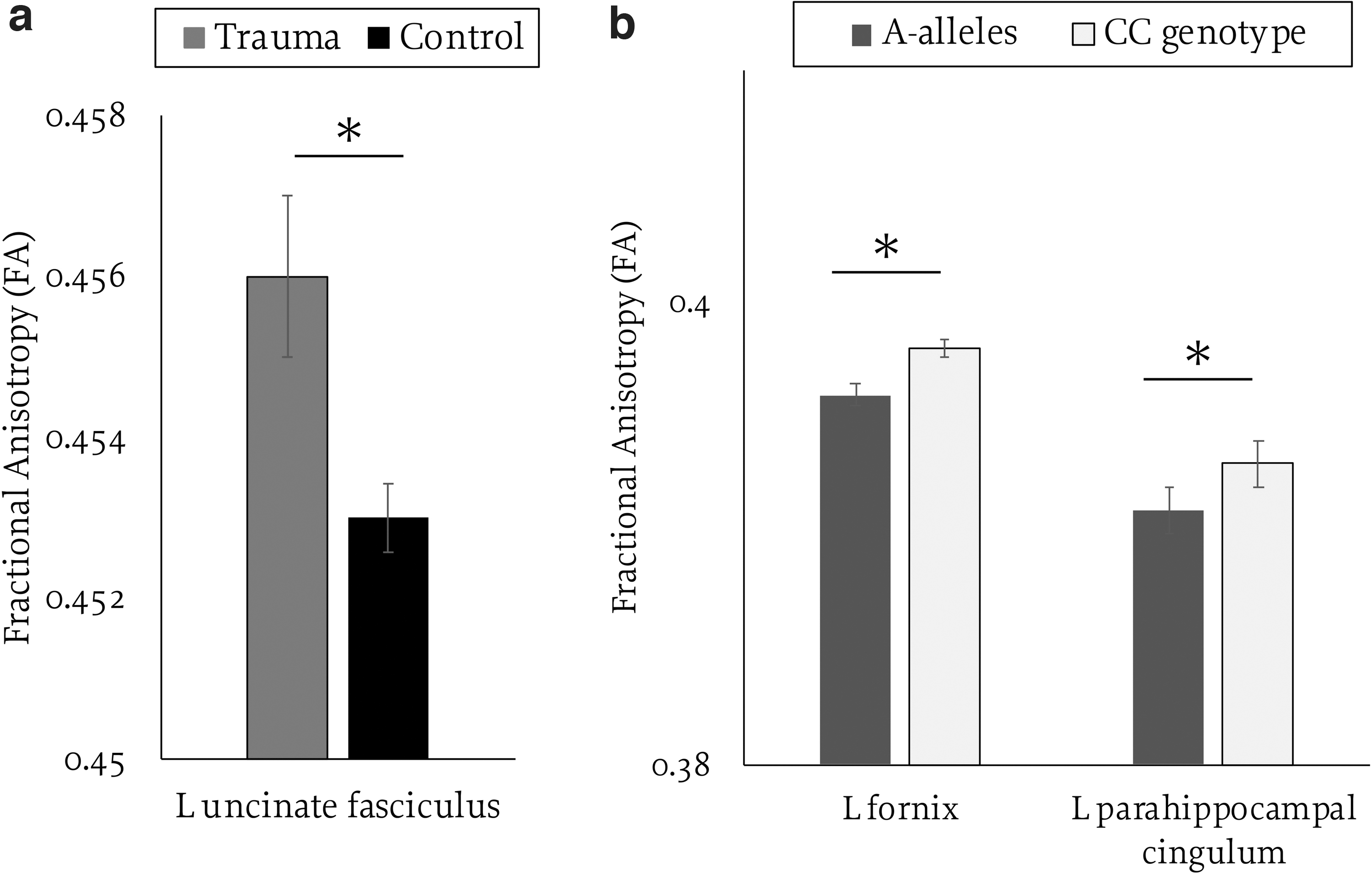
Main effects of trauma exposure
Main effects of FAAH
With the inclusion of tier 1 covariates, the FAAH C385A variant was associated with lower FA in the left fornix [overall model: F(3, 9455)=27.43, p<0.001; β=0.002, t=3.5, p<0.001], right fornix [F(3, 9455)=26.04, p<0.001; β=0.002, t=2.5, p=0.01], right [F(3, 9455)=9.944, p<0.001; β=0.002, t=2.37, p=0.02], and left parahippocampal cingulum [F(3, 9453)=16.41, p<0.001; β=0.003, t=3.59, p<0.001]. With the addition of tier 2 covariates, the FAAH C385A variant was associated with lower FA of the left fornix [F(6, 8633)=20.12, p<0.001; β=0.001, t=2.39, p=0.017] and left parahippocampal cingulum [F(6, 8631)=13.25, p<0.001; β=0.002, t=2.64, p=0.008; Fig. 2b].
