
Abstract
Introduction:
CD19-chimeric antigen receptor (CAR) T cell therapy is a promising immunotherapy for cancer treatment that has shown remarkable clinical responses, leading to approval by the FDA for relapsed and refractory B cell hematological malignancy treatment. Cannabidiol (CBD) is a nonpsychoactive cannabinoid compound that has been utilized as a palliative treatment in cancer patients due to its immunosuppressive properties. Currently, studies on using CBD during immunotherapy have gained increasing attention. However, the possible interaction between CBD and CAR T cell therapy has not been studied. Therefore, in this study, we aimed to examine the direct effects of CBD on CD19-CAR T cell function against hematologic malignancies.
Materials and Methods:
The cytotoxic effect of CBD was determined by a cell proliferation reagent water-soluble tatrazolium salt (WST-1) assay. CAR T cells were generated by retroviral transduction and treated with CBD at a nontoxic dose. The effect of CBD on immune characteristics, including transgene expression, T cell subset, and memory phenotype, was analyzed by flow cytometry. Proliferation, apoptosis, and cell cycle distribution were analyzed with standard methods. The effect on cytotoxic function was evaluated using degranulation assays, and antitumor activity was evaluated using flow cytometry.
Results:
The half-maximum inhibitory concentration (IC50) of CBD on NALM6, Raji, and T cells ranged from 16 to 22 μM. The maximum nontoxic dose of CBD that maintained cell viability at ∼100% was 8 μM. For the generation of CD19-CAR T cells, primary T cells were activated and transduced with a retroviral vector encoding CD19-CAR. CBD did not alter the surface expression or immune characteristics, including the T cell subset and memory phenotype, of CD19-CAR T cells. However, CBD suppressed CD19-CAR T cell proliferation by inducing apoptosis, as evidenced by an increase in the proportion of cells in the Sub-G1 phase in cell cycle arrest. However, the antitumor activity and cytokine secretion of CD19-CAR T cells were not altered by exposure to CBD in this study.
Conclusions:
In this study, a nontoxic dose of CBD affected CD19-CAR T cell proliferation but not its immune characteristics or cytotoxic function.
Introduction
Chimeric antigen receptor (CAR) T cell therapy is an immunotherapy that involves engineering lymphocytes to express CAR capable of binding to surface target molecules on cancer cells. 1 CARs do not require human leukocyte antigen presentation for their activation, 2 making them unaffected by the immune escape mechanism of tumor cells. 3 CAR T cells use a single chain variable fragment (scFV) to recognize and bind to tumor cell antigens. Upon CAR engagement, activated CAR T cells proliferate and secrete soluble mediators and activate the Fas/FasL pathway, leading to tumor lysis. 4 CAR T cells targeting CD19 have shown remarkable results in the treatment of relapsed and refractory acute lymphoblastic leukemia (ALL), 5 resulting in FDA approval for the treatment of relapsed and refractory B cell hematological malignancies.5–9
The therapeutic potential of cannabis in clinical treatment has been used for relieving cancer symptoms and treatment side effects.10,11 Cannabis sativa L. contains major phytocannabinoid compounds, including Δ9-tetrahydrocannabinol and cannabidiol (CBD). CBD is a nonpsychoactive cannabinoid that has low affinity for both cannabinoid receptors type 1 and 212,13 and has been classified as a negative allosteric modulator.14,15 CBD acts as an antagonist of G-protein-coupled receptor 55 (GPR55) 16 and an agonist of transient receptor potential vanilloid type 1 (TRPV1)17,18 and peroxisome proliferator-activated receptor gamma receptors. 19 In addition, CBD acts as an inhibitor of fatty acid amide hydrolase action.18,20 By moderating the activity of these receptors, CBD exhibits multiple therapeutic effects, such as anti-inflammatory and immunosuppressive properties.21–25
Studies have reported a correlation between cannabis consumption and decreased time to tumor progression (TTP) and overall survival in cancer patients treated with immune checkpoint inhibitors. 26 However, cannabis use was found to reduce therapy-related immune-related adverse events.26,27 In another retrospective observational study, cannabis was a factor that reduced the response rate in patients treated with nivolumab. 27 Despite research on the effects of CBD on cellular and cytokine mechanisms of immune cells, few studies have investigated its combination with immunotherapy, particularly CAR T cell therapy. This study aims to investigate the pharmacological effects of CBD on the function of CD19-CAR T cells against ALL and B cell lymphoma.
Materials and Methods
Cannabidiol
CBD was prepared from the supercritical carbon dioxide extract of C. sativa L. grown at the Center of Plant-Produced Biopharmaceutical for Dentistry, Faculty of Dentistry, Chulalongkorn University (Bangkok, Thailand). 28 The CBD isolate was recovered with >95% purity. The identification of CBD was performed by comparison to the CBD reference standard (THC Pharm GmbH, Germany). For the biological assay, CBD was further dissolved in dimethyl sulfoxide (DMSO; endotoxin free with ≥99.9% purity) (Sigma-Aldrich, Roche, Indianapolis, IN, USA) and stored at −20°C. CBD was diluted to the desired concentration with phosphate buffered saline (Gibco BRL Life Technologies, Inc.) for subsequent experiments.
