
Abstract
Purpose:
Our previous work demonstrated that the enhancement of cytotoxicity by protoporphyrin IX (Pp) was significantly higher than hematoporphyrin (Hp), when they were compared at the same ultrasound-exposure conditions, since the cytoskeleton plays a crucial role in numerous cell functions and the effect of sonodynamic therapy (SDT) on cytoskeleton has not been reported yet. So, it is very important to investigate whether SDT has some influence on the cytoskeleton, and it would also be interesting to compare Pp-/ and Hp-mediated SDT on the cytoskeleton.
Methods:
Ehrlich ascites carcinoma (EAC) cells were exposed to ultrasound for up to 15 seconds (1.34 MHz, 1 w/cm2) in the presence of 20 μM of Pp or 20 μM of Hp. Cell viability was determined by the trypan blue exclusion test. Morphologic changes were observed under a scanning electron microscope. Cytoskeleton damages were shown by immunofluorescence staining of microtubules, actin microfilaments, and intermediate filaments of vimentin.
Results:
Our experiment showed that the extent of cytoskeleton damage in EAC cells increased as time was prolonged, and also which was more significant in the Pp-SDT group than that in the Hp-SDT group at the same time after exposure.
Conclusions:
Results implied that the ultrastructural changes might account for cell destruction and the cytoskeletal components were involved in the cell-damaging process induced SDT treatment.
Introduction
Sonodynamic therapy (sdt) is an approach based on the synergistic effect of ultrasound and a chemical compound referred to as the “sonosensitizer.” The effect can be localized by focusing ultrasound on a defined region and choosing compounds with tumor affinity. 1,2 The ability to enhance drug cytotoxicity with ultrasound that enables efficient, but localized, effects on a pathologic site with minimal damage to peripheral healthy tissue would be a valuable clinical asset. 3 Possible mechanisms of SDT may include the following: (1) initiating chain peroxidation of membrane lipids via peroxyl and/or alkoxyl radicals derived from sonosensitizers, (2) rendering the cell more susceptible to shear forces because of the cell membrane physically destabilized by irradiation, and (3) enhancing drug transportation across the cell membrane induced by ultrasonic treatment. 4 However, the mechanism of SDT may be influenced by a number of factors, including the sonosensitizer, the ultrasound parameters, and the nature of the biologic model, so the precise bioeffects of SDT on cells or tissues are not fully understood as yet.
Hematoporphyrin (Hp), the most widely used sonosensitizer, in mediated sonodynamic therapy has been well investigated in vivo and in vitro. 5 –7 Protoporphyrin IX (Pp), a component of Hp derivatives, accumulates in rapidly proliferating cancer cells and thus provides selective destruction of tumors. Our previous work demonstrated that the enhancement of the synergistic inhibitory effect by Pp was significantly higher than Hp, when they were compared at the same ultrasound-exposure conditions on sarcoma 180 solid tumors, and the ultrastructure changes might account for inhibiting tumor growth and even inducing cell death. 8 The ultrastructure changes reflected the overall morphologic destruction, which suggested that ultrasound irradiation with Pp and Hp played an important role in damaging the cell-membrane system and the internal morphologic destruction. Since the cytoskeleton plays a crucial role in numerous cell functions, such as sustaining cell normal morphology, cell proliferation, signal transduction, and so forth, 9 –11 the possibility of selective targeting of cytoskeletal proteins [e.g., microtubule (MT), actin microfilament (AF), and intermediate filament of vimentin (VF)] constitutes a very important objective for all cancer therapies. 12
5-aminolevulinic acid (ALA), a precursor of the endogenous photosensitizer, Pp, is widely used in photodynamic therapy (PDT). ALA-PDT-induced changes in the cytoskeleton, cell shape, and adhesion have been reported by researchers. 13 However, so far, the effect of SDT on the cytoskeleton has not been reported as yet. So, it is very important to find out whether the sonodynamic effects have some influence on cytoskeleton, and it would also be interesting to compare the sonodynamic effects of protoporphyrin-IX-mediated sonodynamic therapy (Pp-SDT) with hematoporphyrin-mediated sonodynamic therapy (Hp-SDT) on the cytoskeleton of Ehrlich ascites carcinoma (EAC) cells. In the present study, the trypan blue exclusion test was used to determine cell viability after SDT treatment, and a scanning electron microscope (SEM) observation of the surface of cells was performed to evaluate the morphologic changes induced by ultrasonic irradiation; also, the changes of relative key cytoskeletal elements (MT, AF, and VF) of EAC cells were detected by using the immunofluorescence staining method. The underlying mechanism of cell damage was also evaluated during SDT treatment.
Materials and Methods
Chemicals
Hematoporphyrin and protoporphyrin IX disodium salt were purchased from Sigma Chemical Co. (St. Louis, MO) and had a purity of 60% and 94% high-performance liquid chromatography: (HPLC), respectively. Their chemical structures are shown in Figure 1.