The left fornix [F(7, 8631)=18.36, p<0.001; β=0.001, t=2.37, p=0.018] and left parahippocampal cingulum [overall model: F(7, 8629)=11.55, p<0.001; β=0.002, t=2.67, p=0.008] both remained significant when adding the tier 3 covariate (Fig. 2b) but neither passed FDR correction. Follow-up analyses comparing the three gene groups showed significant differences in left fornix FA only for AC versus CC gene groups [F(7, 8809)=18.2, p<0.001; β=0.027, t=2.14, p=0.017]. No other gene groups were significant with the addition of tier 3 covariates. FAAH genotype was not associated with FA in other frontolimbic pathways.
Trauma-by-FAAH interactions
With the inclusion of tier 2 covariates, there were no significant trauma-by-FAAH genotype interactions on frontolimbic FA (F change p's>0.9).
Main effects of anxiety symptoms
With the inclusion of tier 1 covariates, anxiety was not associated with FA in any of the tested frontolimbic pathways (p's>0.1). With the addition of tier 2 covariates, anxiety was positively associated with FA in the left fornix [F(6, 8631)=20.47, p<0.001, β=0.03, t=2.78, p=0.005; see Fig. 3a]. Controlling for effects of trauma exposure and FAAH genotype (tier 3), anxiety remained a significant positive predictor of FA in the left fornix [F(8, 8631)=16.42, p<0.001, β=0.028, t=2.56, p=0.01], suggesting that results are robust to trauma and FAAH. Results for the left fornix passed FDR correction and remained significant when considering anxiety as a group variable (low, high) rather than a continuous variable (F(8, 8631)=16.18, p<0.001, β=0.023, t=2.16, p=0.031).
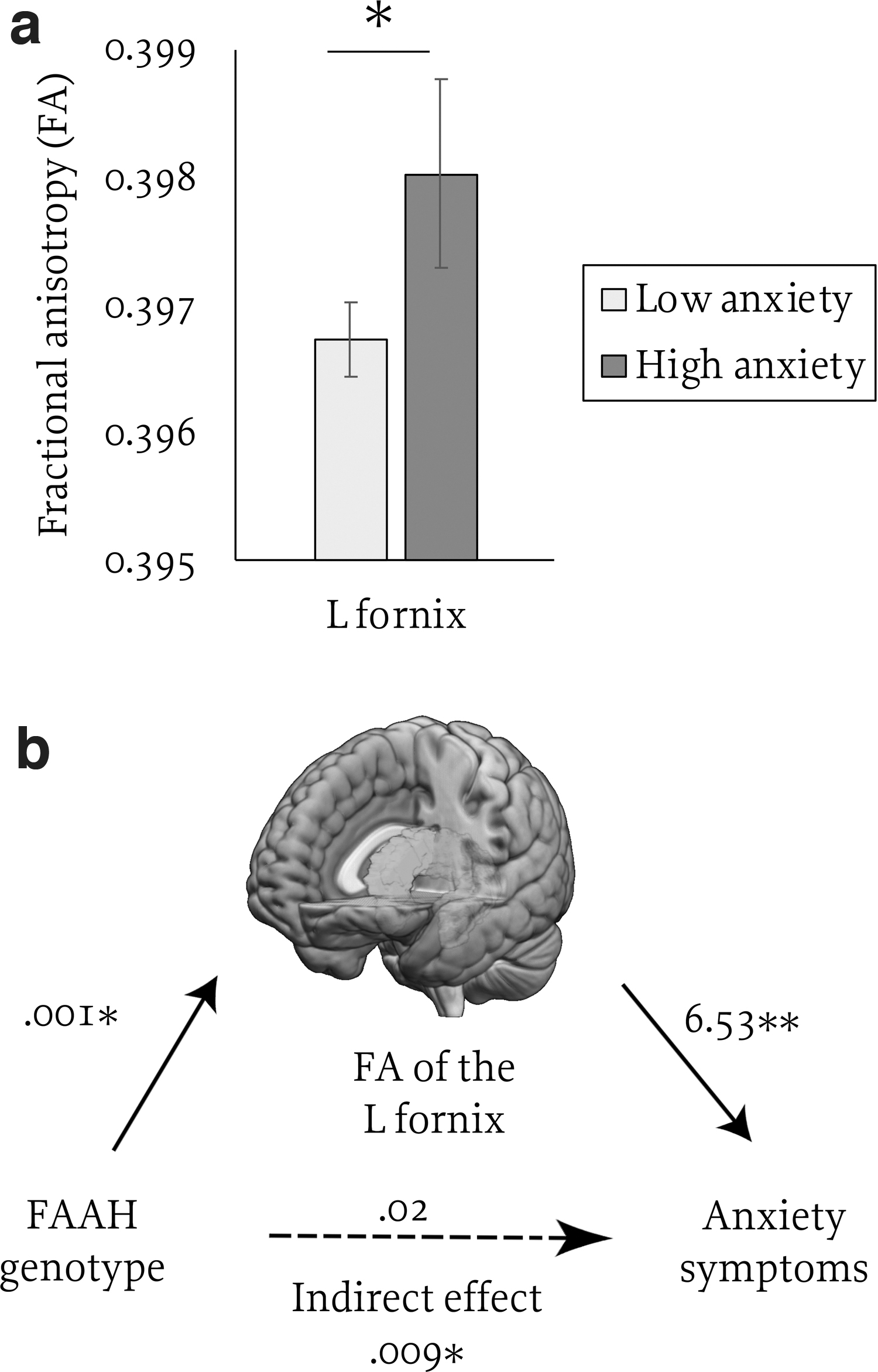
Effects of anxiety symptoms
Mediation