Primary cells and cell lines
NALM6 cells, Raji cells, and 293T human kidney embryonic cells were cultured in RPMI 1640 or DMEM supplemented with 10% heat-inactivated fetal bovine serum (FBS), 1% GlutaMAX™, and 1% Penstrep (Gibco BRL Life Technologies, Inc.), respectively. Peripheral blood mononuclear cells (PBMCs) were obtained from healthy volunteers after obtaining signed informed consent that was approved by the institutional review board of the Faculty of Medicine, Chulalongkorn University (IRB No. 633/64). All experiments were performed following guidelines and regulations.
Cytotoxicity of CBD
Primary T cells and CD19+ tumor cells, including NALM6 and Raji cells, were treated with CBD at varying concentrations of 0–30 μM for 24 h. Cell viability was evaluated using a water-soluble tatrazolium salt (WST-1) assay (Sigma–Aldrich, Roche) with vehicle containing 0.1% DMSO as a control.
Preparation of retroviral vector
The CD19-CAR construct is composed of CD19-specific scFV (FMC63), an IgG2-derived CH3 hinge, and the intracellular signaling domain of CD28 and CD3ζ. To produce a retroviral vector, 293T cells were transfected with a MomLV gag-pol plasmid, RD114 envelope plasmid, and plasmid encoding the CD19-CAR construct using GeneJuice® as the DNA transfection reagent (Novagen, Billerica, MA). Retroviral supernatants were harvested, filtered, and snap frozen at 48 and 72 h after transfection. 29
Generation of CD19-CAR T cells
CD19-CAR T cells were generated as previously described. 29 Briefly, PBMCs were isolated and activated with T Cell TransAct™ (Miltenyi Biotec, Auburn, CA, USA) for 72 h. Activated cells were expanded in TexMACS medium (Miltenyi Biotec, Auburn, CA, USA) supplemented with 5% heat-inactivated FBS.
To generate CD19-CAR T cells, RetroNectin (FN CH-296; Takara Shuzo, Otsu, Japan)-coated plates were loaded with retroviral supernatant and centrifuged at 2000 g for 90 min. Activated T cells were added to the plate and incubated for 4 days. Activated T cells added to the precoated well without viral supernatant were included as nontransduced T cells (NT).
Flow cytometry
Biotinylated human CD19 protein and Phycoerythrin (PE)-biotin antibody (Miltenyi Biotec, Bergisch Gladbach, Germany) were used for CD19-CAR detection. CD3-PerCP (OKT3), CD4-APC (OKT4), CD8-PE (SK1), CD3-PerCP (OKT3), CD62L-FITC (DREG-56), CD45RO-APC (UCHL1), monensin, CD107-PerCP (H4A3), CD3-APC (OKT3), CD19-APC (HIB19), CD3-FITC (OKT3) (BioLegend, San Jose, CA), and 7-aminoactinomycin D (7-AAD) (BD Biosciences) were used to detect immune characteristics and cytotoxic function. Flow cytometry was performed using a BD Accuri™ C6 Plus Flow Cytometer (BD Biosciences).
Apoptosis assay and cell cycle analysis
CD19-CAR T cells were treated with CBD (2–8 μM) for 96 h. Apoptosis assays were performed by staining with an Annexin V-Propidium Iodide (PI) Apoptosis Kit (Sigma-Aldrich, Roche). The DNA content of the cell cycle was determined by staining with the BD Cycletest™ Plus DNA Reagent Kit (BD Biosciences) according to the manufacturer's instructions. The percentage of apoptotic cells and the cell cycle were analyzed by flow cytometry.
Degranulation assay
CD19-CAR T cells were stained with monensin and anti-human CD107a and cocultured with target cells at an effector/target (E:T) ratio of 1:1 in the presence of CBD (2–8 μM) for 4 h. Cells were harvested, stained with anti-human CD3, and analyzed for CD107a expression.
Coculture experiments
CD19-CAR T cells were cocultured with target cells at an E:T ratio of 1:2 in the presence of CBD (2–8 μM) for 24 h. Cells were harvested and quantified by flow cytometry using 7-AAD, anti-human CD3, and CD19. The percentage inhibition was calculated using the following formula:
Cytokine detection
CD19-CAR T cells were cocultured with target cells at an E:T ratio of 1:1 in complete RPMI 1640 medium in the presence or absence of CBD (2–8 μM) for 24 h. Culture supernatants were harvested, and the cytokine levels of tumor necrosis factor-α (TNF-α), interferon-γ (IFN-γ), interleukin-2 (IL-2), and granulocyte-macrophage colony-stimulating factor (GM-CSF) were measured by ELISA Kits according to manufacturer's instruction (88-7346, 88-7316, and 88-7025; Invitrogen) (DGM00; R&D Systems).
Statistics
All results are reported as the mean±SEM. The statistical analysis was performed using GraphPad Prism software (Prism, version 8.4.0; La Jolla, CA). One-way ANOVA was used to determine statistical significance among groups. p<0.05 was considered statistically significant.
Results
Cytotoxicity of CBD
To study the effect of CBD on the function of CD19-CAR T cells, we first determined the cytotoxic dose of CBD in CD19+ tumor cells and primary T cells in vitro. Cells were treated with CBD (0–30 μM) for 24 h. CBD dose-dependently decreased the percent viability of NALM6, Raji, and activated T cells (Fig. 1A–1C). The maximum nontoxic dose of CBD was 8 μM. The half-maximum inhibitory concentration (IC50) of CBD in NALM6, Raji, and T cells was 21.92±0.78, 19.04±0.56, and 15.61±0.62 μM, respectively (Fig. 1D). The nontoxic doses of 2–8 μM were then chosen for the subsequent experiments.