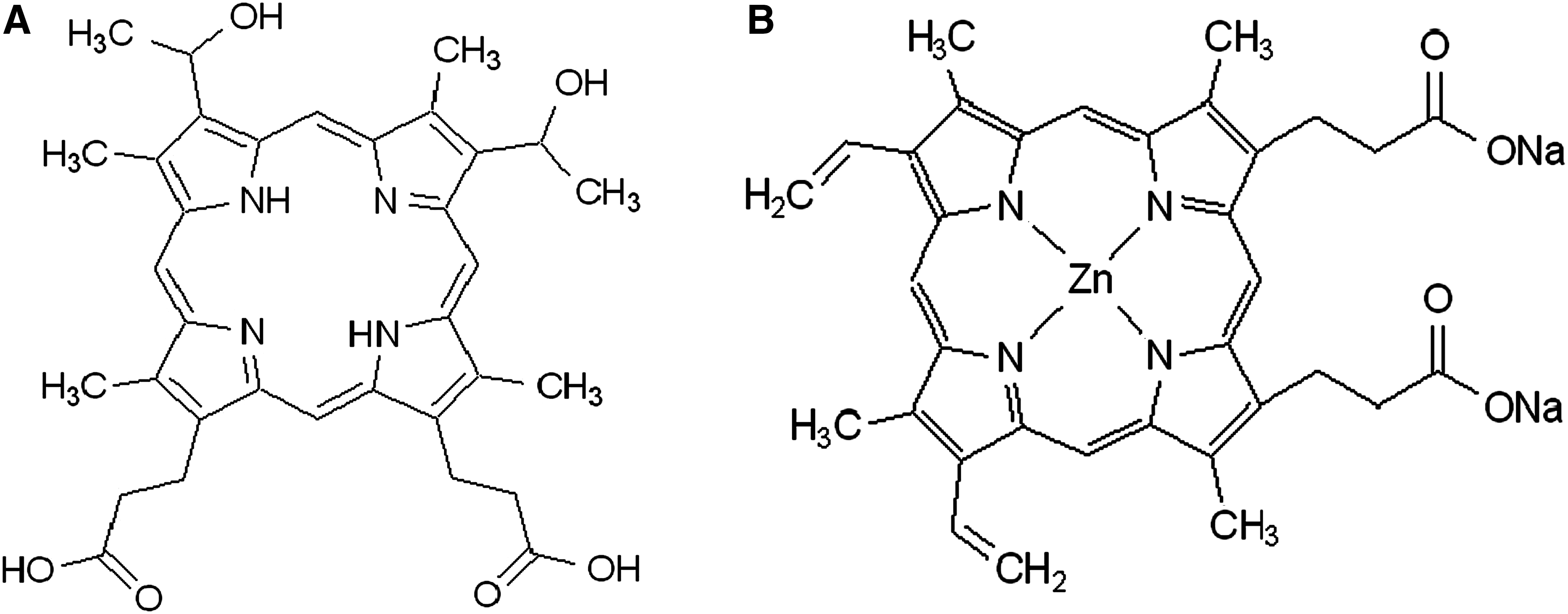
Chemical structure of hematoporphyrin (
Rabbit polyclonal antibody specific for β-tubulin was obtained from Santa Cruz Biotechnology (Santa Cruz, CA). Mouse monoclonal antibody (mAb) (clone V9) specific for vimentin was purchased from Wuhan Boster Biological Technology (Wuhan, China). Fluorescein isothiocyonate (FITC) labeled phalloidin, Cy3-conjugated goat anti-rabbit IgG, and FITC-labeled goat antimouse IgG were all obtained from Sigma.
Ultrasound exposure
The experimental set-up for insonation is shown in Figure 2. An ultrasound transducer with a diameter of 23 mm and a focal length of 43 mm was submerged in the bottom of a glass container filled with cold, degassed water. A polystyrene sample test tube containing a 1.0-mL cell suspension was placed into the focal area of the transducer for insonation.
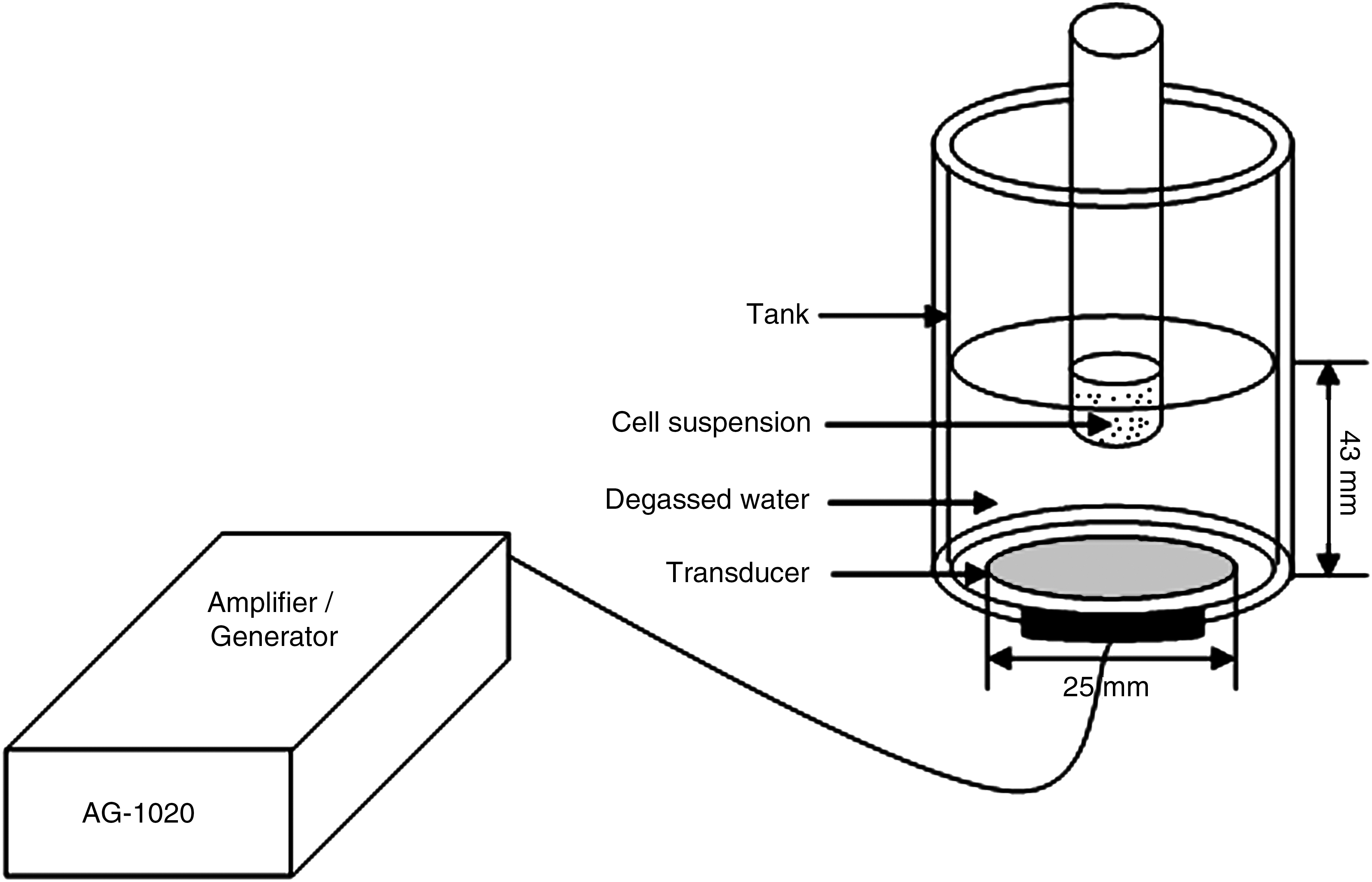
Ultrasonic exposure equipment.
The ultrasound transducer was manufactured by the Institution of Applied Acoustics at Shaanxi Normal University (Xi'an, China). The same transducer was used for all experiments, with a resonant frequency of 1.34 MHz in a standing wave mode, and it was used to convert the electrical power measured by the amplifier (T&C Power Conversion, Inc., Rochester, NY) into acoustic power. In order to specify the intensity in the insonation experiment and have an easy, obvious understanding, we used the reading-output power from the amplifier, representing our spatial average ultrasound intensity in our experiment system. For all experiments, cold, degassed water (4°C) was used as the ultrasonic coupling medium, thus reducing the thermal effect caused by ultrasound irradiation. The temperature within the cell suspensions was checked with a thermometer, and found that the temperature rise was unlikely to induce thermal damage of cells during such a short duration.