We explored indirect effects of FAAH on anxiety through variation in FA. We focused on FA of the left fornix given the observed main effects of both anxiety and FAAH on this pathway. With the inclusion of tier 2 variables, there were significant indirect effects of FAAH genotype on anxiety through FA of the left fornix (β=0.0093, boot standard error [SE]=0.0054, lower level confidence interval [LLCI]=0.0008, upper level confidence interval [ULCI]=0.0218; partially standardized β=0.0015, boot SE=0.0009, LLCI=0.0001, ULCI=0.003; see Fig. 3b).
Reversal of the model (FAAH predicting fornix FA through anxiety) was not significant (LLCI=−0.0003, ULCI=0.00005) indicating that fornix FA mediated the associated between FAAH and anxiety but not the reverse. Results remained consistent when considering anxiety as a group (low, high) rather than a continuous variable (LLCI=0.0009, ULCI=0.022). The indirect effect for FA of left parahippocampal cingulum was not significant (LLCI=−0.0036, ULCI=0.0135).
Moderation
Given that a prior study demonstrated interactive effects of FAAH and functional connectivity in predicting anxiety in the ABCD sample, 39 we explored whether similar interactive effects exist for structural connectivity. With the inclusion of tier 2 variables, there was a significant interaction between FAAH genotype and left cingulum FA in predicting anxiety, [F(8, 8623)=1.41, p<0.001, b=0.95, t=2.82, p=0.0048; Fig. 4]. This effect was driven by a positive association between FA and anxiety in youth with the CC genotype (b=5.33, t=2.83, p=0.0047), but no association between FA and anxiety among A-alleles (b=−2.62, t=−1.2, p=0.21). These results passed FDR correction and remained significant when considering anxiety as a group rather than a continuous variable (p=0.02). No other pathways showed significant interactive effects with FAAH.
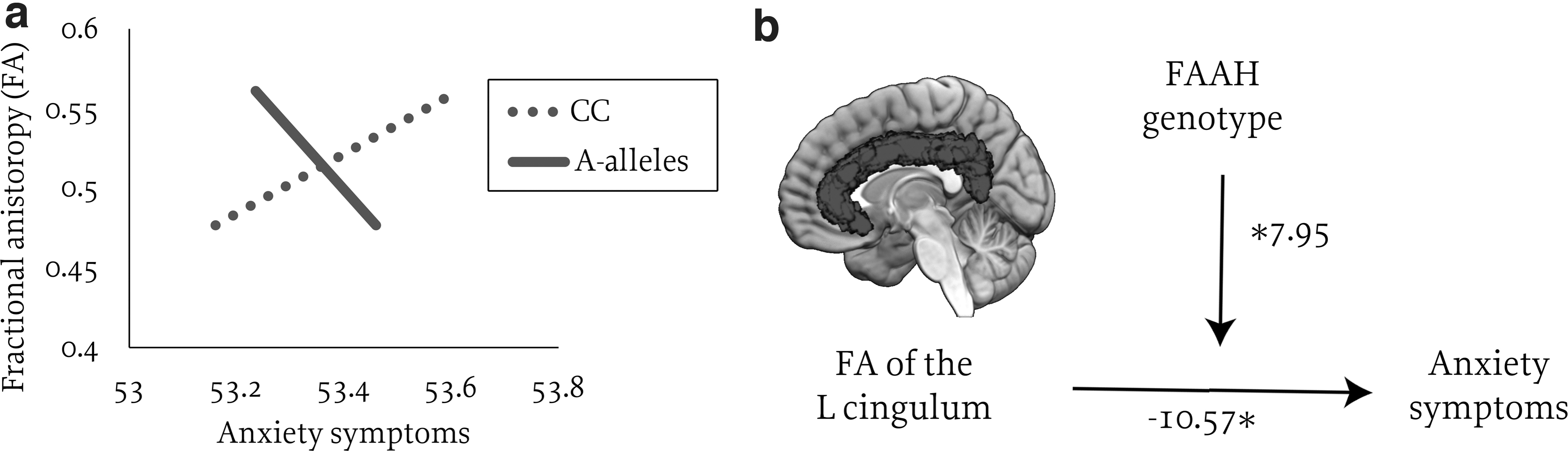
Interactive effects of FAAH genotype and FA of the left cingulum on anxiety symptoms.