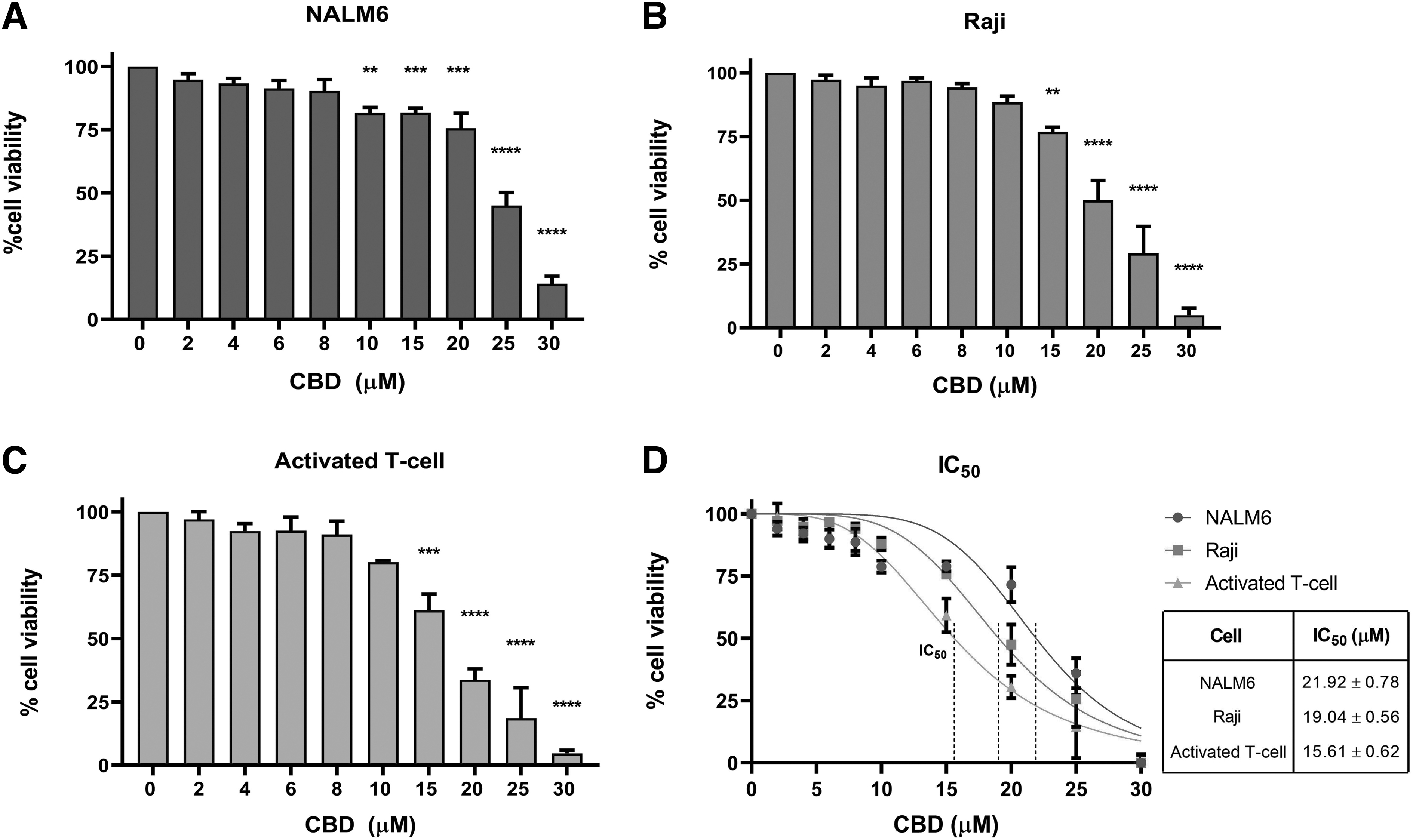
Cytotoxicity of CBD in hematologic tumor cell lines and primary cells.
Generation of CD19-CAR T cells for B cell leukemia and lymphoma
First, we generated CD19-CAR T cells by retroviral transduction, as illustrated in Figure 2A. NT were generated as a control. CD19-CAR constructs consisted of an anti-CD19 scFV sequence linked with a hinge, followed by the intracellular domain of costimulatory CD28 fused with the CD3ζ chain, as shown in Figure 2B. CAR T cells were expanded in the presence of IL-7 and IL-15, and transgene expression was analyzed on days 4 and 11 after transduction. CD19-CAR T cells exhibited transgene expression of 95.4±1.32% and 94.6±1.58% on days 4 and 11 after transduction, respectively (Fig. 2C, D).