Tumor cells and animals
The EAC cells and Institute of Cancer Research (ICR) mice were supplied by the Experimental Animal Center of Shaanxi of the Chinese Traditional Medicine Institution. The cell lines were passaged weekly through ICR mice with a body weight of 18–22 g in the form of ascites. After 7–10 days of in vivo inoculation, the cells were harvested from the abdomen, washed 3 times by phosphate buffer solution (PBS; pH 7.4), collected by centrifugation at 2000 rpm for five min, then resuspended in PBS at a concentration of 3 × 106 cells/mL.
Treatmental protocol
The cell suspensions were put in polystyrene sample test tubes (1 mL). These tubes were randomly divided into 6 groups as follows: (1) control group (CT); (2) those treated with hematoporphyrin (Hp); (3) protoporphyrin IX (Pp); (4) ultrasound (Us); (5) ultrasound+Hp (Uh), and (6) ultrasound+Pp (Up). Each group had four test tubes. The final concentrations of Hp and Pp were 20 μM, while instead of Hp or Pp, equivalent PBS was administered to tubes in the CT and Us groups. All tubes were incubated at 37°C for 30 minutes; then, the Us, Uh and Up groups were exposed to ultrasound for up to 15 seconds. Postultrasonic treatment, the cells were washed by PBS and resuspended in fresh RPMI-1640 medium with 10% fetal bovine serum (FBS) for incubation at 37°C in a humidified 5% CO2 atmosphere. A series of experiments were done at 0, 2, and 4 hours after SDT treatment, respectively.
Cell survival assay
Immediately after SDT treatment, the cell viability was detected by the trypan blue exclusion test. A 100-μL cell suspension was taken from the sample test tube and mixed with 200 μL of 0.4% trypan blue solution. Cell integrity was determined by counting the number of unstained cells on a hemocytometer glass plate using an optical microscope. The cell-survival rate was obtained from the following formula: cell survival rate = the number of unstained cells/the number of cells in CT group × 100%.
SEM observation
At different times after treatment, cells for SEM were fixed by 2.5% glutaraldehyde in 0.1 M of PBS (pH 7.4) and postfixed by 1% osmium tetroxide, then washed by PBS, dehydrated by graded alcohol, displaced, dried at the critical point, gold evaporated, and observed by using an SEM (S-570, Hitachi, Tokyo, Japan).
Fluorescence staining
Indirect immunofluorescence analysis
After different treatments, EAC cells were placed on glass slide, fixed by 4% fresh paraformaldehyde (PFA) in PBS (pH 7.4) for 10 minutes at 37°C. For the β-tubulin study, the fixed cells were permeabilized by incubating with 0.1% Triton X-100 in a MT-stabilizing buffer, PEM (100 mM Pipes, 2 mM EGTA, 1 mM MgSO4; pH 6.8), for 10 minutes at 37°C, while for VF detection, the fixed cells were permeabilized with 0.1% Triton X-100 in PBS for 10 minutes at 37°C; then, cells were washed by PBS 3 times, blocked with normal goat serum for 45 minutes, and incubated with the primary antibodies of β-tubulin (1:50) or vimentin (1:50) overnight at 4°C. Substitutions of PBS for the primary antibodies were used as negative controls. After washing by PBS, cells were incubated with the corresponding specific secondary antibodies (Cy3-IgG for β-tubulin and FITC-IgG for vimentin, respectively) overnight at 4°C, rinsed in coverslips with PBS, and mounted on slides with glycerol-PBS (1:1), the samples were observed with a Leica (Wetzlar, Germany) TCS SP5 inverted confocal laser-scanning microscopy equipped with a 100 × oil immersion lenses, and cell morphology was documented by differential interference contrast (DIC).
FITC-phalloidin-labeled F-actin
For F-actin, PFA-fixed cells were washed extensively in PBS, permeabilized with 0.1% Triton X-100 in PBS, and then washed again in PBS. Cells were stained with a 0.5-μg/mL FITC-phalloidin in PBS for 1 hour at room temperature and washed several times in PBS to remove unbound phalloidin conjugate. F-actin was imaged by fluorescence microscopy, using a Nikon E600 microscope with 100 × oil immersion lenses. The images were captured by a CCD camera equipment with the same exposure settings. Status of F-actin polymerization was quantitated by randomly choosing at least 10 microscopic fields (40 ×) and counting cells in the field as either normal (F-actin uniformaly localized at the cellular periphery) or abnormal (diffused and decreased F-actin flurescence signal or F-actin in the membrane blebs) cells. The normal F-actin ratio could be expressed as the number of normal cells/the total number of cells.
Results
Cell viability assay
The viability of EAC cells in the presence of Hp or Pp after a 15-seconds exposure to ultrasound was detected immediately post-treatment. As shown in Figure 3, no cell damage was observed in the untreated CT group, cell viability in Hp and Pp groups had no difference with CT, and the ultrasound-alone treatment group had only a slight antitumor effect, while cell survival rates in the synergistic treatment groups significantly decreased and the cell killing effect was more obvious in the Up than in the Uh group (P < 0.01).