Discussion
This study tested the hypothesis that the FAAH C385A variant—associated with higher eCB signaling—buffers the effects of childhood trauma on anxiety and frontolimbic pathways in 9- to 11-year-old children. We did not find evidence for interactive effects of trauma exposure and FAAH genotype on childhood anxiety symptoms nor frontolimbic white matter microstructure. We did, however, observe distinct effects of trauma exposure and FAAH genotype on anxiety and frontolimbic white matter. In particular, trauma-exposed youth (vs. controls) showed elevated anxiety and higher FA of the left uncincate. The FAAH A-allele (vs. CC genotype) was associated with lower FA in the left fornix and left parahippocampal cingulum, and there was an indirect effect of FAAH genotype on anxiety through FA of the left fornix.
Moreover, FAAH genotype moderated the association between FA of the left cingulum bundle and anxiety, suggesting a possible brain phenotype of anxiety risk, which parallels the reported patterns observed in resting-state functional connectivity. 39 Together, these findings demonstrate that both trauma and the FAAH C385A variant impact frontolimbic circuitry and anxiety risk during preadolescence. In particular, alterations in white matter microstructure may influence functioning in neuronal circuits that govern stress and anxiety regulation. Results also add to prior studies, suggesting that the reported buffering effects of eCBs against stress-related effects on frontolimbic circuitry and anxiety risk emerge later, during adolescence (ages 12+).
We observed elevated anxiety in children with histories of trauma exposure relative to controls, which is consistent with the extant literature.11,13 Neuroimaging studies also frequently report altered white matter microstructure of the uncincate after childhood trauma; however, the direction of these effects are inconsistent. Indeed, several studies report lower FA in trauma-exposed adolescents and adults with histories of childhood trauma as compared with controls.20,49 Here, we observed higher FA in the uncincate in trauma-exposed children (vs. controls), which is consistent with a prior study in children in the same age range. 5
Elevated FA of the uncincate in trauma-exposed youth may reflect enhancements within medial temporal-prefrontal pathways, which may contribute to altered hippocampal memory functioning. 50 The observed pattern is broadly consistent with prior studies reporting accelerated maturation of amygdala-prefrontal circuits following early life adversity, which may suggest an adaptive response. 51 However, accelerated developmental may have consequences for later in life. 52 Prior studies also show a shift in amygdala-prefrontal circuits around age 12, and that altered patterns of white matter neurodevelopment across childhood and adolescence may reflect psychiatric risk.19,38 Therefore, microstructural alterations described in this article may reflect a brain phenotype that is specific to preadolescence. 38 Future waves of ABCD will provide better insights into whether the observed patterns are time limited or persistent.
We observed lower FA in A-alleles (vs. CC youth) in the left fornix and left parahippocampal cingulum, tracts that connect the hippocampus to the hypothalamus and medial prefrontal regions, respectively. Prior studies demonstrate that hippocampal-prefrontal circuits are critical for stress responding and emotion regulation, 53 including dampening amygdala output, fear extinction, and contextual processing.54,55
Further, there is a high density of cannabinoid type 1 receptors in both the hippocampus and prefrontal cortex, 56 and a prior study in adults links genetic variation in eCB signaling to altered hippocampal activity during fear extinction. 47 Our findings demonstrate that FAAH genotype modulates frontolimbic pathways during preadolescence—earlier than previously demonstrated. We observed effects in the fornix and parahippocampal cingulum, which are implicated in fear, anxiety, and episodic memory.57,58 We did not observe main effects of FAAH on FA of the uncincate, which is consistent with prior studies showing effects of FAAH on this pathway emerge after age 12.