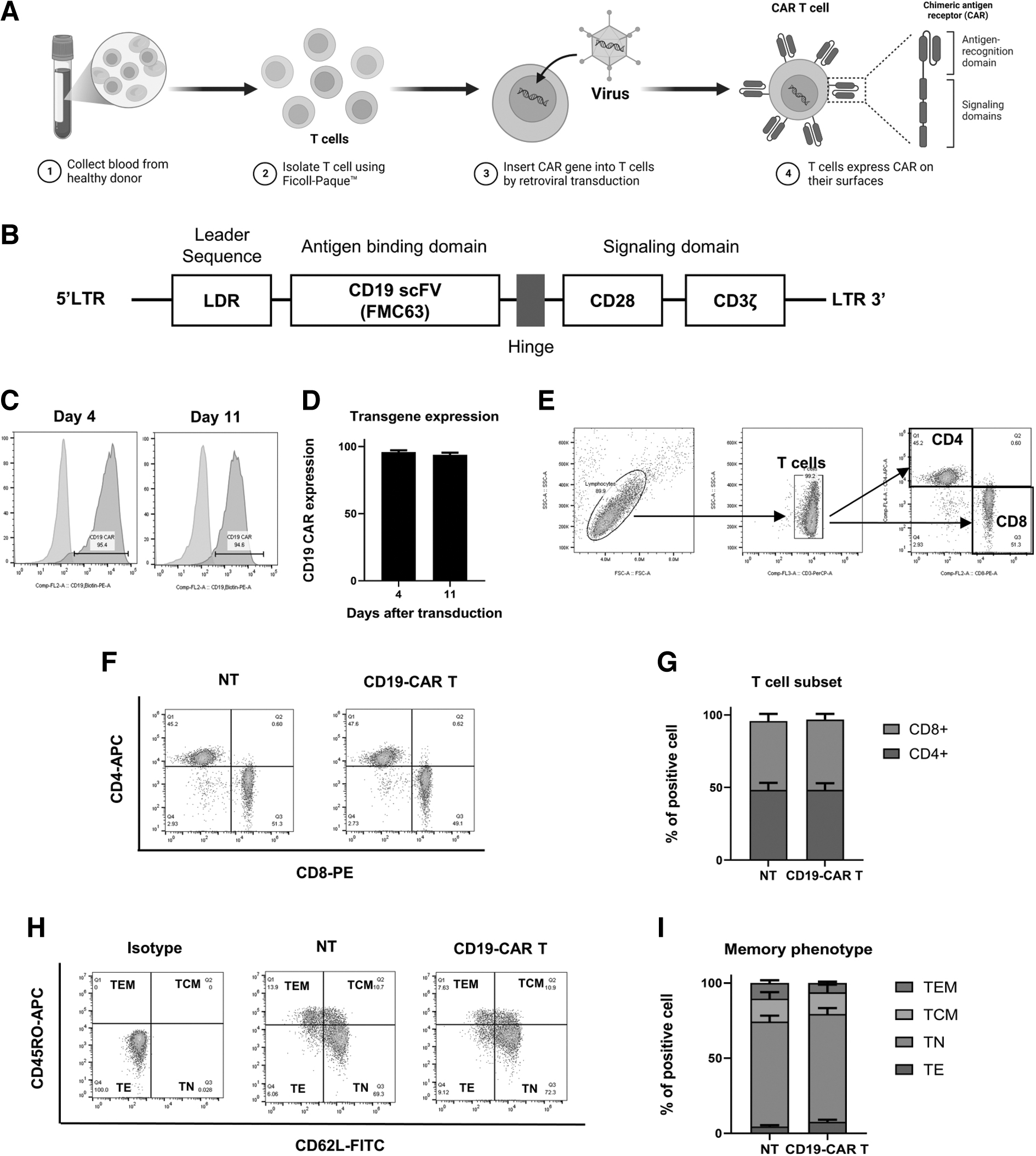
Generation of CD19-CAR T cell using retroviral transduction.
As it has been shown that therapeutic efficacy is related to the memory phenotype of the manufactured product, CD19-CAR T cells were harvested to analyze the T cell subset and memory phenotype on day 11 after transduction. Figure 2E illustrates the gating strategy of the T cell subset, including CD4 and CD8 T cells. The results revealed that the ratio between CD4 and CD8 T cells was ∼1:1 in both NT and CD19-CAR T cells (Fig. 2F, G). As shown in Figure 2H and I, NT and CD19-CAR T cells contained ∼70% naive T cells, followed by central memory, effector memory, and effector subpopulations.
CBD did not alter the immune characteristics of CAR T cells
To investigate the effect of CBD on immune characteristics, we treated CD19-CAR T cells with 2, 4, and 8 μM CBD for 7 days. As shown in Figure 3A and B, CBD did not alter CAR transgene expression. Analysis of T cell subsets also revealed that the ratio between CD4 and CD8 T cells was maintained at 1:1 in all groups (Fig. 3C). Moreover, CBD treatment did not affect the memory phenotype of CD19-CAR T cells. These results suggest that CBD at 2, 4, and 8 μM did not alter transgene expression or immune characteristics, including the T cell subset and memory phenotypes of CD19-CAR T cells.