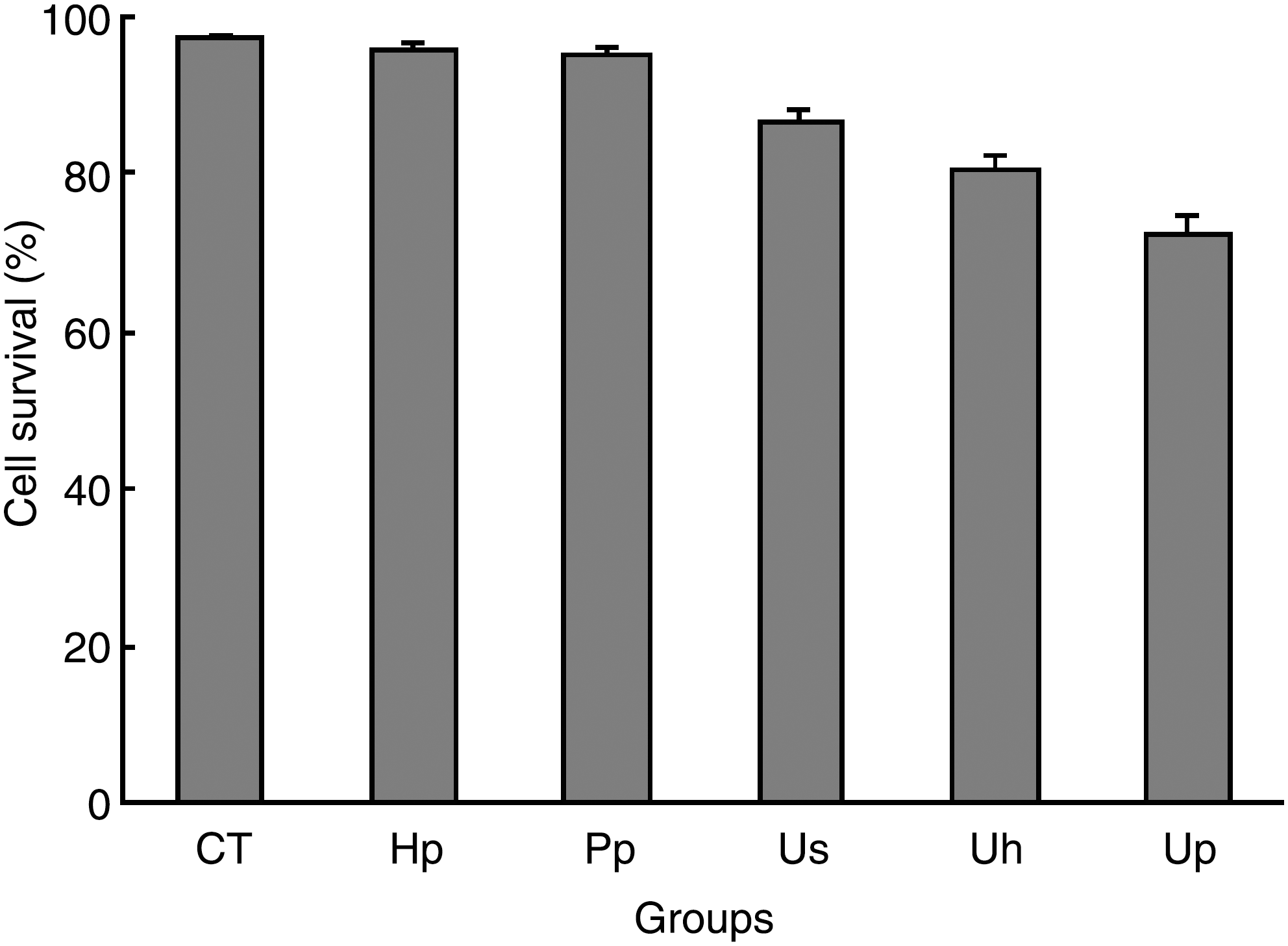
The viability of EAC cells after different treatments. CT, control group, with no treatment; Hp, 20 μM Hp alone; Pp, 20 μM Pp alone; Us, ultrasound alone; Uh, ultrasound plus 20 μM Hp; Up, ultrasound plus 20 μM Pp.
Results of SEM observation
When observed under SEM, untreated cells showed a typical surface morphology, with numerous randomly distributed microvill (Fig. 4A). Hp (Fig. 4D) or Pp (Fig. 4G) alone had only a slight effect on the surface of the cells until 4 hour post-treatment and had no obvious difference with the CT group. Immediately after treatment, cells subjected to ultrasound showed no significant difference with CT group cells, while after 2 hour of incubation, the number of microvilli on most cell membranes was remarkably decreased (Fig. 4B), and the surface of many cells became relatively smooth with no obvious microvilli; several small craters were also seen on some cell membranes until 4 hour post-treatment (Fig. 4C). In the synergistic treatment groups, the extent of cells damaging was increased in a time-dependent manner. In both the Uh and Up groups, the number of microvilli remarkably decreased at 2 hour after treatment (Fig. 4E and 4H, respectively) and had apparent deformation 4 hour post-treatment (Fig. 4F and 4I, respectively). However, cells in the Up group were more seriously damaged than those in the Uh group, and after 4 hour of incubation post-treatment, cells in the Up group with some irregular papillous protuberances were seen in the surface where the cytoplasm seemed to have extruded through the membrane boundary (Fig. 4I).
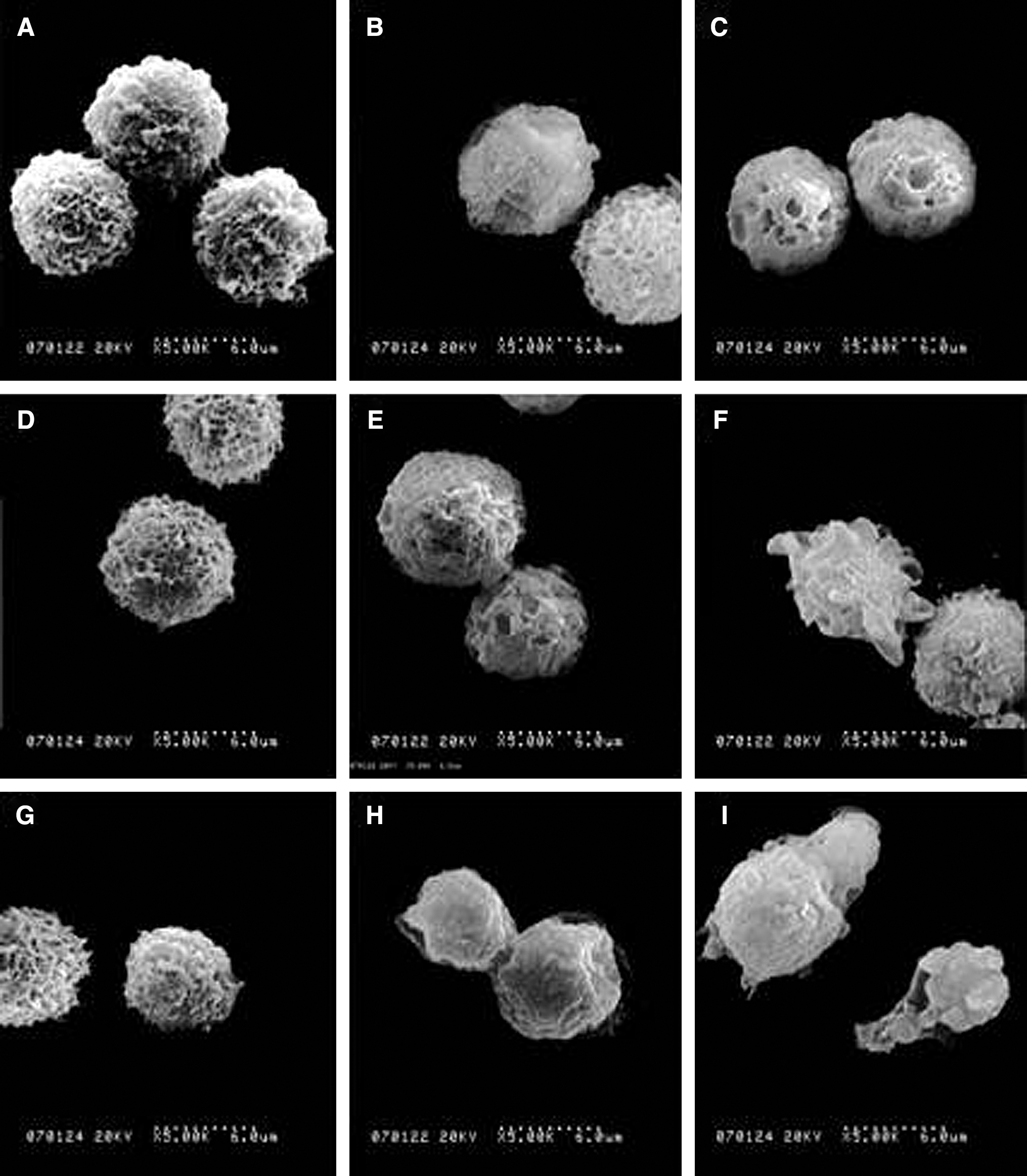
SEM images of EAC cells. (
Sonodynamic effect on the microtubule cytoskeleton
The polymerization state of microtubers (MTs) was examined by immunofluorescence staining of β-tubulin immediately and several hours of culture after treatment. As shown in Figure 5, control EAC cells showed a well-developed MT network around the nucleus (Fig. 5A) and had no changes after 4 hour of incubation (Fig. 5C). The MTs polymerization state of the Hp (Fig. 5D) and Pp (Fig. 5B) groups cells had no time-dependent changes, while the MT fluorescence signal in the Us alone group was partially decreased at 4 hour after treatment (Fig. 5F). In the synergistic treatment groups, immediately after treatment, most cells in the Uh group were minimally affected and were quite similar to controls either in the appearance of MT network or in the overall morphology, and only a small part of cells had diffused fluorescence (Fig. 5G), while the volume of most cells became larger and the MT network diffused