We observed indirect (but not direct) effects of FAAH on anxiety in this preadolescent sample, which is consistent with prior studies showing that these effects emerge during adolescence.38,39 Interestingly, we found evidence for an indirect association between FAAH and anxiety through white matter integrity of the fornix, which has been previously associated with anxiety risk in adults. 59 Similar to a recent study of functional connectivity in the ABCD sample, 39 we found that FAAH interacted with structural connectivity of the cingulum to predict anxiety. In both studies, the effects of brain connectivity on anxiety were only apparent with the CC group, suggesting that the A-allele (associated with elevated eCB signaling) may confer some benefit against anxiety. Given that altered FA in the cingulum is frequently reported in adults and youths with anxiety, 57 our findings suggest that FAAH modulates frontolimbic pathways involved in anxiety risk.
Strengths and limitations of this study warrant mention. Strengths of this study include the use of a large, nationwide study, and inclusion of both trauma and FAAH on anxiety and frontolimbic development. One limitation is that the proportion of youth with exposure to one or more types of potentially traumatic events was relatively low, which may limit comparison with other studies of trauma. For example, our prior community-based studies of lower income urban areas find higher rates of exposure among children of a similar age (e.g., 45%). 60
The relatively low rate of trauma in the baseline ABCD sample may be related to the high representation of youth from white and higher income families, or related to the age. Indeed, rates of trauma exposure have been shown to dramatically increase around age 12. 11 Therefore, future studies should aim to examine the effects of age-of-onset, trauma severity, frequency, or trauma types on frontolimbic development. Also, the mediation analyses are cross-sectional and should be considered exploratory. Future studies incorporating longitudinal releases of ABCD data should help distinguish between mediation and spuriousness (i.e., caused by another covariate) associations among trauma, anxiety, and FA. 48
Further, we relied on DTI, which is a nonspecific measure of white matter integrity. FA is influenced by a number of factors, including axon density, diameter, damage, myelination, and permeability, and by crossing fibers, fiber orientation, or number.61,62 Therefore, future studies should employ different approaches to better characterize the observed effects. In addition, although the FAAH C385A variant has consistently associated with higher circulating levels of the eCB anandamide, there is variation in eCB levels within genotype groups. Future studies measuring circulating concentrations of eCBs in youth, particularly preadolescent youth, are needed.
Conclusions
This study builds on our knowledge of the neural correlates of anxiety risk in youth. In particular, our findings demonstrate distinct effects of trauma exposure and the FAAH C385A variant on white matter microstructure and subsequent anxiety risk in preadolescent children. This line of work should provide important insights into potential neurodevelopmental mechanisms leading to anxiety risk, and potential targets for intervention.
Footnotes
Acknowledgments
Authors' Contributions
H.A.M. contributed to conceptualization (lead); writing—original draft (lead); formal analysis (lead); writing—review and editing. J.E. and S.D. contributed to writing—review and editing. C.A.R. performed writing—review and editing; conceptualization (supporting). All authors approved the final version of the article.
Author Disclosure Statement
C.A.R. disclosed consulting fees from Lundbeck. All other authors reported no biomedical financial or potential conflicts of interest.
Funding Information
H.A.M. is supported by K01MH119241 and C.A.R. is supported by R33MH111935. Data used in the preparation of this article were obtained from the ABCD study (https://abcdstudy.org), held in the NDA. This is a multisite, longitudinal study designed to recruit >10,000 children age 9–10 and follow them over 10 years into early adulthood. The ABCD study is supported by the National Institutes of Health and additional federal partners under award numbers U01DA041022, U01DA041028, U01DA041048, U01DA041089, U01DA041106, U01DA041117, U01DA041120, U01DA041134, U01DA041148, U01DA041156, U01DA041174, U24DA041123, U24DA041147, U01DA041093, and U01DA041025. A full list of supporters is available at https://abcdstudy.org/nih-collaborators A listing of participating sites, and a complete listing of the study investigators can be found at ![]() investigators.html ABCD consortium investigators designed and implemented the study and/or provided data, but did not participate in analysis or writing of this report. This article reflects the views of the authors, and may not reflect the opinions or views of the NIH or ABCD consortium investigators. The ABCD data repository grows and changes over time.
investigators.html ABCD consortium investigators designed and implemented the study and/or provided data, but did not participate in analysis or writing of this report. This article reflects the views of the authors, and may not reflect the opinions or views of the NIH or ABCD consortium investigators. The ABCD data repository grows and changes over time.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.