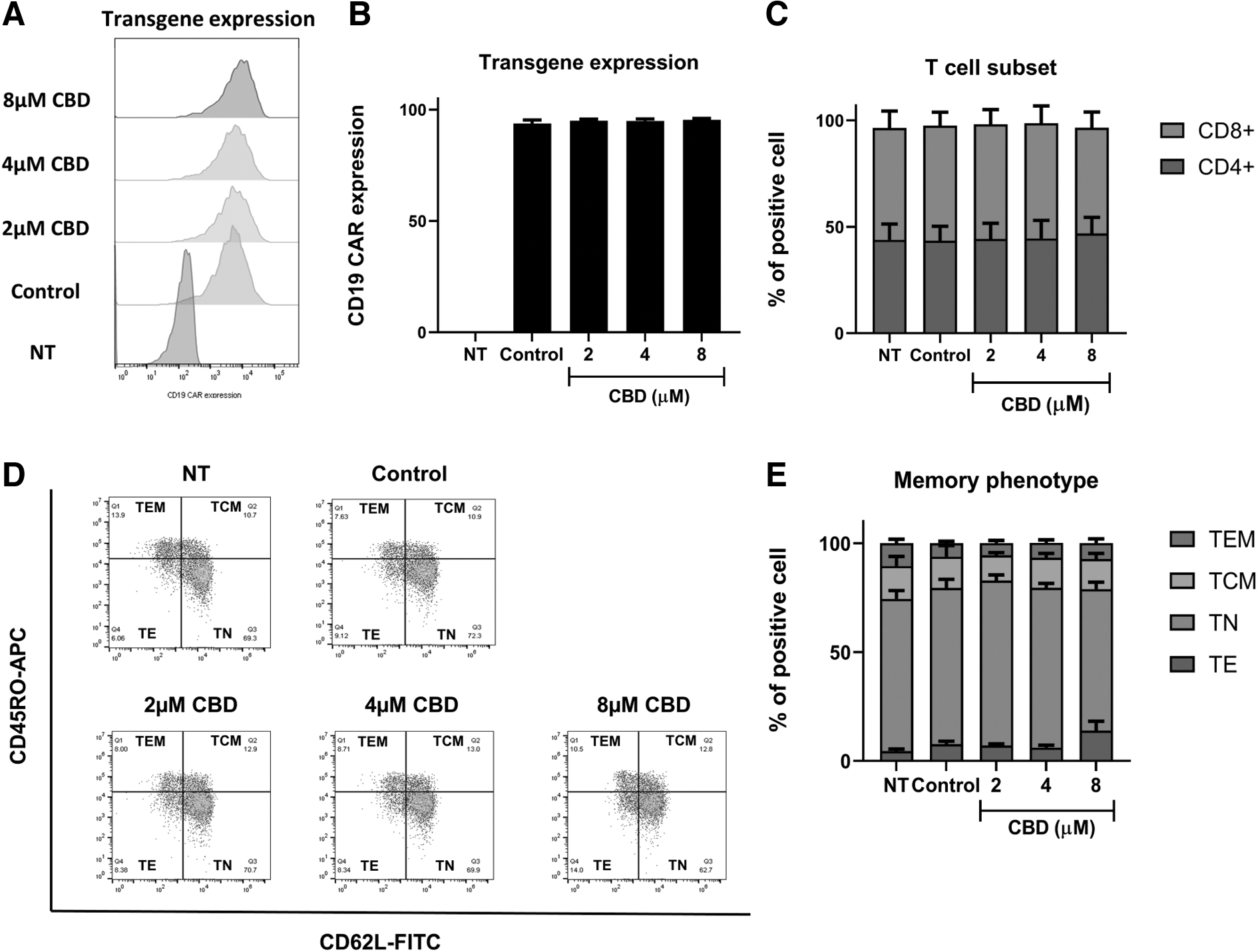
CBD did not alter immune characteristics of CAR T cells. NT and CD19-CAR T cells were treated with CBD at 0, 2, 4, and 8 μM for 7 days.
Treatment with CBD decreases CAR T cell proliferation and induces apoptosis
We next examined the impact of CBD on CAR T cell proliferation. CD19-CAR T cells were expanded in the presence of CBD at nontoxic doses ranging from 2 to 8 μM for 7 days. At concentrations of 2 and 4 μM, CBD did not affect the proliferation of nontransduced and CD19-CAR T cells. However, we observed that on day 4 of culture, CBD at 8 μM led to a reduction in CAR T cell proliferation. This effect was statistically significant on days 6 and 7 of the culture period (Fig. 4A). To investigate the inhibitory effect of CBD on cell proliferation, we performed an apoptosis assay on the fourth day of CBD treatment using annexin V-PI staining. We found that at 8 μM CBD, the number of apoptotic cells was significantly increased compared with that in untreated CD19-CAR T cells (Fig. 4B, C). We next assessed the effect of CBD on the cell cycle of CD19-CAR T cells and found that 8 μM CBD induced cells to enter the Sub-G1 phase, which is the apoptotic phase, and decreased the number of cells in the G2/M phase of the cell cycle (Fig. 4D).