in the Up group (Fig. 5J). After 2 hour of culture, the overall morphology of most cells in the Uh group were slightly changed under DIC microscopy (Fig. 5E), with the fluorescent signal reduced and diffused (Fig. 5H), and it was scarcely visible in some parts of cells. This phenomenon was also been seen in the Up groups at the same time (Fig. 5K). Further, 4 hour post-treatment, cells in the Uh group were corresponded with a variable amount of perinuclear vacuoles with a disorganized MT network (Fig. 5I); however, the morphology of most cells in the Up group were deformed and some extracellular blebs containing tubulin were also could be seen on the surface of cells (Fig. 5L).
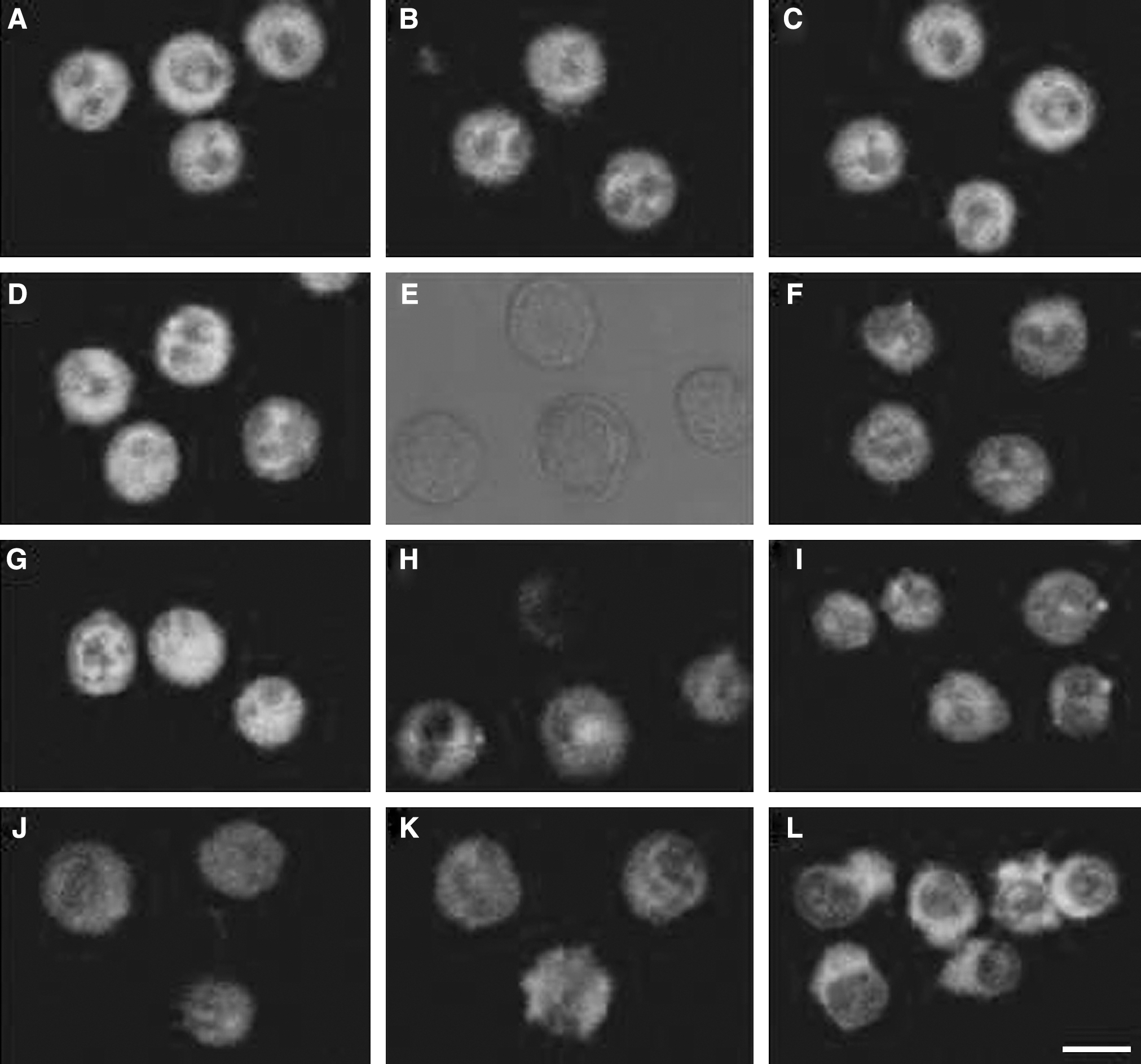
Immunofluorescence staining of β-tubulin in EAC cells subjected to Hp-SDT and Pp-SDT, respectively. (
Sonodynamic effect on the intracellular vimentin cytoskeleton
The fluorescence study revealed a restructuring of the intermediate filament networks represented by vimentin elements. Cells showed increasing vimentin alterations as time was prolonged, and which was more serious in Pp-SDT group cells than that in Hp-SDT group cells at the same experiment conditions. As shown in Figure 6, vimentin cytoskeleton of CT groups cells formed a network around the nucleus (Fig. 6A), and it still distributed well after 4 hour of incubation (Fig. 6C). Changes in Hp (Fig. 6D) and Pp (Fig. 6B) group cells were slight until 4 hour of culture, while the vemintin fluorescence in the Us group was a part of attenuation at 4 hour after treatment (Fig. 6F). Immediately after Hp-SDT treatment, cells retained the morphology of control and the vimentin cytoskeleton of most cells was minimally affected; it was disturbed and diffused in a small part of cells (Fig. 6G); 2 hour later, there were many vacuoles in some parts of cells (Fig. 6E) with scarcely visible vimetin network (Fig. 6H), and there were small blebs on some cell membranes at 4 hour post-treatment (Fig. 6I). In the corresponding Up group, cells were just similar to those of Uh group, but the volume of them was larger immediately after treatment (Fig. 6J). After 2 hour of incubation, a diffused vimetin network and vimetin in grains dispersed over the cytoplasm were observed in some cells, while plasm a-membrane blebs that contained vimetin could be seen in a few cells (Fig. 6K). Severe alterations in morphology and distribution of vimentin could be detected at 4 hour after treatment. Most of cells exhibited alterations of the plasma membrane, and numerous blebs showing homogeneous fluorescence due to vimentin could be seen on the cell surface (Fig. 6L).
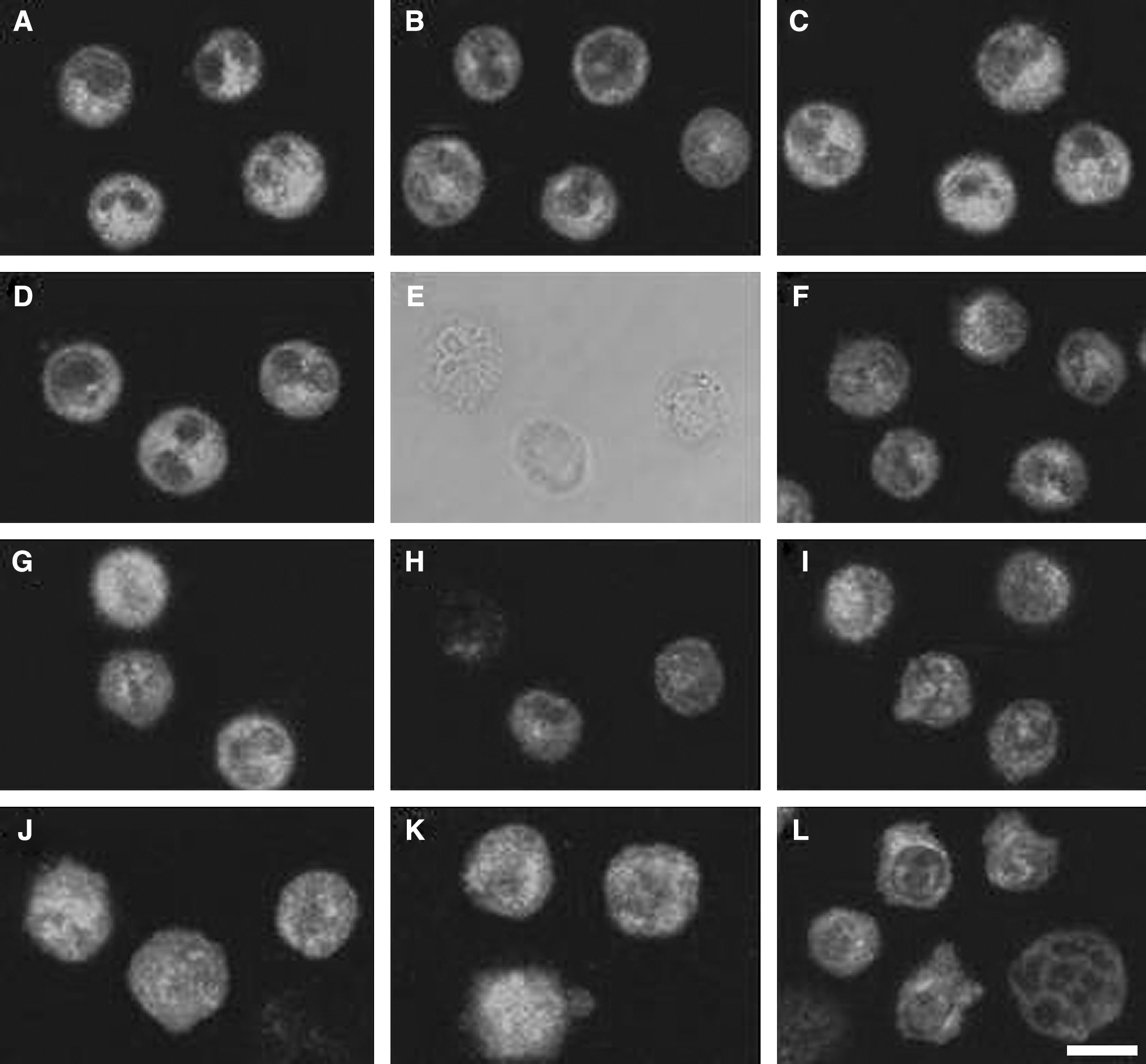
Sonodynamic effects by Hp-SDT and Pp-SDT on the vimentin fibers. (
Sonodynamic effect on the intracellular actin cytoskeleton
The actin cytoskeleton was revealed by the fluorescence of FITC-phalloidin, which selectively stains F-actin. The control pattern of F-actin mainly localized at the cellular periphery and had no obvious time-dependent changes (Fig. 7A–C). Cells in the Hp (Fig. 7D) and Pp (Fig. 7E) groups were similar to control, while the Us group cells had slightly changes after 4 hours of incubation (Fig. 7F). As in the case of MT and VF, SDT induced a variable disorganization and reduction in the number of AF, and the sonodynamic effects of Pp-SDT on F-actin of EAC cells was stronger than that of Hp-SDT. There were at least three different cell populations that could be seen both in the Uh and Up groups. The first population was composed of cells quite similar to control either in the appearance of F-actin network or in the overall morphology; the second corresponded to cells larger in cell volume with a diffused and clustered F-actin network; and the third was formed by cells with blebs on the plasma membrane or the intense fluorescent signal of F-actin presented at the base of blebs. Immediately after synergistic treatments, cells in both the Uh (Fig. 7G) and Up (Fig. 7J) groups were mainly of the first and second types; after 2 hours of incubation, most cells in the Uh (Fig. 7H) and Up (Fig. 7K) groups were the second and third types; cells with blebs on the plasma membrane in the Uh (Fig. 7I) and Up (Fig. 7L) groups increased 4 hours post-treatment. So, the percentage of normal F-actin decreased in a time-dependent manner after SDT treatment, as shown in Figure 8, where the percentage of normal F-actin was remarkably decreased in Uh-group cells with incubation time (0 hour: 78%, 2 hour: 72%, 4 hour: 64%, respectively), and which was more significantly decreased in Up-group cells with time (0 hour: 74%; 2 hour: 62%; 4 hour: 49%, respectively).