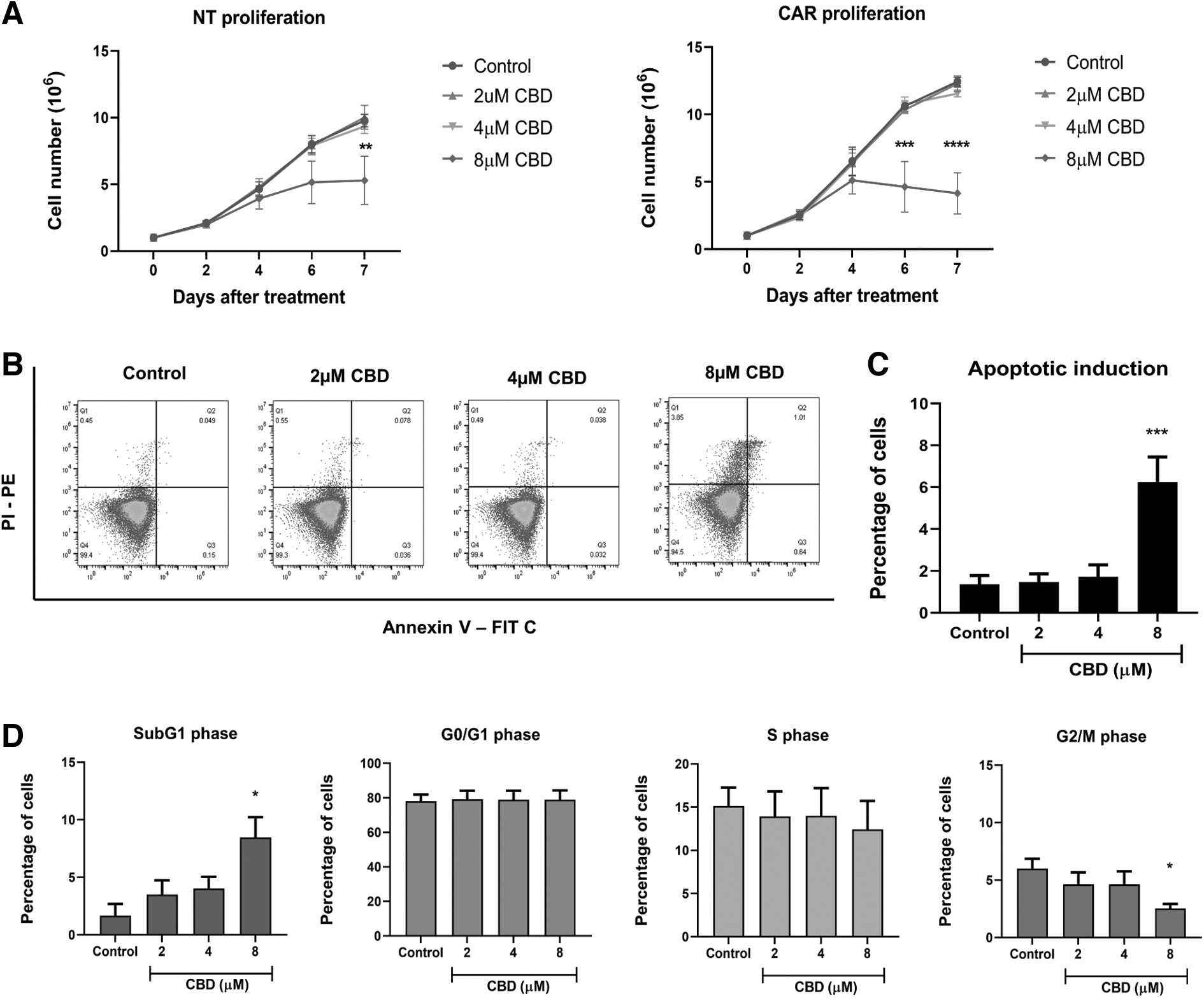
Treatment of CBD decreases CAR T cell proliferation and induces apoptosis and Sub-G1 cell cycle arrest. NT and CD19-CAR T cells were treated with CBD at 0, 2, 4, and 8 μM for 7 days. Cell proliferation was monitored every 2 days by Trypan blue exclusion.
CBD did not alter the cytotoxic function of CAR T cells
Having observed that CBD at a nontoxic dose induced CAR T cell apoptosis and decreased proliferation, we sought to investigate its effect on CAR T cell responses to the target antigen. We examined the effect of CBD on the degranulation of CD19-CAR T cells when stimulated with CD19+ tumor cells (NALM6 and Raji) by quantifying CD107a expression. Our results showed that CBD at 2–8 μM did not suppress the degranulation of CD19-CAR T cells when cocultured with tumor cells (Fig. 5A–C). Moreover, we investigated the effect of CBD on the antitumor activity of CD19-CAR T cells. CAR T cells were cocultured with target cells at an E:T ratio of 1:2 in the presence or absence of CBD for 24 h (Fig. 5D). Figure 5E illustrates the gating strategy of the killing assay using 7-AAD to exclude dead cells. The results showed that CBD did not alter the antitumor activity of CD19-CAR T cells when cocultured with NALM6 and Raji cells (Fig. 5F–H).
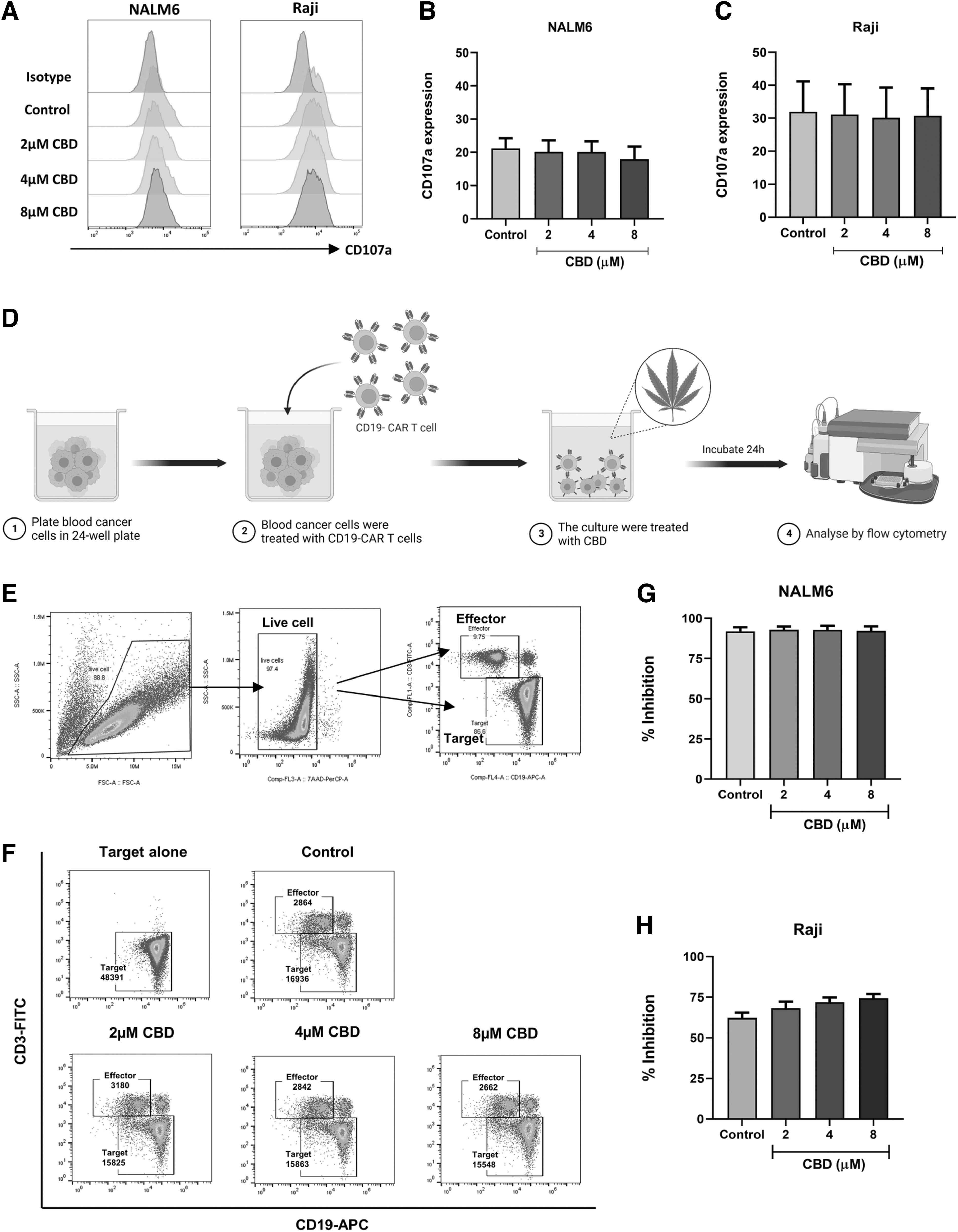
CBD did not alter cytotoxic function of CAR T cells. Degranulation was determined by stimulating CD19-CAR T cell with NALM6 and Raji cells at E:T ratio of 1:1 for 4 h.
In addition, we conducted cytokine assays encompassing four conditions: (i) target cells alone, (ii) CD19-CAR T cells alone, (iii) coculture of CD19-CAR T cells and tumor cells with a control vehicle, and (iv) coculture of CD19-CAR T cells and tumor cells with CBD (2–8 μM). As shown in Supplementary Figure S1, stimulating CD19-CAR T cells with CD19+ target cells (vehicle condition) significantly upregulated several cytokines, including TNF-α, IFN-γ, IL-2, and GM-CSF. Notably, CBD concentrations of 2–8 μM did not impede cytokine production from CD19-CAR T cells stimulated by tumor cells. In conclusion, the administration of CBD at nontoxic dosages did not compromise the cytolytic function or antitumor activity of CD19-CAR T cells.
Discussion
In this study, we examined the effect of CBD on the function of CD19-CAR T cells against ALL and B cell lymphoma by investigating immune characteristics, proliferation, and cytotoxic function. We found that nontoxic doses of CBD did not affect transgene expression, the T cell subset, or the memory phenotype. However, at a concentration of 8 μM, CBD suppressed CD19-CAR T cell proliferation by inducing apoptosis, as evidenced by the increase in Sub-G1 phase cell cycle arrest. However, CBD did not affect the cytolytic function, antitumor activity, or cytokine production of CD19-CAR T cells when cultured with tumor cells.