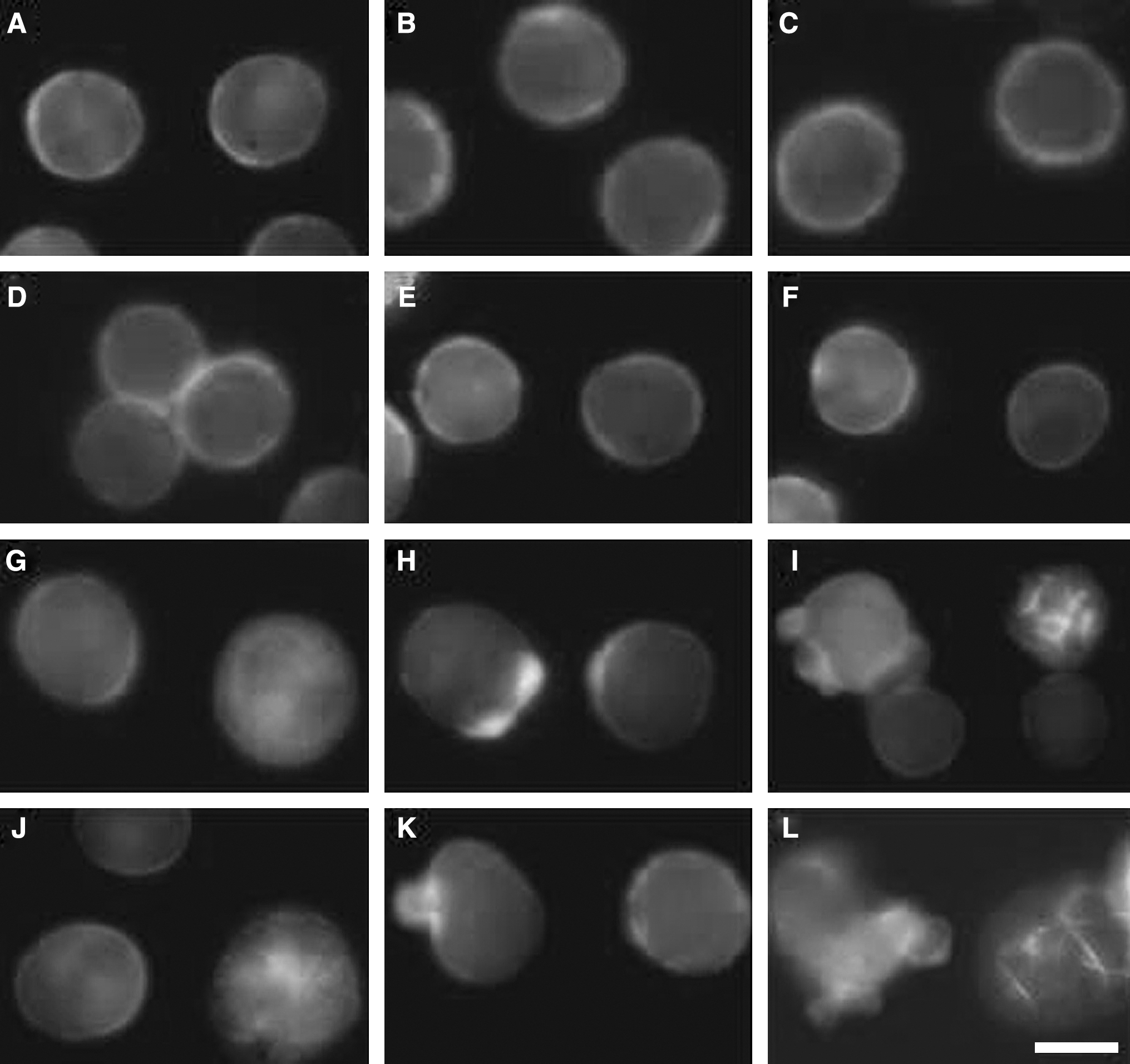
Fluorescence staining of F-actin after sonodynamic treatment, labeled by FITC-phalloidin. (
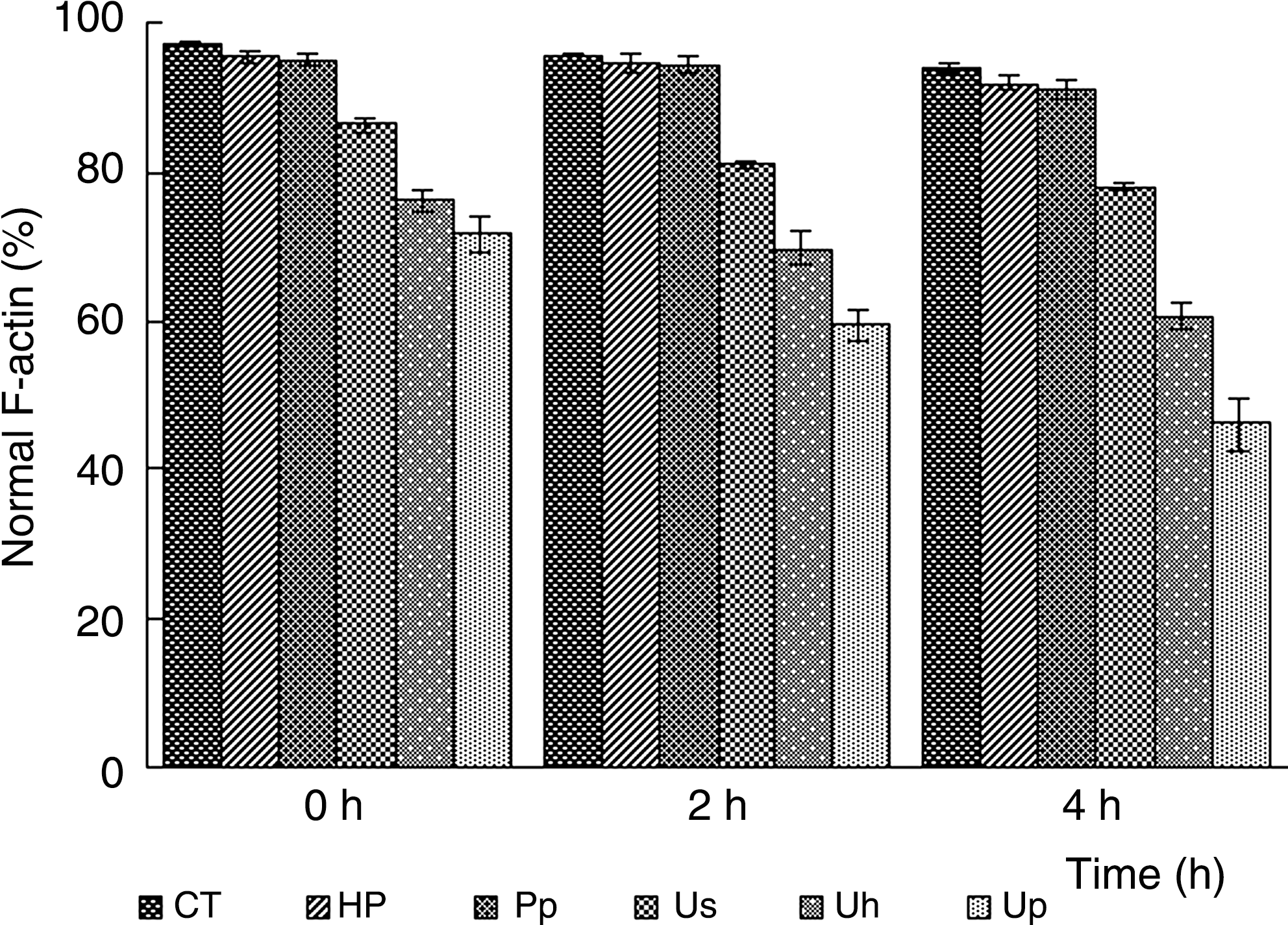
The normal F-actin ratios of EAC cells after different treatments. Each point corresponds to the mean value ± standard deviation from at least three independent experiments.
Discussion
In our previous work, we have found that Pp could induce more potential cytotoxicity than Hp when irradiated with ultrasound at the same experiment conditions both in vivo and in vitro. 8 And the ultrastructure changes suggested that ultrasound irradiation with Pp or Hp played an important role in damaging the cell-membrane system and the internal structures. In the present study, it was very interesting to investigate whether the sonodynamic effects would have some influence on the cytoskeleton and also to compare the difference of Pp-SDT treatment with Hp-SDT treatment on EAC cytoskeletons.
First, the cell viability after different treatments was determined by the trypan blue exclusion test. Results showed the synergistic treatments had significant antitumor effects, in comparison with other sole treatments, and the cell damaging was more obvious in the Up than in the Uh group at the same experiment conditions (P < 0.01). Then, the overall morphologic changes induced by ultrasonic irradiation were detected by SEM observation. Figure 4 shows that the synergistic treatment group cells were significantly influenced, and the morphologic changes also showed a time-dependent manner, mainly including the decreasing of microvilli and the deformation of cells. These phenomena in the Up group were more serious than that in the Uh-group cells at the same time point. The results implied that cell structure was remarkably damaged, and the membrane deformation seemed to be filled with the extent of cytoskeleton distribution. So, finally, the changes of relatively key cytoskeletal elements (MT, AF and VF) of EAC cells were detected by using the immunofluorescence staining method to evaluate the underlying mechanism of cell damaging during irradiation. After SDT treatment, changes of three cytoskeletal elements all presented time-dependent manners, MTs showed a diffused fluorescence and presented an aberrant intracellular distribution, vimentin fibers reacted slightly to its antibody, most F-actin were diffused and clustered, and these alterations were particularly obvious from 2 to 4 hours post-treatment. The disintegration of these cytoskeletal components could be due to the depolymerization of MT to α-and β-tubulins and AF to G-actin, or to a direct breakage of these structures. Thus, the damage effects observed in cytoskeletal elements could be induced either by a direct effect of the certain mechanical stress of SDT on the cytoskeletal proteins or by indirect effect induced by the free radicals generated after ultrsonic exposure. However, it seems very difficult to separate mechanical actions due to shear stress from chemical actions due to free radicals as the cause of cellular cytoskeleton damage after SDT treatment.