CBD has been known to exert various immunomodulatory effects. Previous studies have reported that CBD suppresses the proliferation of many immune cells, such as B cells, 21 T cells, 24 murine splenocytes, 30 and mouse microglia. 31 In addition, CBD has been shown to suppress cytokine production from lymphocytes, including IL-2, IFN-γ, TNF-α, IL-17A, and GM-CSF.23,24 The suppression of IL-2, a key cytokine regulating T cell proliferation, is linked to activating protein-1 (AP-1) and nuclear factor of activated T cells (NFAT) transcription factor suppression.21,24
Earlier studies, however, have indicated that CBD treatment at a nontoxic dose of 8 μM for up to 3 days did not exert suppressive effects on the proliferation of activated T cells, as measured by assays involving bromodeoxyuridine incorporation and intracellular dye carboxyfluorescein succinimidyl ester (CFSE) cell proliferation.23,24 Our findings aligned with these previous observations, highlighting the absence of a substantial inhibition effect within the initial 3-day timeframe. However, the proliferation of CAR T cells treated with CBD exhibited a decline on the fourth day and maintained a consistent decrease over subsequent days. This suggests a time-dependent response, where the inhibitory effect of CBD is dependent on its concentration and duration of exposure. Prolong exposure to the nontoxic dose of CBD appears to result in reduced T cell proliferation.
To further study the inhibitory effect of CBD on CAR T cell proliferation, we performed apoptosis assays and cell cycle arrest analyses. Although 8 μM did not show cytotoxicity as determined by the WST-1 assay, it suppressed cell expansion and induced apoptosis after long-term treatment. Apoptosis is a form of programmed cell death involved in many physiological processes to maintain cellular homeostasis. There are two pathways of apoptosis: (i) the intrinsic pathway through mitochondria-mediated caspase-9 activation and (ii) the extrinsic pathway through Fas death receptor-mediated caspase-8 activation. Both pathways lead to caspase-3 activation, which induces apoptosis. 32
Some mechanisms of CBD-induced cellular apoptosis have been proposed. A previous study reported that CBD induced apoptosis of lymphocytes through the formation of reactive oxygen species and caspase-8/caspase-3 activation. 33 CBD also acts as an agonist of TRPV1 and TRPV2 receptors, 34 in which triggering of TRPV1 and TRPV2 receptors induces the extrinsic pathway through Fas-mediated apoptosis.34,35 In addition, CBD, an antagonist of the GPR55 receptor, can inhibit the activation of the ERK pathway, which regulates cell cycle progression and cellular activation,36,37 leading to cell cycle arrest.
We also investigated the effect of CBD on the response of CD19-CAR T cells to the target antigen. Although CBD suppressed cell proliferation, it did not affect the cytolytic function or cytokine secretion of CD19-CAR T cells upon antigen stimulation. To activate T cells, endogenous T cell receptors recognize antigens presented on endogenous major histocompatibility complex (MHC) molecules. 38 Thus, in vitro stimulation of T cells is generally achieved by anti-CD3/anti-CD28 antibody stimulation. In contrast, CAR T cells are activated upon the binding of antigens with the scFV of CARs, leading to the activation of intracellular signaling components in the CAR construct. 2 Studies investigating the effect of CBD on T cell function have reported that CBD suppresses activation markers, cytokine production, and AP-1 and NFAT activity, leading to the inhibition of degranulation.21–24 Interestingly, we did not observe an inhibitory effect of CBD on the antitumor activities of CAR T cells, which could be attributed to the different signaling rewiring between CARs and T cell receptors.
In the present study, we reported the inhibitory effects of CBD on CAR T cells at a dose of 8 μM (∼2515 ng/mL). A pharmacokinetic analysis of CBD revealed that its plasma concentration is influenced by factors such as dosage, formulation, and route of administration. 39 Following oral administration (ranging from 5 to 50 mg/kg), plasma concentrations between 7.1 and 1200 ng/mL were observed.40,41 While this physiological range appears to be lower than the concentration at which inhibitory effects were observed in our in vitro experiments, it is worth noting that in a pre-clinical study, rats orally administered 50 mg/kg of CBD in a nano-emulsion formulation exhibited plasma levels of up to ∼3000 ng/mL. 42
Growing evidence indicated that the successful outcomes in patients undergoing CAR T cell treatment rely on the cells' ability to expand and persist after infusion, 43 and insufficient persistence of CAR T cells in vivo is a significant cause of relapse. 44 Considering that the persistence of CAR T cells critically influences the clinical response in CAR T cell therapy, 45 conducting further investigations to assess the impact of CBD on CAR T cells over extended periods and its potential effect on the in vivo antitumor activity of CAR T cells becomes essential. Such studies are warranted to elucidate the plausible clinical implications of combining CBD with immunotherapy.
Conclusion
To our knowledge, this is the first in vitro investigation on CBD's interaction with CAR T cell. We found that CBD affects CD19-CAR T cell proliferation but not their phenotypes or cytotoxic function. Our findings suggest that the immunomodulatory effects of CBD on the immune system should be carefully considered when developing combination therapies. Further studies are needed to better understand the potential benefits and risks of combining CBD with CAR T cell therapy.
Footnotes
Authors' Contributions
N.C.: Investigation, formal analysis, visualization, writing—original draft. K.C.S.P.: Visualization, writing—reviewing and editing. K.S.: Conceptualization, methodology, writing—reviewing and editing, funding acquisition. S.V.: Resources, writing—reviewing and editing. S.T.: Conceptualization, methodology, supervision, writing—reviewing and editing, funding acquisition.
Author Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This research is funded by the Thailand Science Research and Innovation Fund Chulalongkorn University (HEA663300010), The National Research Council of Thailand (NRCT), and H.M. the King Bhumibhol Adulyadej's 72nd Birthday Anniversary Scholarship, Chulalongkorn University, Bangkok, Thailand.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.