In addition, plasma-membrane blebbing became a prominent phenomenon post-SDT treatment. We could observe the tubulin-/ and vimentin-containing blebs formed in most of cells under the confocal laser-scanning microscopy at 4 hours after Pp-SDT treatment, and F-actin presented with blebs could be clearly seen in both Hp-SDT and Pp-SDT group cells from 2 to 4 hours postexposure. The majority of studies examining the formation of membrane blebs have focused on the role of key cytoskeletal proteins. 14 Blebbing of the cell surface is often accompanied by the disruption of microtubules 15,16 and the reorganization of actin microfilaments. 17 Therefore, our results implied that not only destruction, but also reconstruction of some cytoskeleton systems were necessary for cell damaging induced by SDT treatment. Also, membrane blebbing was reported to be associated with apoptosis. 18 –23 Numerous studies have focused directly on the role of actin in the apoptotic membrane changes, 24 –26 and the concentration of F-actin is correlated with bleb size. 27 In this study, we could see F-actin presented at membrane blebs or at the base of blebs after SDT treatment, which may have indicated the apoptosis action induced by ultrasonic exposure. Also, actin microfilaments in cells with synergistic treatments appeared as clusters contrary to uniformaly stained control cells, presumely because changes of several cytoskeleton-organizing proteins due to SDT treatment. However, futher studies need to be done to confirm the conclusion.
Conclusions
In summary, we showed, for the first time, that the extent of cytoskeleton damage increased as time was prolonged, and the inhibitory effect was stronger in the Pp-SDT group than that in Hp-SDT group at the same time point after SDT treatment in EAC cells. The comparative results indicated that the cytoskeletal components are involved in cell damaging induced by SDT, and several reasons may account for the different impacts between Hp and Pp on SDT, including there different structures and distinct subcellular localization patterns. The intracellular cytoskeletal components may constitute basic, important targets of SDT, and the sonodynamically induced cytoskeleton changes may account for cell deformation and even cell death; however, the detailed mechanism still needs further investigation.
Footnotes
Acknowledgments
This work was supported by the Natural Science Foundation for the Youth (no. 10904087) and the Excellent Doctor Innovation Project of Shaanxi Normal University.
Disclosure Statement
No competing financial interests exist.